







-
定向刨花板(oriented strand board, OSB)是以大片木质刨花为原料,经施胶并定向铺装后热压而成。OSB是一种生物基材料,水分会软化木纤维并影响OSB的力学强度[1-3]。木纤维的干缩湿胀易造成OSB内部结构变化[4-5]。已有研究[6-7]表明:相较于65%相对空气湿度(relative humidity, RH),85%RH条件下OSB的弹性模量、静曲强度更低,且变异性增加。通过开展循环吸放水实验,LI等[8]发现多次吸放水会导致OSB变形,并降低其弹性模量。HODOUSEK等[9]研究表明:吸水及含水率增加均会降低OSB的透气性。提高OSB的耐水性和力学强度是拓展其应用范围的重要途径。木材材性在很大程度上决定了OSB的物理力学性能[10],疏水化改性木材刨花可以极大提高OSB的耐水性[11],石蜡等耐水添加剂的使用也能够改善OSB的尺寸稳定性[12]。吸湿影响OSB物理性能的直观表现是厚度膨胀,尺寸稳定性也是衡量OSB耐水性的重要指标,厚度方向上的尺寸变化会直接影响OSB的抗压强度,理解OSB抗压强度变化的发生机制有助于提高OSB的耐水性。数字散斑相关分析技术是一种动态记录材料面内应变分布的可靠手段[13]。该技术通过分析由高速相机捕捉的一系列照片来获得材料的表面变形,并进一步量化面内应变分布[14]。数字散斑相关分析技术可以有效记录载荷作用下木质材料的破坏过程和探索力学损失的发生机制[15-17]。本研究旨在系统探索水分对OSB抗压强度的影响规律,并基于应变分布分析影响规律的发生机制,研究结果可以为优化OSB加工工艺提供必要的理论支撑。
-
2类OSB购自加拿大Norbord公司,分别采用杨木Populus euramericana刨花和异氰酸酯胶黏剂为原料生产,气干条件下厚度和密度分别为13.0 mm、594.4 kg·m−3和19.0 mm、605.1 kg·m−3。每种OSB制备36个截面尺寸为(35.1±0.2) mm×(35.1±0.2) mm的试件,将每种厚度的试件均分为6组,对应不同的水分条件。水分条件包括对照组,(65±3)%相对湿度(RH);A,(83±3)%RH;B,(95±3)%RH;C,1次吸放水循环;D,2次吸放水循环;E,8次吸放水循环。1次吸放水循环将试件置于(20±1) ℃水中1 h,后取出放在40 ℃的烘箱中干燥2 h,之后在(65±3)%RH和(20±1) ℃条件下陈放2 d。这一干燥条件旨在模拟实际应用环境下的风干条件。每次吸放水循环前后,采集试件质量和厚度信息。将3组试件分别在(20±1) ℃和3种不同空气湿度条件下陈放4周,采集陈放前后试件质量和厚度信息。力学测试完成后,记录所有试件的绝干质量。
-
压缩试验对试件质量的影响忽略不计,计算试件含水率: CM=[(m−m0)/m0]×100%。其中:CM是试件的含水率(%),m是试件的质量(g),m0是试件的绝干质量(g)。不同水分条件下试件的厚度变化CT=(T−T0)×100%/T0。其中:CT是试件的厚度变化(%),T是不同水分条件下试件的厚度(mm),T0是对照条件下试件的厚度(mm)。试件压缩强度SC=F/S。其中:SC是试件的压缩强度(N·mm−2),F是加载载荷(N),S是试件的接触面积(mm2)。
试件沿厚度方向变形时,应变正值和负值分别表示拉伸应变和压缩应变。剪切应变是试件发生倾斜滑移产生的变形,剪切应变的正值和负值分别表示顺时针剪切和逆时针剪切。试件沿厚度方向每毫米的应变分布,根据
${{S}}=\left(\displaystyle \sum_{{i}}^{{{i}} + 1} {{{P_{{\rm{S}}i}}}} \right)/{{n}}$ 计算获得。试件沿厚度方向的剪切应变分布,根据${{{{S}}_{{{{{\rm{S}}}}}}}}=\left(\displaystyle \sum_{{i}}^{{{i}} + 1}\left| {{{P_{{\rm{SS}}i}}}} \right| \right)/{{n}}$ 计算得出。其中:S是厚度方向的平均压缩应变,$P_{{\rm{S}}i} $ 是厚度i和 (i+1) 间的压缩应变,SS是厚度i和 (i+1) 间的平均剪切应变,PSSi是厚度i和 (i+1) 间的剪切应变,n是单位厚度间的应变值个数。 -
使用细砂纸轻轻砂光OSB试件一个侧面,后用0.5 mm黑色中性笔绘制散斑。为防止吸放水过程影响散斑质量,C、D、E组试件的散斑是在吸放水过程后绘制。A、B组试件的散斑是在陈放前绘制。
本研究聚焦于试件的压缩强度测试,使用Instron万能力学试验机对试件平面加压。加载过程中压头以0.2 mm·min−1匀速位移,每100 ms记录1次压头高度(±0.001 mm)和载荷(±0.001 N)。加载流程:手动调整压头初始载荷至980.0 N (0.8 N·mm−2),之后开始匀速自动加载,载荷达到8 575.0 N (7.0 N·mm−2)时加载结束。自动加载过程中,使用数字散斑相关分析技术同步记录试件表面应变分布。具体为使用高速摄像机(M2514-MP2)每100 ms拍摄1张试件表面照片,并使用Correlated Solutions软件计算散斑位置变化和应变分布。
-
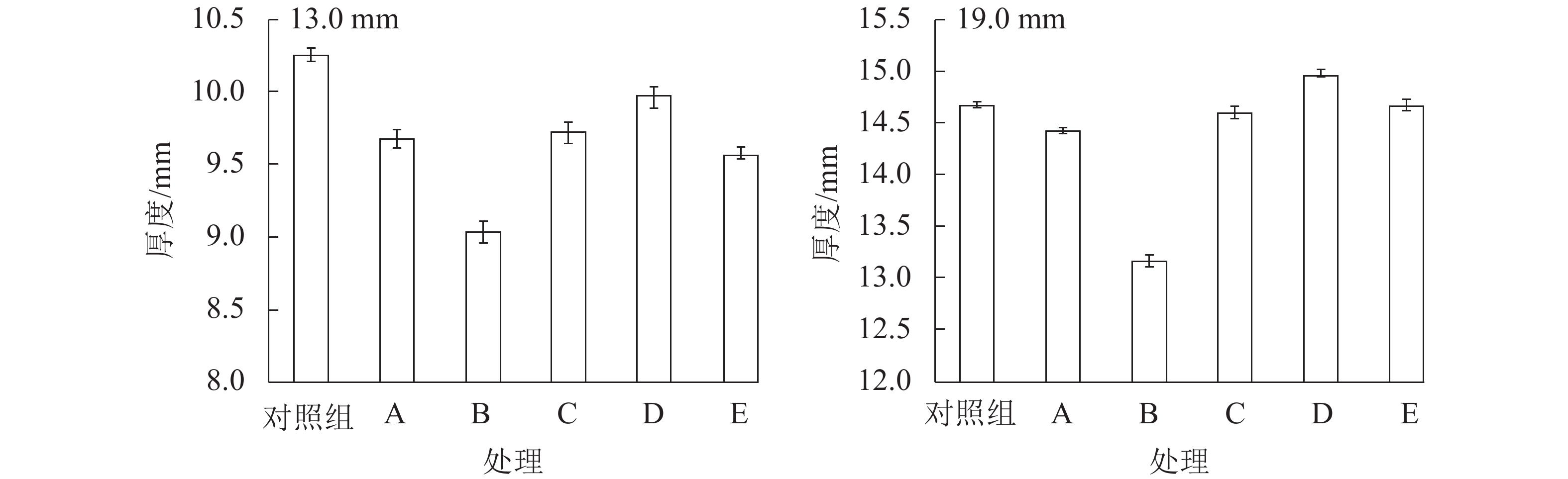
如图1所示:相较于C组试件,E组试件的厚度变化更大,这说明多次吸放水会引起试件内部结构变化。刨花的干缩湿胀会释放固定在OSB内部的热压应力,并使得刨花间孔隙扩大[4]。吸放水循环8次后,E组试件13.0和19.0 mm的含水率分别升高4.38%和3.81%,这归因于吸湿滞后现象。含水率升高是造成多次吸放水循环后试件厚度增加的另一重要原因。吸放水循环会增加样品的厚度,循环次数与厚度变化正相关。相较于其他试件,B组试件的含水率最高,厚度变化也最大。
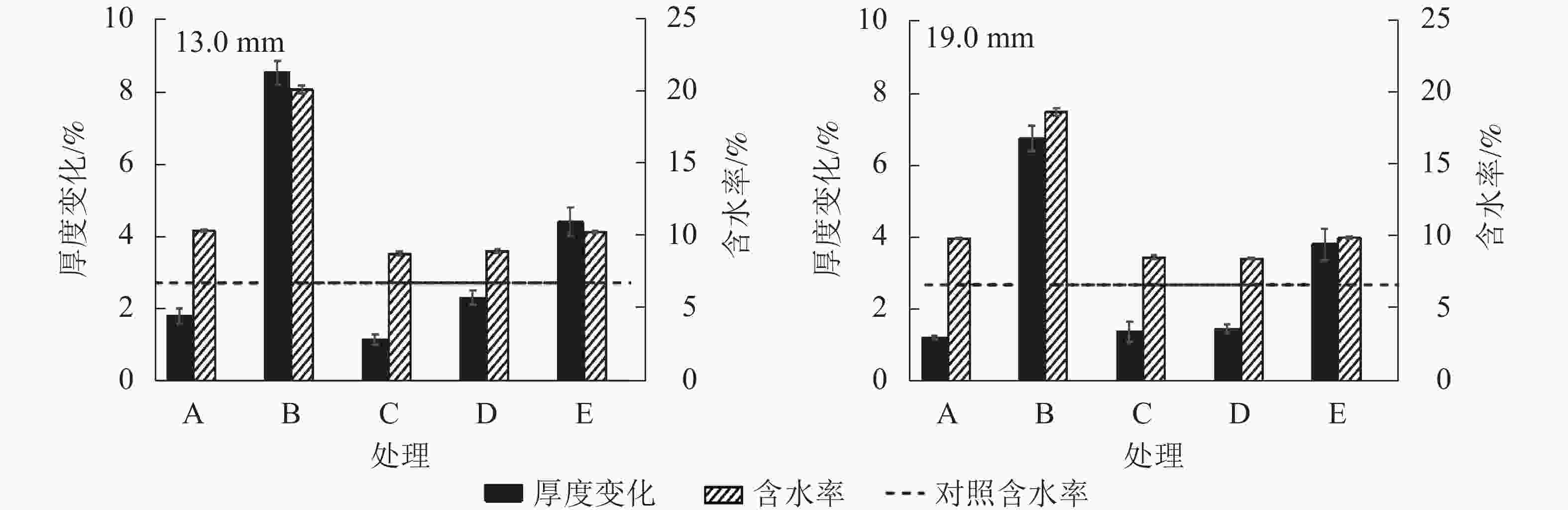
图 1 不同水分条件下试件的厚度变化和含水率
Figure 1. Thickness change and moisture content of samples at different water situations
-
表1显示不同水分条件下试件变形的差异性,试件变形(加压头位移)越大代表试件压缩强度越低。其中,B组试件的变形显著增加(P<0.05),说明(95±3)%RH条件下OSB的压缩强度下降显著(P<0.05)。E组试件也发生显著变形,说明多次吸放水循环同样会明显降低OSB的压缩强度。相较于19.0 mm试件,13.0 mm试件更易受环境湿度和吸放水循环影响。试件厚度变化比例大是13.0 mm试件位移显著增加的重要原因。具体表现为(83±3)%RH条件下和1个吸放水循环后,位移已经有显著性增加。较大的原始厚度使得19.0 mm试件厚度变化比例较小,因此仅在(95±3)%RH条件下和8个吸放水循环后位移会有显著性增加。
表 1 各组试件在 7 MPa载荷条件下变形的t检验结果
Table 1. T-test results of the displacement at 7 MPa
厚度/mm P 对照组/A 对照组/B 对照组/C 对照组/D 对照组/E 13.0 7.87E−03* 6.91E−06* 2.56E−02* 7.32E−02 3.05E−04* 19.0 1.26E−01 1.47E−05* 4.53E−01 9.36E−01 9.39E−03* 说明:*表示差异显著(P<0.05) 图2表明:7 MPa载荷条件下,B组试件的最终厚度最小。这说明含水率增加软化了木材刨花,并造成载荷条件下木材刨花的大幅变形。吸放水循环后,厚度为13.0 mm试件的最终厚度均比对照组低。吸湿会软化刨花纤维,吸放水循环可以引起刨花间孔隙放大和扩散,这均会降低OSB试件的压缩强度[18]。以上理论可以解释13.0 mm试件的最终厚度结果,但并不能完全解释19.0 mm试件的最终厚度变化。与对照试件相比,吸放水循环后,载荷条件下试件的最终厚度甚至小幅增加。该现象的原因是吸放水循环释放了热压时聚集在试件内部的应力,这使得试件压缩变形困难。厚OSB加工成型往往需要更大的压力,因此吸放水循环释放出的内部应力也更大。
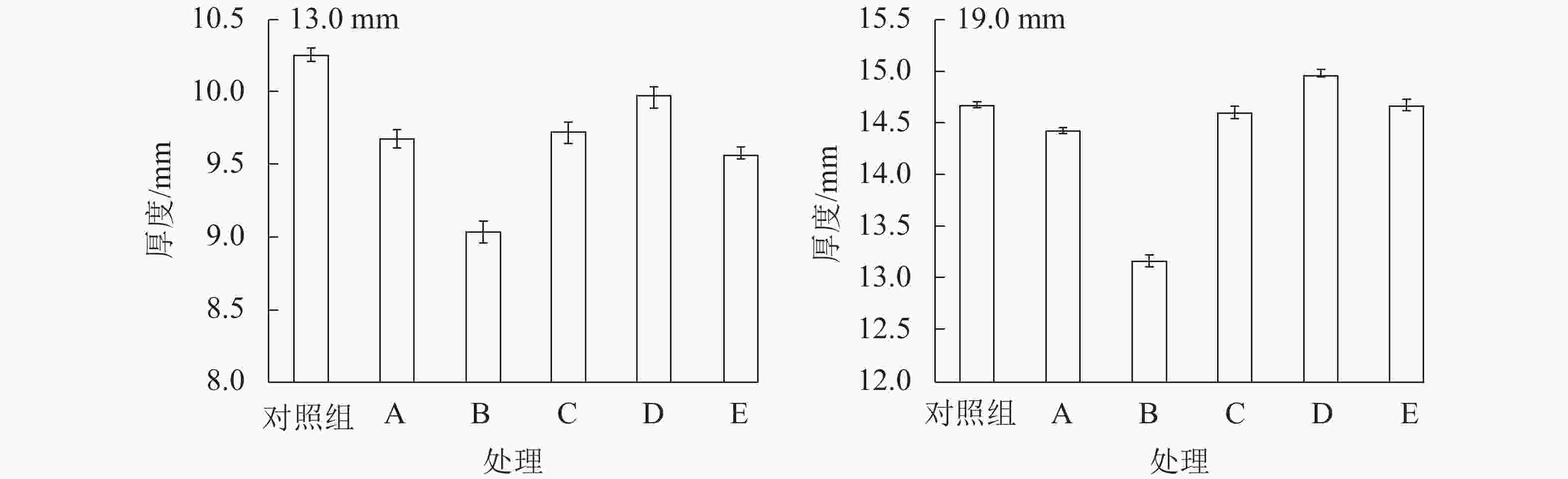
图 2 7 MPa载荷时试件的厚度
Figure 2. Thickness of samples at 7 MPa compressive strength
以厚度为19.0 mm试件为例,相较于对照组,B组试件压缩变形增加约70%,E组试件压缩变形增加约30%。为进一步探索试件压缩强度的发生机制,需要分析试件压缩强度与厚度方向变形(加压头位移)之间的关系(图3)。不同水分行为下,试件发生了内部结构变化和刨花软化,这会影响强度与变形关系曲线。吸放水循环和高含水率均弱化了曲线的拐点,这说明试件的弹性形变区间变窄,更多变形是由塑性形变造成。循环吸放水使得刨花间孔隙放大,刨花的完整形态被破坏,刨花在负载条件下的承载能力随之降低。高含水率条件下[(95±3)%RH],B组试件极易发生变形,变形曲线没有明显拐点,这表明水分使刨花软化,降低了刨花刚度,使刨花更易于压缩变形[19]。
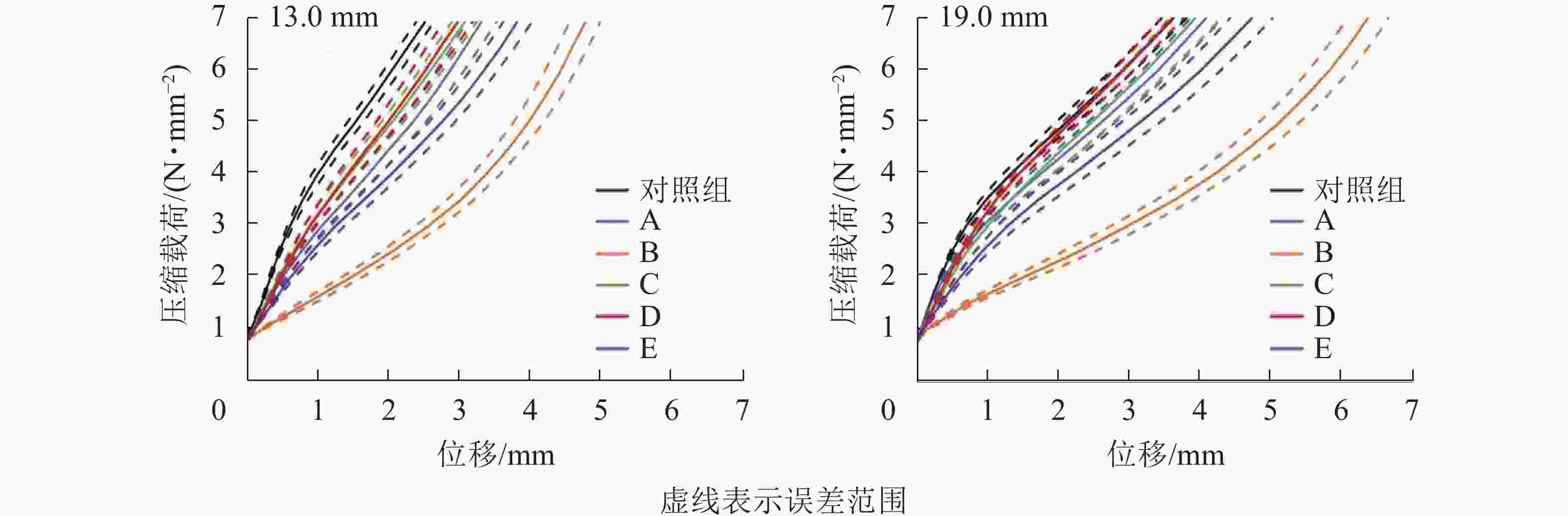
图 3 不同水分条件下试件压缩载荷与变形之间的关系
Figure 3. Relationship between compressive strength and displacement of samples at different water conditions
-
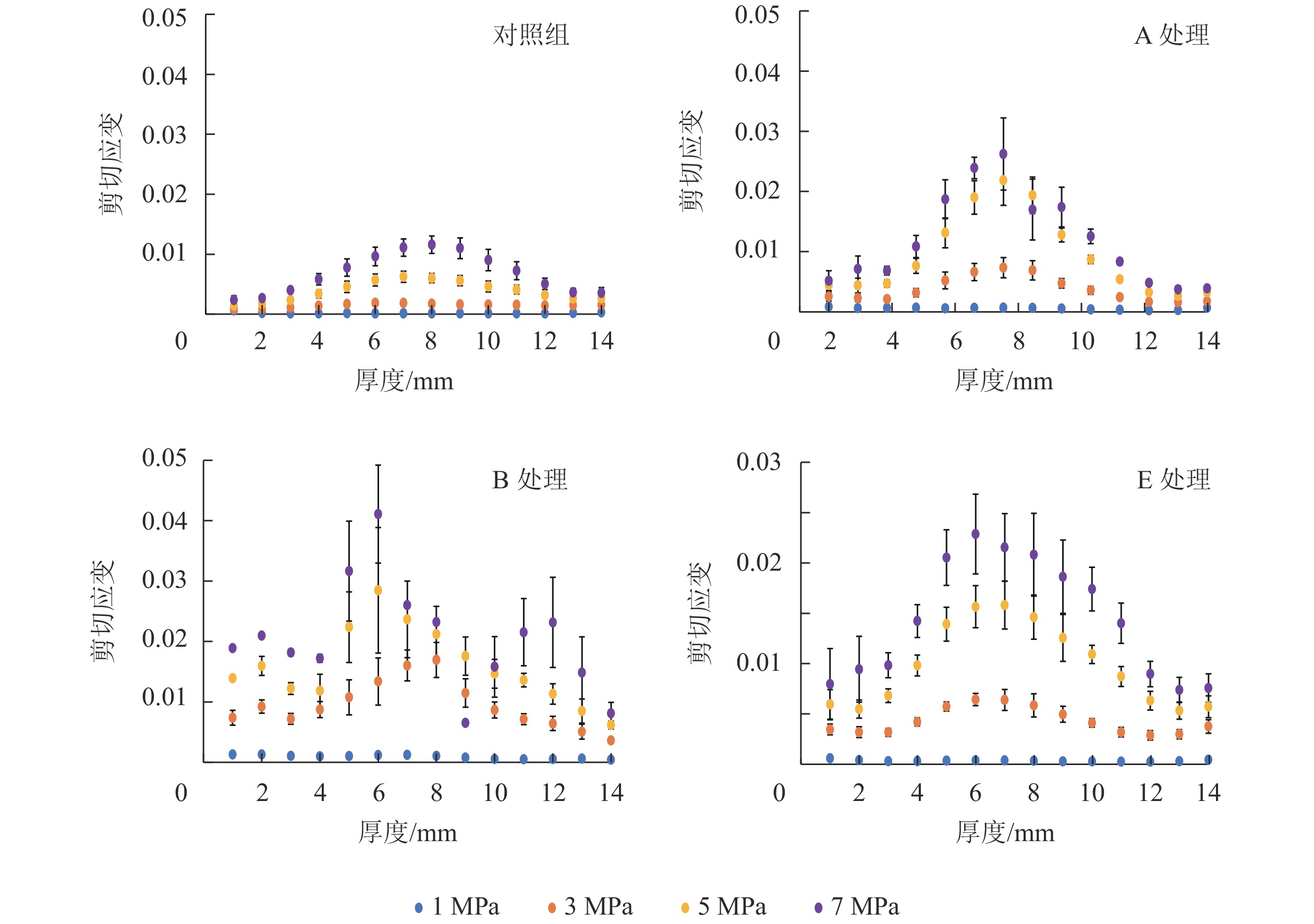
2种厚度试件的应变分布相似,以13.0 mm试件为分析对象。图4和图5结果显示:压缩应变主要集中在沿试件厚度方向的中心区域,这归因于OSB中心区域低密度和表层区域高密度[20]。木质复合材料的密度和刚度往往正相关[21]。因此,应变容易发生在试件沿厚度方向上的中心区域。吸放水循环次数增加,试件内应变增加,但应变分布变化较小。含水率升高会使试件内应变增加,同时改变应变分布。木材刨花会因为含水率升高而软化,这会降低木材刚度。OSB中心区域密度低,单位体积内刨花数量少,因此含水率对该区域应变的影响被弱化。此外,胶黏剂分布对试件内的应变分布也有重要影响,胶黏剂往往可以改善木材的力学性能和尺寸稳定性[22]。OSB表层施胶量通常比中心层多,因此试件表层区域力学性能受吸放水或含水率影响较小,具体体现为B组和E组试件表层区域的应变普遍较小。
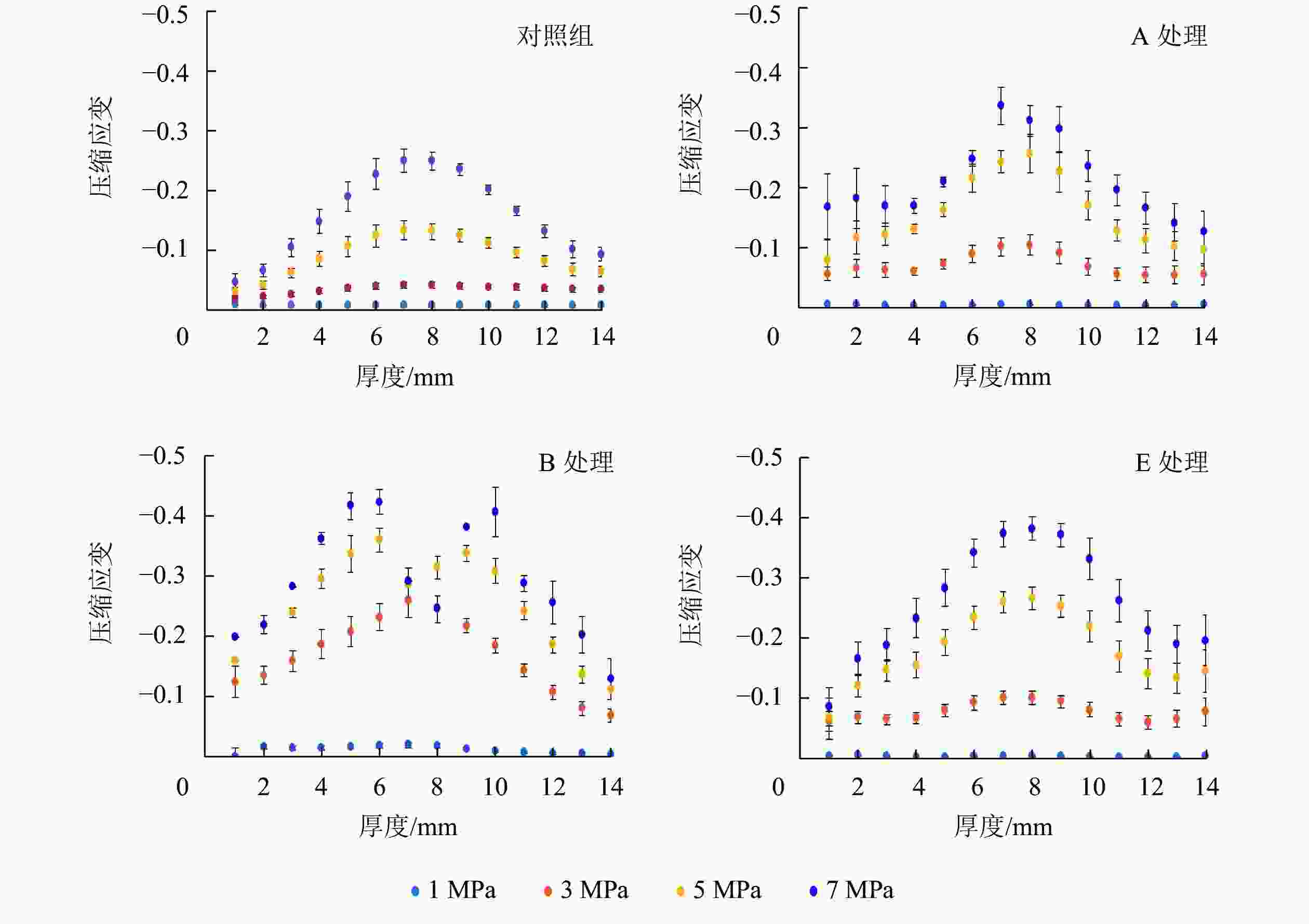
图 4 不同压缩载荷(1、 3、 5、 7 MPa)下13.0 mm试件沿厚度方向的压缩应变分布
Figure 4. Compression strain distribution along the thickness direction of the 13.0 mm samples under four compression strengths(CS=1, 3, 5, 7 MPa)
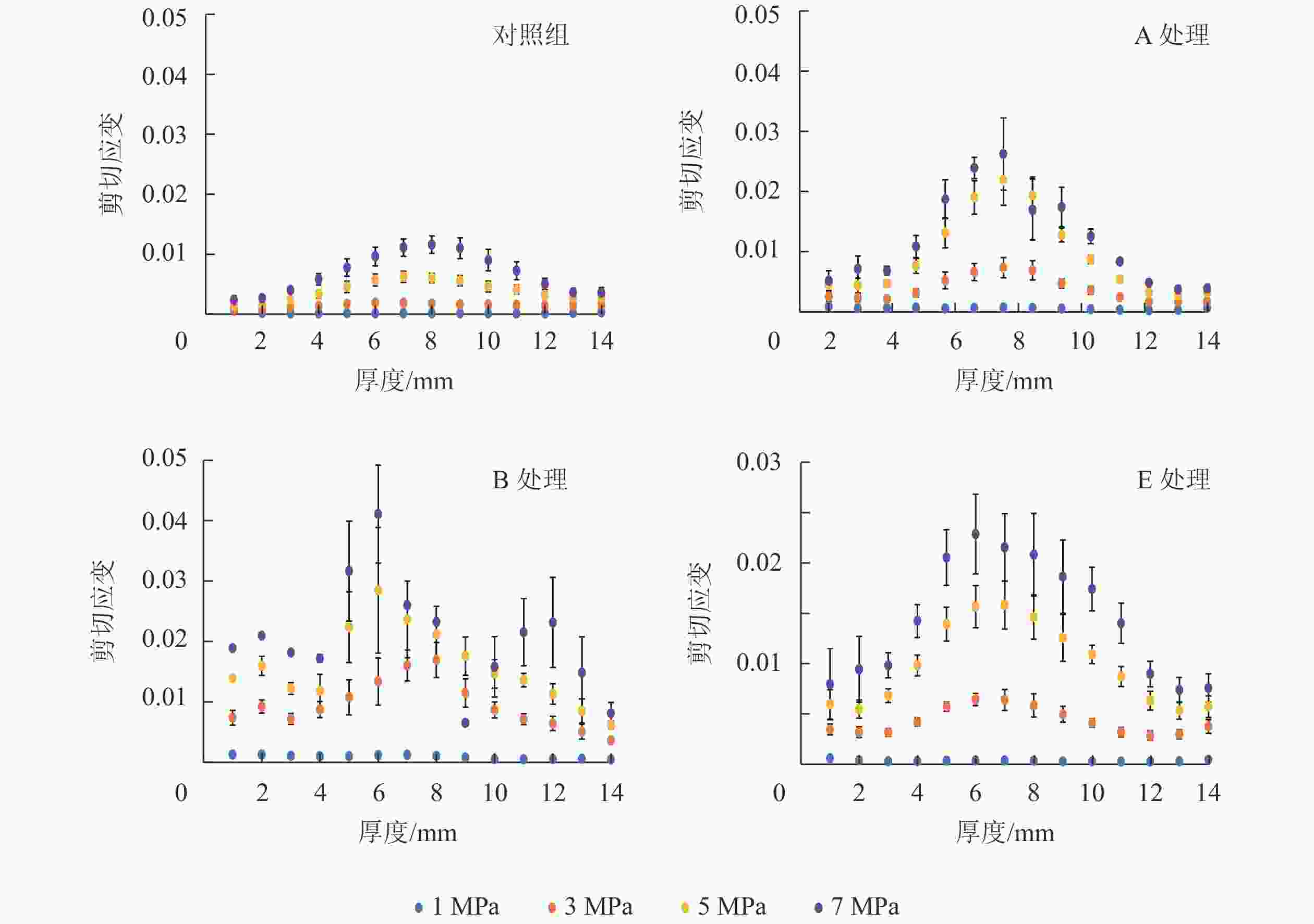
图 5 不同压缩载荷下(1、3、 5、 7 MPa)13.0 mm试件沿厚度方向的剪切应变分布
Figure 5. Shear strain distribution along the thickness direction of the 13.0 mm samples under four compression strengths (CS=1, 3, 5, 7 MPa)
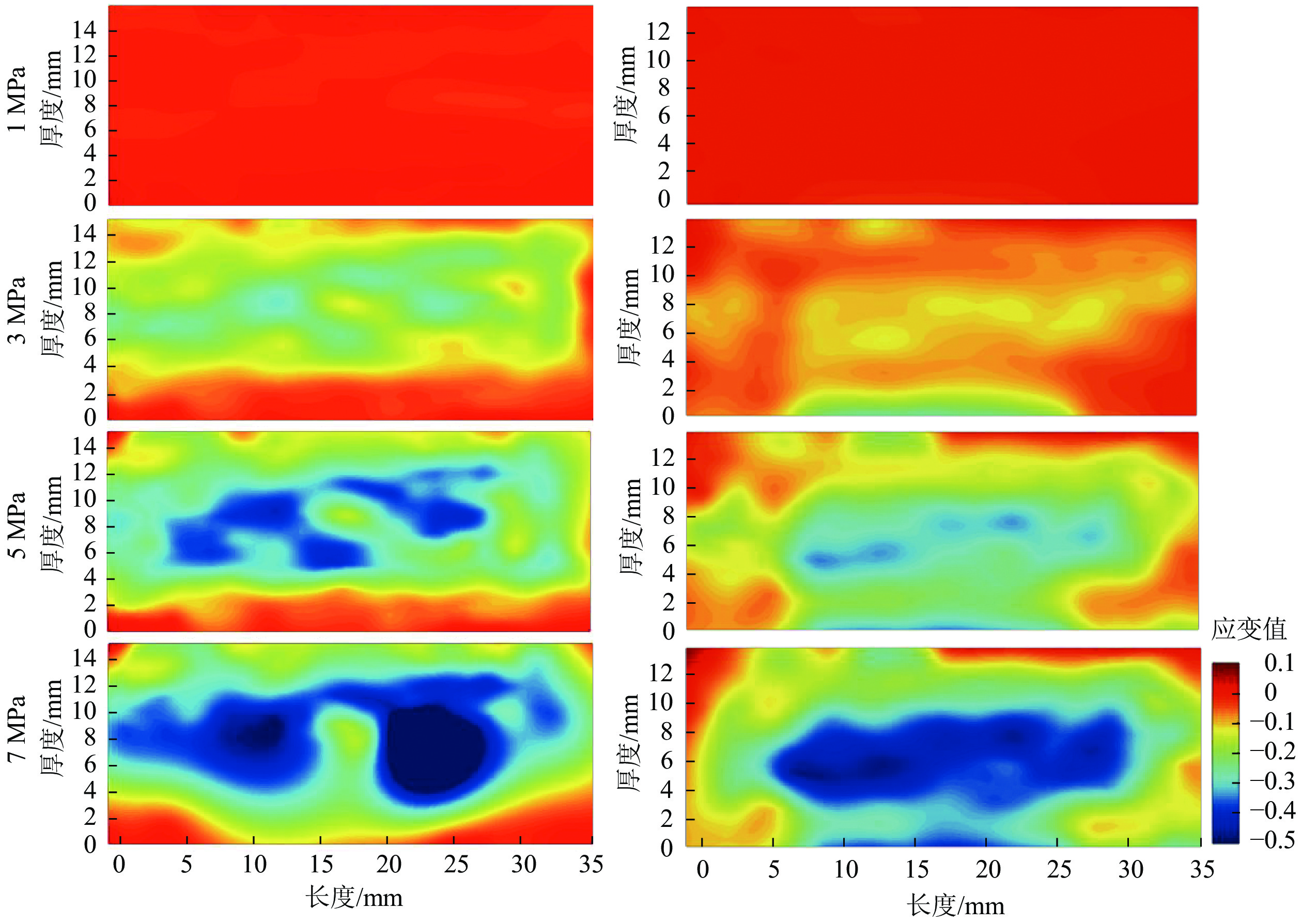
为进一步探索试件内应变的动态迁移过程,区分B组和E组试件的应变分布,由图6和图7结果显示:载荷低于3 MPa时,2组试件内均未出现明显应变,且应变均匀分布。载荷增加时,试件内部结构持续变化,部分区域出现明显的应变集中(图6蓝色区域)。结合图3可知:压缩应变随压缩强度的增大而增加,且主要集中在试件的中心区域。B组试件的应变始于中心区域,但E组试件的应变则从下表面开始。自由水易积聚在近OSB试件侧面和上下表面的孔隙中,或者沿着长刨花向试件内部迁移[4]。多次吸放水循环使得E组试件下表面的结构不再密实,载荷作用下,这一区域容易被再次压缩变形,并产生应变集中。该区域密实化后,应变集中则更多发生在试件中心区域。
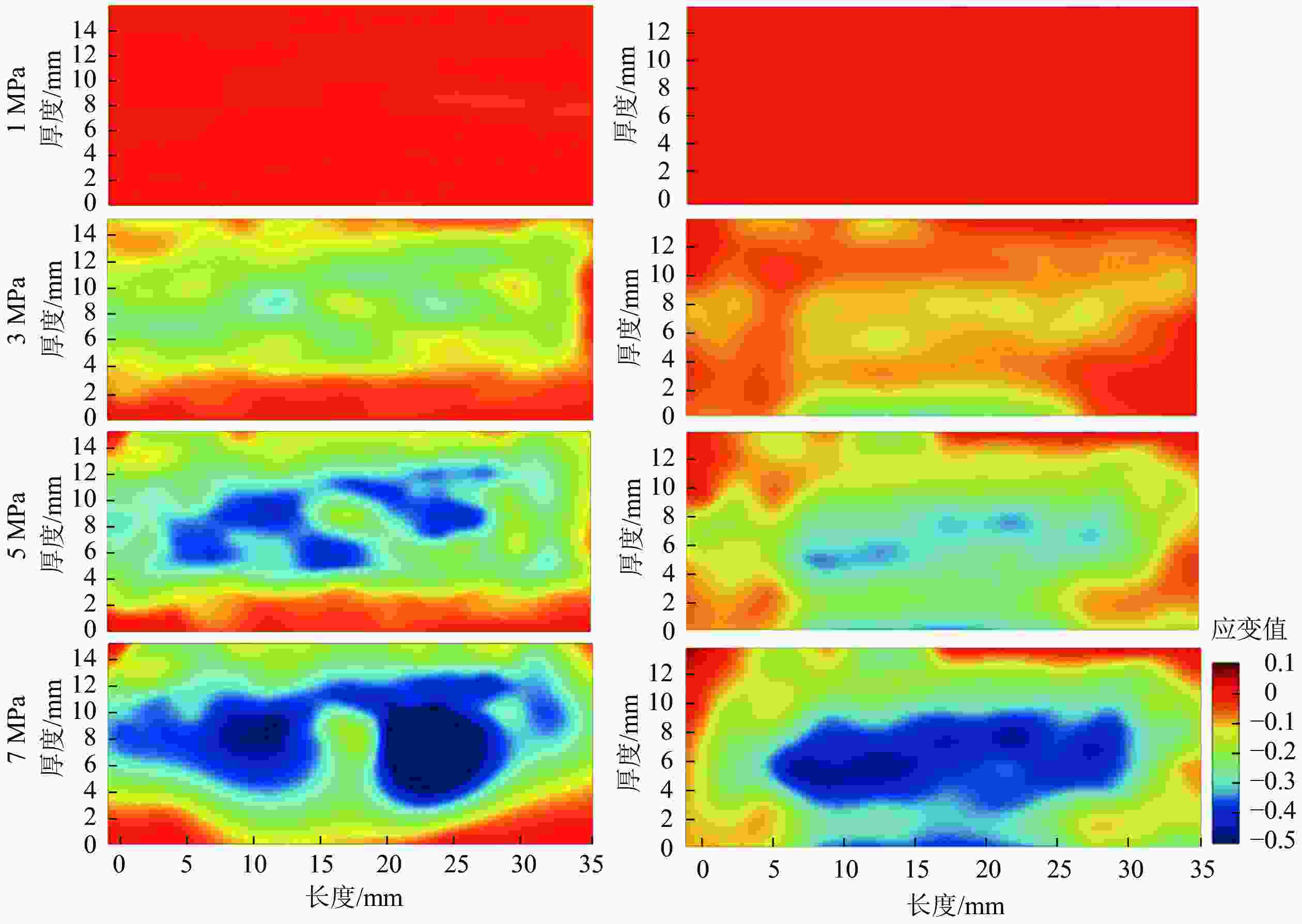
图 6 不同压缩载荷下13.0 mm试件B(左列)和E(右列)的压缩应变分布
Figure 6. Compression strain distribution of 13.0 mm sample B (left) and sample E (right) under four compressive strengths

图 7 不同压缩载荷下13.0 mm试件B(左列)和E(右列)的剪切应变分布
Figure 7. Shear strain distribution of 13.0 mm sample B (left) and sample E (right) under different compression strengths
图7所示:剪切应变分布不连续且主要集中于中心区域。剪切应变往往造成试件内部结构变化,具体表现为木材刨花间相对位置滑移。多孔的内部结构使刨花在载荷状态下易发生开胶和错位,并产生剪切应变。由于MDI胶黏剂的耐水性和稳定性,水分吸着很难将胶黏着的木材刨花分离[23],因此试件上下表面紧凑的内部结构和充分的胶黏剂渗透,可以有效避免水分行为对内部结构的影响,这表现为上下表面区域剪切应变较小。
-
本研究结果表明:①含水率的升高和吸放水循环会使OSB厚度增加和压缩强度降低。薄OSB的厚度和压缩强度更容易受含水率和吸放水循环影响。②含水率增加会使木材刨花软化润胀,含水率20%时,较未吸湿试件,OSB试件压缩变形增加约70%。含水率增加使试件厚度方向上的应变增加且应变分布改变,具体表现为试件厚度中心区域的应变集中现象弱化。③较未吸放水试件,8次吸放水后,OSB压缩变形增加约30%。试件厚度方向上的应变增加,但应变分布变化不明显。④压缩载荷作用下,压缩和剪切应变均多始于OSB试件厚度方向上的中心区域。吸放水循环会释放OSB表层内应力,这引起试件表层区域结构疏松和载荷作用下表层压缩应变集中。
Effect of water on the mechanical performance of OSB in compression tests
-
摘要:
目的 旨在深入研究水分对定向刨花板(oriented strand board, OSB)力学性能的影响规律。 方法 选取13和19 mm共2种厚度的OSB为研究对象,调整试件含水率及对试件进行吸放水循环处理,使用万能力学试验机压缩试件,同步使用数字散斑相关分析技术记录试件的面内应变分布。 结果 含水率和吸放水循环次数增加,均会增加试件厚度和压缩变形。压缩载荷作用下,压缩和剪切应变均多始于OSB试件厚度方向上的中心区域。吸放水循环会释放OSB表层内应力,造成试件表层区域结构疏松和载荷作用下表层压缩应变集中。含水率为20%时,试件压缩变形增加70%,且面内应变分布规律改变,具体表现为试件中心区域应变集中现象弱化。吸放水循环使试件面内应变增加,但对应变分布规律影响较小。8次吸放水循环后,试件压缩变形增加约30%。 结论 含水率和吸放水循环次数的增加均使OSB的抗压强度下降和厚度方向上的应变增加,含水率增加会造成应变分布改变但吸放水循环对应变分布影响不明显。图7表1参23 Abstract:Objective Oriented strand board (OSB) is an environmental- and eco-friendly material produced with renewable, mainly fresh wood from logs with either small or large diameter logs. Composed of cross-oriented layers consisting of thin and rectangular wooden flakes or strands that are compressed and bonded together with synthetic resins, it is an important engineered wood product widely applied in load bearing situations and its mechanical performance is largely dependent on the moisture content (MC) and water sorption/desorption. Therefore, with an investigation of the compressive strength (CS) of samples at different MC and water sorption/desorption cycles, this study is aimed to research the effect of water on mechanical performances of OSB. Method With samples selected from two types of OSB panels with different thicknesses, conditioned to three different MCs and treated with multiple water sorption/desorption cycles, supervision was conducted of the dimensional changes, CS and strain distribution of samples were monitored. Then the CS was measured using an Instron universal testing machine with the strain distribution recorded using digital image correlation (DIC) simultaneously. Result (1) An increase in both MC and water sorption/desorption cycle could lead to an increase in thickness and compressive deformation; (2) The CS of the samples was significantly decreased when they were conditioned with a relative humidity of 95% and a MC of approximately 20%; (3) High MC could reduce the final thickness of OSB after compressing whereas water sorption/desorption cycles could slightly decrease the final thickness of OSB after compressing; (4) High MC and multiple water sorption/desorption cycles were able to change the relationship between CS and displacement from two domains linear to almost linear which attributed to the disappearance of the samples’ plastic deformation domain; (5) High MC can soften wood strands and enlarge internal voids, which could lead to strain increase and the change of strain distribution profile while water sorption/desorption cycles degraded the internal structure of samples and (6) Water sorption/desorption cycles might cause compression strain accumulation in surface layers. Conclusion Water affects the CS of OSB in various aspects including the softening of wood strands, enlargement of voids among wood strands, diminishing of trapped stress from hot-pressing with different MCs. Such findings will help with the generation of effective manufacturing strategies that aim at good mechanical performance of OSB at different water conditions as well as optimizing the application methods and extending the application fields of OSB. [Ch, 7 fig. 1 tab. 23 ref.] -
图 1 不同水分条件下试件的厚度变化和含水率
Figure 1 Thickness change and moisture content of samples at different water situations
图 3 不同水分条件下试件压缩载荷与变形之间的关系
Figure 3 Relationship between compressive strength and displacement of samples at different water conditions
图 4 不同压缩载荷(1、 3、 5、 7 MPa)下13.0 mm试件沿厚度方向的压缩应变分布
Figure 4 Compression strain distribution along the thickness direction of the 13.0 mm samples under four compression strengths(CS=1, 3, 5, 7 MPa)
图 5 不同压缩载荷下(1、3、 5、 7 MPa)13.0 mm试件沿厚度方向的剪切应变分布
Figure 5 Shear strain distribution along the thickness direction of the 13.0 mm samples under four compression strengths (CS=1, 3, 5, 7 MPa)
图 6 不同压缩载荷下13.0 mm试件B(左列)和E(右列)的压缩应变分布
Figure 6 Compression strain distribution of 13.0 mm sample B (left) and sample E (right) under four compressive strengths
图 7 不同压缩载荷下13.0 mm试件B(左列)和E(右列)的剪切应变分布
Figure 7 Shear strain distribution of 13.0 mm sample B (left) and sample E (right) under different compression strengths
表 1 各组试件在 7 MPa载荷条件下变形的t检验结果
Table 1. T-test results of the displacement at 7 MPa
厚度/mm P 对照组/A 对照组/B 对照组/C 对照组/D 对照组/E 13.0 7.87E−03* 6.91E−06* 2.56E−02* 7.32E−02 3.05E−04* 19.0 1.26E−01 1.47E−05* 4.53E−01 9.36E−01 9.39E−03* 说明:*表示差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
-
[1] HSIEH T Y, CHANG F C. Effects of moisture content and temperature on wood creep [J]. Holzforschung, 2018, 72(12): 1071 − 1078. [2] ZHAN Tianyi, JIANG Jiali, LU Jianxiong, et al. Temperature-humidity-time equivalence and relaxation in dynamic viscoelastic response of Chinese fir wood [J/OL]. Constr Build Mater, 2019, 227: 116637[2021-03-15]. doi: 10.1016/j.conbuildmat.2019.08.018. [3] LI Rongrong, CAO Pingxiang, XU Wei, et al. Experimental and numerical study of moisture-induced stress formation in hexagonal glulam using X-ray computed tomography and finite-element analysis [J]. BioResources, 2018, 13(4): 7395 − 7403. [4] LI Wanzhao, van den BULCKE J, DHAENE J, et al. Investigating the interaction between internal structural changes and water sorption of MDF and OSB using X-ray computed tomography [J]. Wood Sci Technol, 2018, 52(3): 701 − 716. [5] 李万兆, 詹先旭, 杨勇, 等. 木质人造板吸水过程中内部单元体尺寸及相对位置的变化[J]. 林业工程学报, 2018, 3(3): 24 − 28. LI Wanzhao, ZHAN Xianxu, YANG Yong, et al. Changes of size and relative position of the internal unit during water uptake of wood-based panels [J]. J For Eng, 2018, 3(3): 24 − 28. [6] PRITCHARD J, ANSELL M P, THOMPSON R J H, et al. Effect of two relative humidity environments on the performance properties of MDF, OSB and chipboard. Part 1. MOR, MOE and fatigue life performance [J]. Wood Sci Technol, 2001, 35(5): 95 − 403. [7] PRITCHARD J, ANSELL M P, THOMPSON R J H, et al. Effect of two relative humidity environments on the performance properties of MDF, OSB and chipboard. Part 2: fatigue and creep performance [J]. Wood Sci Technol, 2001, 35(5): 405 − 423. [8] LI Wanzhao, CHEN Chaoyi, SHI Jiangtao, et al. Understanding the mechanical performance of OSB in compression tests[J/OL]. Constr Build Mater, 2020, 260: 119837[2021-03-25]. doi: 10.1016/j.conbuildmat.2020.119837. [9] HODOUSEK M, BOHM M, SOUCKOVA A, et al. Effect of moisture content on the air permeability of oriented strand boards [J]. BioResources, 2018, 13(3): 4856 − 4869. [10] DUMITRASCU A E, LUNGULEASA A, SALACA E A, et al. Evaluation of selected properties of oriented strand boards made from fast growing wood species [J]. BioResources, 2020, 15(1): 199 − 210. [11] PIPISKA T, PARIL P, CERMAK P, et al. Effect of chemical and thermal modification, and material replacement on strand board properties [J]. Eur J Wood Wood Prod, 2020, 78(3): 565 − 575. [12] ZHAO Sisi, GAO Wei, YI Tengfei, et al. Fabrication of nano-cupric oxide in phenol–formaldehyde resin adhesive: synthesis, curing characteristics, and morphology [J]. Wood Sci Technol, 2019, 53(4): 819 − 835. [13] OSCARSSON J, OLSSON A, ENQUIST B. Strain fields around knots in Norway spruce specimens exposed to tensile forces [J]. Wood Sci Technol, 2012, 46(4): 593 − 610. [14] SUTTON M A, ORTEU J J, SCHREIER H. Image Correlation for Shape, motion and Deformation Measurements: Basic Concepts, Theory and Applications[M]. New York: Springer Science & Business Media, 2009: 81 − 116. [15] CHING D J, KAMKE F A, BAY B K. Methodology for comparing wood adhesive bond load transfer using digital volume correlation [J]. Wood Sci Technol, 2018, 52: 1569 − 1587. [16] MCKINLEY P, SINHA A, KAMKE F A. Understanding the effect of weathering on adhesive bonds for wood composites using digital image correlation (DIC) [J]. Holzforschung, 2019, 73: 155 − 164. [17] 李万兆, 陈玉瑜, 梅长彤. 胶合板层间水分迁移规律的研究[J]. 林业工程学报, 2019, 4(5): 29 – 33. LI Wanzhao, CHEN Yuyu, MEI Changtong. Exploration of water transport behavior among layers in plywood[J]. J For Eng, 2019, 4(5): 29 – 33. [18] LI Wanzhao, MEI Changtong, van den BULCKE J, et al. The effect of water sorption/desorption on fatigue deflection of OSB [J]. Constr Build Mater, 2019, 223: 1196 − 1203. [19] 李任, 黄荣凤. 木材湿热软化径向压缩变形特征研究进展[J]. 世界林业研究, 2019, 32(1): 65 − 69. LI Ren, HUANG Rongfeng. Research progress in the deformation properties of radial compression wood under hydro-thermal condition [J]. World For Res, 2019, 32(1): 65 − 69. [20] GARCIA P J, WANG S Q, MOSCHIER W W. Moisture effects on in-situ vertical density profile monitoring during OSB hot-pressing [J]. For Prod J, 2008, 58(10): 47 − 51. [21] WANG Junfeng, WANG Xuan, ZHAN Tianyi, et al. Preparation of hydro-thermal surface-densified plywood inspired by the stiffness difference in “sandwich structure” of wood [J]. Constr Build Mater, 2018, 177: 83 − 90. [22] WANG Xinzhou, CHEN Xuanzong, XIE Xuqin, et al. Multi-scale evaluation of the effect of phenol formaldehyde resin impregnation on the dimensional stability and mechanical properties of Pinus massoniana Lamb [J/OL]. Forests, 2019, 10(8): 646[2021-04-10]. doi: 10.3390/f10080646. [23] LI Wanzhao, van den BULCKE J, de SCHRUVER T, et al. Investigating water transport in MDF and OSB using a gantry-based X-ray CT scanning system [J]. Wood Sci Technol, 2016, 50(6): 1197 − 1211. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210291
 点击查看大图
点击查看大图

计量
- 文章访问数: 1548
- HTML全文浏览量: 498
- PDF下载量: 27
- 被引次数: 0



