








-
潮滩是海岸带地区高潮时淹没、低潮时露出的区域,也是受陆海交互作用强烈影响的生态过渡地带[1]。由于独特的水文环境和生态系统动态特点,潮滩湿地是地球上生产力最高、生态服务价值最大的生态系统之一[2]。典型潮滩湿地有淤泥质海滩湿地、潮间盐水沼泽湿地、潮间森林沼泽湿地、红树林湿地等。中国海岸带湿地每年总固碳量大于4×105 t·a-1,潮滩湿地中的盐沼植被作为滨海湿地的重要组成部分,是重要的碳汇[3]。盐沼湿地植被位于海岸带缓冲区,不仅为水鸟等生物提供丰富的食源、安全的隐蔽场所和繁殖地[4-5],也具有消浪护岸、保滩促淤、水质净化等重要的生态服务功能,其演化直接关系到沿海区域社会经济可持续发展和生态安全[6-9]。近年来,由于海平面上升[8]、海岸带围垦[9]、外来物种入侵[10]等自然和人为因素的影响,潮滩盐沼植被面积萎缩、功能退化,受到的威胁日趋严重,给中国沿海城市带来巨大生态威胁和环境风险[11-12]。因此,需要构建完善、科学的潮滩盐沼湿地监测与评估体系,掌握潮滩植物现状和动态变化。由于潮滩盐沼植被受潮汐周期性影响,现场大范围调查困难。因此,常采用遥感监测的手段,大面积、快速地获取其种类、数量及其空间分布信息并进行相应的生物物理参数遥感反演估算。国内研究者多利用Landsat等多光谱遥感影像对中国盐沼植被分布及时空变化进行研究,由于多光谱遥感影像的光谱分辨率低,较难满足对潮滩盐沼植被识别的需求[13-14]。高光谱卫星影像的发展为盐沼植被遥感识别提供了新的手段,它一般具有几十、几百个波段信息,使其在盐沼植被识别分类方面有独特的优势[15-16]。BELLUCO等[17]利用ROSIS和CASI等多种遥感影像对意大利东北部潮间带盐沼植被进行分类,并结合实地测量,发现高光谱影像的分类精度明显高于多光谱影像。然而,高光谱遥感影像信息量大,冗余信息多,有效光谱波段信息提取难度大。如何选择合适的高光谱影像波段,研究多波段的有效信息提取,是其应用的技术关键[18]。地物光谱仪以其灵活测量的特点,能够设计大量实验现场观测不同植被生理过程的有效光谱信息,通过分析并选择地物的特征波段,与高光谱遥感影像波段结合,为高光谱遥感影像识别地物提供依据[19-21]。盐沼植被光谱差异不仅受植被生长季相影响[22],还受植被盖度、下垫面等不同因素影响[23-24]。以往在对潮滩盐沼植被的光谱研究中,没有充分考虑潮滩土壤粒径和土壤湿度快速动态变化对植被光谱的影响,特别是涨退潮期间水体背景对植被光谱反射的影响[25]。因此,为提高遥感影像信息提取精度,针对潮滩盐沼植被生境特点,以长江口潮滩地带为研究区域,使用地物光谱仪测定潮滩地带典型盐沼植被及其下垫面的光谱反射率,获得不同类型和盖度盐沼植被光谱反射特征,以及不同土壤类型和潮位背景影响下的盐沼植被光谱反射率的变化规律,建立潮滩盐沼植被光谱特征库,为潮滩湿地盐沼植被高光谱遥感影像解译提供依据,实现遥感手段快速准确获取潮滩湿地植被的时空变化及生态环境变化。
-
长江口是长江在中国东部入海的河口,因受径流、水下地形等因素影响。该河口内为不正规半日潮。潮滩湿地因受长江口特殊的气候、水文、地质、地貌、泥沙,尤其是人为因素影响,潮滩格局、地形及植被经长期演替均发生较大变化。
长江口潮滩湿地资源极其丰富,潮滩上的盐沼植被以芦苇Phragmites australis群落、海三棱藨草Scirpus mariqueter群落、藨草Scirpus triqueter群落、糙叶薹草Carex scabrifolia群落和互花米草Spartina alterniflora群落为主[23]。芦苇群落是长江口潮滩优势种群,主要分布于高潮滩区域。互花米草是外来入侵种,自上海市引入以来,在崇明岛及九段沙大肆蔓延[12]。海三棱藨草主要分布于低潮滩,是中国的特有种。由于潮滩高程差异,不同高程潮滩的水淹时间不同,海三棱藨草群落密度随高程的增加而增大。在光滩区域群落密度低,一般为10%~20%,植株一般较低矮,通常在20 cm以下。海三棱藨草在高潮时几乎完全被淹没,低潮时露出。海三棱藨草的遥感监测因其植株低矮稀疏,下垫面易受潮汐影响。
本研究选择崇明东滩(CMDT),崇明北八滧(CMBBX)和南汇边滩(NHBT)等3个长江口典型潮滩湿地。崇明东滩(31°24′59.72″~31°37′54.37″N,121°45′44.52″~122°04′47.78″E)位于上海市崇明岛东部,南临长江入海口,东濒东海,主要由长江入海泥沙堆积形成,是典型的淤泥质潮滩,主要分布有海三棱藨草群落和芦苇群落。崇明北八滧(31°40′55.20″~31°36′57.60″N,121°41′2.40″~121°51′18″E)位于崇明岛北沿,濒临长江北支,水流平缓,潮滩淤涨快,湿地资源丰富,主要分布有海三棱藨草群落和互花米草群落。南汇边滩(30°51′43.20″~30°50′52.80″N,121°56′2.40″~121°51′57.59″E)位于上海南部,杭州湾北部,受长江口径流及杭州湾北岸水流影响大,其优越的地理位置使其同时受到长江带来的丰富泥沙和杭州湾北部潮流的影响,形成了广阔的砂质潮滩,主要分布有海三棱藨草群落和芦苇群落。
-
潮滩盐沼植物光谱测量样带与样方设置需充分考虑潮滩植物生态学特征,植被类型、形态决定其光谱反射特征,同时又受到物候现象与生长环境的影响。不同季相潮滩盐沼植被的光谱反射差异比较明显。春季为芦苇和互花米草的生长期,光谱上表现出绿色植物的特征,而海三棱藨草由于较晚进入生长期,绝大多数呈枯黄,其中低盖度的光谱特征类似光滩;夏至初秋为潮滩盐沼植被的生长旺季,在该物候期测量的光谱数据更具代表性;秋末冬初,海三棱藨草的“红边”特征几乎消失,芦苇和互花米草的“红边”陡坡大幅下降[23]。
本研究选择盐沼植被生长茂盛期进行光谱反射特征研究,分别在长江口崇明东滩、崇明北八滧和南汇边滩等3个潮滩湿地进行野外光谱测量。其中崇明北八滧测点样方13个,崇明东滩测点样方39个,南汇测点样方30个。样带设置采取由陆向海方向设置样方,样方大小为1 m × 1 m,并在崇明东滩设置水淹实验样方4个。样方设置充分考虑植被盖度、植被类型、土壤湿度、土壤类型等生态因子和环境因子。盐沼植被类型主要为高潮滩的芦苇、中低潮滩的海三棱藨草、互花米草。土壤类型有粉砂、极细砂、细粉砂这3种长江口潮滩典型土壤类型。低潮滩光滩由于长时间被海水覆盖湿度较高,故在光滩沿潮滩土壤湿度梯度设置光谱采样点。
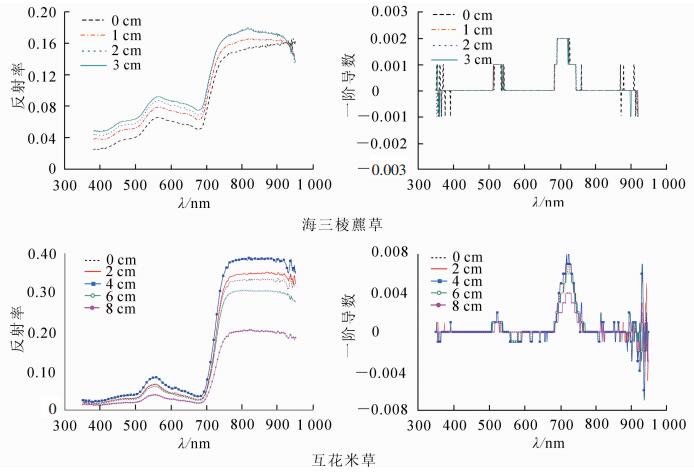
考虑到潮滩湿地盐沼植被测量中潮汐水位高低对植被冠层光谱反射的影响比较复杂,因此对崇明东滩潮滩前沿的海三棱藨草和互花米草进行不同潮位情况模拟实验。通过对同一样方盐沼植被加水来模拟涨潮,获取在其他外部条件完全一致的情形下盐沼植被受不同潮汐影响时的光谱特征。根据海三棱藨草(10 cm)和互花米草(20 cm)的植株高度设置潮位,分别为0,1,2,3 cm和0,2,4,6,8 cm。
-
测量仪器采用美国ASD公司(Analytical Spectral Devices Inc)生产的手持式野外地物光谱仪,测量波长为325~1 075 nm,波长取样间隔为1.4 nm,光谱分辨率为3 nm,视场角为25°。野外测量选择晴朗无风的条件下,在9:00-14:00进行。各个样方测量2~3次,测量数据15~20个·次-1。根据天气状况及测量点的改变,间隔2~3 min进行环境光谱校正。为使测量数据能与卫星影像、航空影像进行比较,测量时传感器探头垂直向下。同时测量人员面向阳光入射方向,避免阴影落在样方区域。
每个样方测量数据包括经纬度,土壤湿度,植被平均高度,天气情况(包括云量、风速),光谱反射率等。土壤湿度测量使用英国Delta-T公司生产的WET土壤三参数仪,在样方区域均匀选取5个点,分别测量每个点的湿度后取平均值。同时,使用手持式全球定位系统(GPS)测定样方的经纬度坐标。通过相机拍摄样方内植被覆盖情况,利用ArcGIS 10.1的监督分类功能计算植被覆盖像元数与总像元数,得到植被盖度。将ASD地物光谱仪测量的光谱数据通过HH2 Sync导出,并在Viewspec Pro 6.2.0软件里对测量数据进行处理。考虑到仪器噪声为保证测量数据的精确性,仅保留350~950 nm范围内的光谱数据。
-
光谱分析主要分为以下几个部分:①分析同等盖度不同植被类型的光谱反射特征,分别对样方盖度为60%±5%的海三棱藨草、芦苇、互花米草的光谱特征进行分析;②分析同种植被不同盖度情况下的光谱反射特征,对海三棱藨草、芦苇、互花米草在不同盖度条件下的光谱特征进行分析,植被盖度受其生长环境影响大,实际测量盐沼的植被盖度见表 1;③分析在不同土壤背景下盐沼的光谱特征,选择长江口3种典型盐沼植被及其下垫面土壤进行分析(表 2);④分析潮滩前沿地带海三棱藨草和互花米草在不同潮位影响下的光谱特征。潮滩前沿植被受潮位影响最为显著,前沿地带海三棱藨草植株高度多为6~12 cm,植被盖度较低,通常为10%~30%。该处互花米草的植株高度为15~24 cm,植被盖度较海三棱藨草要高,一般为20%~45%。本研究分析主要选择潮滩前沿具有代表性的海三棱藨草群落(植株高度为10 cm,盖度为30%)和互花米草群落(植株高度为20 cm,盖度为40%)。根据植株的高度设置实验潮位。具体实验潮位见表 3。
表 1 不同盖度植被样地
Table 1. Sample plots of different coverage saltmarsh
芦苇 海三棱藨草 互花米草 植被盖度/% 样地数/个 植被盖度/% 样地数/个 植被盖度/% 样地数/个 60 4 10 12 50 5 70 10 30 15 60 7 90 6 65 7 70 8 表 2 不同下垫面土壤植被样地数
Table 2. Sample plots of different underlying
测量地点 样地数/个 芦苇 海三棱藨草 互花米草 崇明东滩 14 20 0 南汇边滩 6 9 0 崇明北八滧 0 5 20 表 3 不同潮位植被样地
Table 3. Sample plots of different tidal
组别 海水淹没深度/cm 海三棱藨草 互花米草 1 0 0 2 1 2 3 2 4 4 3 6 5 8 光谱分析采用光谱曲线法和一阶导数法,光谱曲线主要反映在350~950 nm波段不同地物的反射和吸收特征,一阶导数主要用于分析在350~950 nm波段地物的反射和吸收变化速率,增强植被光谱变化特征。一阶导数正值越大表明光谱反射率增速越大。相关性分析法用来衡量2个变量之间的密切程度,通过相关性分析确定植被盖度、土壤粒径、潮位与盐沼植被反射光谱之间的密切程度[24]。在软件Origin 9.0和Excel中进行处理分析。
-
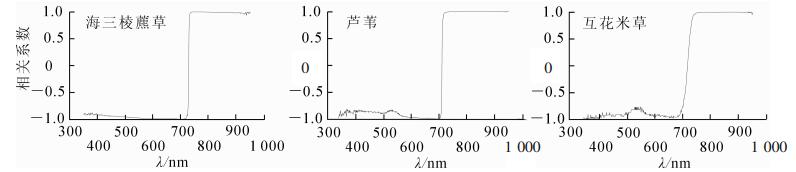
图 1是生长季节长江口3种主要潮滩盐沼植被海三棱藨草、芦苇和互花米草约60%盖度下的光谱反射和一阶导数曲线。在波长550 nm附近海三棱藨草、芦苇和互花米草均有1个次反射峰值,在波长680 nm附近有1个吸收谷,且光谱反射率均小于0.05。这是由于叶片中的叶绿素发生光合作用,在此处强烈吸收。反射率到波长700 nm附近急剧增大,增长速率同时也增大。在350~950 nm波长范围内,这3种潮滩盐沼植被的光谱特征差异很大。一阶导数图可以看出互花米草在“红边”位置(700~750 nm)的反射率增速高于另外2种潮滩盐沼植被。在533~560和683~751 nm波长范围内,这3种潮滩盐沼植被的光谱差异相对明显。

图 1 相近盖度不同盐沼植被光谱反射率曲线及其一阶导数
Figure 1. Curve map of spectral reflectance and first derivative of different saltmarsh with similar coverage
-
在崇明东滩选择相近土壤湿度不同盖度的潮滩盐沼植被进行光谱测量,结果见图 2。海三棱藨草、芦苇、互花米草在350~730 nm波段光谱反射率随盖度增大而减小。到730 nm后3种植被反射率随盖度增大而增大,且不同盖度的芦苇和互花米草光谱反射率变化趋势同海三棱藨草基本一致。由反射光谱一阶导数曲线得到3种盐沼植被在绿峰波段(519~528 nm),红谷波段(675~685 nm),红边波段(727~738 nm)有明显的峰、谷特点。绿峰波段是叶绿素反射区域,盖度高的叶绿素反射作用强。位于680 nm附近的波谷是叶绿素吸收红光形成的,植被盖度越高在该处的吸收作用越强,反射率越低。盐沼植被在红边的增速越快,表明该种群的生长环境比较适宜。这些波段能很好地反映反射光谱随植被盖度的变化特征,是判断植被盖度的有效波段范围。

图 2 不同盖度下盐沼植被的光谱反射率曲线及其一阶导数
Figure 2. Curve map of spectral reflectance and first derivative of saltmarsh with different coverages
从图 3可知:在可见光波段,反射率与植被盖度呈负相关,主要是在该波段范围土壤的高反射及植被在可见光波段有强烈吸收作用;在近红外波段,反射率与植被盖度呈正相关性,相关性绝对值均接近1,是由于植被在近红外波段高反射率且反射率随植被盖度的增大而增大。故盐沼植被盖度对其反射光谱影响很大。当利用遥感手段对盐沼植被进行信息提取时,不可忽略植被盖度的影响。

图 3 不同盖度下盐沼光谱相关性分析
Figure 3. Spectral correlation analysis of saltmarsh with different coverages
-
从图 4的样地土壤反射光谱曲线可以看出:3种土壤反射光谱总体呈现随波长增大反射率增大的趋势,在350~600 nm波长范围内,土壤的反射率随波长增大而增大的趋势明显;在波长600 nm附近出现第1个反射峰;到波长600 nm后,土壤反射光谱随波长变化反射率变化不明显,呈缓慢增大或减小的趋势。

图 4 不同土壤的光谱反射率曲线及其一阶导数曲线
Figure 4. Curve map of spectral reflectance and first derivative of different kinds of soil
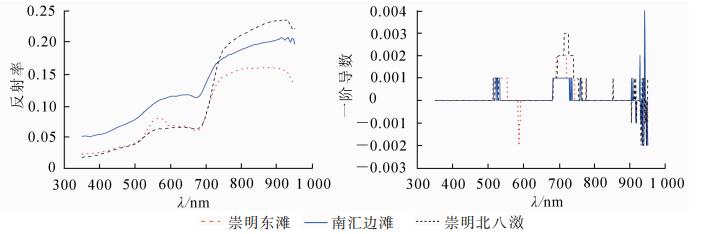
崇明东滩、南汇边滩、崇明北八滧3个研究区的土壤光谱反射特征有明显差异,相同盖度的同种植被在不同样地的光谱反射率表现出较大差异,不论是低盖度还是中等盖度,在可见光波段,海三棱藨草光谱反射率均表现为南汇边滩的最高,崇明北八滧的最低,崇明东滩的居中;近红外波段则反之;可以认为盐沼植被的这种光谱差异很大程度上是由下垫面土壤导致的。
低盖度30%±5%海三棱藨草光谱曲线(图 5)整体呈随波长增大反射率增大的趋势,只在红光波段出现小的波谷,与健康植被光谱曲线相比,植被光谱特征不明显。图 5显示:崇明北八滧和南汇边滩的低盖度海三棱藨草光谱曲线均未出现绿光反射峰,仅有崇明东滩的绿峰和红谷2个植被特征均呈现。而3个样带的中等盖度60%±5%海三棱藨草光谱曲线(图 6)都呈健康植被反射率特征。对于不同的样带,土壤下垫面对植被光谱影响程度不同。

图 5 低盖度海三棱藨草在不同潮滩土壤上的光谱反射率曲线及一阶导数曲线
Figure 5. Curve map of spectral reflectance and first derivative of low coverage Scirpus mariqueter in different tidal flat

图 6 中等盖度海三棱藨草在不同潮滩土壤上的光谱反射率曲线及其一阶导数曲线
Figure 6. Curve map of spectral reflectance and first derivative of medium coverage Scirpus mariqueter in different tidal flat
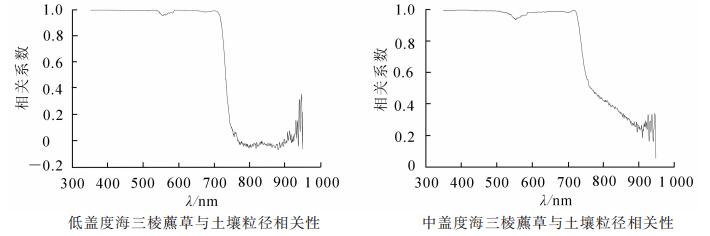
从图 7相关性分析中可以看出:在可见光波段海三棱藨草光谱反射率与土壤下垫面的相关性更高,相关系数在0.94~0.99范围内波动,相关系数在波长550 nm附近有一个最小值,是由于550 nm为绿色植被的“绿峰”,表现为盐沼植被叶绿素的强反射作用,土壤的影响相对较小;相关性到波长750 nm后明显下降,与土壤到750 nm后光谱反射率趋于稳定有关,近红外波段是植被的敏感波段,主要由植被自身生长状况决定。

图 7 不同潮滩土壤背景下海三棱藨草光谱相关性分析
Figure 7. Spectral correlation analysis of Scirpus mariqueter in different tidal flat
由上述结果分析可见:土壤下垫面对海三棱藨草光谱反射率在可见光波段的影响较大,而在近红外波段的影响则较小;海三棱藨草盖度越小,其反射光谱受土壤下垫面影响越大,当盖度过小时,海三棱藨草反射光谱的植被特征将消失,并呈现土壤的反射光谱特点。
-
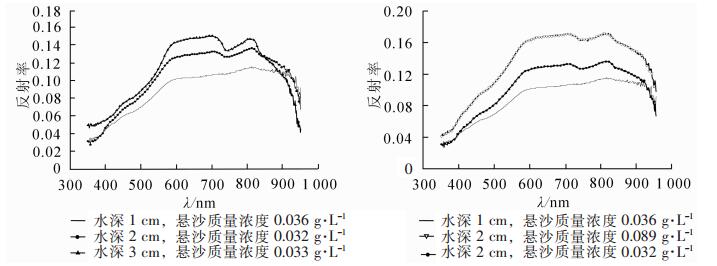
潮滩盐沼植被由于受到周期性水文过程影响,对涨潮时潮滩水体进行的光谱测量发现,光谱反射率随水深的增加增大,但在近红外波段水深大的反射率下降速率明显大于水深浅的水体;相同水深,悬沙质量浓度大的反射率大(图 8)。对研究区潮滩前沿的海三棱藨草群落和互花米草群落受不同潮位影响时的光谱特征进行分析,水体悬沙质量浓度控制在0.032~0.036 g·L-1范围内。

图 8 崇明东滩不同水深和悬沙浓度水体的光谱反射率曲线
Figure 8. Curve map of spectral reflectance of water with changing water level and suspended sediment concentration
如图 9所示:在350~950 nm波长范围内,不同潮位海三棱藨草反射率曲线走势大体一致,并随潮位的增加反射率增大。波段830~950 nm,表面有水体覆盖的海三棱藨草反射率随波长增大而呈下降趋势,与830~950 nm波段水体的光谱特征相似,而表面没有水体淹没的在近红外波段仍表现为反射率随波长的增大而上升趋势,与健康植被反射光谱特征相同。潮位为2和3 cm时的海三棱藨草在920 nm附近反射率几乎完全重合。由潮位和海三棱藨草植被反射光谱之间的相关性分析可以看出,两者相关性相当高,350~950 nm波段的相关系数绝对值基本都在0.97以上(图 10)。

图 9 盐沼植被在不同潮位下的光谱反射率曲线及一阶导数曲线
Figure 9. Curve map of spectral reflectance and first derivative of saltmarsh with changing water level

图 10 不同潮位与盐沼植被光谱相关性分析
Figure 10. Spectral correlation analysis of saltmarsh with changing water level
盖度相对较高的互花米草群落在潮位在0~4 cm时,有潮汐影响的反射光谱高于无潮汐影响的,且反射光谱随潮位升高而增加。当潮位超过4 cm后,反射光谱均小于无潮汐时的反射光谱,呈现反射光谱随潮位升高而下降的趋势。在可见光和近红外波段,互花米草群落的反射率与潮位均有一定相关性,为-0.37~-0.78,当波长超过700 nm后,两者间的相关系数值更大,即反射光谱与近红外波段潮位之间相关性更高(图 10)。互花米草植被的反射光谱及其与潮位相关性分析结果显示,当下垫面被较薄水体覆盖发生镜面反射时,植被反射光谱随潮位升高而增大。而当潮位超过一定值时,出露的植被较少,传感器视野范围内主要为水体和湿土下垫面,植被反射光谱受到水体下垫面的影响较大,表现为下垫面的光谱特征,植被反射光谱随潮位升高而下降。
在潮汐影响下,潮位变化极大地影响了盐沼植被光谱反射率。当土壤表面有较薄水体覆盖时,植被光谱反射率在350~950 nm范围内随潮位升高而增大;潮位继续升高到将植株逐渐淹没后,光谱反射率急剧下降。由于海三棱藨草和互花米草的植被盖度、植株高度不同,光谱反射率下降的潮位也不同。在相同潮位下,植株高、盖度高的植被露出水面的部分更多,传感器获得更多的植被光谱信息。海三棱藨草光谱反射率与潮位间的相关性高于互花米草,还受到两者植株形态的影响。
-
本研究利用ASD手持式地物光谱仪测量了长江口不同盐沼植被的反射光谱,针对植被生长茂盛期,分析不同潮滩土壤、潮位、植被盖度条件下的盐沼植被光谱反射特征,主要得出以下结论:①不同类型盐沼植被的光谱反射率差异较大,这种差异主要是植被本身和下垫面共同造成的。②盐沼植被的反射光谱受盖度的影响极大,在可见光波段两者呈负相关,在近红外波段两者呈正相关,相关系数接近0.99。盐沼植被的反射率在可见光波段随盖度增大而降低,在近红外波段则相反。③土壤下垫面对盐沼植被反射光谱影响极大。植被盖度越小,土壤下垫面影响越大。土壤下垫面对植被光谱反射率在可见光波段的影响较大,而在近红外波段的影响则较小。④盐沼植被反射光谱与潮位相关性显著。当潮位较低时,植被反射率高于无潮汐时的反射率,且呈随潮位升高反射率增大的趋势。当潮位上涨到一定高度,植被反射率低于无潮汐时的反射率,且随潮位的升高植被反射率呈下降的趋势。
-
本研究对不同潮位与植被反射光谱的相关性与刘光等[24]研究不同水深对水生植被冠层光谱特征影响得到的结论是一致的,可见光波段和近红外波段水深对植被光谱反射率的影响有明显差异。从植被和水体的光谱特征分析,水体的反射主要在蓝绿波段,在近红外波段吸收作用强;植被叶绿素对蓝光和红光吸收作用强,对绿光反射作用强,其叶细胞结构对近红外有高反射,所以在不同波段范围,对水深的波谱响应不同。同理,盖度对植被光谱特征的影响包括了植被自身叶绿素含量等因素。同一植被盖度高的叶绿素含量高,在可见光红波段的吸收作用就更强,金仲辉[26]在研究绿色植物光谱特征时也得到这样的结果。对土壤下垫面的分析结果,土壤在可见光和近红外波段变化幅度不大,不同土壤背景对于盐沼植被光谱反射率是有差异的。故在陆生植被的遥感指数研究中,HUETE[27]提出了土壤调节植被指数,来修正下垫面土壤的影响。
本研究在设计中没有考虑土壤盐度等对盐沼植被光谱的影响,在潮位对盐沼植被光谱影响中也没有设计不同悬沙浓度与植被光谱反射之间的关系。因此今后的研究将充分考虑土壤和水体中的具体影响因子,结合室内控制实验,对盐沼植被光谱与其影响因子之间的相应关系进行定量研究,建立盐沼植被反射光谱模型。
Spectral analysis of an intertidal saltmarsh in the Yangtze Estuary
-
摘要: 以长江口典型盐沼植被为研究对象,考虑不同盐沼植被类型、盖度、土壤类型、土壤湿度以及高中低潮滩地形分布因素,分别在崇明东滩国家级自然保护区、崇明北湖边滩、南汇边滩设置光谱测量样带和样方,使用ASD光谱仪进行现场光谱测量,分析不同植被盖度、土壤下垫面、潮位条件下典型盐沼植被的光谱特征反射曲线。研究结果表明:同一类型盐沼植被的光谱反射率差异是由植被盖度和下垫面共同导致的。在相同土壤背景下,光谱反射率与植被盖度在可见光波段相关系数接近-1.00,呈负相关,在近红外波段相关系数接近0.99,呈正相关。不同潮滩土壤背景,盐沼植被光谱反射率有明显差异。在可见光波段,土壤下垫面对植被光谱反射率影响较大,而在近红外波段的影响则较小。植被盖度越小,其反射光谱受土壤下垫面影响越大。盐沼植被与潮位的相关系数最大可达0.97,低潮位时植被反射光谱高于无潮汐时的光谱,且呈现随潮位升高反射率增大的趋势;当潮位上涨到一定高度,植被反射光谱低于无潮汐时的光谱,且呈现随潮位的升高而下降的趋势。Abstract: To consider different vegetation types, coverage, soil types, soil moisture, and gradient elevation of a tidal flat so as to determine the effect of the tide, three typical saltmarsh vegetation types in the Yangtze Estuary were chosen as research objects. Vegetation transects and samples(1 m×1 m) were selected for the above factors, and the growth environment of the saltmarsh was simulated. An ASD Fieldspec HandHeld 2 was used to measure the spectrum of the saltmarsh vegetation canopy; then the spectral reflectance and the first derivative (350-950 nm) were calculated, and correlations between coverage/soil/tidal level and spectral reflectance were analyzed. Results showed that vegetation coverage and the underlying surface greatly affected spectral reflectance of the saltmarsh. For the same soil conditions, spectral reflectance of different saltmarsh vegetation coverages in the visible light band was negatively correlated (r≈-1.00) with coverage and in the near infrared positively correlated (r≈0.99). For both low coverages and high coverages, spectral reflectance of different soil types in the visible light band was positively correlated (r≈0.97) but in the near infrared was less correlated. The saltmarsh and tidal level were significantly correlated (r≈0.97). Also spectral reflectance of the saltmarsh at high tide was higher than at low tide, and the reflectance increased as the tide rose. When the tide level rose to a certain height, spectral reflectance of the saltmarsh was lower than without a tide, and this generally decreased as the tide level fell. The effect of vetegation coverage, underlying(soil types and soil moisture) should not be ignored when monitoring saltmarsh vegetation.
-
Key words:
- plant ecology /
- saltmarsh /
- spectral reflectance /
- tide level /
- soil /
- Yangtze Estuary
-
图 1 相近盖度不同盐沼植被光谱反射率曲线及其一阶导数
Figure 1 Curve map of spectral reflectance and first derivative of different saltmarsh with similar coverage
图 2 不同盖度下盐沼植被的光谱反射率曲线及其一阶导数
Figure 2 Curve map of spectral reflectance and first derivative of saltmarsh with different coverages
图 3 不同盖度下盐沼光谱相关性分析
Figure 3 Spectral correlation analysis of saltmarsh with different coverages
图 4 不同土壤的光谱反射率曲线及其一阶导数曲线
Figure 4 Curve map of spectral reflectance and first derivative of different kinds of soil
图 5 低盖度海三棱藨草在不同潮滩土壤上的光谱反射率曲线及一阶导数曲线
Figure 5 Curve map of spectral reflectance and first derivative of low coverage Scirpus mariqueter in different tidal flat
图 6 中等盖度海三棱藨草在不同潮滩土壤上的光谱反射率曲线及其一阶导数曲线
Figure 6 Curve map of spectral reflectance and first derivative of medium coverage Scirpus mariqueter in different tidal flat
图 7 不同潮滩土壤背景下海三棱藨草光谱相关性分析
Figure 7 Spectral correlation analysis of Scirpus mariqueter in different tidal flat
图 8 崇明东滩不同水深和悬沙浓度水体的光谱反射率曲线
Figure 8 Curve map of spectral reflectance of water with changing water level and suspended sediment concentration
图 9 盐沼植被在不同潮位下的光谱反射率曲线及一阶导数曲线
Figure 9 Curve map of spectral reflectance and first derivative of saltmarsh with changing water level
图 10 不同潮位与盐沼植被光谱相关性分析
Figure 10 Spectral correlation analysis of saltmarsh with changing water level
表 1 不同盖度植被样地
Table 1. Sample plots of different coverage saltmarsh
芦苇 海三棱藨草 互花米草 植被盖度/% 样地数/个 植被盖度/% 样地数/个 植被盖度/% 样地数/个 60 4 10 12 50 5 70 10 30 15 60 7 90 6 65 7 70 8  下载: 导出CSV
下载: 导出CSV
表 2 不同下垫面土壤植被样地数
Table 2. Sample plots of different underlying
测量地点 样地数/个 芦苇 海三棱藨草 互花米草 崇明东滩 14 20 0 南汇边滩 6 9 0 崇明北八滧 0 5 20
下载: 导出CSV
表 3 不同潮位植被样地
Table 3. Sample plots of different tidal
组别 海水淹没深度/cm 海三棱藨草 互花米草 1 0 0 2 1 2 3 2 4 4 3 6 5 8
下载: 导出CSV
-
[1] 陈吉余, 程和琴, 戴志军.滩涂湿地利用与保护的协调发展探讨:以上海市为例[J].中国工程科学, 2007, 9(6):11-17. CHEN Jiyu, CHENG Heqin, DAI Zhijun. Compatibility of utilization and protection of tidal flat and wetland:a case study in Shanghai Area[J]. Eng Sci, 2007, 9(6):11-17. [2] COSTANZA R, d'ARGE R, de GROOT R, et al. The value of the world's ecosystem services and natural capital[J]. Ecol Econ, 1998, 25(1):3-15. [3] 马安娜, 陆健健.湿地生态系统碳通量研究进展[J].湿地科学, 2008, 2(6):116-123. MA Anna, LU Jianjian. The progress of research on carbon flux in wetland ecosystems[J]. Wetland Sci, 2008, 2(6):116-123. [4] 胡嘉东, 郑丙辉, 万峻.潮间带湿地栖息地功能退化评价方法研究与应用[J].环境科学研究, 2009, 22(2):171-175. HU Jiadong, ZHENG Binghui, WAN Jun. Studies and application of an assessment model of the habitat function degradation in the intertidal wetland[J]. Res Environ Sci, 2009, 22(2):171-175. [5] 陈秀芝, 孙瑛.中国特有盐沼植物:海三棱藨草的现状及保护利用[J].湿地科学与管理, 2011, 7(1):60-63. CHEN Xiuzhi, SUN Ying. Chinese endemic salt marsh piant:bulrush(Scirpus mariqueter), current status of its conservation and utilization[J]. Wetlang Sci Manage, 2011, 7(1):60-63. [6] 张晓龙, 李培英, 李萍, 等.中国滨海湿地研究现状与展望[J].海洋科学进展, 2005, 23(1):87-95. ZHANG Xiaolong, LI Peiying, LI Ping, et al. Present conditions and prospects of study on coastal wetlands in China[J]. Adv Marine Sci, 2005, 23(1):87-95. [7] 李青山, 张华鹏, 崔勇, 等.湿地功能的研究进展[J].科学技术与工程, 2004, 4(11):41-42. LI Qingshan, ZHANG Huapeng, CUI Yong, et al. A prospect for study on functions of wetland[J]. Sci Tecchnol Eng, 2004, 4(11):41-42. [8] 季子修, 蒋自巽, 朱季文, 等.海平面上升对长江三角洲和苏北滨海平原海岸侵蚀的可能影响[J].地理学报, 1993, 4(6):516-526. JI Zixiu, JIANG Zixun, ZHU Jiwen, et al. Impacts of sea level rise on coastal erosion in the Changjiang Delata and North Jiangsu Coastal Plain[J]. Acta Geogr Sin, 1993, 4(6):516-526. [9] TIAN Bo, WU Wenting, YANG Zhaoqing, et al. Drivers, trends, and potential impacts of long-term coastal reclamation in China from 1985 to 2010[J]. Estuarine Coastal Shelf Sci, 2016, 170:83-90. [10] 黄华梅.上海滩涂盐沼植被的分布格局和时空动态研究[D].上海: 华东师范大学, 2009. HUANG Huamei. A Research on Spatial-temporal Dynamics of Salt Marsh Vegetation at the Intertidal Zone in Shanghai[D]. Shanghai: East China Normal University, 2009. [11] 朱春娇, 田波, 周云轩, 等.基于Markov和CLUE-S模型的浦东新区湿地演变遥感分析与预测[J].复旦学报(自然科学版), 2015, 54(4):431-438. ZHU Chunjiao, TIAN Bo, ZHOU Yunxuan, et al. Wetland change analysis and forecasting in Pudong New Area Using Markov and CLUE-S Model[J]. J Fudan Univ Nat Sci, 2015, 54(4):431-438. [12] 赖婷, 杨为民, 田波.遥感与GIS支持下杭州湾北岸湿地空间威胁性分析与评价研究[J].复旦学报(自然科学版), 2013, 52(3):356-362. LAI Ting, YANG Weimin, TIAN Bo. A GIS and remote sensing based analysis of wetland spatial threat in the North Hangzhou Bay[J]. J Fudan Univ Nat Sci, 2013, 52(3):356-362. [13] 黄华梅, 张利权, 袁琳.崇明东滩自然保护区盐沼植被的时空动态[J].生态学报, 2007, 27(10):4166-4172. HUANG Huamei, ZHANG Liquan, YUAN Lin. The spatio-temporal dynamics of salt marsh vegetation for Chongming Dongtan National Nature Reserve, Shanghai[J]. Acta Ecol Sin, 2007, 27(10):4166-4172. [14] 张杰.长江口潮滩植被检测及时空变化的遥感研究[D].上海: 华东师范大学, 2007. ZHANG Jie. Remote Sensing Study on Vegetation Detection and Temporal Change of Tidal Flat in the Yangtze River Estuary[D]. Shanghai: East China Normal University, 2007. [15] 刘克, 赵文吉, 郭逍宇, 等.野鸭湖典型湿地植物光谱特征[J].生态学报, 2010, 30(21):5853-5861. LIU Ke, ZHAO Wenji, GUO Xiaoyu, et al. Spectral bands of typical wetland vegetation in the Wild Duck Lake[J]. Acta Ecol Sin, 2010, 30(21):5853-5861. [16] GOVENDER M, CHETTY K, BULCOCK H. A review of hyperspectral remote sensing and its application in vegetation and water resource studies[J]. Water S A, 2007, 33(2):145-151. [17] BELLUCO E, CAMUFFO M, FERRARI S, et al. Mapping salt-marsh vegetation by multispectral and hyperspectral remote sensing[J]. Remote Sense Environ, 2006, 105(1):54-67. [18] 刘建平, 赵英时.高光谱遥感数据解译的最佳波段选择方法研究[J].中国科学院大学学报, 1999, 16(2):153-161. LIU Jianping, ZHAO Yingshi. Methods on optimal bands seletion in hyperspectral remote sensing data interpretation[J]. J Grad Sch Acad Sin, 1999, 16(2):153-161. [19] SCHMIDT K S, SKIDMORE A K. Spectral discrimination of vegetation types in a coastal wetland[J]. Remote Sens Environ, 2003, 85(1):92-108. [20] 林川, 宫兆宁, 赵文吉, 等.基于光谱特征变量的湿地典型植物生态类型识别方法:以北京野鸭湖湿地为例[J].生态学报, 2013, 33(4):1172-1185. LIN Chuan, GONG Zhaoning, ZHAO Wenji, et al. Identifying typical plant ecological types based on spectral characteristic variables:a case study in Wild Duck Lake wetland, Beijing[J]. Acta Ecol Sin, 2013, 33(4):1172-1185. [21] 李亚丹, 杜华强, 周国模, 等.雷竹叶绿素与高光谱植被指数关系及其反演模型[J].浙江农林大学学报, 2015, 32(3):335-345. LI Yadan, DU Huaqiang, ZHOU Guomo, et al. Chlorophyll content in Phyllostachys violascens related to hyper-spectral vegetation indices and development of an inversion model[J]. J Zhejiang A & F Univ, 2015, 32(3):335-345. [22] 高占国, 张利权.上海盐沼植被的多季相地面光谱测量与分析[J].生态学报, 2006, 26(3):793-800. GAO Zhanguo, ZHANG Liquan. Measuring and analyzing of the multi-seasonal spectral characteristics for saltmarsh vegetation in Shanghai[J]. Acta Ecol Sin, 2006, 26(3):793-800. [23] 高占国.长江口盐沼植被的光谱特征研究[D].上海: 华东师范大学, 2006. GAO Zhanguo. Spectral Characteristics of Salt Marsh Vegetation in the Yangtze River Estuary[D]. Shanghai: East China Normal University, 2006. [24] 刘光, 唐鹏, 蔡占庆, 等.水体背景对湿地水生植物冠层光谱影响研究[J].光谱学与光谱分析, 2015, 35(10):2970-2976. LIU Guang, TANG Peng, CAI Zhanqing, et al. A study on effect of water background on canopy spectral of wetland aquatic plant[J]. Spectrosc Spectrol Anal, 2015, 35(10):2970-2976. [25] 舒敏彦.海岸带盐沼植被指数构建研究[D].上海: 华东师范大学, 2017. SHU Minyan. A New Remote Sensing Vegetation Index for Coastal Saltmarsh Wetland[D]. Shanghai: East China Normal University, 2017 [26] 金仲辉.绿色植物反射光谱的特征及其在监测农作物生长中的应用[J].物理, 1993, 22(11):673-678. JIN Zhonghui. Characteristics of reflectance spectra for green plants and their application in inspecting the growth of the crop[J]. Physics, 1993, 22(11):673-678. [27] HUETE A R. A soil-adjusted vegetation index (SAVI)[J]. Remote Sense Environ, 1988, 25(3):295-309 -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.01.014
 点击查看大图
点击查看大图

计量
- 文章访问数: 4237
- HTML全文浏览量: 1036
- PDF下载量: 276
- 被引次数: 0
