




-
城市森林不仅具有一般森林调节水、大气等生态循环的功能,同时还具有直接服务于居民的生态系统服务功能[1],其结构对其生态效益发挥有着直接影响。作为林地物种避难所、自然和半自然栖息地的残余,城市森林为城市鸟类提供了异质性的栖息地和友好的生存条件。而城市鸟类作为城市破碎森林中常见和显著的动物,为植物提供授粉、害虫控制、营养物质运输等生态服务功能[2-3],是城市生物多样性的重要组成部分,也是评价城市环境的因素之一[4-5]。以往对城市森林的研究更多地注重于城市森林植物群落景观效果、绿化遮阴、户外游憩功能等,在生态服务功能上也多着重植物群落主体的生态和社会效益,忽视了动物特别是与鸟类之间密切联系。国内外有关城市鸟类的研究也多以城市化为背景,探讨以城市鸟类为典型代表物种所受城市环境变化的影响[6-8],并通过调查鸟类群落、不同生境鸟类群落结构对比[9-10]、大小尺度上城市梯度效应和栖息地特征[11-13]等方面,分析城市化对鸟类群落结构变化及同质化程度的影响[14-15]。研究样地多集中在城市公园等各类城市绿地斑块中,但各斑块差异大,研究较少从城市森林结构层次分析对鸟类群落的影响。因而本研究以城市鸟类为指示物种,参考狭义上城市森林定义,选取安徽省合肥市老城区21处面积为0.5~2.5 hm2的城市森林斑块进行城市森林结构和鸟类指标调查。从树种组成、空间、基础服务设施、景观结构特征层次上分析与鸟类间的关系,探明优化城市森林结构,提高生态质量和生物多样性的方法,进而更好地发挥城市森林的生态功能,尤其是作为鸟类等野生动物栖息地的生态服务功能。
-
合肥市位于华东地区长江三角洲西端,江淮之间,30°56'~32°33'N,116°40'~117°58'E,属亚热带和暖温带过渡性的副热带季风气候区,地带性植被为落叶与常绿阔叶混交林,是首批国家园林城市之一。老城区位于合肥市中心地带,20世纪80年代以环城公园为核心,构筑了优良的绿化基础,具有良好的生态本底。随着合肥市“一湖一岭、两扇两翼、一核四区、多廊多点”[16]的森林城市空间布局建设,老城区整体绿化也得到了提升,现建成区绿地率达40.3%,绿化覆盖率达46.0%。
合肥市位于中国3条鸟类迁徙通道中的“东亚—澳大利亚”候鸟迁徙通道北缘[17],根据王歧山等[18]和吴海龙等[19]划分的安徽省5个动物地理区,合肥南北跨江淮丘陵区与沿江平原区,西接大别山区,独特的地理位置使动物尤其是鸟类多样性丰富。詹双侯等[20]对合肥地区鸟类最新研究显示:目前合肥地区共记录鸟类19目68科187属374种3亚种,是安徽省鸟类种类最多的城市。
-
合肥市具有良好的城市森林本底,样地选择兼顾森林斑块分布和鸟类活动特征,以向城市居民开放的城市森林绿地为主。参考狭义上城市森林规模的定义,即“面积大于0.5 hm2[21],地域群落的树干基部断面积之和达到5.5~28.0 m2·hm−2[22],且林木树冠覆盖度在10%~30%及以上”,在合肥市区一环内及其附近研究区域内初步选择21处相互间距大于300 m或大于100 m且有主干道阻隔,面积为0.5~2.5 hm2的城市森林斑块作为样地(表1)。
表 1 城市森林样地概况
Table 1. Overview of urban forest sample plots
编号 样地名称 面积/hm2 群落 主要群落组成 1 琥珀街道社区游园 1.10 针阔混交 水杉Metasequoia glyptostroboides+乌桕Sapium sebiferum+樟树Cinnamomun camphora-海桐Pittosporum tobira+山茶Camellia sinensis-地毯草Axonopus compressus 2 清溪东路环城林带 1.55 针阔混交 龙柏Sabina chinensis+法桐Platanus × acerifolia+构树Broussonetia papyrifera-桂花Osmanthus fragrans+紫薇Lagerstroemia indica-麦冬Ophiopogon japonicus 3 拱辰桥游园 0.85 针阔混交 构树+栾树Koelreuteria paniculata+女贞Ligustrum lucidum+石楠Photinia serratifolia-桂花+海桐-麦冬 4 古井桥与交通厅桥间林带 0.60 阔叶混交 构树+皂荚Gleditsia sinensis+刺槐Robinia pseudoacacia-棕榈Trachycarpus fortunei-麦冬 5 寿春路街旁游园 0.76 针阔混交 栾树+三角槭Acer buergerianum +桂花-石榴Punica granatum-玉簪Hosta plantaginea+麦冬 6 江淮风情园 1.10 针阔混交 广玉兰Magnolia grandiflora+桂花+樟树-海桐+红叶石楠Photinia × fraseri 7 西园新村社区公园 2.10 针阔混交 樟树+圆柏Juniperus chinensis -桂花+夹竹桃Juniperus chinensis +石榴Punica granatum-沿阶草Ophiopogon bodinieri+麦冬 8 合燃公园 0.86 针阔混交 水杉+樟树+银杏Ginkgo biloba-紫薇+红叶石楠-地毯草 9 飞虹小区游园 0.53 阔叶混交 樟树+紫叶李Prunus cerasifera f. atropurpurea -石榴+八角金盘Fatsia japonica -麦冬 10 四里河防护林带 1.10 针阔混交 矮冬青Ilex lohfauensis +石楠+樟树-桂花-早熟禾Poa annua+葱莲Zephyranthes candida 11 长岗游园 0.93 阔叶混交 栾树+樟树+鸡爪槭Acer palmatum -金边黄杨Euonymus japonicus ‘Aureo-marginatus’ +红叶石楠-萱草Hemerocallis fulva 12 环城西路莲花庵林带 1.05 针阔混交 女贞+枫杨Pterocarya stenoptera -棕榈+臭牡丹Clerodendrum bungei -麦冬+一年蓬Erigeron annuus 13 环城西路与淮河西路林带 0.83 阔叶混交 棕榈+刺槐+樟树-八角金盘+红叶石楠-蔓长春花Vinca major+沿阶草 14 环城南路防护林带 0.80 阔叶混交 女贞+樟树+栾树-棕榈+八角金盘-沿阶草 15 滁洲路与凤凰桥街旁游园 0.60 阔叶混交 桂花+广玉兰+栾树-夹竹桃Nerium oleander+女贞-沿阶草+狗牙根Cynodon dactylon 16 淮河路街旁游园 0.76 针阔混交 水杉+雪松Cedrus deodara +广玉兰-金边黄杨+日本珊瑚树Viburnum odoratissimum var. awabuki -蔓长春花 17 巢湖南路滨水公园 2.00 针阔混交 银杏+樟树+雪松-桂花+小叶女贞Ligustrum quihoui-早熟禾+狗牙根 18 小花园 0.52 针阔混交 樟树+法桐-桂花+南天竹Nandina domestica-沿阶草+麦冬 19 环城公园北路林带 0.77 针阔混交 女贞+刺槐+构树-棕榈-麦冬 20 五里墩立交桥林带 0.58 针阔混交 栾树+构树+紫叶李-桂花-麦冬 21 商鼎公园 1.15 针阔混交 银杏+无患子Sapindus mukorossi -杜鹃Rhododendron simsii+红叶石楠-早熟禾 -
于2020年6-12月对选定的斑块采用样方、样带法进行植物群落结构调查。依据形状面积大小,在0.5~1.5 hm2规模样地设立1个20 m×20 m的标准样方或长不小于20 m、面积375~425 m2的样带,1.5~2.5 hm2规模设立2个标准样方或样带,共设有样方或样带24 个。借助布鲁莱斯测高器(Blume-Leiss)、卷尺等工具获取样地乔灌草组成、冠幅、盖度等数据。采用经典抽样法记录灌木层和草本层植物种名、高度、覆盖度等特征,并手持全球定位仪(GPS)结合手机奥维地图软件,对样地内道路布局、长度、宽度标记和测量,对游憩设施座椅、环卫设施个数进行记录。
-
鸟类调查参考HJ 710.4—2014《生物多样性观测技术导则 鸟类》,结合常规调查手段,在2020年9月中旬至12上旬,以固定样线和样点结合的形式对21处样地鸟类组成特征进行观测,平均每个样地每月调查1~2次。
-
城市森林结构包括树种组成、空间、基础设施、景观结构特征[1,23]。选用合肥建成区2018年Spot卫星遥感影像图,参考2019年高德地图遥感卫星影像图,借助地理信息系统ArcGIS 10.2进行影像图拼接、地理坐标配准,并对各研究样地进行目视解译。树种组成结构包括植物物种、乔灌草丰富度、乔灌木多样性指数;空间结构包括林木树冠、绿化覆盖率、林分密度(胸高断面积和乔木密度);基础服务设施结构包括道路密度、道路及硬质铺装用地面积比例、建筑用地面积比例、游憩服务设施密度、道路围合率;景观结构包括斑块周长、斑块面积、形状指数(分维数)以及景观连通性,景观连通性指数通过ArcGIS与Conefor Inputs插件及 Conefor 2.6软件计算获得。以老城区中临泉路、潜山路、望江路、铜陵路围合成的约49 km2区域为背景,借助ArcGIS和卫星遥感影像对区域内城市森林景观斑块的数量、面积、分布进行统计。利用Conefor 2.6软件计算各斑块的斑块对整体连通性重要值(dIIC)和可能连通性重要值(dPC),参考相关研究[24]和一般林鸟日常活动范围,将斑块间的距离连通阈值分别设置在500、1 000、1 500 m,连通概率皆设置为0.5。量化区域内的栖息地斑块和链接对景观连通性的重要性。
-
分析鸟类丰富度、多度、Shannon-Wiener多样性指数、Pielou均匀度指数表示,并对鸟类群落结构进行优势种划分和相似度分析[25-26]。
-
以城市鸟类丰富度、多度、多样性指数、均匀度指数作为反映鸟类群落结构组成的指标,与城市森林组成、空间、基础服务设施和景观结构指标的相关因子进行Spearman相关性分析,结合主成分分析、回归分析探讨城市森林结构因子对城市森林中鸟类群落结构产生的影响,建立回归模型。
-
调查发现∶乔灌草90种,隶属51科76属;其中,乔木42种,灌木38种(含8种灌生形态乔木),草本18种。各样地植物丰富度为8~26种,其中:乔木丰富度为4~12种,多样性1.10~2.43;灌木丰富度为1~13种,多样性0~2.01;草本地被丰富度为0~6种。各样地植物组成差异比较明显,总体上以功能需求为主导。以游憩功能为主的样地,灌木、草本植物相比以防护功能为主的样地更为种类丰富。样地群落结构多为乔灌草模式,以针阔混交林、阔叶混交林为主,棕榈、樟树、桂花、雪松、女贞、紫薇等树种在样地中具有较高的应用。
-
垂直空间结构上,样地乔木树高5~10 m居多,平均树高为9.3 m;胸径以10~20 cm、20~30 cm居多,平均胸径为21.4 cm;平均冠幅为4.1 m,平均枝下高为2.6 m,森林斑块中大乔木规格的林木偏少。水平空间林分结构上,样地乔木密度为225~1 400株·hm−2,平均为678株·hm−2,胸高断面积为3.56~56.71 m2·hm−2,平均为26.63 m2·hm−2。其中样地21(商鼎公园)为近2 a新建商业公园,乔木多为小径级,胸高断面积仅为3.56 m2·hm−2,尚未达到5.5 m2·hm−2。各样地林木树冠覆盖面积为1 842~9 855 m2,不含水域面积各样地林木树冠覆盖率为16.1%~85.7%。除样地21外,其余皆达到了城市森林林木树冠覆盖率标准。
-
各样地道路密度为0~615 m·hm−2,道路与样地铺装面积为178~7 586 m2,道路与样地铺装率为3.1%~66.2%,样地建筑用地面积为68~2 540 m2,占地比例为0%~17.8%,主干道围合率为7.4%~65.6%,游憩设施密度为3~73个·hm−2。
-
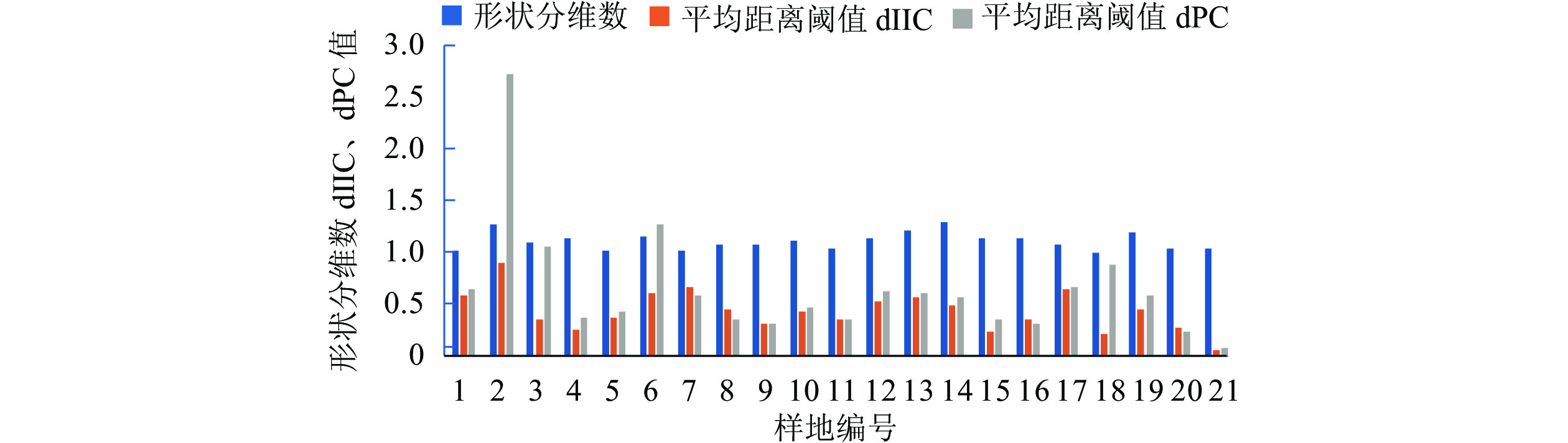
样地长宽比为1.4~55.0,形状分维数为1.0~1.3。老城区约49 km2区域内共1 095块城市森林景观斑块,斑块总面积约为432 hm2,其中最小斑块面积为189 m2,最大斑块12.1 hm2,平均斑块面积3 950 m2。各样地平均dIIC值为0.072~0.907,平均dPC值为0.084~2.743 (图1和图2),以往研究显示面积越大的斑块其dIIC值和dPC值也相对较高。本研究的样地斑块在整体上dIIC值和dPC值都偏低,对区域整体连通性贡献小。

图 1 研究区域景观结构指标特征图
Figure 1. Characteristic map of regional landscape structure index
-
由表2可见:研究区共记录鸟类8目22科38种,占合肥市记录鸟种的10.08%,其中雀形目Passeriformes 29种,非雀形目9种,最大值保留法计1 213只。居留型上,留鸟24种(63.16%),是合肥城市鸟类群落主要组成部分,夏候鸟1种(2.63%),冬候鸟9种(23.68%),旅鸟4种(10.53%);区系组成上,古北界种19种(50%),东洋界种14种(36.84%),广布种5种(13.16%)。古北界种与东洋界种相互渗透,这反映了合肥地理区系上的特殊性。
表 2 样地鸟类统计表
Table 2. Statistical table of birds in sample plots
目、科、种 居留型 地理型 保护级别 食性 出现样地编号 遇见率 鷉目 Podicipediformes 1 鷉科 Podicipedidae 小鷉 Tachybaptus ruficollis R 广 LC / 10、17 + 鹤形目 Gruiformes 2 秧鸡科 Rallidae 黑水鸡 Gallinula chloropus R 东 LC / 1 + 鸽形目 Columbiformes 3 鸠鸽科 Columbidae 山斑鸠 Streptopelia orientalis R 古 LC G 1、3、6、7、12、13、15~17、19~20 ++ 珠颈斑鸠 S. chinensis R 东 LC G 1~21 ++ 鹃形目 Cuculiformes 4 杜鹃科 Cuculidae 噪鹃 Eudynamys scolopacea S 东 LC/省一 O 18 + 佛法僧目 Coraciiformes 5 翠鸟科 Alcedinidae 普通翠鸟 Alcedo atthis R 广 LC / 1、7 + 戴胜目 Upupiformes 6 戴胜科 Upupidae 戴胜 Upupa epops R 广 LC I 3、11 + 鴷形目 Piciformes 7 啄木鸟科 Picidae 星头啄木鸟 Dendrocopos canicapillus R 古 LC/省一 I 3、13、17、20 + 大斑啄木鸟 D. major R 古 LC/省一 I 13 + 雀形目 Passeriformes 8 鹡鸰科 Motacillidae 白鹡鸰 Motacilla alba R 广 LC I 1~3、6、10、17、18、21 ++ 灰鹡鸰 M. cinerea P 古 LC I 7、17 + 9 鹎科 Pycnontidae 领雀嘴鹎 Spizixos semitorques R 东 LC O 4 + 白头鹎 Pycnonotus sinensis R 东 LC O 1~20 +++ 10 伯劳科 Laniidae 棕背伯劳 Lanius schach R 东 LC/省二 P 5、10、16、17 + 11 椋鸟科Sturnidae 八哥 Acridotheres cristatellus R 东 LC O 2、7、17 + 丝光椋鸟 Sturnus sericeus R 东 LC I 13 + 12 鸦科 Corvidae 灰喜鹊 Cyanopica cyana R 古 LC/省一 O 1~4、6~21 +++ 喜鹊 Pica pica R 广 LC O 2、4~8、11、14、18、19 ++ 13 鸫科 Turdidae 红尾歌鸲 Luscinia sibilans W 古 LC I 19 + 红胁蓝尾鸲 Tarsiger cyanurus W 古 LC I 13、16 + 鹊鸲 Copsychus saularis R 东 LC I 1~9、11~20 ++ 北红尾鸲 Phoenicurus auroreus W 古 LC I 9~12、17 ++ 灰背鸫 Turdus hortulorum W 古 LC I 7、12、14 + 乌鸫 T. merula R 东 LC I 1~8、10、12~17、19、20 ++ 14 鹟科 Muscicapidae 鸲姬鹟 Ficedula mugimaki P 古 LC I 12、13 + 15 画眉科 Timallidae 黑脸噪鹛 Garrulax perspicillatus R 东 LC O 2、12、20 ++ 16 鸦雀科 Paradoxornithidae 棕头鸦雀 Paradoxornis webbianus R 东 LC I 2 + 17 莺科 Sylviidae 黄腰柳莺 Phylloscopus proregulus W 古 LC I 1~3、7、9、17、20 ++ 黄眉柳莺 P. inornatus P 古 LC I 12~15、18 ++ 极北柳莺 P. borealis P 古 LC I 20 + 18 长尾山雀科 Aegithalidae 银喉长尾山雀 Aegithalos caudatus R 古 LC I 6 + 红头长尾山雀 A. concinnus R 东 LC I 6、20 ++ 19 山雀科 Paridae 黄腹山雀 Parus venustulus W 东 LC I 6、7 ++ 大山雀 P. major R 古 LC I 6 + 20 雀科 Passeridae 树麻雀 Passer montanus R 古 LC O 1~3、5~9、11、18、20、21 +++ 21 燕雀科 Fringillidae 燕雀 Fringilla montifringilla W 古 LC G 2 ++ 黑尾蜡嘴雀 Eophona migratoria W 古 LC G 4、6、7、12~14、20 ++ 22 鹀科 Emberizidae 白眉鹀 Emberiza tristrami W 古 LC G 19 + 说明:居留型中,S为夏候鸟、W为冬候鸟、R为留鸟、P为旅鸟;地理型中,广为广布种、东为东洋界种、古为古北界种;保护级别 中,LC为世界自然保护联盟(IUCN)中无危物种,省一为安徽省一级保护动物,省二为安徽省二级保护动物;遇见率中,+++为 优势种;++为常见种,+少见种;食性中,I为食虫鸟类,G为食谷鸟类,O为杂食鸟类,P为食肉性,/为水域性鸟类 -
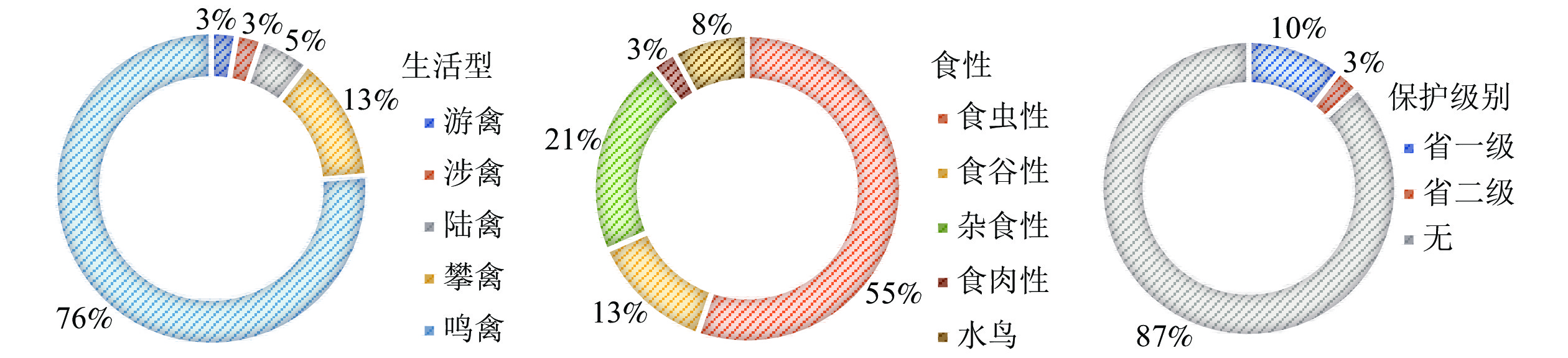
从食性上分析(不考虑水鸟),食虫性鸟类与杂食性鸟类占比高达76%(图2),可见,城市森林为食虫性和杂食性鸟类提供了丰富的食物来源。从保护级别分析,以常见鸟类为主,38种鸟类为无危(LC)物种,安徽省省级保护鸟类仅有5种。优势种为白头鹎Pycnonotus sinensis、灰喜鹊Cyanopica cyana、树麻雀Passer montanus,皆为杂食性、集群性鸟类。城市背景下,它们对城市化环境具有较强的适应性,相比其他鸟类更具有生存优势。在与城市居民相处中,此类鸟更爱伴人而居,典型的以树麻雀为代表,实际调查中远离居住区或以防护为主的森林斑块并不受树麻雀欢迎。
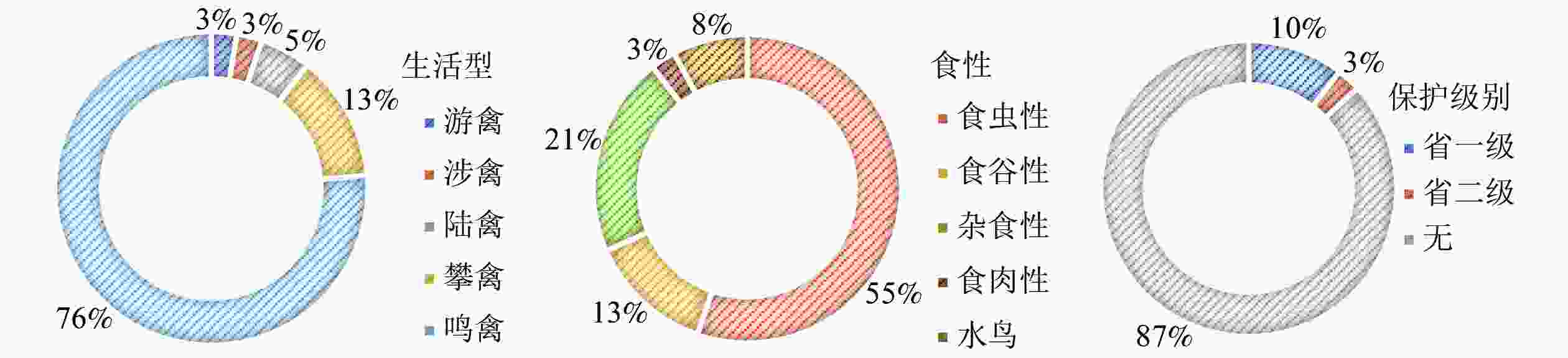
图 2 城市鸟类生活型、食性即保护级别构成
Figure 2. Composition of life style, feeding habit and protection level of urban birds
-
样地鸟类丰富度4~15种,多度11~122,多样性指数为1.34~2.50,均匀度指数为0.37~0.69。其中样地21未达到城市森林标准,其鸟类丰富度、多度、多样性及均匀度指数都远低于其他样地。相似度分析(表3)显示:城市森林斑块在鸟类组成上具有较高的相似性,相似度值为0.381~0.857,均值为0.588。鸟类组成上除却常见鸟种外,部分鸟类只出现在特定的生境中,例如红尾歌鸲Luscinia sibilans、红胁蓝尾鸲Tarsiger cyanurus、白眉鹀Emberiza tristrami和鸲姬鹟Ficedula mugimaki只在环城公园林木密集隐蔽的样地中出现。
表 3 样地鸟类相似度矩阵
Table 3. Bird similarity matrix chart in sample plot
样地 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 1 1 0.667 0.818 0.526 0.556 0.640 0.692 0.667 0.667 0.526 0.526 0.609 0.583 0.600 0.778 0.632 0.640 0.700 0.600 0.667 0.533 2 1 0.667 0.571 0.600 0.593 0.643 0.700 0.600 0.476 0.571 0.480 0.385 0.455 0.500 0.476 0.593 0.636 0.545 0.615 0.471 3 1 0.526 0.556 0.640 0.615 0.667 0.667 0.526 0.632 0.522 0.583 0.500 0.667 0.632 0.720 0.600 0.600 0.750 0.533 4 1 0.667 0.636 0.609 0.800 0.533 0.500 0.625 0.600 0.571 0.824 0.667 0.625 0.455 0.588 0.706 0.571 0.333 5 1 0.571 0.545 0.857 0.571 0.533 0.667 0.421 0.400 0.625 0.571 0.667 0.476 0.625 0.625 0.500 0.364 6 1 0.690 0.667 0.476 0.455 0.545 0.538 0.519 0.609 0.571 0.545 0.500 0.609 0.609 0.667 0.444 7 1 0.636 0.545 0.348 0.522 0.593 0.500 0.667 0.545 0.522 0.621 0.500 0.583 0.643 0.316 8 1 0.714 0.533 0.800 0.526 0.500 0.750 0.714 0.667 0.476 0.750 0.750 0.600 0.545 9 1 0.533 0.800 0.526 0.400 0.500 0.571 0.533 0.571 0.625 0.500 0.600 0.545 10 1 0.500 0.500 0.381 0.471 0.533 0.625 0.727 0.471 0.471 0.381 0.500 11 1 0.500 0.381 0.588 0.533 0.500 0.455 0.706 0.588 0.476 0.533 12 1 0.720 0.762 0.737 0.600 0.538 0.476 0.571 0.640 0.250 13 1 0.636 0.700 0.667 0.519 0.455 0.545 0.615 0.235 14 1 0.750 0.588 0.435 0.667 0.667 0.545 0.308 15 1 0.800 0.571 0.625 0.750 0.600 0.364 16 1 0.636 0.471 0.706 0.571 0.333 17 1 0.435 0.522 0.519 0.333 18 1 0.556 0.455 0.615 19 1 0.545 0.308 20 1 0.353 21 1 -
对符合城市森林定义的20个样地按照有无水域划分为2组,近水、含水域有样地1~4、6、7、10、15、17、18,无水域有样地5、8、9、11~14、16、19、20。利用SPSS 25.0进行非参数秩和检验,结果显示:有无水域类别上,鸟类丰富度、多度、多样性以及均匀度整体上并无显著差异。有水域样地为临近南淝河或含有小面积水域斑块,不足以为众多水鸟提供栖息资源,且南淝河驳岸皆为硬质化驳岸,植物覆盖低,不利于鸟类生存。实际观测中,有水域的样地仅有小鷉、普通翠鸟、黑水鸡、白鹡鸰、灰鹡鸰等5种喜水域鸟类,数量稀少。
-
Spearman相关性分析显示:鸟类丰富度与树种组成结构中的乔木物种丰富度(r=0.664,P<0.01),乔木多样性(r=0.570,P<0.01)极显著正相关。与空间结构中林木树冠覆盖面积(r=0.696,P<0.01)、绿化覆盖面积(r=0.577,P<0.01)极显著正相关,与样地乔木密度(r=0.484,P<0.05)显著正相关。与基础设施结构中道路密度(r=−0.459,P<0.05)显著负相关。与景观结构中样地面积(r=0.585,P<0.01)、斑块对整体连通性重要值(dIIC)(r=0.638,P<0.01)、可能连通性重要值(dPC)(r=0.634,P<0.01)极显著正相关。
-
鸟类多度在树种组成结构上与乔木物种丰富度(r=0.489,P<0.05)显著正相关;在空间结构上与林木树冠覆盖面积(r=0.657,P<0.01)极显著正相关,与绿化覆盖面积(r=0.549,P<0.05)显著正相关;在基础设施结构上与主干道围合率(r=−0.471,P<0.05)显著负相关;在景观结构上与样地面积(r=0.625,P<0.01)、dIIC值(r=0.635,P<0.01)极显著正相关,与dPC值(r=0.473,P<0.05)显著正相关。
-
鸟类多样性在树种组成结构上与乔木物种丰富度(r=0.601,P<0.01)极显著正相关,与乔木多样性(r=0.504,P<0.05)显著正相关;在空间结构上与林木树冠覆盖面积(r=0.625,P<0.01)极显著正相关,与乔木密度(r=0.487,P<0.05)、绿化覆盖面积(r=0.482,P<0.05)显著正相关;在基础设施结构上与道路密度(r=−0.526,P<0.05)显著负相关;在景观结构上与dIIC值(r=0.615,P<0.01)极显著正相关,与dPC值(r=0.515,P<0.05)显著正相关。
-
鸟类均匀度在树种组成结构上与乔木物种丰富度(r=0.594,P<0.01)极显著正相关,与乔木多样性(r=0.504,P<0.05)显著正相关;在空间结构上与林木树冠覆盖面积(r=0.617,P<0.01)极显著正相关,与乔木密度(r=0.484,P<0.05)、绿化覆盖面积(r=0.464,P<0.05)显著正相关;在基础设施结构上与道路密度(r=−0.512,P<0.05)显著负相关;在景观结构上与dIIC值(r=0.617,P<0.01)极显著正相关,与dPC值(r=0.512,P<0.05)显著正相关。
-
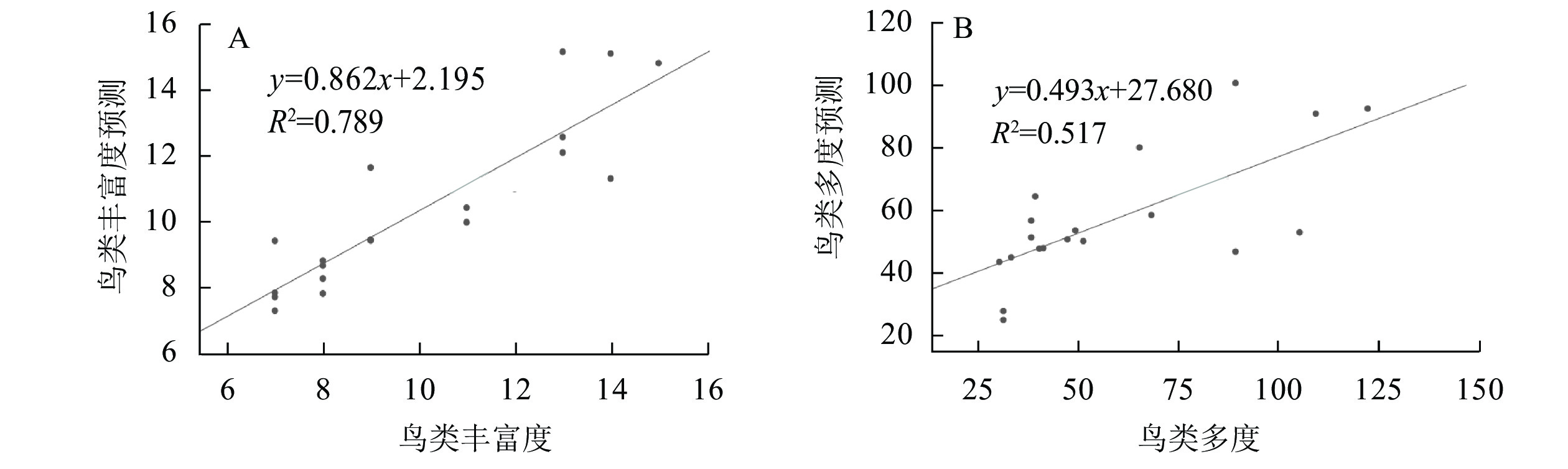
①城市森林结构与鸟类丰富度间的回归分析。对影响样地鸟类丰富度的自变量进行主成分分析,同时利用逐步回归法得到最优方程,其变量与主成分分析一致:RB=0.399RT+0.005DT+3.176A+0.755。其中:RB为鸟类丰富度(种);RT为乔木丰富度;DT为乔木密度(株·hm−2);A为样地面积(hm2)。模型拟合度为0.888,R2为0.789,调整后的R2为0.749,方差检验结果显示P<0.01,回归效果较好,方差扩大因子法(VIF)检验不存在共线性。将原数据代入回归模型进行检验,得到的鸟类丰富度预测值与观测值进行相关分析,在0.01显著水平下预测值与观测值相关系数为0.908,两者显著相关。将预测值(y)与观测值(x)做回归分析,模型结果为y`=0.802x+2.195,拟合优度R2为0.789,观测值在回归直线附近波动,说明回归效果较好(图3A)。②城市森林结构与鸟类多度间的回归分析。对影响样地鸟类多度的自变量进行主成分分析,同时利用逐步回归法得到最优方程:AB=30A−0.945RER+73.26。其中:AB为鸟类多度(只);RA为样地面积(hm2); RRE为道路围合率(%)。模型拟合度为0.720,R2为0.518,调整后的R2为0.462,方差检验结果显示P<0.01,方差扩大因子法(VIF)检验不存在共线性。原数据代入回归模型进行检验,得到的鸟类多度预测值与观测值进行相关分析,在0.01显著水平下预测值与观测值相关系数为0.710,两者显著相关,将预测值(y)与观测值(x)做回归分析模型结果为y`=0.493x+27.68,拟合优度R2为0.517,观测值在回归直线附近波动,说明回归效果较好(图3B)。③城市森林结构与鸟类多样性、均匀度间的回归分析。样地鸟类多样性和均匀度的相关性回归分析中,自变量一致,对自变量进行主成分分析,利用逐步回归法得到关于城市森林结构与样地鸟类多样性、均匀度的回归方程显示R2<0.5,回归模型并不能起到解释因变量的作用,故将方程舍弃。
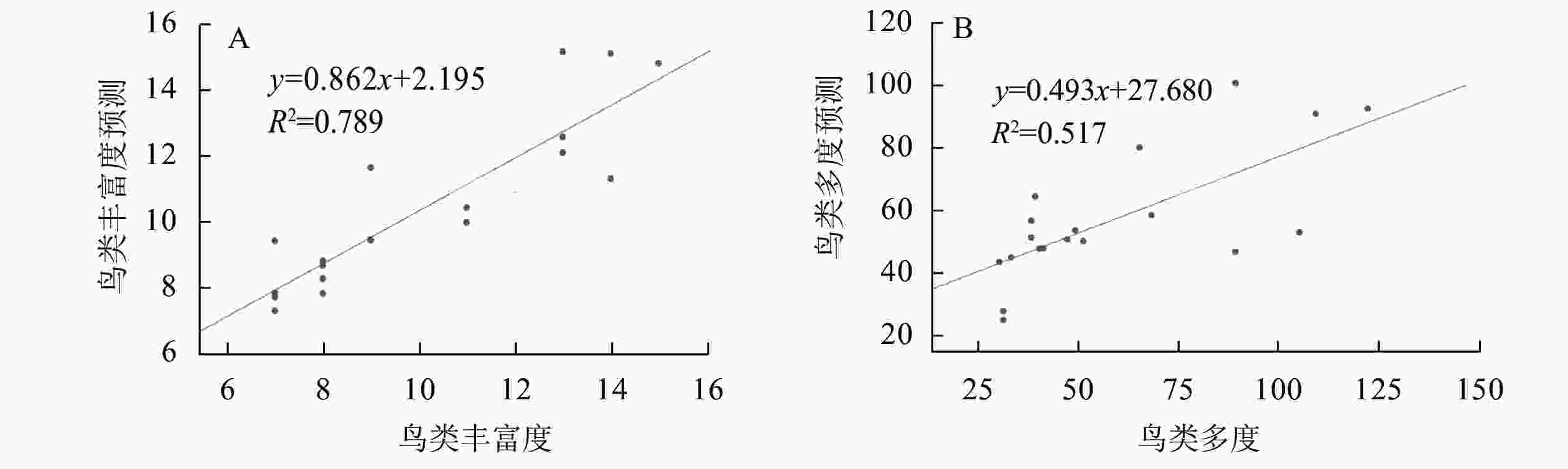
图 3 鸟类丰富度回归检验(A)和鸟类多度回归检验(B)
Figure 3. Regression analysis of bird richness (A) and bird abundance (B)
-
本研究记录乔灌草90种,总体上样地乔灌草丰富度由其功能需求决定,以游憩服务功能为主的斑块其灌木、草本种类更为丰富。样地基础设施密度都维持在较低的水平,受面积影响,各斑块对区域整体景观连通性贡献偏小。样地鸟类共记录8目22科38种,计1 213只。食虫性与杂食性鸟类占比高达76%,为城市鸟类群落主要组成成分。城市森林提供了丰富的食物来源,因而杂食性、集群性的白头鹎、灰喜鹊和树麻雀成为优势种。各样地鸟类组成具有较高的相似性,相似度均值为0.588,存在同质化现象。环城公园一带乔木密度高、斑块连通性高的样地能为红尾歌鸲等少见鸟种提供栖息地。
从分析结果来看,城市森林对城市鸟类的吸引力取决于多个结构层次,尽管有无水域对鸟类群落影响不显著,但树种组成、空间、基础设施、景观结构对鸟类都有着不同程度的影响,其中,样地乔木丰富度、林木树冠覆盖面积、样地绿化覆盖面积、斑块的景观连通性重要值(dIIC)、可能连通性重要值(dPC)对样地鸟类群落丰富度、多度、多样性、均匀度影响显著。进一步分析发现,影响鸟类丰富度的因素可以归为3个方面:一是斑块面积大小和区域内斑块的连通性,这代表着城市森林样地中所拥有的资源量和潜在可获取的资源;二是乔木组成的丰富性和多样性;三是样地空间上的利用,如道路密度与乔木密度,两者表达了单位面积内斑块所拥有的资源富集程度。
观测鸟类是监测城市森林栖息地生境变化和生态服务功能的一种有效手段。本研究从城市森林结构层次分析证实了城市森林与城市鸟类之间的密切联系,也是对合肥城市生物多样性研究的重要补充。鉴于研究样本容量有限,得出的回归方程只能在一定程度上起到解释作用。鸟类相比其他动物具有更为宽广的活动范围,往后研究可侧重于区域背景下城市森林斑块之间的连通、布局等因素对城市鸟类种群迁飞、联系、觅食活动的影响。
Relationship between urban forest structure and bird habitat in autumn in Hefei City
-
摘要:
目的 为优化城市森林结构,提高城市森林斑块生态服务功能,以鸟类为指标,阐述以城市森林结构为基础的城市鸟类栖息响应机制。 方法 以安徽省合肥市为研究区域,选取21处面积在0.5~2.5 hm2的城市森林斑块为样地,采用样方法进行植物群落及结构特征调查,以固定样线和样点结合的形式在秋季观测记录选取的样地鸟类群落组成。结合ArcGIS与卫星影像图,量化城市森林树种组成、空间、基础设施、景观结构相关指标,利用相关分析、主成分分析、回归分析明确与城市鸟类丰富度、多度、Shannon指数、Pielou均匀度指标间的潜在关系。 结果 ①研究记录样地植被90种,隶属51科76属;鸟类8目22科38种,占合肥市记录鸟种的10.08%,最大值保留法计1 213只;城市鸟类优势种为白头鹎Pycnonotus sinensis、灰喜鹊Cyanopica cyana、树麻雀Passer montanus,各斑块之间的鸟类组成有着较高的相似度,相似度均值为0.588。②非参数秩和检验表明在有无水域条件上,鸟类群落差异不明显,城市森林树种组成、空间、基础设施、景观结构对鸟类指标都有着不同程度的影响。③利用主成分分析进一步阐述影响因素,可将影响鸟类丰富度的因素归为城市森林斑块所拥有的资源量和潜在可获取的资源、乔木的丰富度和多样性、单位面积下斑块的资源富集程度3个方面,进一步分析得到关于鸟类丰富度与鸟类多度的方程。 结论 城市森林对城市鸟类的吸引力取决于多个结构层次,能反映城市森林拥有的资源量和潜在可获取的资源、乔木丰富度和多样性、单位面积资源富集程度的指标是影响鸟类栖息的重要因素。图3表3参26 Abstract:Objective This study is designed to explore the response mechanism of urban bird habitat based on urban forest structure with birds as indicators, so as to optimize urban forest structure and improve the ecological service function of urban forest patches. Method Taking Hefei City as the study area, 21 urban forest patches with an area of 0.5−2.5 hm2 were selected as sample plots. The characteristics of plant community and structure were investigated by sampling method, and the composition of bird community in selected sample plots was observed and recorded in the form of fixed sample line and sample point in autumn. Combined with ArcGIS and satellite images, the relevant indicators of urban forest tree species composition, space, infrastructure and landscape structure were quantified, and correlation analysis, principal component analysis and regression analysis were used to clarify the potential relationship with urban bird richness, abundance, Shannon index and Pielou evenness. Result (1) 90 species of vegetation belonging to 51 families and 76 genera were recorded. 38 species of birds in 22 families and 8 orders were recorded, accounting for 10.08% of the recorded bird species in Hefei, and 1213 birds were counted by maximum retention method. The dominant species of urban birds were Pycnonotus sinensis, Cyanopica cyana and Passer montanus, and the bird composition between patches had a high similarity, with an average similarity of 0.588. (2) Non-parametric rank sum test showed that there was no obvious difference in bird community with or without water area, and the composition of urban forest tree species, space, infrastructure and landscape structure had different degrees of impact on bird indicators. (3) Principal component analysis showed that the factors affecting bird richness could be classified into three aspects: the amount of resources and potentially available resources of urban forest patches, the richness and diversity of trees, and the resource enrichment degree of patches per unit area, and the equations of bird richness and bird abundance could be obtained by further analysis. Conclusion The attraction of urban forest to urban birds depends on multiple structural levels. The indicators that can reflect the amount of resources and the potentially available resources of urban forest, tree richness and diversity, and resource enrichment degree per unit area are important factors affecting bird habitat. [Ch, 3 fig. 3 tab. 26 ref.] -
Key words:
- urban forest /
- urban birds /
- forest structure /
- biodiversity /
- Hefei City
-
图 2 城市鸟类生活型、食性即保护级别构成
Figure 2 Composition of life style, feeding habit and protection level of urban birds
图 3 鸟类丰富度回归检验(A)和鸟类多度回归检验(B)
Figure 3 Regression analysis of bird richness (A) and bird abundance (B)
表 1 城市森林样地概况
Table 1. Overview of urban forest sample plots
编号 样地名称 面积/hm2 群落 主要群落组成 1 琥珀街道社区游园 1.10 针阔混交 水杉Metasequoia glyptostroboides+乌桕Sapium sebiferum+樟树Cinnamomun camphora-海桐Pittosporum tobira+山茶Camellia sinensis-地毯草Axonopus compressus 2 清溪东路环城林带 1.55 针阔混交 龙柏Sabina chinensis+法桐Platanus × acerifolia+构树Broussonetia papyrifera-桂花Osmanthus fragrans+紫薇Lagerstroemia indica-麦冬Ophiopogon japonicus 3 拱辰桥游园 0.85 针阔混交 构树+栾树Koelreuteria paniculata+女贞Ligustrum lucidum+石楠Photinia serratifolia-桂花+海桐-麦冬 4 古井桥与交通厅桥间林带 0.60 阔叶混交 构树+皂荚Gleditsia sinensis+刺槐Robinia pseudoacacia-棕榈Trachycarpus fortunei-麦冬 5 寿春路街旁游园 0.76 针阔混交 栾树+三角槭Acer buergerianum +桂花-石榴Punica granatum-玉簪Hosta plantaginea+麦冬 6 江淮风情园 1.10 针阔混交 广玉兰Magnolia grandiflora+桂花+樟树-海桐+红叶石楠Photinia × fraseri 7 西园新村社区公园 2.10 针阔混交 樟树+圆柏Juniperus chinensis -桂花+夹竹桃Juniperus chinensis +石榴Punica granatum-沿阶草Ophiopogon bodinieri+麦冬 8 合燃公园 0.86 针阔混交 水杉+樟树+银杏Ginkgo biloba-紫薇+红叶石楠-地毯草 9 飞虹小区游园 0.53 阔叶混交 樟树+紫叶李Prunus cerasifera f. atropurpurea -石榴+八角金盘Fatsia japonica -麦冬 10 四里河防护林带 1.10 针阔混交 矮冬青Ilex lohfauensis +石楠+樟树-桂花-早熟禾Poa annua+葱莲Zephyranthes candida 11 长岗游园 0.93 阔叶混交 栾树+樟树+鸡爪槭Acer palmatum -金边黄杨Euonymus japonicus ‘Aureo-marginatus’ +红叶石楠-萱草Hemerocallis fulva 12 环城西路莲花庵林带 1.05 针阔混交 女贞+枫杨Pterocarya stenoptera -棕榈+臭牡丹Clerodendrum bungei -麦冬+一年蓬Erigeron annuus 13 环城西路与淮河西路林带 0.83 阔叶混交 棕榈+刺槐+樟树-八角金盘+红叶石楠-蔓长春花Vinca major+沿阶草 14 环城南路防护林带 0.80 阔叶混交 女贞+樟树+栾树-棕榈+八角金盘-沿阶草 15 滁洲路与凤凰桥街旁游园 0.60 阔叶混交 桂花+广玉兰+栾树-夹竹桃Nerium oleander+女贞-沿阶草+狗牙根Cynodon dactylon 16 淮河路街旁游园 0.76 针阔混交 水杉+雪松Cedrus deodara +广玉兰-金边黄杨+日本珊瑚树Viburnum odoratissimum var. awabuki -蔓长春花 17 巢湖南路滨水公园 2.00 针阔混交 银杏+樟树+雪松-桂花+小叶女贞Ligustrum quihoui-早熟禾+狗牙根 18 小花园 0.52 针阔混交 樟树+法桐-桂花+南天竹Nandina domestica-沿阶草+麦冬 19 环城公园北路林带 0.77 针阔混交 女贞+刺槐+构树-棕榈-麦冬 20 五里墩立交桥林带 0.58 针阔混交 栾树+构树+紫叶李-桂花-麦冬 21 商鼎公园 1.15 针阔混交 银杏+无患子Sapindus mukorossi -杜鹃Rhododendron simsii+红叶石楠-早熟禾  下载: 导出CSV
下载: 导出CSV
表 2 样地鸟类统计表
Table 2. Statistical table of birds in sample plots
目、科、种 居留型 地理型 保护级别 食性 出现样地编号 遇见率 鷉目 Podicipediformes 1 鷉科 Podicipedidae 小鷉 Tachybaptus ruficollis R 广 LC / 10、17 + 鹤形目 Gruiformes 2 秧鸡科 Rallidae 黑水鸡 Gallinula chloropus R 东 LC / 1 + 鸽形目 Columbiformes 3 鸠鸽科 Columbidae 山斑鸠 Streptopelia orientalis R 古 LC G 1、3、6、7、12、13、15~17、19~20 ++ 珠颈斑鸠 S. chinensis R 东 LC G 1~21 ++ 鹃形目 Cuculiformes 4 杜鹃科 Cuculidae 噪鹃 Eudynamys scolopacea S 东 LC/省一 O 18 + 佛法僧目 Coraciiformes 5 翠鸟科 Alcedinidae 普通翠鸟 Alcedo atthis R 广 LC / 1、7 + 戴胜目 Upupiformes 6 戴胜科 Upupidae 戴胜 Upupa epops R 广 LC I 3、11 + 鴷形目 Piciformes 7 啄木鸟科 Picidae 星头啄木鸟 Dendrocopos canicapillus R 古 LC/省一 I 3、13、17、20 + 大斑啄木鸟 D. major R 古 LC/省一 I 13 + 雀形目 Passeriformes 8 鹡鸰科 Motacillidae 白鹡鸰 Motacilla alba R 广 LC I 1~3、6、10、17、18、21 ++ 灰鹡鸰 M. cinerea P 古 LC I 7、17 + 9 鹎科 Pycnontidae 领雀嘴鹎 Spizixos semitorques R 东 LC O 4 + 白头鹎 Pycnonotus sinensis R 东 LC O 1~20 +++ 10 伯劳科 Laniidae 棕背伯劳 Lanius schach R 东 LC/省二 P 5、10、16、17 + 11 椋鸟科Sturnidae 八哥 Acridotheres cristatellus R 东 LC O 2、7、17 + 丝光椋鸟 Sturnus sericeus R 东 LC I 13 + 12 鸦科 Corvidae 灰喜鹊 Cyanopica cyana R 古 LC/省一 O 1~4、6~21 +++ 喜鹊 Pica pica R 广 LC O 2、4~8、11、14、18、19 ++ 13 鸫科 Turdidae 红尾歌鸲 Luscinia sibilans W 古 LC I 19 + 红胁蓝尾鸲 Tarsiger cyanurus W 古 LC I 13、16 + 鹊鸲 Copsychus saularis R 东 LC I 1~9、11~20 ++ 北红尾鸲 Phoenicurus auroreus W 古 LC I 9~12、17 ++ 灰背鸫 Turdus hortulorum W 古 LC I 7、12、14 + 乌鸫 T. merula R 东 LC I 1~8、10、12~17、19、20 ++ 14 鹟科 Muscicapidae 鸲姬鹟 Ficedula mugimaki P 古 LC I 12、13 + 15 画眉科 Timallidae 黑脸噪鹛 Garrulax perspicillatus R 东 LC O 2、12、20 ++ 16 鸦雀科 Paradoxornithidae 棕头鸦雀 Paradoxornis webbianus R 东 LC I 2 + 17 莺科 Sylviidae 黄腰柳莺 Phylloscopus proregulus W 古 LC I 1~3、7、9、17、20 ++ 黄眉柳莺 P. inornatus P 古 LC I 12~15、18 ++ 极北柳莺 P. borealis P 古 LC I 20 + 18 长尾山雀科 Aegithalidae 银喉长尾山雀 Aegithalos caudatus R 古 LC I 6 + 红头长尾山雀 A. concinnus R 东 LC I 6、20 ++ 19 山雀科 Paridae 黄腹山雀 Parus venustulus W 东 LC I 6、7 ++ 大山雀 P. major R 古 LC I 6 + 20 雀科 Passeridae 树麻雀 Passer montanus R 古 LC O 1~3、5~9、11、18、20、21 +++ 21 燕雀科 Fringillidae 燕雀 Fringilla montifringilla W 古 LC G 2 ++ 黑尾蜡嘴雀 Eophona migratoria W 古 LC G 4、6、7、12~14、20 ++ 22 鹀科 Emberizidae 白眉鹀 Emberiza tristrami W 古 LC G 19 + 说明:居留型中,S为夏候鸟、W为冬候鸟、R为留鸟、P为旅鸟;地理型中,广为广布种、东为东洋界种、古为古北界种;保护级别 中,LC为世界自然保护联盟(IUCN)中无危物种,省一为安徽省一级保护动物,省二为安徽省二级保护动物;遇见率中,+++为 优势种;++为常见种,+少见种;食性中,I为食虫鸟类,G为食谷鸟类,O为杂食鸟类,P为食肉性,/为水域性鸟类
下载: 导出CSV
表 3 样地鸟类相似度矩阵
Table 3. Bird similarity matrix chart in sample plot
样地 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 1 1 0.667 0.818 0.526 0.556 0.640 0.692 0.667 0.667 0.526 0.526 0.609 0.583 0.600 0.778 0.632 0.640 0.700 0.600 0.667 0.533 2 1 0.667 0.571 0.600 0.593 0.643 0.700 0.600 0.476 0.571 0.480 0.385 0.455 0.500 0.476 0.593 0.636 0.545 0.615 0.471 3 1 0.526 0.556 0.640 0.615 0.667 0.667 0.526 0.632 0.522 0.583 0.500 0.667 0.632 0.720 0.600 0.600 0.750 0.533 4 1 0.667 0.636 0.609 0.800 0.533 0.500 0.625 0.600 0.571 0.824 0.667 0.625 0.455 0.588 0.706 0.571 0.333 5 1 0.571 0.545 0.857 0.571 0.533 0.667 0.421 0.400 0.625 0.571 0.667 0.476 0.625 0.625 0.500 0.364 6 1 0.690 0.667 0.476 0.455 0.545 0.538 0.519 0.609 0.571 0.545 0.500 0.609 0.609 0.667 0.444 7 1 0.636 0.545 0.348 0.522 0.593 0.500 0.667 0.545 0.522 0.621 0.500 0.583 0.643 0.316 8 1 0.714 0.533 0.800 0.526 0.500 0.750 0.714 0.667 0.476 0.750 0.750 0.600 0.545 9 1 0.533 0.800 0.526 0.400 0.500 0.571 0.533 0.571 0.625 0.500 0.600 0.545 10 1 0.500 0.500 0.381 0.471 0.533 0.625 0.727 0.471 0.471 0.381 0.500 11 1 0.500 0.381 0.588 0.533 0.500 0.455 0.706 0.588 0.476 0.533 12 1 0.720 0.762 0.737 0.600 0.538 0.476 0.571 0.640 0.250 13 1 0.636 0.700 0.667 0.519 0.455 0.545 0.615 0.235 14 1 0.750 0.588 0.435 0.667 0.667 0.545 0.308 15 1 0.800 0.571 0.625 0.750 0.600 0.364 16 1 0.636 0.471 0.706 0.571 0.333 17 1 0.435 0.522 0.519 0.333 18 1 0.556 0.455 0.615 19 1 0.545 0.308 20 1 0.353 21 1
下载: 导出CSV
-
[1] 龚岚. 北京城区典型城市森林结构特点分析[D]. 北京: 北京林业大学, 2015. GONG Lan. Analysis of Forest Structure Characteristics of Typical Cities in Beijing Urban Area [D]. Beijing: Beijing Forestry University, 2015. [2] FUJITA M, KOIKE F. Landscape effects on ecosystems: birds as active vectors of nutrient transport to fragmented urban forests versus forest-dominated landscapes [J]. Ecosystems, 2009, 12(3): 391 − 400. [3] LUNDBERG J, MOBERG F. Mobile link organisms and ecosystem functioning: implications for ecosystem resilience and management [J]. Ecosystems, 2003, 6(1): 87 − 98. [4] 张征恺, 黄甘霖. 中国城市鸟类学研究进展[J]. 生态学报, 2018, 38(10): 3357 − 3367. ZHANG Zhengkai, HUANG Ganlin. Urban ornithological research in China: a review [J]. Acta Ecol Sin, 2018, 38(10): 3357 − 3367. [5] HOSTETLER M, KNOWLES-YANEZ K. Land use, scale, and bird distributionsin the Phoenix metropolitan area [J]. Landscape Urban Plann, 2003, 62(2): 55 − 68. [6] 谢世林, 曹垒, 逯非, 等. 鸟类对城市化的适应[J]. 生态学报, 2016, 36(21): 6696 − 6707. XIE Shilin, CAO Lei, LU Fei, et al. Adaptation of birds to urbanization [J]. Acta Ecol Sin, 2016, 36(21): 6696 − 6707. [7] NORDT A, KLENKE R. Sleepless in town-drivers of the temporal shift in dawn song in urban European blackbirds[J/OL]. PLoS One, 2013, 8(8): e71476 [2021-04-18]. doi: 10.1371/journal.pone.0071476. [8] WARREN P S, KATTI M, ERMANN M, BRAZEL A. Urban bioacoustics: it’s not just noise [J]. Anim Behav, 2006, 71(3): 491 − 502. [9] 侯建华, 李川, 安哲, 等. 城市园林与夏季鸟类群落结构研究[J]. 河北农业大学学报, 2004, 27(4): 93 − 95. HOU Jianhua, LI Chuan, AN Zhe, et al. The study of urban gardens and bird community structure in summer [J]. J Hebei Agric Univ, 2004, 27(4): 93 − 95. [10] 孙丰硕. 不同城市化水平鸟类群落特征及城区鸟类对生境因子的选择[D]. 北京: 北京林业大学, 2016. SUN Fengshuo. Characteristics of Bird Community under Different Urbanization Levels and Urban Birds’ Selection to Environmental Factors [D]. Beijing: Beijing Forestry University, 2016. [11] FILLOY J, GUSTAVO A Z, BELLOCQ M I. Bird diversity in urban ecosystems: the role of the biome and land use along urbanization gradients [J]. Ecosystems, 2019, 22(1): 213 − 227. [12] SANZ V, CAULA S. Assessing bird assemblages along an urban gradient in a Caribbean Island (Margarita, Venezuela) [J]. Urban Ecosyst, 2015, 18(3): 729 − 746. [13] 黄越. 北京城市绿地鸟类生境规划与营造方法研究[D]. 北京: 清华大学, 2015. HUANG Yue. The Methodology of Bird Habitats’ Making and Planning at Beijing Urban Green Spaces [D]. Beijing: Tsinghua University, 2015. [14] 庄艳美, 孔繁花, 尹海伟, 等. 城市绿地空间格局对鸟类群落影响的研究进展[J]. 南京林业大学学报(自然科学版), 2012, 36(3): 131 − 136. ZHUANG Yanmei, KONG Fanhua, YIN Haiwei, et al. A review on the urban green space pattern affecting avian community [J]. J Nanjing For Univ Nat Sci Ed, 2012, 36(3): 131 − 136. [15] SANTIAGO-ALARCON D, MACGREGOR-FORS I, KÜHNERT K, et al. Avian haemosporidian parasites in an urban forest and their relationship to bird size and abundance [J]. Urban Ecosyst, 2016, 19(1): 331 − 346. [16] 江洪. 围绕“六个”创新大力推动森林城市建设[J]. 安徽林业科技, 2015, 41(3): 3 − 4. JIANG Hong. Strongly promoting forest city construction focusing on 6 innovations [J]. Anhui For Sci Technol, 2015, 41(3): 3 − 4. [17] 张孚允, 杨若莉. 中国鸟类迁徙研究[M]. 北京: 中国林业出版社, 1997. ZHANG Fuyun, YANG Ruoli. Research on Bird Migration in China[M]. Beijing: China Forestry Press, 1997. [18] 王岐山, 胡小龙. 合肥市及其附近地区鸟类调查报告[J]. 安徽大学学报(自然科学版), 1979, 3(2): 60 − 88. WANG Qishan, HU Xiaolong. Investigation Report on Birds in Hefei and Its Adjacent Areas [J]. J Anhui Univ Nat Sci Ed, 1979, 3(2): 60 − 88. [19] 吴海龙, 顾长明. 安徽鸟类图志[M]. 芜湖: 安徽师范大学出版社, 2017. WU Hailong, GU Changming. A Guide to Birds of Anhui[M]. Wuhu: Press of Anhui Normal University, 2017. [20] 詹双侯, 吴翠珍. 合肥地区鸟类调查和多样性分析[J]. 现代农业科技, 2020(17): 201 − 202, 205. ZHAN Shuanghou, WU Cuizhen. Bird survey and diversity analysis in Hefei area [J]. Mod Agric Sci Technol, 2020(17): 201 − 202, 205. [21] 刘常富, 李海梅, 何兴元, 等. 城市森林概念探析[J]. 生态学杂志, 2003, 22(5): 146 − 149. LIU Changfu, LI Haimei, HE Xingyuan, et al. Concept discussion and analysis of urban forest [J]. Chin J Ecol, 2003, 22(5): 146 − 149. [22] ROWANTREE R A. Ecology of the urban forest-introduction to PartⅠ [J]. Urban Ecol, 1984, 8(1/2): 1 − 11. [23] 连军营. 城市森林结构与效益研究[D]. 西安: 西北大学, 2008. LIAN Junying. Study on Urban Forest Structure and Benefits [D]. Xi’an: Northwest University, 2008. [24] 熊春妮, 魏虹, 兰明娟. 重庆市都市区绿地景观的连通性[J]. 生态学报, 2008, 28(5): 2237 − 2244. XIONG Chunni, WEI Hong, LAN Mingjuan. Analysis of connectivity on greenland landscape in metropolitan region of Chongqing City [J]. Acta Ecol Sin, 2008, 28(5): 2237 − 2244. [25] 郑猛, 刘君宜, 刘清平, 等. 咸宁城市鸟类春季群落结构初步研究[J]. 湖北农业科学, 2018, 57(3): 64 − 70. ZHENG Meng, LIU Junyi, LIU Qingping, et al. Preliminary study on the spring avian community structure in Xianning City [J]. Hubei Agric Sci, 2018, 57(3): 64 − 70. [26] 隋金玲. 北京市区绿化隔离带内鸟类群落结构及其影响因素研究[D]. 北京: 北京林业大学, 2005. SUI Jinling. Studies on Avian Community Structures and Affecting Factors in the Greenbelts of Beijing Urban Area[D]. Beijing: Beijing Forestry University, 2005. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210453
 点击查看大图
点击查看大图
计量
- 文章访问数: 1671
- HTML全文浏览量: 478
- PDF下载量: 35
- 被引次数: 0
