




-
土壤水是自然界水循环中的重要环节,是地表水转化为地下水的纽带,同时也是植物所需水分的主要来源,干旱半干旱地区土壤水是制约植被恢复的关键性因素[1]。黄土高原地处中国中部偏北,受特殊地理条件及气候因素的影响,区域内水土流失严重,是世界上著名的生态脆弱区。为治理黄土高原水土流失,该地区实施了一系列的系统治理,生态环境得到明显改善[2]。
在水资源匮乏的黄土高原,大规模植树造林在遏制水土流失、改善生态环境的同时,一系列新的生态问题凸显,其中人工林土壤水分变化特征是学者们重点关注的科学问题。马婧怡等[3]研究了黄土丘陵区6种不同植被类型0~3 m土层的土壤含水量变化特征,指出乔灌林能消耗更多的深层水分;姬王佳等[4]在陕北地区用同位素分析法探究了不同植被土壤水的运移过程,指出土壤水的补充主要来自降水,不同植被对降水的响应存在差异,小降水事件能引起农地和草地土壤含水量增长,而人工林土壤含水量增长主要依靠夏秋季暴雨补充;BAI等[5]利用Hydrus-1D模型模拟了不同植被0~4 m土层土壤水的变化过程,指出随恢复年限的增加,在耕地转化为林地7~13 a后,人工植被可使土壤含水量出现负增长;杨敏等[6]对半干旱偏旱区、半干旱区、半湿润区的刺槐Robinia pseudoacacia和柠条Caragana korshinskii进行研究指出:植被对水分的利用策略不同,柠条会对更深层次的水分产生影响;张永旺等[7]对陕北雨季和非雨季的乔、灌植被土壤水分亏缺特征进行了定量评价,指出乔木林地土壤水分亏缺较为严重,植被类型是影响土壤水分的重要因素。这一系列研究从不同视角提高了对黄土高原土壤水的分布情况及变化规律的认识,同时也指出土壤含水量受植被类型影响明显,人工植被在恢复过程中会加剧对深层水分的消耗,导致土壤出现低湿现象,严重制约了人工植被的高质量恢复。因此,明确区域典型人工植被土壤含水量的特征是实现区域生态可持续发展的关键。
在晋西黄土区,刺槐、侧柏Platycladus orientalis和油松Pinus tabuliformis是被广泛栽植的树种。为此,本研究以刺槐、侧柏、油松、刺槐-侧柏、刺槐-油松5种典型人工林作为研究对象,对5种典型林分土壤含水量进行测定,探究不同人工林土壤含水量的垂直特征及变化规律,以期为区域植被建设可持续发展提供理论参考。
-
研究区位于山西省吉县蔡家川流域(36°14′27″~36°18′23″N,110°39′45″~10°47′45″E),地处黄土高原东南部,为典型的黄土残塬、梁峁侵蚀地形,海拔为898~1 574 m,流域呈西—东走向,主沟道长为12.15 km,流域面积为39.33 km2。该地区属黄土高原东南部暖温带半湿润区,冬季严寒,夏季炎热,年平均气温为10.0 ℃,年均降水量为533.5 mm,年均潜在蒸散量为1 724.0 mm,降水年际变化较大,且年内分配不均,降水主要发生在4—10月,土壤凋萎湿度为7.96%[8]。流域内人工植被主要为刺槐、油松、侧柏。林下灌木主要有黄刺玫Rosa xanthina、茅莓Rubus parvifolius、六道木Abelia biflora等。林下草本主要有茜草Rubia cordifolia、委陵菜Potentilla chinensis、白莲蒿Artemisia sacrorum等。
-
选择阳坡中坡位置,林龄为30 a,立地条件基本一致的刺槐、侧柏、油松、刺槐-侧柏、刺槐-油松人工林为研究对象,布设20 m×20 m的标准样方,每种林分设置3个重复,进行土壤水分定点观测,不同林分样地间距控制在400 m范围内。
-
于2021年7月布设样地,记录样地基本信息(表1),同时开展土壤物理性质的测定。以10 cm为间隔使用环刀分层测定0~100 cm土层土壤容重、土壤孔隙度。于2022年4—10月每月下旬使用直径为5 cm的土钻测定人工林0~500 cm土层土壤含水量,每个样方沿对角线设置5个取样点,取样深度为500 cm,0~100 cm取样间隔为10 cm,100~500 cm取样间隔为20 cm,共计30个土层。取出的样品一部分用马尔文激光粒度仪测定土壤颗粒组成,另一部分在105°烘干至恒量测定土壤含水量,同一样地不同月份取样距离不超过20 cm,以尽可能减小土壤空间变异带来的影响。降水量数据由布设在样地周围平坦且无遮蔽的自计雨量筒采集。
表 1 样地基本情况
Table 1. Basic information of the sample plot
人工林类型 样地坐标 海拔/
m坡度/
(°)郁闭度/
%林分密度/
(株·hm−2)凋落物蓄积量/
(t·hm−2)土壤容重/
(g·cm−3)有机质/
(g·kg−1)刺槐 36°16′23″N,110°45′39″E 1 090 25 65 1 825 8.30 1.17±0.12 6.50±1.60 侧柏 36°16′20″N,110°45′23″E 1 150 25 32 1 625 3.69 1.26±0.11 9.87±1.26 油松 36°16′21″N,110°45′45″E 1 140 28 53 1 700 14.08 1.25±0.09 6.38±0.52 刺槐-侧柏 36°16′24″N,110°45′49″E 1 120 26 45 1 725 9.75 1.20±0.09 9.83±1.34 刺槐-油松 36°16′24″N,110°45′43″E 1 120 27 55 1 800 11.14 1.17±0.08 11.00±3.35 说明:土壤容重、有机质数据为平均值±标准差。 -
土壤含水量是指土壤中所含水的质量与烘干土质量的比值,计算公式为:
$$ {W}_{i}=({W}_{1}-{W}_{2})/({W}_{2}-{W}_{0})\times 100\% 。 $$ (1) 式(1)中:$ {W}_{i} $为第$ i $层土壤含水量(g);$ {W}_{1} $为湿土+铝盒的质量(g);$ {W}_{2} $为烘干土+铝盒的质量(g);$ {W}_{0} $为铝盒的质量(g)。
变异系数和标准差计算公式为:
$$ {C}_{\mathrm{V}}=\frac{\sigma }{\mu }\times 100\%\text{;} $$ (2) $$ \mathrm{\sigma }=\sqrt{\sum _{{{i=1}}}^{{n}}\frac{{({W}_{i}-\mu )}^{2}}{n}\mathrm{。}} $$ (3) 式(2)~(3)中:$ {C}_{\mathrm{V}} $为变异系数;$ \sigma $为标准差,$ \mu $为平均值,i为土壤层数,n为土壤总层数。
-
利用Excel 2010处理数据,SPSS 20.0分析数据,Origin Pro 2022制图;运用有序聚类法[9]对土壤含水量垂直层次进行划分,通过DPS 9.01数据处理系统对土壤含水量、标准差及变异系数进行有序聚类,确定最优分层结果。
-
由图1可知:刺槐、侧柏、油松、刺槐-侧柏、刺槐-油松人工林0~100 cm土层土壤容重均值分别为1.20、1.20、1.25、1.20、1.14 g·cm−3,土壤毛管孔隙度为42.08%~51.83%、41.70%~47.14%、41.80%~51.63%、34.65%~48.17%、43.00%~50.91%。刺槐、侧柏、油松、刺槐-侧柏、刺槐-油松人工林0~500 cm土层土壤黏粒质量百分比为2.09%~3.31%、4.37%~10.21%、2.68%~4.87%、5.13%~7.52%、2.51%~4.98%,粉粒质量百分比为33.18%~51.52%、26.68%~37.21%、33.05%~49.33%、36.38%~43.72%、42.96%~53.50%,砂粒质量百分比为45.38%~65.10%、53.23%~68.52%、46.48%~63.76%、50.14%~57.99%%、43.16~53.75%。将人工林土壤颗粒组成按国际制土壤质地分类标准进行划分,5种人工林土壤为砂质壤土。
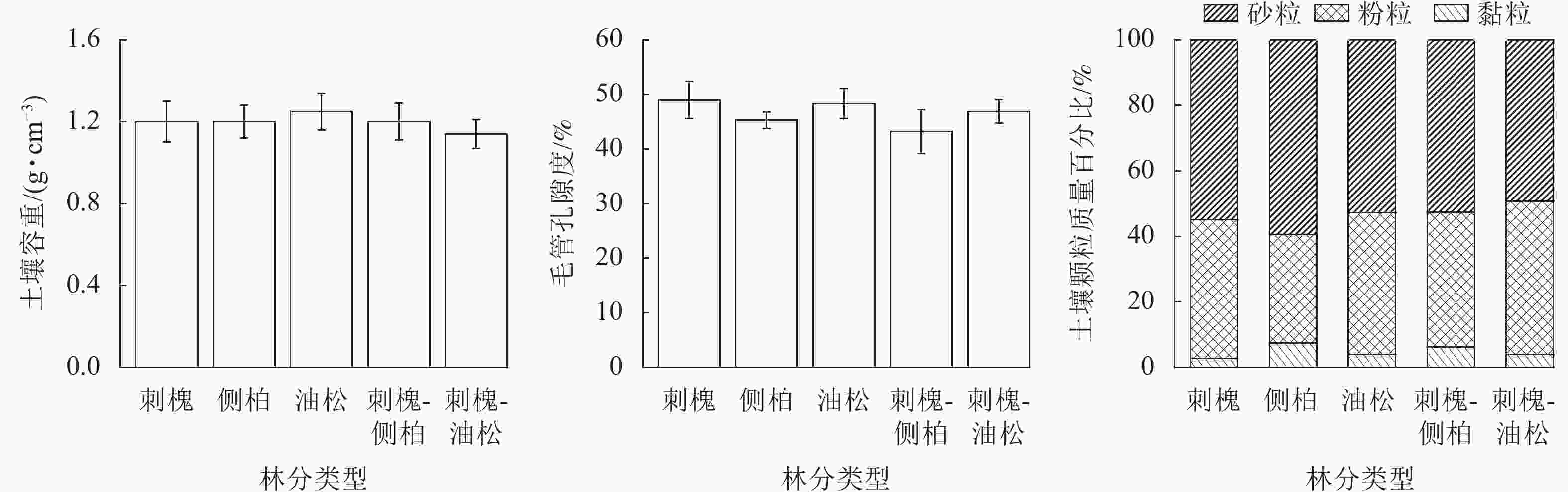
图 1 人工林土壤容重、毛管孔隙度及颗粒组成特征
Figure 1. Soil bulk density, porosity and texture characteristics of plantation
-
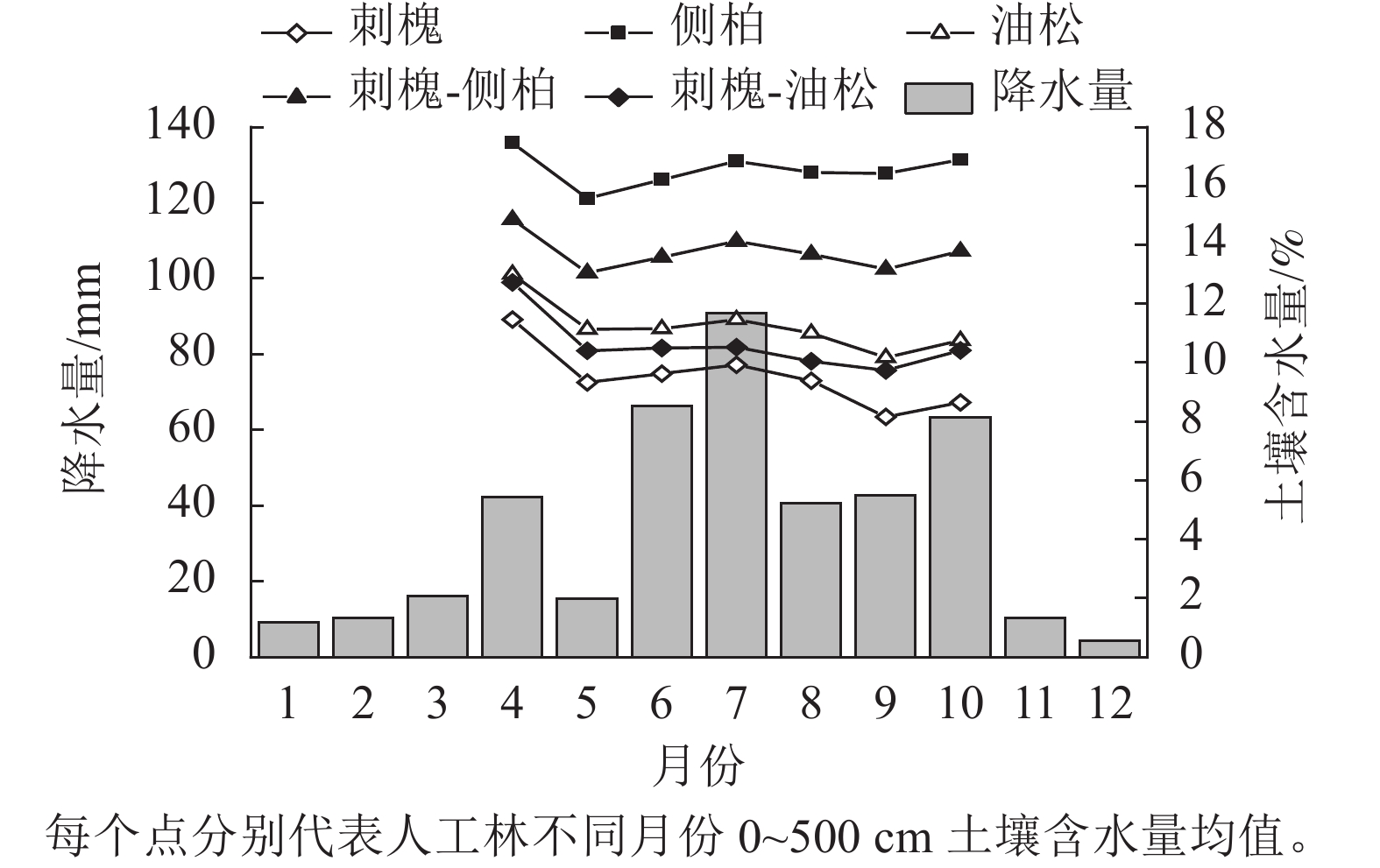
研究区2022年降水量为411.2 mm,低于区域多年平均降水量(533.5 mm)。2022年生长季(4—10月)降水量为361.2 mm,占全年降水的87.84%。
监测期内,人工林土壤含水量的增减变化规律和降水的年内分配保持一致(图2)。人工林土壤含水量在年内有2次增减变化,4—5月和8—9月土壤含水量表现为下降趋势。2个时期中,刺槐林土壤含水量分别减少了2.14%、1.77%,为下降最多的林分,侧柏林土壤含水量分别减少了1.86%、0.42%,为下降最少的林分,表明3种纯林中刺槐耗水最多,其次为油松,侧柏耗水最小。

图 2 典型人工林土壤含水量随时间的变化
Figure 2. Variation of soil moisture content over time in a typical plantation
对比监测期内林地土壤含水量情况,生长季末平均土壤含水量小于生长季初,人工林土壤含水量均处于减少状态,同期雨季结束,年内土壤水分无法得到有效恢复,不良的水分状况将持续到次年生长季。
-
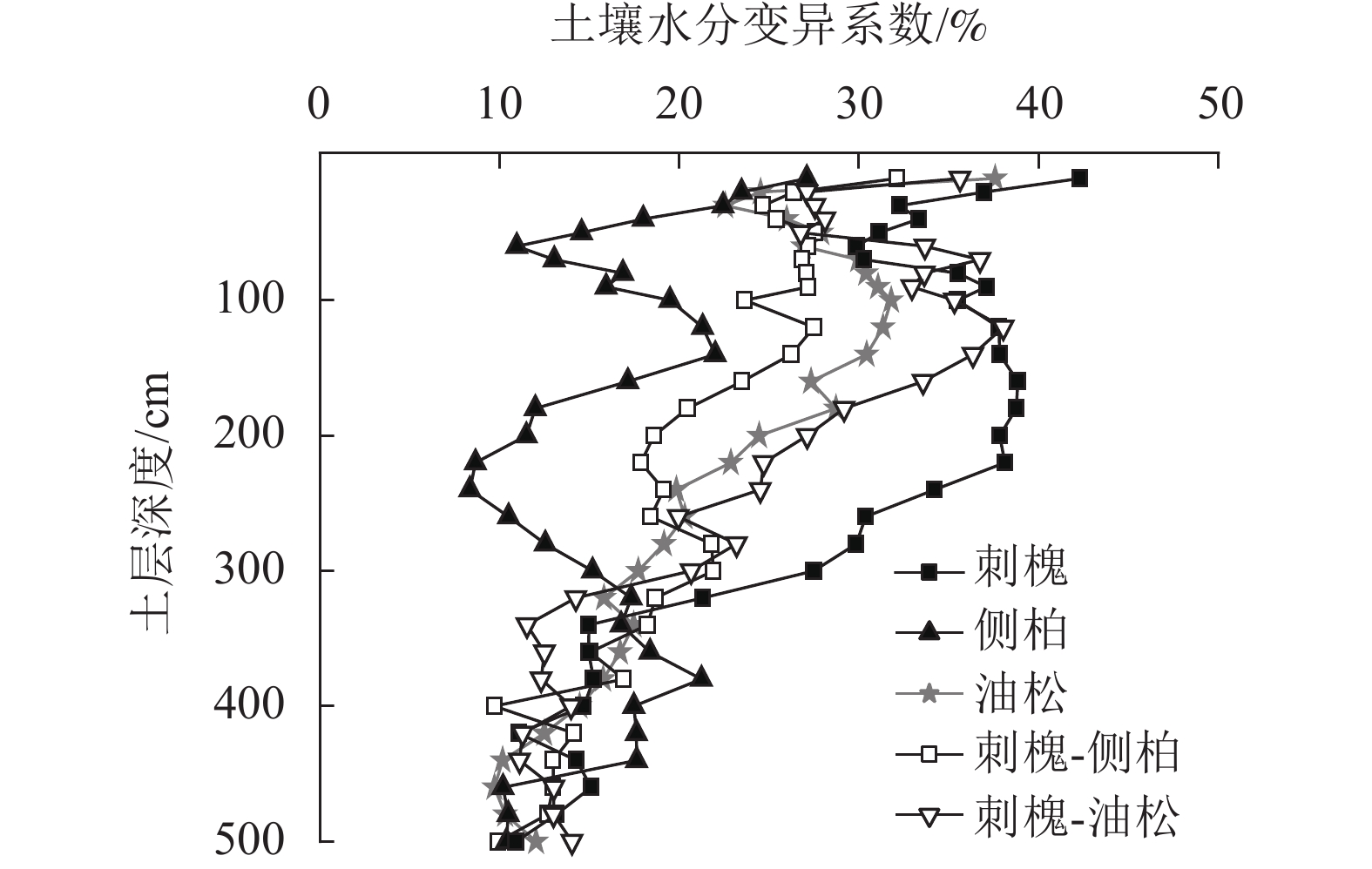
由图3可知:人工林土壤含水量具有相似的垂直变化特征,上层土壤含水量变化剧烈,随土层深度的加深,水分变化趋势逐渐减弱。经计算5种人工林土壤含水量变异系数为15.93%~28.02%,属中等变异,0~500 cm土壤含水量变异系数从大到小依次为刺槐(28.02%)、刺槐-油松(24.05%)、油松(22.18%)、刺槐-侧柏(20.81%)、侧柏(15.93%)。
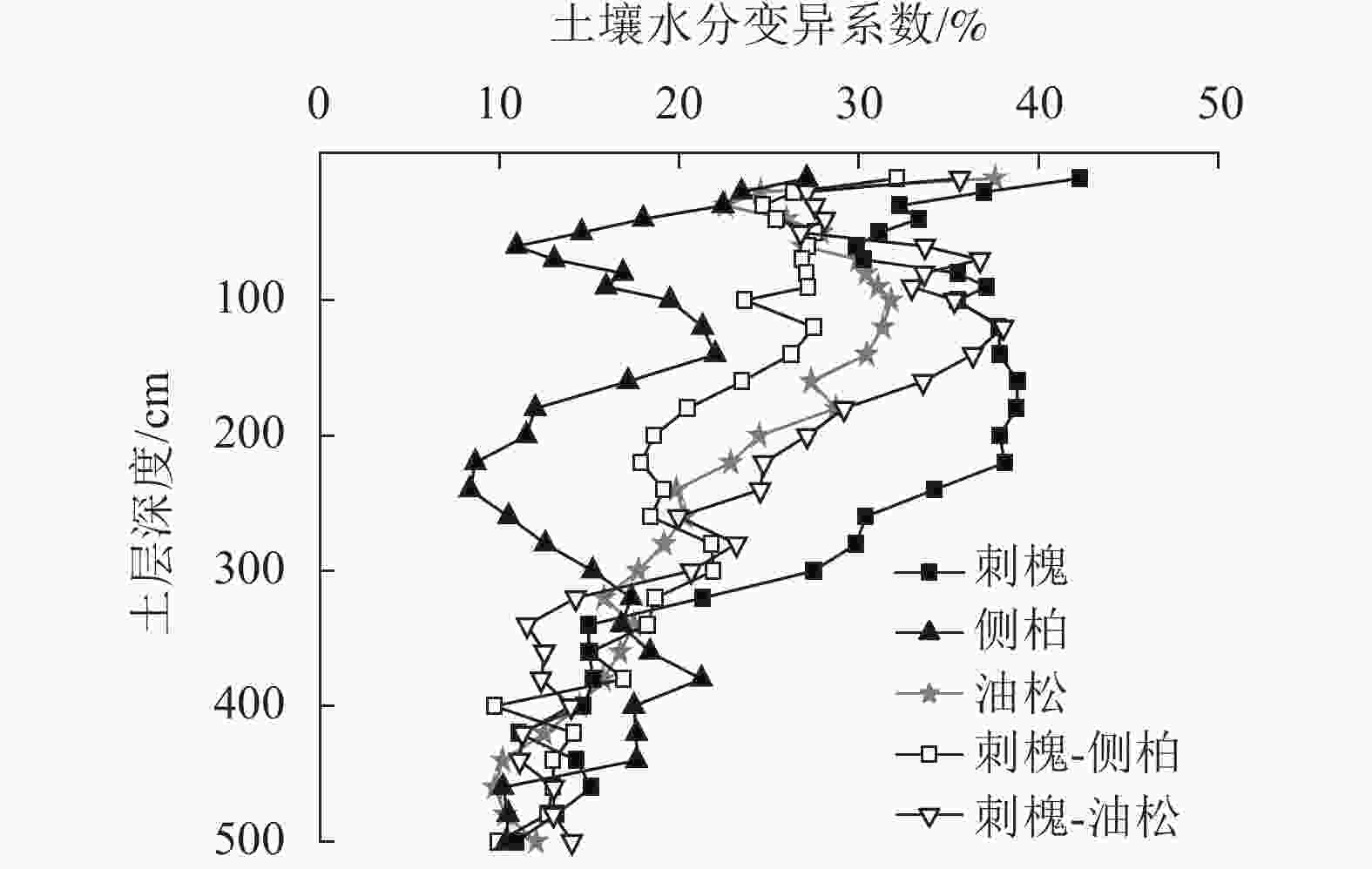
图 3 人工林土壤含水量垂直变异
Figure 3. Vertical variation of soil water content in plantation
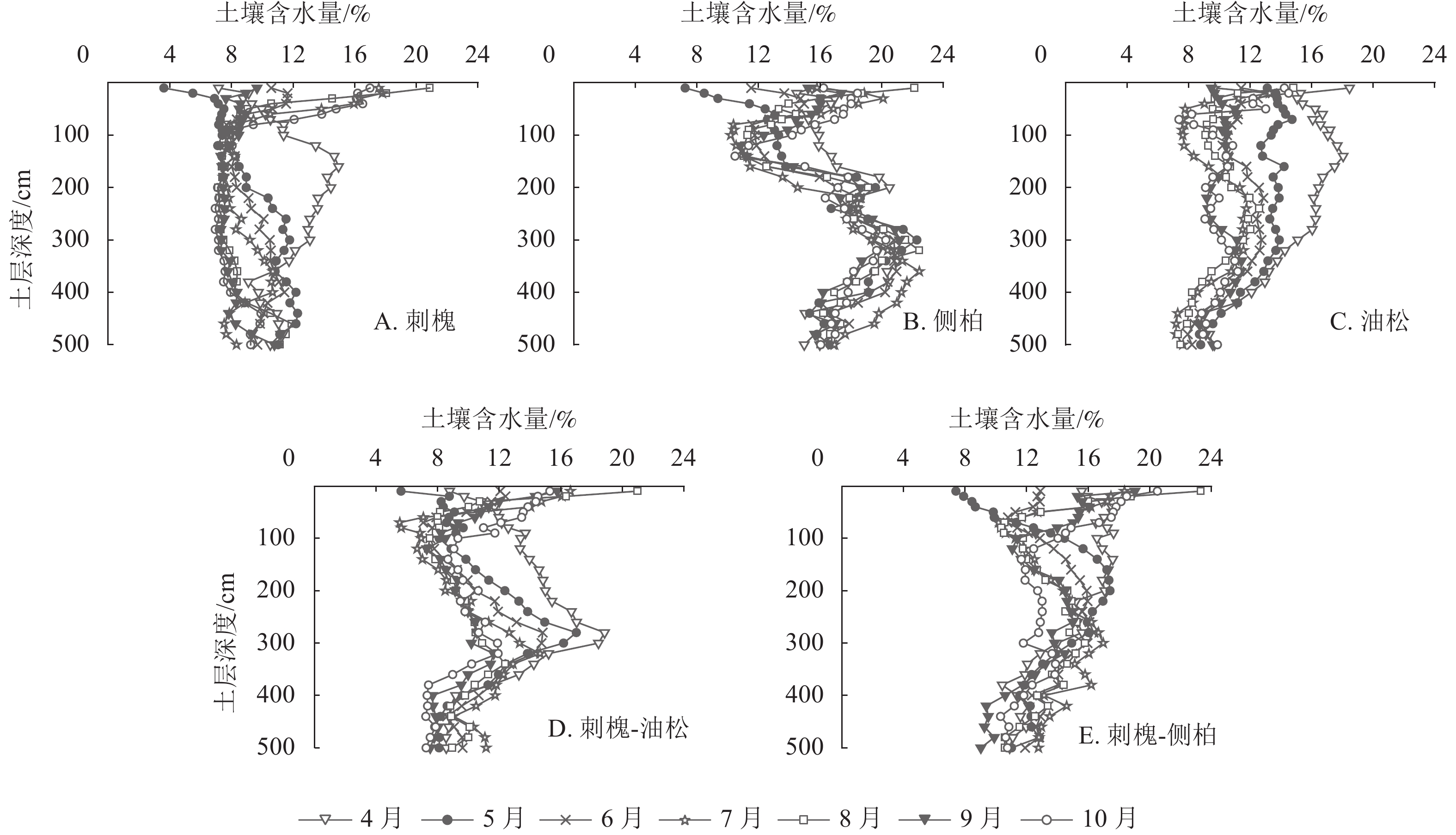
由图4可知:5种人工林土壤含水量垂直分布存在明显差异。刺槐林0~100 cm土层土壤含水量较高,水分变化较为活跃,随土壤深度的加深,变化趋势减弱,约在340 cm土层趋于稳定。侧柏和刺槐-油松人工林土壤含水量在垂直方向上呈S形变化,土壤含水量变化拐点分别出现在100和300 cm土层。以300 cm为界,侧柏林深层土壤含水量明显高于浅层,刺槐-油松林深层土壤含水量低于浅层。油松林在0~300 cm土层土壤含水量有波动,在300~500 cm内土壤含水量出现明显减少。刺槐-侧柏林土壤含水量在0~100 cm土层土壤含水量变化较为活跃,在100~500 cm土层土壤含水量随深度增加呈递减趋势。
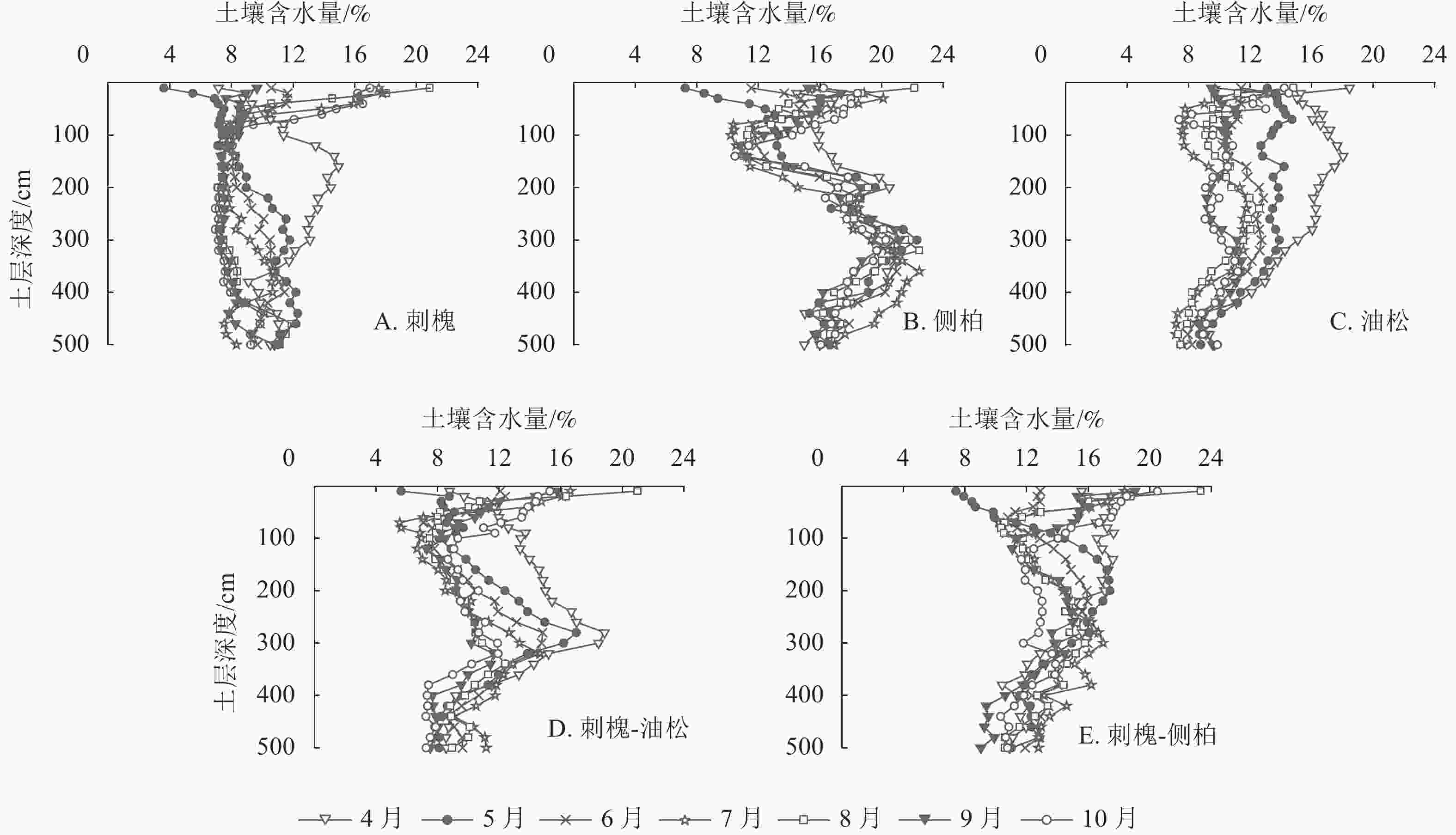
图 4 人工林土壤含水量垂直分布
Figure 4. Vertical distribution of soil water content in plantation
以4—5月水分变化为例,此时气温回升,植被处于萌芽展叶阶段,需消耗大量水分,5月降水量仅为15.4 mm,无法满足植被生长所需。干旱发生时,刺槐0~320 cm、侧柏0~200 cm、油松0~300 cm、刺槐-侧柏0~120 cm、刺槐-油松0~280 cm不同层次土壤含水量下降明显(图4),表明在干旱条件下刺槐、油松、刺槐-油松会加剧对深层水分的消耗,混交造林能有效缓解植被对深层水分的消耗。
监测期内5种人工林0~500 cm土层土壤平均含水量由大到小依次为侧柏(16.56%)、刺槐-侧柏(13.76%)、油松(11.25%)、刺槐-油松(10.63%)、刺槐(9.51%)。单因素方差分析表明:5种人工林0~500 cm土层土壤含水量差异显著(P<0.05)。
-
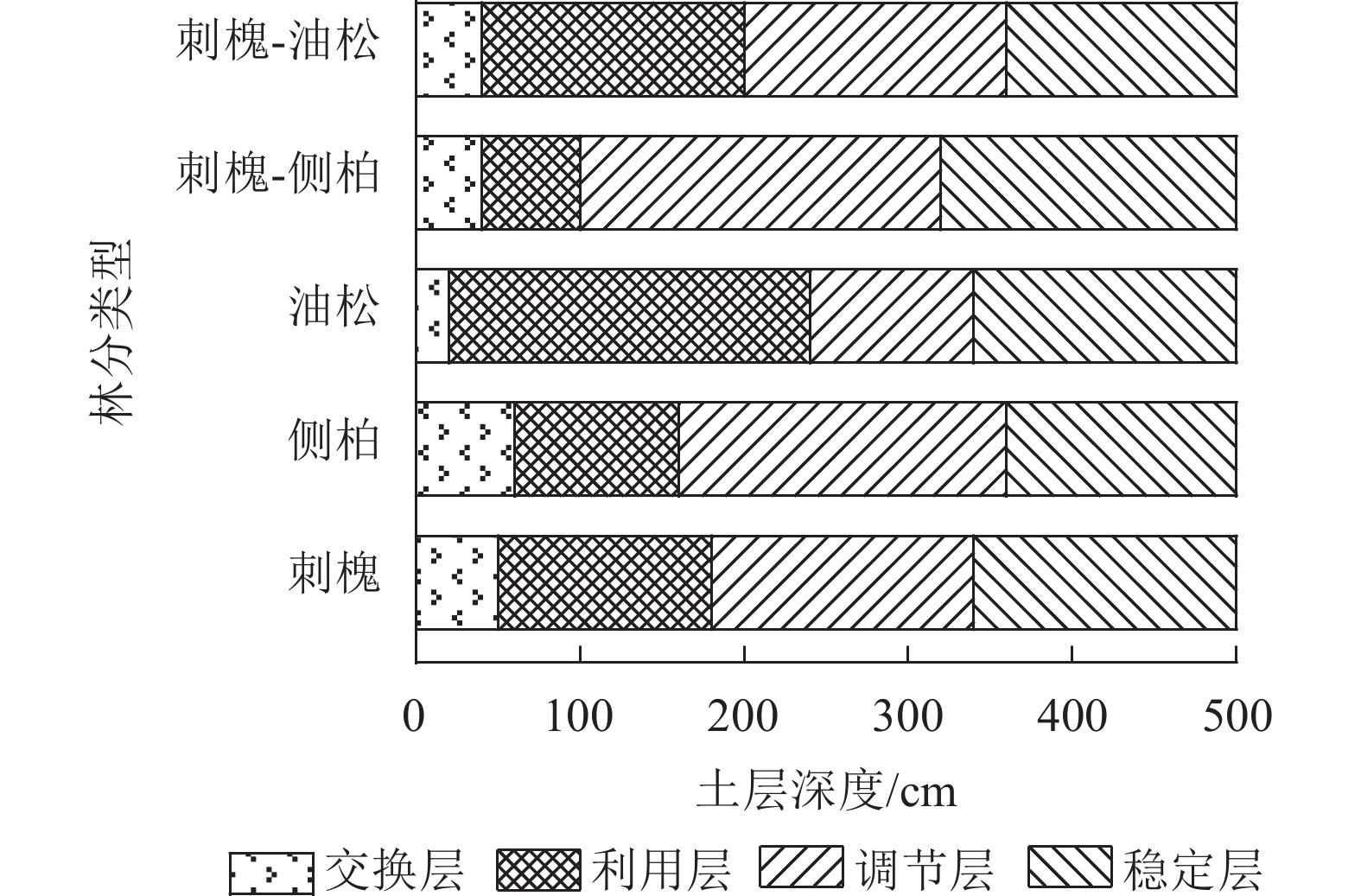
有研究表明:人工林土壤含水量垂直变化表现出层次性[10]。为直观体现土壤水分的垂直运动规律,采用有序聚类法对目标人工林0~500 cm土层土壤水分层次进行划分。依据最优分割结果,将水分垂直层次分为4层,即交换层、利用层、调节层和稳定层[11−12]。受入渗速率、导水率、植物根系等因素影响,不同林分划分结果不同(图5)。
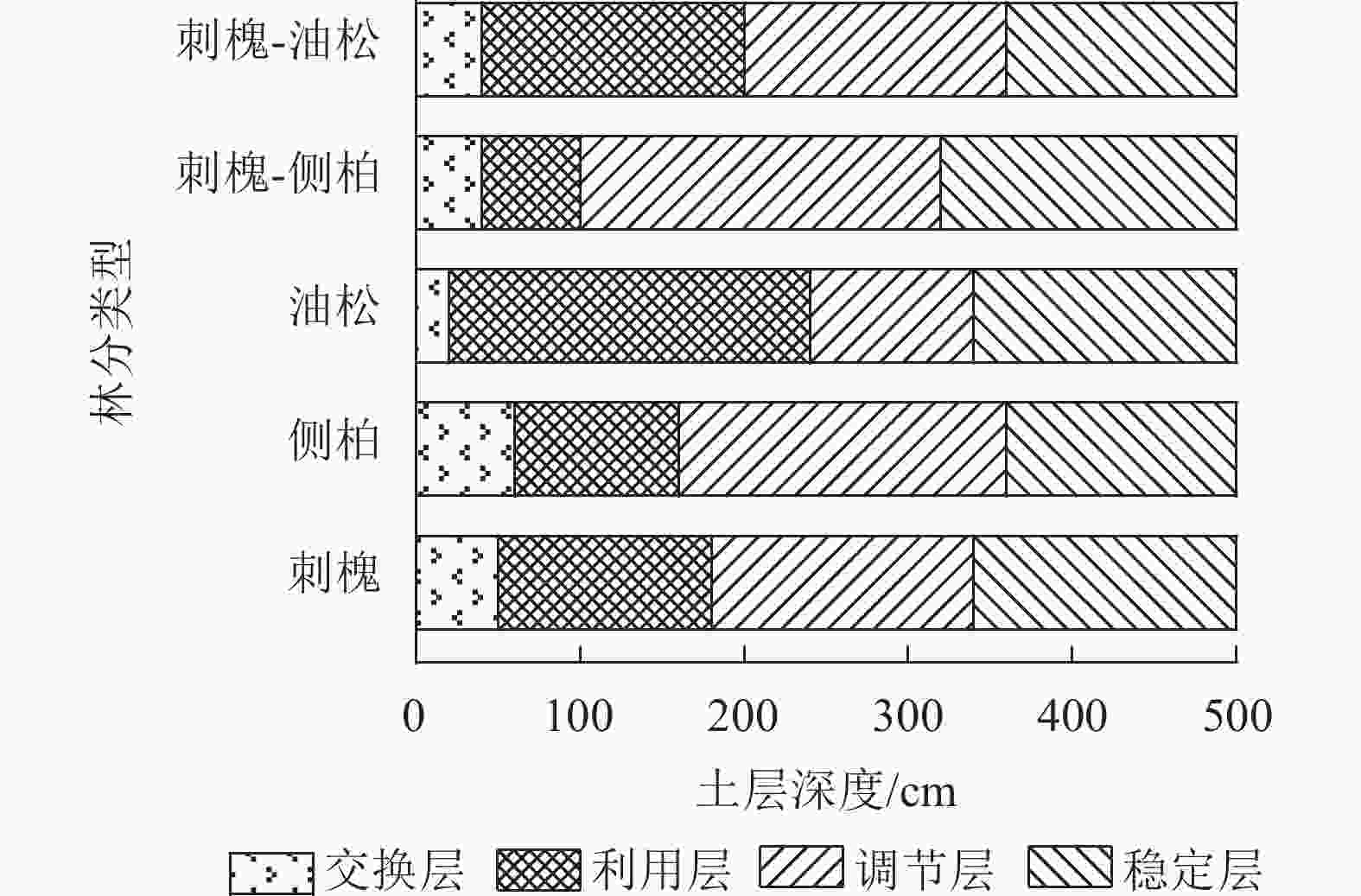
图 5 人工林土壤含水量垂直有序聚类分层结果
Figure 5. Results of vertical ordered cluster stratification of soil water content in plantation
刺槐林交换层为0~50 cm土层,利用层为50~180 cm土层,调节层为180~340 cm土层,稳定层为340~500 cm土层;侧柏林交换层为0~60 cm土层,利用层为60~160 cm土层,调节层为160~360 cm土层,稳定层为360~500 cm土层;油松林交换层为0~20 cm土层,利用层为20~240 cm土层,调节层为240~340 cm土层,稳定层为340~500 cm土层;刺槐-侧柏林交换层为0~40 cm土层,利用层为40~100 cm土层,调节层为100~320 cm土层,稳定层为320~500 cm土层;刺槐-油松林交换层为0~40 cm土层,利用层为40~200 cm土层,调节层为200~360 cm土层,稳定层为360~500 cm土层。总的来看,交换层占比较小,仅占剖面的4%~12%;油松林水分利用空间范围最大,为220 cm,刺槐-侧柏林最小,为60 cm;侧柏和刺槐-侧柏林水分调节空间最大,均为220 cm;稳定层集中在320~360 cm土层内。
-
由表2可知:5种人工林土壤含水量与土壤颗粒组成表现出明显的相关关系,其中土壤含水量与土壤黏粒、粉粒呈显著正相关(P<0.05),与土壤砂粒呈显著负相关(P<0.05),刺槐人工林土壤含水量与土壤容重呈显著负相关(P<0.05),5种人工林土壤含水量与毛管孔隙度无相关。
表 2 人工林土壤含水量及其影响因子皮尔逊相关系数
Table 2. Pearson correlation coefficient of soil moisture content in plantation and its influencing factors
影响因素 刺槐 侧柏 油松 刺槐-侧柏 刺槐-油松 黏粒 0.537** 0.791** 0.512** 0.432* 0.775** 粉粒 0.452* 0.687** 0.425* −0.489* 0.431* 砂粒 −0.467** −0.749** −0.436* −0.545** −0.442* 土壤容重 −0.604** 0.177 0.079 0.159 −0.099 毛管孔隙度 0.310 0.184 0.208 0.405 0.093 说明:**代表极显著相关(P<0.01);*代表显著相关(P<0.05)。 -
大气降水作为黄土高原地区最主要的水资源补充来源,对区域水环境产生明显影响。本研究发现:晋西黄土区人工林土壤含水量时间动态受降水年内分配影响明显,人工林土壤含水量的变化规律和降水量的变化趋势保持一致,这与张晨成[13]的研究结果一致。本研究的5种人工林土壤含水量差异显著(P<0.05),监测期内人工林土壤含水量从大到小依次为侧柏、刺槐-侧柏、油松、刺槐-油松、刺槐。土壤颗粒组成能显著影响土壤含水量(P<0.05),这与前人研究结果一致[14]。土壤颗粒组成通过影响土壤结构,提高或降低土壤持水性和渗透性,从而影响土壤含水量[15]。植被恢复前,地表多为裸露状态,可认为土壤含水量初始值近似,垂直方向上形成的水分差异与植被类型密切相关[16]。植被自身的耗水特性与根系分布均能对土壤含水量产生较大影响。本研究发现:在干旱半干旱的背景下,侧柏林深层土壤含水量较高,其原因在于侧柏作为低耗水树种所消耗的水分较少[17],同时,侧柏细根在水平方向上较为发达,具有明显的表聚性[18],能充分利用浅层土壤水,而深层水分未出现明显消耗,使得林地土壤含水量处于高值。植被根系可借助水势差将深层水分抽取至上层土壤[19],保持根区相对湿润以应对干旱。刺槐、油松作为高耗水树种,生长需要消耗大量水分,其树冠面积较大,林冠截留导致可供补给的水分减少,植被加剧对深层水分的消耗,由此导致刺槐、油松、刺槐-油松3种人工林土壤含水量在剖面上明显减少。任婧宇等[20]研究指出:混交林中不同植被根系穿插改善了土壤环境,使水分入渗速率加快,有效缓解单一树种对深层水分的过度利用,从而改善林地水分状况,这与本研究结果相同。综合5种人工林土壤含水量的情况,刺槐林深层土壤低湿现象最为明显,80 cm以下土层土壤含水量接近研究区凋萎湿度(7.96%)。马建业等[21]研究认为:刺槐所消耗的水分中,约60%来自深层水分上升补给,导致林下深层土壤含水量长期处于低值。王宁等[22]通过土壤水分植被承载力模型计算得出:晋西黄土区刺槐人工林土壤水分植被承载力为1 224株·hm−2,若林分密度超过此阈值,会加剧对深层水分的消耗。本研究中刺槐人工林密度(1 825株·hm−2)已超出区域土壤水分植被承载力阈值。刺槐被广泛栽植于晋西黄土区,在气候暖干化背景下,刺槐林下深层水分长期保持低值,使其难以持续发挥生态服务功能。从土壤含水量的角度出发,在今后造林经营中应减少营造刺槐林,对于现有水分状况较差的刺槐林,可通过间伐减小林分密度,或考虑混交造林,以改善林下土壤低湿的现状。
黄土区土壤水在垂直方向上的动态变化表现出明显层次性,这是降水、植被、土壤等因素综合作用的结果。研究表明:黄土区土壤水分垂直层次划分为速变层、活跃层、次活跃层、相对稳定层[23−24]。本研究采用有序聚类法将研究区人工林0~500 cm土层土壤水分层次由上到下划分为交换层、利用层、调节层和稳定层,划分结果与刘新春等[25]的类似。交换层内水分受光照、降水等外界环境影响较大,层内土壤含水量变化剧烈。油松林交换层分布在0~20 cm土层,为5种人工林中最薄,其原因在于油松林下常年覆盖有致密的凋落物层,这可增强对降水的拦蓄,对水分向下再分配产生影响。人工植被根系在利用层内广泛分布[26],该层土壤易得到降水补给[27],有效地为植被生长发育提供水分,受降水补给和植被消耗的影响,层内土壤具有明显的干湿变化过程。调节层与利用层联系紧密,降水充沛时蓄积水分,干旱时释水供给植物利用,起到良好的调节作用[28]。稳定层受重力影响,层内土质紧实,水分对降水响应滞缓,且地下水难以穿过黄土层向上补给,层内土壤水分较为稳定。综合来看,人工林360 cm以下土层土壤含水量较为稳定,5种人工林在0~360 cm土层内土壤含水量存在较大差异,表明该范围内的土壤含水量受林分类型影响明显,这与黄土高原地区相关研究结果相似[20, 25]。
-
晋西黄土区植被类型差异导致0~500 cm土层土壤含水量存在显著差异。侧柏林土壤含水量显著高于其他4种人工林,刺槐-侧柏林其次,油松、刺槐-油松林较低,刺槐林最低。根据土壤水分监测结果,人工林土壤水分垂直变化可划分为交换层、利用层、调节层和稳定层4个层次,上层水分变化剧烈,随土壤深度的加深,水分变化趋势逐渐减弱,受植被自身利用的影响,刺槐、油松、刺槐-油松林会加剧对深层土壤水分的消耗。因此,在今后造林经营中,应充分考虑植被类型对土壤含水量的影响,选择合理的植被造林,防止深层土壤水分出现过耗的现象。
Vertical distribution of soil moisture in typical plantation in the loess region of western Shanxi Province
-
摘要:
目的 探究晋西黄土区人工林恢复过程中0~500 cm土层土壤含水量垂直特征,为改善人工林土壤水环境,实现林水平衡提供理论依据。 方法 以晋西黄土区蔡家川流域5种典型人工林(刺槐Robinia pseudoacacia、侧柏Platycladus orientalis、油松Pinus tabuliformis、刺槐-侧柏、刺槐-油松)为研究对象,通过土钻烘干法测定0~500 cm土层土壤含水量,分析人工林土壤含水量的垂直特征。 结果 研究区典型人工林平均土壤含水量由大到小依次为侧柏、刺槐-侧柏、油松、刺槐-油松、刺槐,5种人工林土壤含水量差异显著(P<0.05),降水量及土壤质地是影响人工林土壤含水量的主要因素;在垂直方向上,人工林土壤含水量变异程度均为中等变异,变异系数随土层深度的增加呈减小趋势,约在360 cm土层形成稳定变化,人工植被主要影响0~360 cm土层内的土壤水,5种人工林中刺槐林地土壤水分变异程度最大,侧柏林地土壤水分最为稳定;采用有序聚类法将人工林土壤水分垂直层次划分为交换层、利用层、调节层和稳定层,不同林分间层次划分深度存在一定差异。 结论 研究区人工林可导致土壤含水量一定程度的减少,刺槐、油松、刺槐-油松林会加剧对深层土壤水分的消耗,其中刺槐林对深层土壤水分的消耗最为明显,建议在林业生态工程建设时,适当减少刺槐林面积,提高侧柏栽植比例。对现存土壤含水量较低的刺槐林,通过间伐降低林分密度,或将其改造为混交林,以期改善土壤水环境,实现林水平衡。图5表2参28 Abstract:Objective This study, with an investigation of the dynamic characteristics of 0 − 500 cm soil moisture in the restoration process of artificial forests in the loess region of western Shanxi Province, is aimed to provide theoretical basis for improving soil water environment of plantation and realizing forest water balance. Method First, with five typical plantations (Robinia pseudoacacia, Platycladus orientalis, Pinus tabuliformis, R. pseudoacacia-Platycladus orientalis, R. pseudoacacia-Pinus tabuliformis) selected as the research objects, oil drill drying method was employed to measure the soil moisture content of 0 − 500 cm before an anlysis was conducted of the vertical distribution and dynamic change characteristics of soil moisture in artificial forests. Result The average soil water content of plantations from large to small is Platycladus orientalis>R. pseudoacacia-Platycladus orientalis> Pinus tabuliformis>R. pseudoacacia-Pinus tabuliformis> R. pseudoacacia, showing a significant difference in the soil content and that precipitation and soil texture were the main factors affecting soil water content in plantations (P<0.05). Vertically, the variation degree of soil water content was moderate, and the variation coefficient decreased with the increase of soil depth with the variation of soil water content of each plantation becoming steady at a depth of about 360 cm whereas the artificial vegetation mainly affected the soil water in the range of 0 − 360 cm with the water variation of R. pseudoacacia forest being the highest, and that of Platycladus orientalis forest being the most stable. The vertical layers of soil water in the plantation were divided into exchange layer, use layer, regulation layer and stability layer by using the ordered clustering method with some differences in the depth of the layers among different stands. Conclusion The artificial forest land can reduce soil moisture to some extent in the study area and the consumption of deep soil water increases with the occurrence of drought in R. pseudoacacia, Pinus tabuliformis, R. pseudoacacia-Pinus tabuliformis, especially R. pseudoacacia forest. Therefore, it is suggested that during the construction of forestry ecological engineering, efforts should be made to reduce the planting of R. pseudoacacia forest and increase the planting of Platycladus orientalis in an appropriate manner, as well as transform the existing R. pseudoacacia forest with poor soil water status into a mixed forest, so as to improve the soil water environment and achieve forest water balance. [Ch, 5 fig. 2 tab. 28 ref.] -
图 1 人工林土壤容重、毛管孔隙度及颗粒组成特征
Figure 1 Soil bulk density, porosity and texture characteristics of plantation
图 2 典型人工林土壤含水量随时间的变化
Figure 2 Variation of soil moisture content over time in a typical plantation
图 5 人工林土壤含水量垂直有序聚类分层结果
Figure 5 Results of vertical ordered cluster stratification of soil water content in plantation
表 1 样地基本情况
Table 1. Basic information of the sample plot
人工林类型 样地坐标 海拔/
m坡度/
(°)郁闭度/
%林分密度/
(株·hm−2)凋落物蓄积量/
(t·hm−2)土壤容重/
(g·cm−3)有机质/
(g·kg−1)刺槐 36°16′23″N,110°45′39″E 1 090 25 65 1 825 8.30 1.17±0.12 6.50±1.60 侧柏 36°16′20″N,110°45′23″E 1 150 25 32 1 625 3.69 1.26±0.11 9.87±1.26 油松 36°16′21″N,110°45′45″E 1 140 28 53 1 700 14.08 1.25±0.09 6.38±0.52 刺槐-侧柏 36°16′24″N,110°45′49″E 1 120 26 45 1 725 9.75 1.20±0.09 9.83±1.34 刺槐-油松 36°16′24″N,110°45′43″E 1 120 27 55 1 800 11.14 1.17±0.08 11.00±3.35 说明:土壤容重、有机质数据为平均值±标准差。  下载: 导出CSV
下载: 导出CSV
表 2 人工林土壤含水量及其影响因子皮尔逊相关系数
Table 2. Pearson correlation coefficient of soil moisture content in plantation and its influencing factors
影响因素 刺槐 侧柏 油松 刺槐-侧柏 刺槐-油松 黏粒 0.537** 0.791** 0.512** 0.432* 0.775** 粉粒 0.452* 0.687** 0.425* −0.489* 0.431* 砂粒 −0.467** −0.749** −0.436* −0.545** −0.442* 土壤容重 −0.604** 0.177 0.079 0.159 −0.099 毛管孔隙度 0.310 0.184 0.208 0.405 0.093 说明:**代表极显著相关(P<0.01);*代表显著相关(P<0.05)。
下载: 导出CSV
-
[1] HUANG Laiming, SHAO Ming’an. Advances and perspectives on soil water research in China’s Loess Plateau [J/OL]. Earth-Science Reviews, 2019, 199: 102962[2023-04-01]. doi: 10.1016/j.earscirev.2019.102962. [2] 杨磊, 张子豪, 李宗善. 黄土高原植被建设与土壤干燥化: 问题与展望[J]. 生态学报, 2019, 39(20): 7382 − 7388. YANG Lei, ZHANG Zihao, LI Zongshan. Effects of large-scale re-vegetation on soil desiccation in the Loess Plateau: problems and perspectives [J]. Acta Ecologica Sinica, 2019, 39(20): 7382 − 7388. [3] 马婧怡, 贾宁凤, 程曼. 黄土丘陵区不同土地利用方式下土壤水分变化特征[J]. 生态学报, 2018, 38(10): 3471 − 3481. MA Jingyi, JIA Ningfeng, CHENG Man. Water characteristics of soil under different land-use types in the Loess Plateau region [J]. Acta Ecologica Sinica, 2018, 38(10): 3471 − 3481. [4] 姬王佳, 黄亚楠, 李冰冰, 等. 陕北黄土区深剖面不同土地利用方式下土壤水氢氧稳定同位素特征[J]. 应用生态学报, 2019, 30(12): 4143 − 4149. JI Wangjia, HUANG Ya’ nan, LI Bingbing, et al. Oxygen and hydrogen stable isotope compositions of soil water in deep loess profile under different land use types of northern Shaanxi, China [J]. Chinese Journal of Applied Ecology, 2019, 30(12): 4143 − 4149. [5] BAI Xiao, JIA Xiaoxu, JIA Yuhua, et al. Modeling long-term soil water dynamics in response to land-use change in a semi-arid area [J/OL]. Journal of Hydrology, 2020, 585: 124824[2023-04-01]. doi: 10.1016/j.jhydrol.2020.124824. [6] 杨敏, 赵西宁, 高晓东, 等. 黄土高原人工林深层土壤水分利用研究[J]. 水土保持研究, 2019, 26(6): 100 − 106. YANG Min, ZHAO Xining, GAO Xiaodong, et al. Deep soil moisture use of planted forests in the Loess Plateau [J]. Research of Soil and Water Conservation, 2019, 26(6): 100 − 106. [7] 张永旺, 万珊珊, 王俊, 等. 黄土高原植被演替过程中土壤水分亏缺[J]. 水土保持研究, 2020, 27(5): 120 − 125. ZHANG Yongwang, WAN Shanshan, WANG Jun, et al. Soil water deficit during vegetation succession on the Loess Plateau [J]. Research of Soil and Water Conservation, 2020, 27(5): 120 − 125. [8] 孙中峰, 周玉喜, 朱金兆, 等. 晋西黄土丘陵区坡面刺槐林地土壤水分研究[J]. 中国水土保持科学, 2007, 5(5): 43 − 49. SUN Zhongfeng, ZHOU Yuxi, ZHU Jinzhao, et al. Soil moisture of Robinia seudoacacia stands in the loess area in west of Shanxi Province [J]. Science of Soil and Water Conservation, 2007, 5(5): 43 − 49. [9] 李俊, 毕华兴, 李笑吟, 等. 有序聚类法在土壤水分垂直分层中的应用[J]. 北京林业大学学报, 2007, 29(1): 98 − 101. LI Jun, BI Huaxing, LI Xiaoyin, et al. Application of sequential clustering method in vertical dividing of soil moisture profiles [J]. Journal of Beijing Forestry University, 2007, 29(1): 98 − 101. [10] 于洋, 卫伟, 陈利顶, 等. 黄土丘陵区坡面整地和植被耦合下的土壤水分特征[J]. 生态学报, 2016, 36(11): 3441 − 3449. YU Yang, WEI Wei, CHENG Liding, et al. Coupling effects of different land preparation and vegetation on soil moisture characteristics in a semi-arid loess hilly region [J]. Acta Ecologica Sinica, 2016, 36(11): 3441 − 3449. [11] 陈洪松, 邵明安, 王克林. 黄土区荒草地和裸地土壤水分的循环特征[J]. 应用生态学报, 2005, 16(10): 1853 − 1857. CHEN Hongsong, SHAO Ming’ an, WANG Kelin. Water cycling characteristics of grassland and bare land soils on Loess Plateau [J]. Chinese Journal of Applied Ecology, 2005, 16(10): 1853 − 1857. [12] 何子淼, 肖培青, 郝仕龙, 等. 黄丘区野外坡面土壤水分变化对次降雨过程的响应[J]. 中国水土保持科学, 2018, 16(4): 16 − 24. HE Zimiao, XIAO Peiqing, HAO Shilong, et al. Response of soil moisture variation to individual rainfall on the field slope in the loessial hilly-gully region [J]. Science of Soil and Water Conservation, 2018, 16(4): 16 − 24. [13] 张晨成. 黄土高原退耕还林的土壤水分效应研究[D]. 北京: 中国科学院大学, 2017. ZHANG Chencheng. Study on Soil Water Effects of Returning Farmland to Forest on the Loess Plateau of China [D]. Beijiing: University of Chinese Academy of Sciences, 2017. [14] 施政乐, 张建军, 申明爽, 等. 晋西黄土区不同土地利用类型对土壤水分的影响[J]. 水土保持学报, 2021, 35(6): 190 − 197. SHI Zhengle, ZHANG Jianjun, SHEN Mingshuang, et al. Effects of different land use types on soil moisture in the loess region of western Shanxi Province [J]. Journal of Soil and Water Conservation, 2021, 35(6): 190 − 197. [15] 谭学进, 穆兴民, 高鹏, 等. 黄土区植被恢复对土壤物理性质的影响[J]. 中国环境科学, 2019, 39(2): 713 − 722. TAN Xuejin, MU Xingmin, GAO Peng, et al. Effects of vegetation restoration on changes to soil physical properties on the Loess Plateau [J]. China Environmental Science, 2019, 39(2): 713 − 722. [16] 冯棋, 杨磊, 王晶, 等. 黄土丘陵区植被恢复的土壤碳水效应[J]. 生态学报, 2019, 39(18): 6598 − 6609. FENG Qi, YANG Lei, WANG Jing, et al. Response of soil moisture and soil organic carbon to vegetation restoration in deep soil profiles in loess hilly region [J]. Acta Ecologica Sinica, 2019, 39(18): 6598 − 6609. [17] 杨良辰, 张健强, 杨新兵, 等. 华北土石山区7种优势乔木树种耗水分析[J]. 中南林业科技大学学报, 2019, 39(3): 69 − 75. YANG Liangchen, ZHANG Jianqiang, YANG Xinbin, et al. Water consumption analysis on seven dominant arbor tree species in earth and rock mountains in northern China [J]. Journal of Central South University of Forestry &Technology, 2019, 39(3): 69 − 75. [18] 刘新春, 赵勇钢, 刘小芳, 等. 晋西黄土区人工林细根与土壤水碳的耦合关系[J]. 生态学报, 2019, 39(21): 7987 − 7995. LIU Xinchun, ZHAO Yonggang, LIU Xiaofang, et al. Coupling fine roots with soil moisture and organic carbon in artificial forests in loess region of western Shanxi Province [J]. Acta Ecologica Sinica, 2019, 39(21): 7987 − 7995. [19] 席本野, 邸楠, 曹治国, 等. 树木吸收利用深层土壤水的特征与机制: 对人工林培育的启示[J]. 植物生态学报, 2018, 42(9): 885 − 905. XI Benye, DI Nan, CAO Zhiguo, et al. Characteristics and underlying mechanisms of plant deep soil water uptake and utilization: implication for the cultivation of plantation trees [J]. Chinese Journal of Plant Ecology, 2018, 42(9): 885 − 905. [20] 任婧宇, 乔雨宁, 闫璐瑶, 等. 黄土丘陵区主要树种土壤水分动态变化特征及影响因子[J]. 水土保持研究, 2022, 29(6): 73 − 80. RENG Jingyu, QIAO Yuning, YAN Luyao, et al. Dynamic changes and influencing factors of soil moisture of main tree species in loess hilly region [J]. Research of Soil and Water Conservation, 2022, 29(6): 73 − 80. [21] 马建业, 李占斌, 马波, 等. 黄土区小流域植被类型对沟坡地土壤水分循环的影响[J]. 生态学报, 2020, 40(8): 2698 − 2706. MA Jianye, LI Zhanbin, MA Bo, et al. Effects of vegetation types in small watershed on soil water cycle in gully-slope land of loess region [J]. Acta Ecologica Sinica, 2020, 40(8): 2698 − 2706. [22] 王宁, 毕华兴, 郭孟霞, 等. 晋西黄土残塬沟壑区刺槐人工林土壤水分植被承载力研究[J]. 水土保持学报, 2019, 33(6): 213 − 219. WANG Ning, BI Huaxing, GUO Mengxia, et al. Study on soil water vegetation carrying capacity of Robinia pseudoacacia plantation in the loess gully area of western Shanxi Province [J]. Journal of Soil and Water Conservation, 2019, 33(6): 213 − 219. [23] 邵臻, 张富, 陈瑾, 等. 陇中黄土丘陵沟壑区不同土地利用下土壤水分变化分析[J]. 干旱区资源与环境, 2017, 31(12): 129 − 135. SHAO Zhen, ZHANG Fu, CHENG Jin, et al. Soil moisture changes for different land use types in Loess Plateau gully and hilly region of central Gansu [J]. Journal of Arid Land Resources and Environment, 2017, 31(12): 129 − 135. [24] 岳宏昌, 王玉, 李缠云, 等. 黄土丘陵沟壑区土壤水分垂直分布研究[J]. 水土保持通报, 2009, 29(1): 66 − 69, 82. YUE Hongchang, WANG Yu, LI Chanyun, et al. Vertical distribution of soil moisture in the loess hilly and gully area [J]. Bulletin of Soil and Water Conservation, 2009, 29(1): 66 − 69, 82. [25] 刘新春, 赵勇钢, 刘小芳, 等. 晋西黄土区典型人工植被生长季深层土壤储水量与细根生物量分布特征[J]. 中国水土保持科学, 2019, 17(1): 95 − 101. LIU Xinchun, ZHAO Yonggang, LIU Xiaofang, et al. Distribution of moisture storage and fine root biomass in deep soil layers under typical artificial vegetation during the growing season in the Loess Plateau of western Shanxi, China [J]. Science of Soil and Water Conservation, 2019, 17(1): 95 − 101. [26] 陆蕴青. 黄土丘陵区高龄人工林土壤水分及根系吸水特征[D]. 杨凌: 西北农林科技大学, 2022. LU Yunqing. Characteristics of Soil Moisture and Root Water Absorption of Aged Plantation in Loess Hilly Region [D]. Yangling: Northwest A&F University, 2022. [27] 孙亚荣, 陈云明, 王亚娟, 等. 黄土丘陵区柠条人工林土壤水分动态变化特征及降雨特征对其影响[J]. 水土保持学报, 2023, 3(1): 1 − 8. SUN Yarong, CHEN Yunming, WANG Yajuan, et al. Dynamic variation characteristics of soil moisture in Caragana korshinskii plantation in loess hilly area and the influence of rainfall characteristics on it [J]. Journal of Soil and Water Conservation, 2023, 3(1): 1 − 8. [28] 王信增, 焦峰. 基于有序聚类法的土壤水分剖面划分[J]. 西北农林科技大学学报(自然科学版), 2011, 39(2): 191 − 196. WANG Xinzeng, JIAO Feng. Partition of soil moisture profiles based on sequential clustering method [J]. Journal of Northwest A&F University (Natural Science Edition), 2011, 39(2): 191 − 196. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230228
 点击查看大图
点击查看大图

计量
- 文章访问数: 2302
- HTML全文浏览量: 590
- PDF下载量: 59
- 被引次数: 0
