







-
树节是树木生长中的正常生理现象,往往存在于树干内部,在用材中统称为节子[1]。节子一般可分为健全节和疏松节,健全节是由活树枝形成,它与树干木质部连接较为紧密,因此基本不影响木材抗压强度和质量,在外观上,其颜色与周围正常的木材相似[2];疏松节是指枝条死亡后被木质部包合起来的部分,这些部分严重破坏了木材内纤维的连续性,导致应力的大量集中,极大地降低木材的强度与抗弯性,影响木材的质量及其加工性能。因此,节子不仅严重影响了木材质量[3-5]和林木产品的利用率[6-7],还会影响树木的材质[8-9]。对节子内部属性和发育规律的研究将有助于了解节子生长过程中的变化规律。有研究表明:节子的大小随着节子着生高度的增加呈先增加后减小的趋势[10-11],并与枝条着生角度成反比[12]。WANG等[13]对不同种植密度下西南桦Betula alnoides的节子特征进行研究发现:高密度种植能有效减小节子基径。HEIN等[14]利用人工修枝与自然整枝条件下的白蜡树Fraxinus excelsior和欧洲槭Acer pseudoplatanus的枝条数据进行分析,发现枝条基径和树干径向生长速率对枝条预测模型的建立起重要作用。陈东升等[15]以落叶松Larix gmelinii人工林为研究对象,建立了落叶松人工林节子模型。贾炜玮等[16]采用混合模型对红松Pinus koraiensis人工林树干内部活节、未包藏死节和包藏死节的体积进行了较好地拟合。然而,中国目前对于节子形成规律及特征的研究主要集中在北方针叶树种,对于南方阔叶树种的研究还较少。米老排Mytilaria laosensis为金缕梅科Hamamelidaceae壳菜果属Mytilaria的常绿阔叶乔木,主要分布于中国云南、广西、广东等省份及东南亚越南、老挝等国,是上述地区的优良速生树种[17],具备材质优良、结构细腻、生长迅速等特点,多用于制造高等家具。但由于其树冠部分枝条较多,自然整枝能力较弱,导致树干上的枯枝无法及时脱落,极大地降低了该树种的质量。若这部分枯枝得不到及时清理,将会逐渐被树干的形成层包裹,形成死节,降低木材可利用率和木材等级[18]。目前,关于米老排的研究主要集中在生物量模型[19]、种苗培育[20]、碳储量[21]、枝条发育规律[22]和凋落物[23]等方面,关于米老排节子分布和发育规律方面研究较为缺乏。据此,本研究通过研究米老排节子时空分布规律,建立节子变色长度预测模型,为米老排无节大径材的培育提供科学依据。
-
中国林业科学研究院热带林业实验中心(22°11′N,106°75′E)地处广西最南部,与越南谅山市接壤,试验区下设哨平、青山、伏波和白云4个分场。米老排人工林位于青山分场,海拔约440 m。该地区属于亚热带季风气候,雨量充沛,夏无酷热,冬无严寒,年平均气温为 22.5 ℃,年平均降水量约1 210 mm,雨季集中在4—10月,约占全年降水量的3/4,全年无霜期约345 d,年平均日照约1 620 h。土壤主要为砖红壤,呈酸性(pH 4.5),土壤有机质质量分数为24.65 g·kg−1,全氮质量分数为1.02 g·kg−1,全磷质量分数为 0.47 g·kg−1,有效氮质量分数为117.20 mg·kg−1,有效磷质量分数为1.20 mg·kg−1,有效钾质量分数为84.20 mg·kg−1。
-
米老排人工林建于2008年,采用来源于同一优良母树的种子培育的实生苗进行造林,初始种植密度为1 667 株·hm−2(株行距2 m×3 m),该样地在培育过程中没有进行间伐和施肥等处理。2019年2月,在该样地中设置了5块20 m×30 m的标准地,对每块样地内的米老排进行编号,并测定每棵树的胸径、树高、枝下高、冠幅等因子。各样地调查数据如表1所示。
表 1 米老排人工林各样地概况
Table 1. Information of different sampled plots of the M. laosensis plantation
样地号 面积/
hm²平均树
高/m平均胸
径/cm平均枝
下高/m平均冠
幅/m坡向 坡度/
(°)M601 0.06 17.8 18.1 5.3 6.8 阳 25 M602 0.06 17.5 17.2 4.9 7.5 半阳 28 M603 0.06 19.4 17.8 4.5 7.9 阳 23 M604 0.06 17.7 18.1 4.5 6.2 半阳 31 M605 0.06 18.4 17.9 4.8 6.4 半阳 22 -
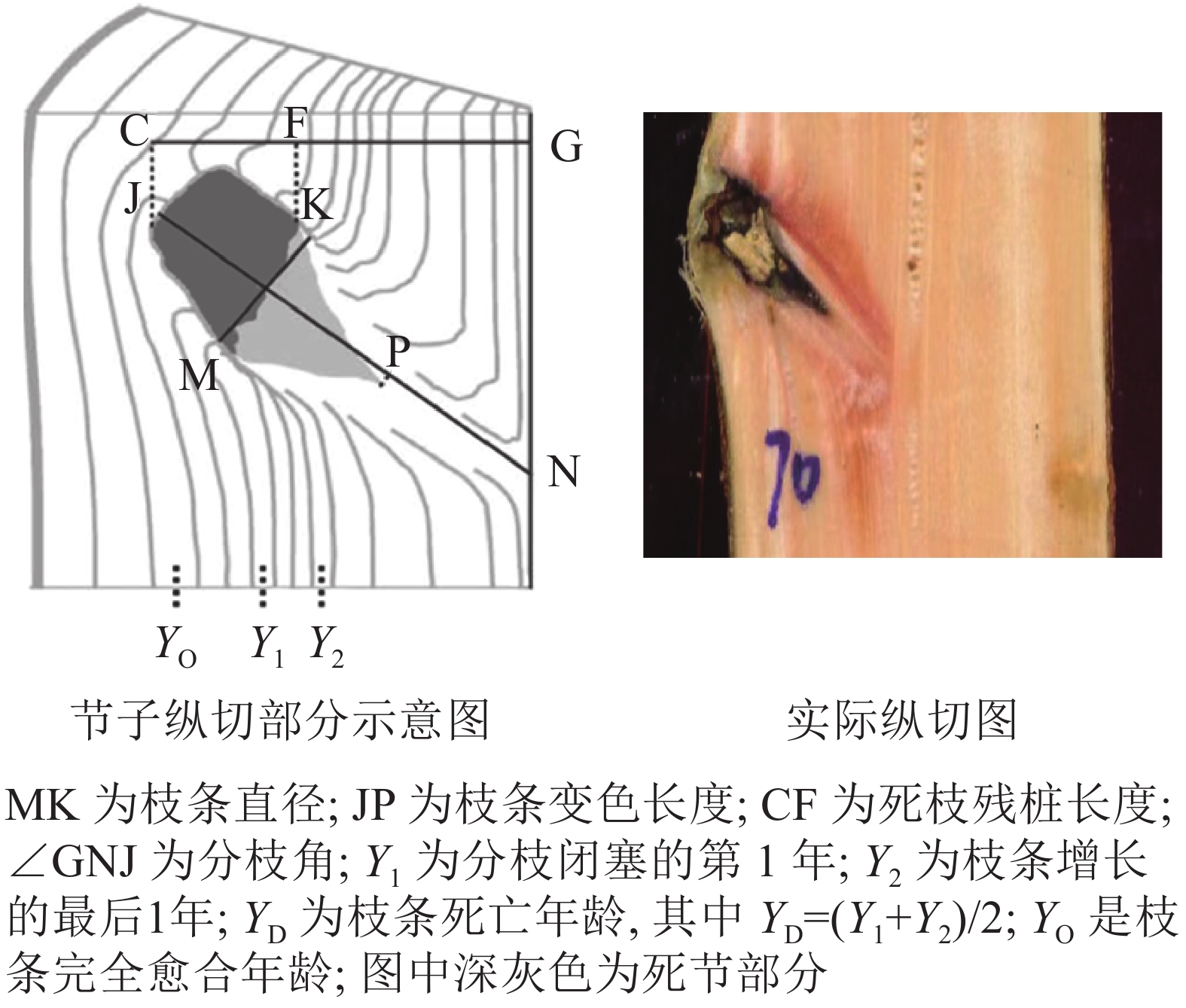
依据每木检尺数据,在每个标准地中分别选出1株优势木、1株亚优势木和2株标准木进行节子解析试验,共计20株。用喷漆标记树干的北方向位置,并用胸径尺测定其胸径后,用油锯将样树伐倒,再次测定其树高、枝下高和冠幅等因子(表2)。以2 m为区分段,用油锯进行切割,分别统计整株节子的数量及其所在高度。随后,将各区分段有节子的部分锯切成 20~30 cm 的圆盘,随后使用油锯沿节子中心部分进行纵切,得到节子剖面(图1),抛光至清晰,最终得到1101个完整的节子。
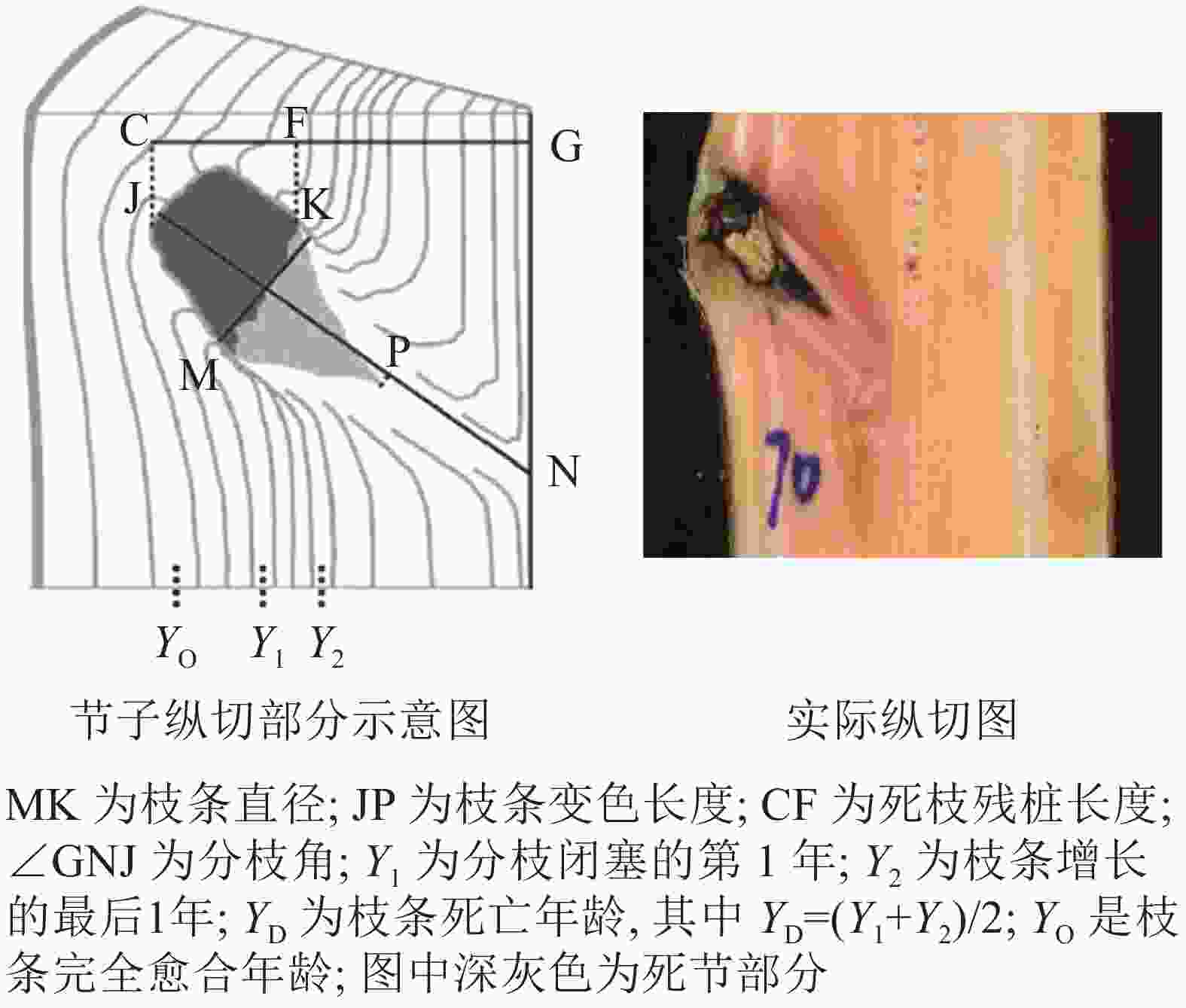
表 2 取样树信息
Table 2. Summary sampled tree statistics
项目 树高/m 胸径/cm 枝下高/m 冠幅/m 数值范围 17.1~22.8 17.7~25.3 4.5~8.8 6.5~10.4 平均值 19.5 21.4 6.9 8.6 标准差 3.9 4.2 1.2 2.3 -
将所测定的节子指标作为自变量(x1~x4),节子变色长度为因变量(y)。进行逐步回归分析,筛选出影响节子变色长度的关键因素。并构建多元线性归回模型:y=a0+a1x1+a2x2+a3x3+a4x4,其中a0、a1、a2、a3、a4为参数。
-
采用SPSS 25.0对数据进行统计分析,使用最小显著性差异法(LSD)进行多重比较,显著性水平为α=0.05,用Excel制表,利用Origin软件制图。
-
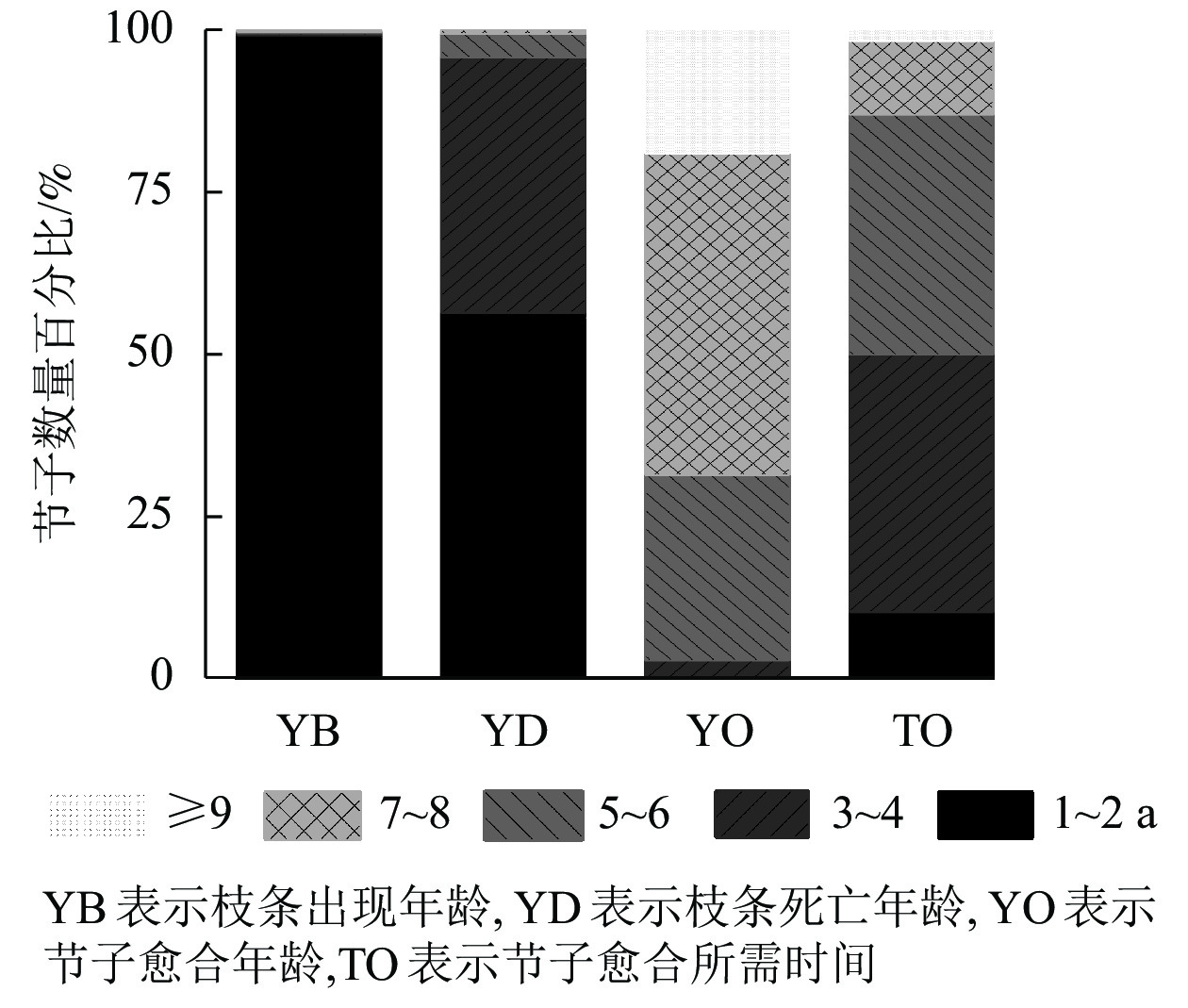
图2显示:米老排大多数枝条在第1~2年开始生长;大量枝条在第3~4年死亡,较少会在第5~8年死亡;第5~8年是节子愈合的高峰期,约占所有节子的87.2%,少部分节子会在第9年痊愈。在枝条死亡脱落后,枝条周围的形成层逐渐将树干内部的枯枝残留包围形成节子,76.7%的节子需要3~6 a的时间才能完全包裹,而能在1~2 a被完全包裹的节子数量较少,仅为10.4%,12.9%的节子需要7 a以上的时间才能完成包裹。
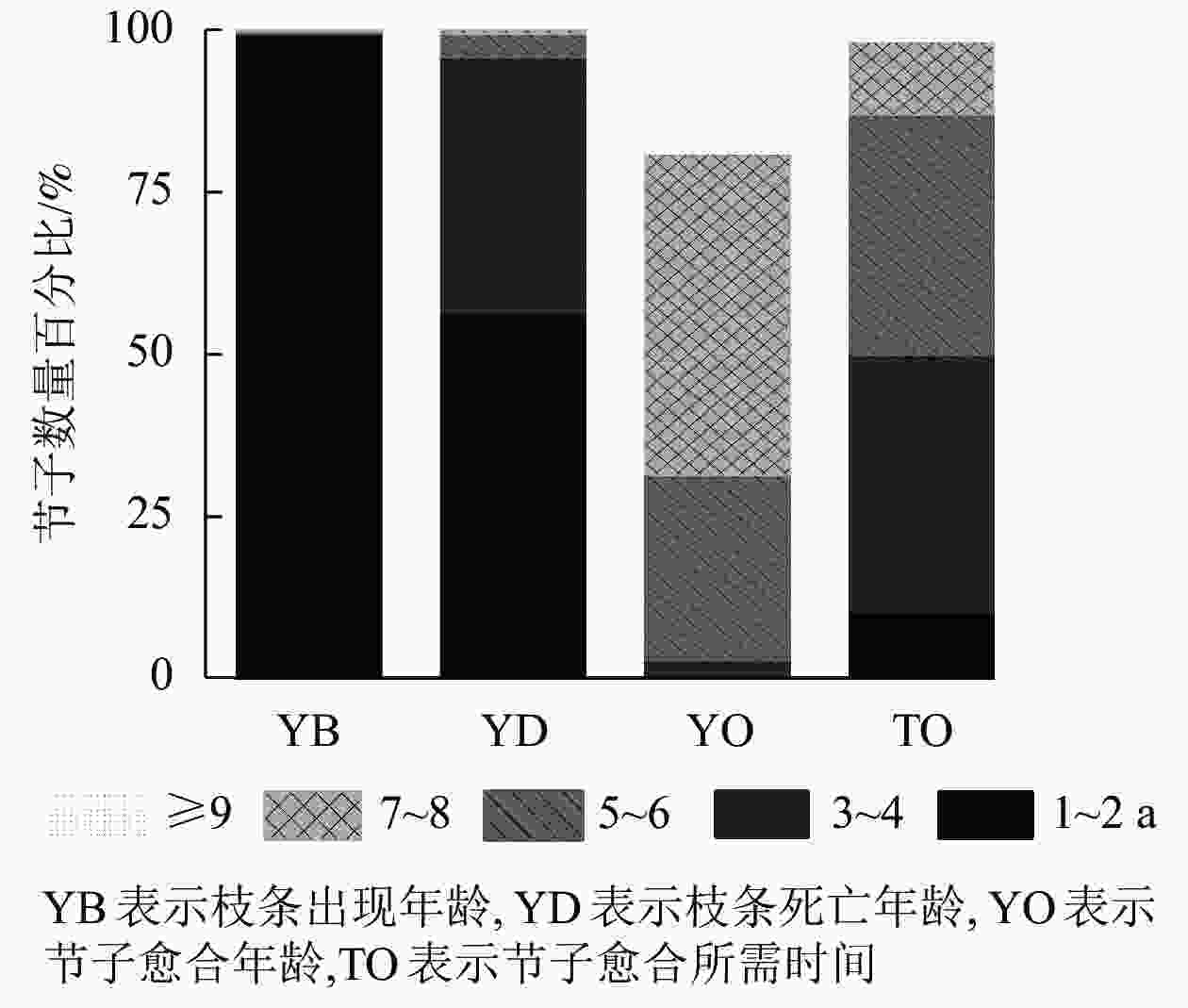
图 2 米老排节子数量的时间分布
Figure 2. Temporal distribution of branch occlusion of M. laosensis
-
如图3所示:在垂直水平上,米老排节子数量随树干高度增加呈先上升后下降的趋势。以2.0 m为区分段,在树干高度为2.0~4.0 m内节子数量百分比最高,约占整株节子总数的17.8%。在2.0~4.0 m之后,随着高度的增加,节子数量开始逐渐减少。从节子数量累计百分比看,61.9%节子集中在0~8.0 m。
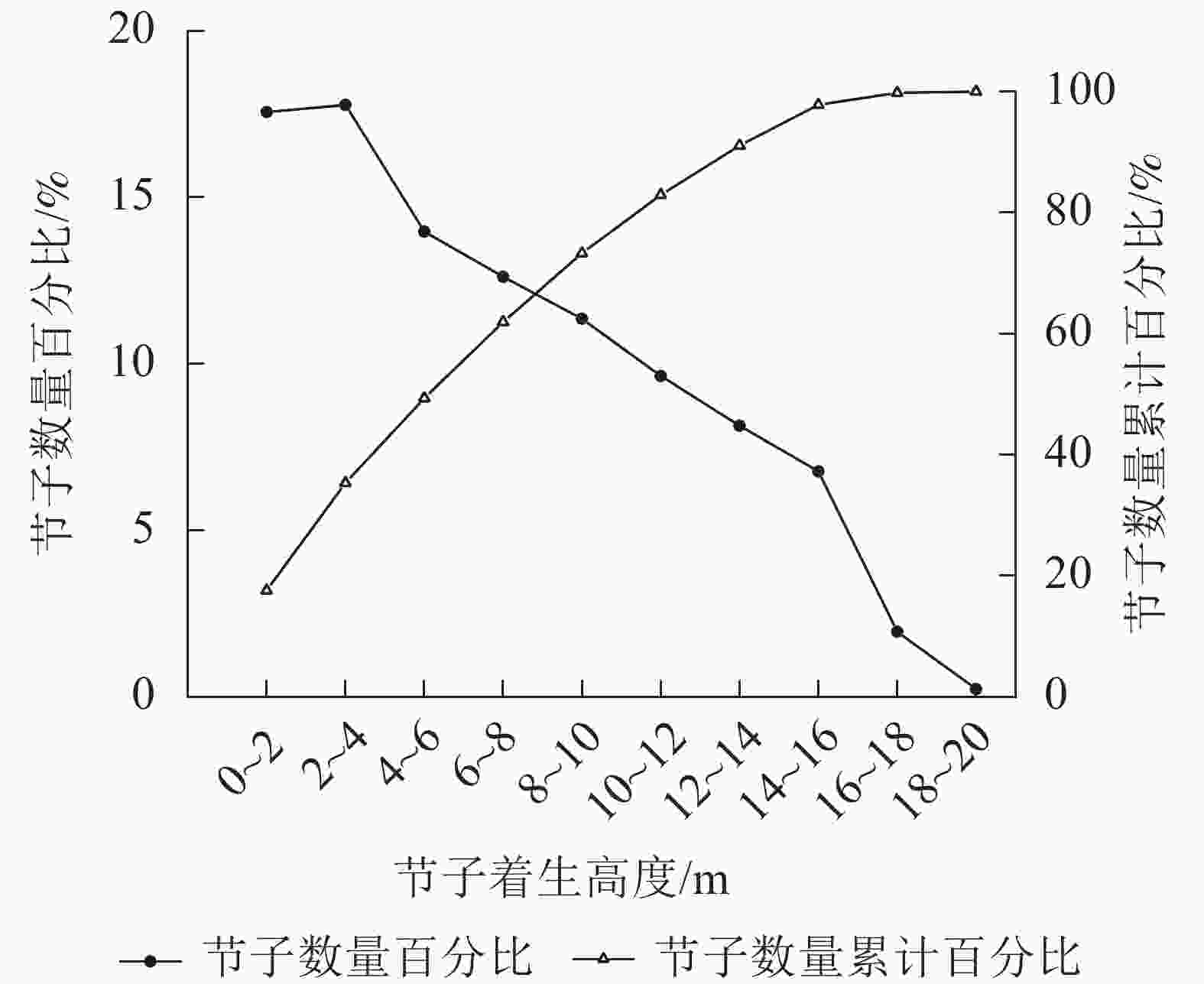
图 3 米老排节子的垂直分布
Figure 3. Vertical distribution of M. laosensis knots
-
由表3可知:节子的平均变色长度达102.5 mm。由图4可知:节子的变色长度与节子直径呈正比。除直径40.0~50.0 mm与直径50.0~60.0 mm间的节子变色长度无显著差异外,其他不同节子直径间变色长度均存在显著差异(P<0.05)。节子直径为0~10.0 mm时节子平均变色长度最小,仅为58.4 mm,而当节子直径为50.0~60.0 mm时,节子平均变色长度最大,为136.0 mm。
表 3 节子信息表
Table 3. Sample branch description for M. laosensis
项目 分枝角度/(°) 枝条死亡
年龄/a节子愈合所
需时间/a节子着生
高度/m节子直径/mm 变色长度/mm 枝条出现
年龄/a节子愈合
年龄/a死节长度/mm 范围 12.9~88.5 2~8 1~10 0.1~18.4 4.7~59.1 17.2~189.6 1~7 3~11 1.4~148.5 平均值 54.1 2.6 4.6 6.8 23.7 102.5 1.1 7.3 51.1 标准差 10.0 0.8 1.6 4.6 8.1 21.1 0.4 1.5 26.1 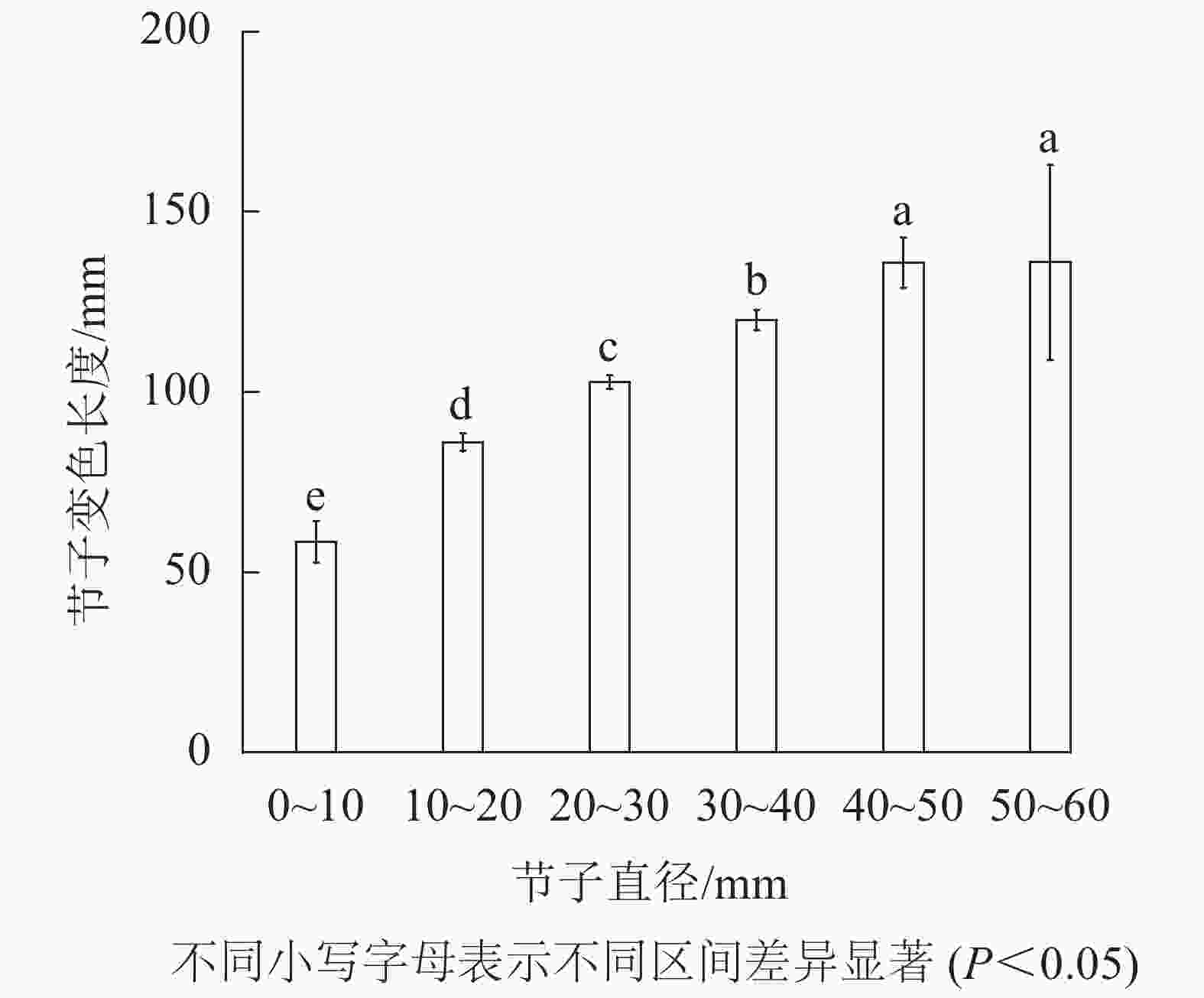
图 4 节子直径对变色长度的影响
Figure 4. Effects of knot diameter on discoloration length
-
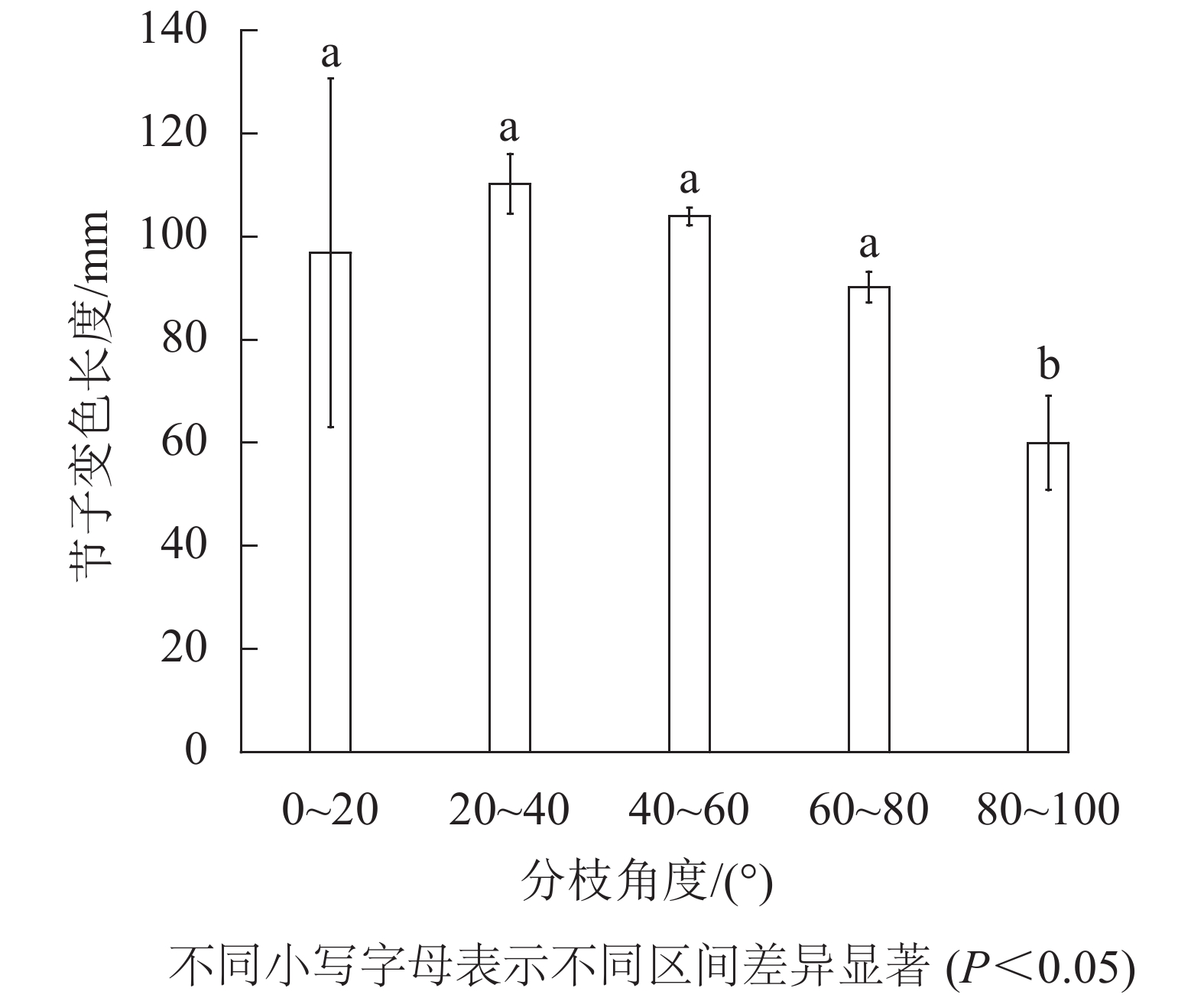
由图5可见:米老排节子变色长度随着分枝角度的增加,呈先增大而后减小的趋势,其中节子分枝角区间在80.0°~100.0°的节子变色长度显著小于其他分支角区间(P<0.05)。当节子的分枝角度在80.0°~100.0°内时,节子平均变色长度最小,仅为60.0 mm;当分枝角度在20.0°~40.0°内时,节子平均变色长度最大,达110.2 mm。
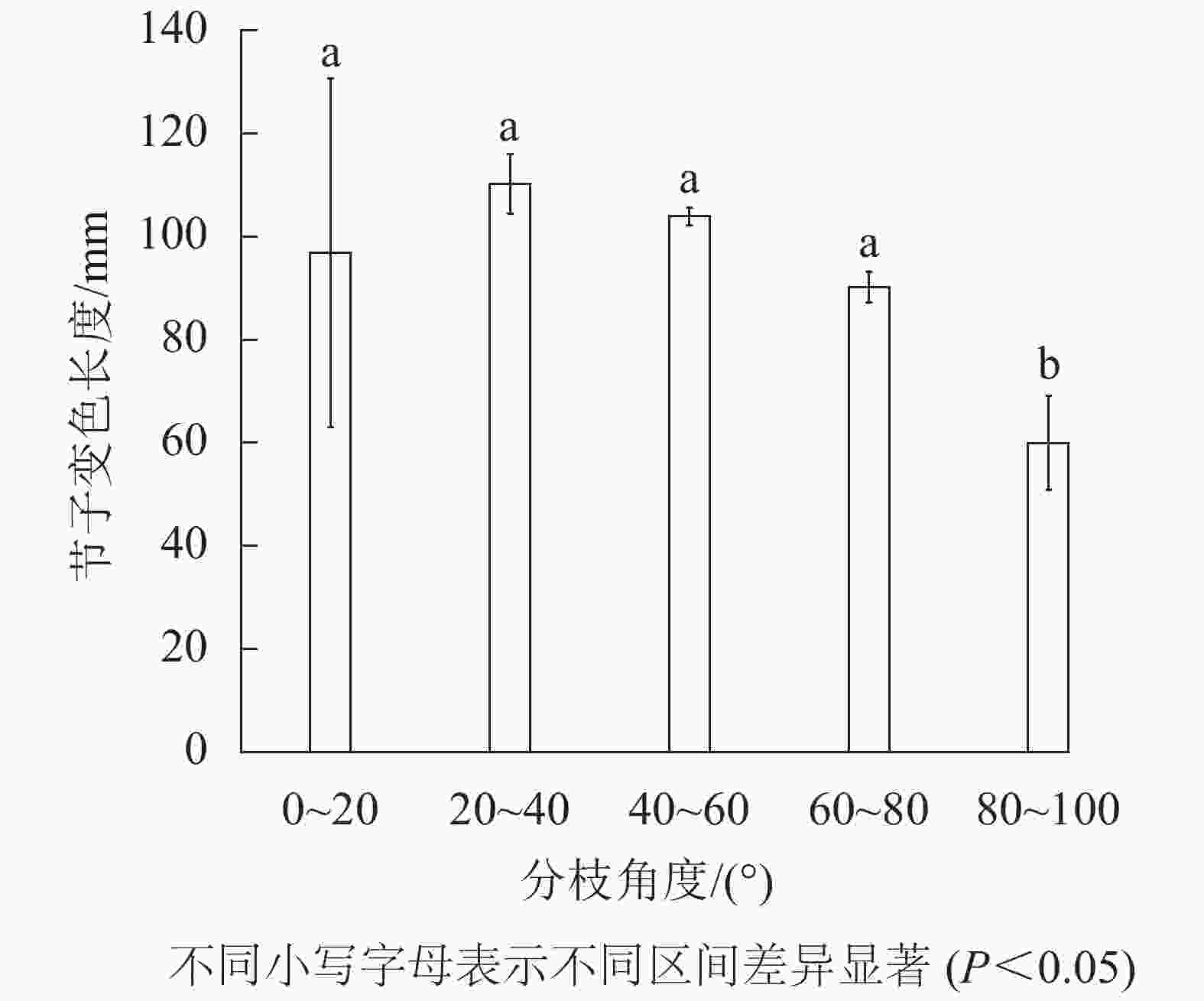
图 5 分枝角度对变色长度的影响
Figure 5. Effects of branch insertion angle on discoloration length
-
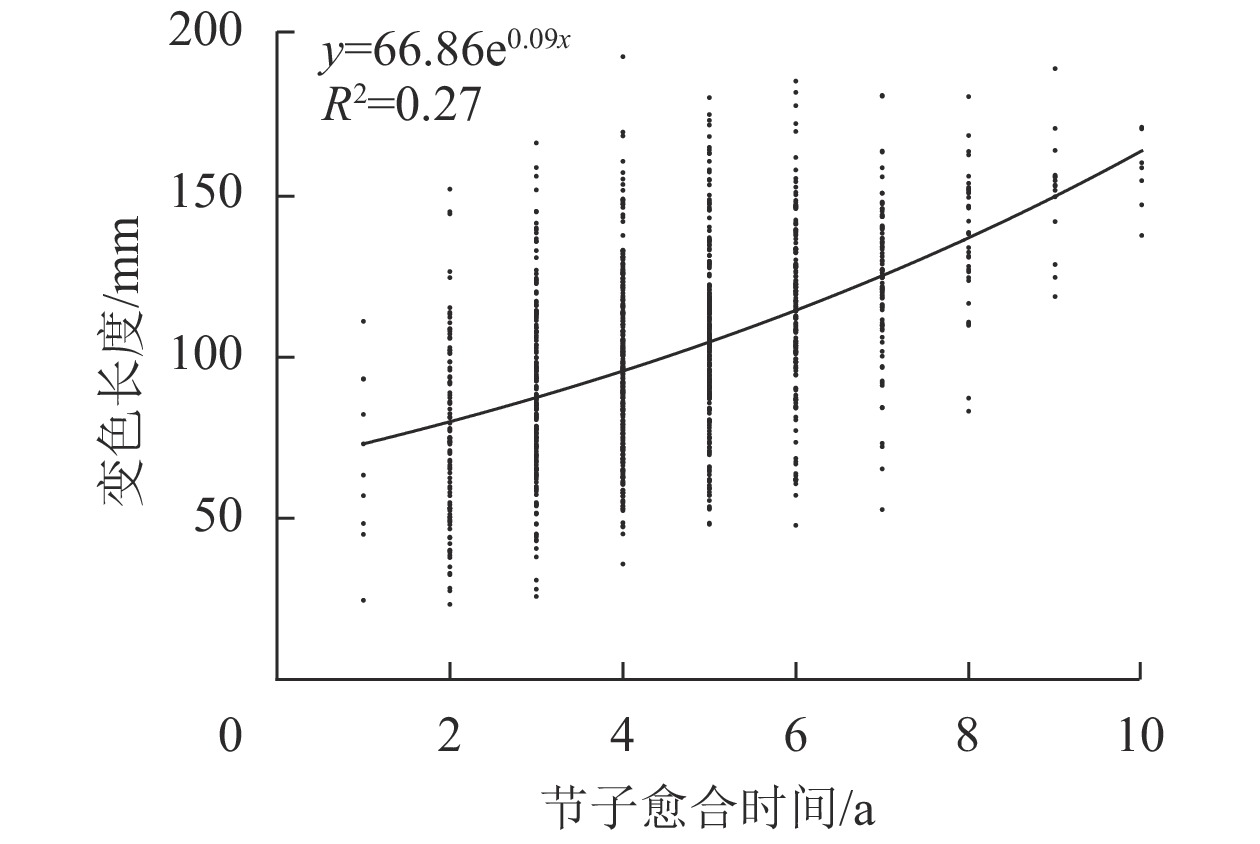
由图6可见:米老排节子变色长度与愈合时间呈正相关,且当节子愈合时间不超过6 a时,变色长度随愈合时间的增加而缓慢增大,而当愈合时间超过6 a,变色长度随愈合时间的增加增速变快。
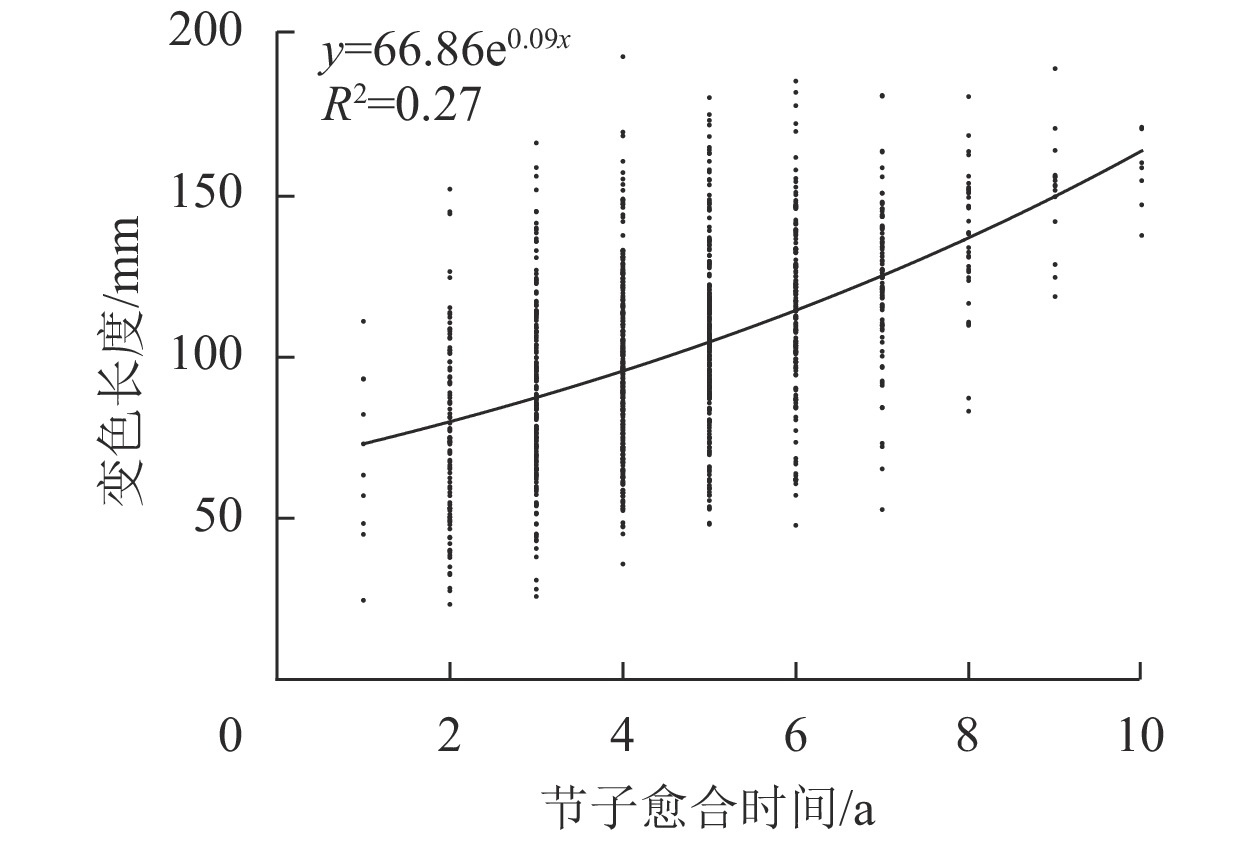
图 6 节子愈合时间对变色长度的影响
Figure 6. Effects of knot occlusion time on discoloration length
-
将枝条出现年龄、枝条死亡年龄、节子愈合时间、分枝角度、死节长度、节子着生高度以及节子直径等7个指标作为自变量,与节子变色长度进行逐步回归分析,得到描述节子变色长度(yDL)的最优模型:yDL=24.388+1.729xKD+0.350xDBL−6.635xYB+5.451xOT。其中:xKD为节子直径,xDBL为死节长度,xYB为枝条出现年龄,xOT为节子愈合时间。模型中各参数均具有统计学意义,模型的R2为0.649(表4)。模型的残差分布均匀,表明方程模拟效果较好(图7)。
表 4 节子变色长度模型参数估计值和拟合统计量
Table 4. Estimated parameter values and fitted statistics values of discoloration length model
参数 估计值 标准差 t P 残差平方和 R² a0 24.388 2.830 8.619 0.057 371 405.423 0.649 a1 1.729 0.064 26.968 0.000 a2 0.350 0.023 14.927 0.000 a3 −6.635 1.485 −4.467 0.000 a4 5.451 0.395 13.805 0.000 说明:a0是常数,a1、a2、a3和a4分别是模型中节子直径、 死节长度、枝条出现年龄和节子愈合时间的参数 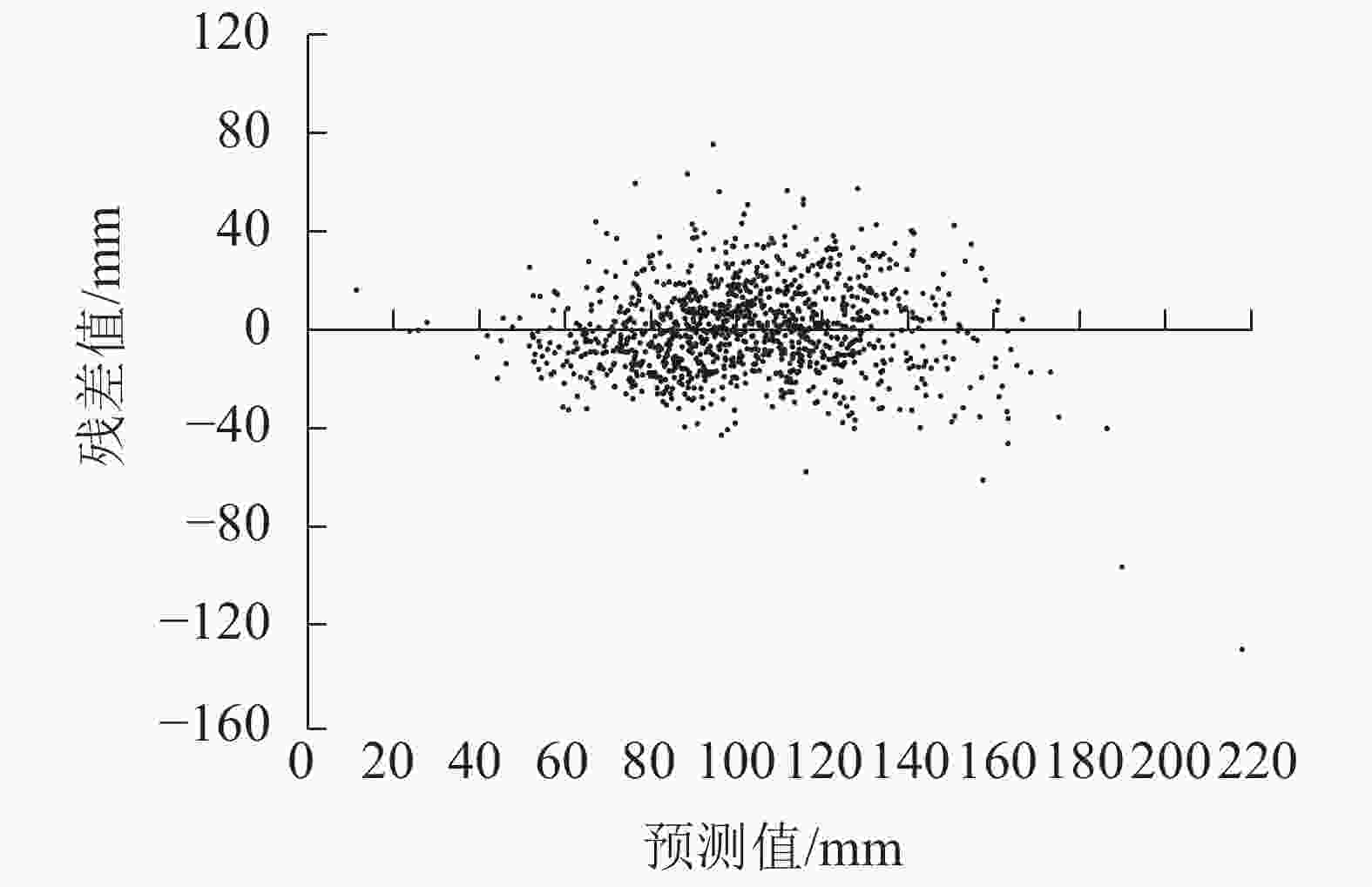
图 7 变色长度预测模型的残差分布
Figure 7. Residual distribution of discoloration length model
-
本研究发现:米老排枝条在第2~4年达到死亡的高峰期,树木上枯枝残留在后期被树干完全包裹形成节子。而水曲柳Fraxinus mandshurica节子形成高峰期在第6~15年[24],格木Erythrophleum fordii在第11~20年是控制节子的关键时期[25]。这些差异主要来源于以下几个原因:一是树种差异,不同树种之间树干径向生长速率不同[13];二是生长立地条件不同,在高密度种植情况下,不同个体之间竞争有限的周围环境资源,会导致个体分配到枝条生长的能量受到影响[26]。本研究将枝条死亡年龄作为米老排修枝的判断依据,在米老排人工林生长第2~4年可对其进行首次修枝。
树干上的枝条死亡后,死枝残桩有时会残留在树干内,阻止树干外部的愈合,延长了枝条愈合所需时间,增大树干内部与外界水分和真菌等接触的时间,从而增加了木材内部受到侵染而发生腐坏的概率[27]。本研究发现:米老排人工林多数节子愈合需要3~6 a,甚至更长的时间。有研究表明:节子愈合时间与节子的直径呈正比[28],与树干径向生长速率呈反比[12],因此,可适当对林木进行间伐或者施肥等措施,以促进树干的径向生长速率,缩短节子愈合所需时间。在米老排垂直分布上,其半数以上节子主要集中在树高8.0 m以下部分,这一结果与楸树Catalpa bungei[29]、水曲柳[24]的研究结果相似,据此可以将8.0 m作为米老排修枝的参考高度。
-
在本研究中:节子变色长度随节子直径的增大而增大,当节子直径不超过50.0 mm时,节子变色长度会随着节子直径的增加而显著增大。关追追等[30]研究发现:当水曲柳节子直径超过15.0 mm时,节子变色长度会随着节子直径的增加而明显增大。而DǍNESCU等[31]认为:欧亚槭Acer pseudoplatanus节子直径超过50.0~60.0 mm时,木材变色率才会明显提高。说明米老排人工林节子在真菌等微生物侵染的情况下更容易发生变色。节子变色长度随分枝角度的增加而减小,且在本研究中当分枝角度大于60.0°时,节子变色长度随分枝角度的增加而显著降低。这与对西南桦[32]的研究结果一致,而与水曲柳[24]的研究结果相反。在本研究中,米老排节子愈合时间超过6 a时,变色长度明显增大。本研究运用逐步回归分析,筛选出节子直径(xKD)、死节长度 (xDBL)、枝条出现年龄(xYB)和节子愈合时间(xOT) 4个关键因子,得到描述节子变色长度(yDL)的最优模型:yDL=24.388+1.729xKD+0.350xDBL−6.635xYB+5.451xOT,相关性达到了极显著水平(P<0.01)。计算结果与实际测量结果之间差异极小,因此可以利用该模型预测米老排枝条形成节子后对木材的影响状况。下一步研究可以对不同年龄、密度和立地条件的林分进行取样分析。
-
米老排人工林在自然整枝条件下,大多数枝条的死亡时间在第2~4年,节子愈合年龄主要集中在第5~8年。节子直径、节子愈合时间、死节长度和枝条出现年龄是决定节子变色长度的关键因子。当节子直径在不超过50.0 mm和愈合时间不超过10 a时,节子变色长度均随直径和年龄的增加显著增加。因此,为了减少节子变色长度,提高米老排木材的质量和出材率,可以在米老排生长的第2~4年,对树干8.0 m以下的米老排人工林进行首次修枝。
Spatial and temporal distribution characteristics and discoloration law of knots in the Mytilaria laosensis plantation
-
摘要:
目的 探明米老排Mytilaria laosensis人工林节子的时空分布特征及变色规律,为培育米老排无节材提供理论参考。 方法 以位于广西凭祥市的中国林业科学研究院热带林业实验中心11年生米老排人工林为研究对象,选取20株米老排样木,利用节子解析的方法,对获得的1 101个完整节子进行研究。 结果 从节子发育年龄上,米老排的枝条主要从第1~2年开始生长,第2~4年是其死亡高峰期(91.9%),这些枝条(87.2%)死亡后在第5~8年内完全愈合并形成节子,其中大部分节子(76.7%)的愈合时间为3~6 a;在第2~4年米老排枝条死亡数量较多,该时段是控制节子形成的关键时期。在节子垂直分布上,节子的数量呈现随树高的增加先上升后急剧下降的趋势,且主要集中在高度为0~8.0 m的树干上。节子变色长度与节子的直径和愈合时间呈显著正相关(P<0.05),但与分枝角度呈显著负相关(P<0.05)。通过逐步回归,得出节子直径、死节长度、节子出现年龄以及愈合时间是影响米老排节子变色长度的关键因子,相关性达极显著水平(P<0.01)。 结论 在米老排种植后的2~4 a,对树干8.0 m以下进行首次修枝,有助于该树种无节材的培育。采取相应的森林培育措施减小枝条基径和缩短节子愈合所需时间,有助于减少节子对木材带来的影响。图7表4参32 Abstract:Objective The objective is to explore the spatial and temporal distribution characteristics and discoloration law of knots in the Mytilaria laosensis plantation, so as to provide theoretical reference for the cultivation of clear wood. Method A total of 1 101 complete knots were obtained by knot analysis from 20 trees of 11-year old M. laosensis plantation in Experimental Center of Tropical Forestry in Pingxiang City of Guangxi. Result In the development age of the knots, the branches of M. laosensis mainly began to grow in the 1st and 2nd year, and the death peak came in the 2nd to 4th year (91.9%). These branches (87.2%) completely healed and formed knots in the 5th to 8th year after death, and the healing time of most of them (76.7%) was 3 to 6 years. More branches of M. laosensis died in the 2nd to 4th year, which was a critical period to control the formation of knots. In the vertical distribution, the number of knots increased first and then decreased sharply with the increase of tree height and they were mainly concentrated on the trunk with a height of 0−8.0 m. The discoloration length of knots was positively correlated with knot diameter and healing time (P<0.05), but negatively correlated with branch angle (P<0.05). Through stepwise regression, it was concluded that knot diameter, dead knot length, knot age and knot healing time were the key factors affecting knot discoloration length, and their correlation was very significant (P<0.01). Conclusion Pruning the trunk below 8.0 m for the first time 2−4 years after planting is beneficial to the cultivation of clear wood. Taking corresponding forest cultivation measures to reduce the base diameter of branches and the time required for knot healing will help ease the impact of knots on wood. [Ch, 7 fig. 4 tab. 32 ref.] -
表 1 米老排人工林各样地概况
Table 1. Information of different sampled plots of the M. laosensis plantation
样地号 面积/
hm²平均树
高/m平均胸
径/cm平均枝
下高/m平均冠
幅/m坡向 坡度/
(°)M601 0.06 17.8 18.1 5.3 6.8 阳 25 M602 0.06 17.5 17.2 4.9 7.5 半阳 28 M603 0.06 19.4 17.8 4.5 7.9 阳 23 M604 0.06 17.7 18.1 4.5 6.2 半阳 31 M605 0.06 18.4 17.9 4.8 6.4 半阳 22  下载: 导出CSV
下载: 导出CSV
表 2 取样树信息
Table 2. Summary sampled tree statistics
项目 树高/m 胸径/cm 枝下高/m 冠幅/m 数值范围 17.1~22.8 17.7~25.3 4.5~8.8 6.5~10.4 平均值 19.5 21.4 6.9 8.6 标准差 3.9 4.2 1.2 2.3
下载: 导出CSV
表 3 节子信息表
Table 3. Sample branch description for M. laosensis
项目 分枝角度/(°) 枝条死亡
年龄/a节子愈合所
需时间/a节子着生
高度/m节子直径/mm 变色长度/mm 枝条出现
年龄/a节子愈合
年龄/a死节长度/mm 范围 12.9~88.5 2~8 1~10 0.1~18.4 4.7~59.1 17.2~189.6 1~7 3~11 1.4~148.5 平均值 54.1 2.6 4.6 6.8 23.7 102.5 1.1 7.3 51.1 标准差 10.0 0.8 1.6 4.6 8.1 21.1 0.4 1.5 26.1
下载: 导出CSV
表 4 节子变色长度模型参数估计值和拟合统计量
Table 4. Estimated parameter values and fitted statistics values of discoloration length model
参数 估计值 标准差 t P 残差平方和 R² a0 24.388 2.830 8.619 0.057 371 405.423 0.649 a1 1.729 0.064 26.968 0.000 a2 0.350 0.023 14.927 0.000 a3 −6.635 1.485 −4.467 0.000 a4 5.451 0.395 13.805 0.000 说明:a0是常数,a1、a2、a3和a4分别是模型中节子直径、 死节长度、枝条出现年龄和节子愈合时间的参数
下载: 导出CSV
-
[1] 李明月, 陈东升, 王雪玉, 等. 节子的性质及其对木材材性的影响[J]. 西北林学院学报, 2020, 35(4): 197 − 204. LI Mingyue, CHEN Dongsheng, WANG Xueyu, et al. Nature of the knot and its impact on wood properties [J]. J Northwest For Univ, 2020, 35(4): 197 − 204. [2] VESTOL G I, HOIBO O A. Prediction of knot diameter in Picea abies (L. ) Karst [J]. Holz Roh Werkst, 2001, 59(1/2): 129 − 136. [3] SHIGO A L. Compartmentalization: a conceptual framework for understanding how trees grow and defend themselves [J]. Annu Rev Phytopathol, 1984, 22(1): 189 − 214. [4] 黄素涌, 王建, 吕建雄, 等. 世界节子研究进展[J]. 林产工业, 2011, 38(5): 3 − 7. HUANG Suyong, WANG Jian, LÜ Jianxiong, et al. Review of overall research on konts [J]. China For Prod Ind, 2011, 38(5): 3 − 7. [5] LOWELL E C, MAGUIRE D A, BRIGGS D G, et al. Effects of silviculture and genetics on branch/knot attributes of coastal pacific northwest Douglas-fir and implications for wood quality: a synthesis [J]. Forests, 2014, 5(7): 1717 − 1736. [6] MACDONALD E, HUBERT J. A review of the effects of silviculture on timber quality of Sitka spruce [J]. Forestry, 2002, 5(2): 107 − 138. [7] BUKSNOWITZ C, HACKSPIEL C, HOFSTETTER K. Knots in trees: strain distribution in a naturally optimized structure [J]. Wood Sci Technol, 2010, 44(3): 389 − 398. [8] MÄKINEN H. Effect of stand density on the branch development of silver birch (Betula pendula Roth) in central Finland [J]. Trees, 2002, 16(4): 346 − 353. [9] ALCORN P J, BAUHUS J, THOMAS D S, et al. Photosynthetic response to green crown pruning in young plantation-grown Eucalyptus pilularis and E. cloeziana [J]. For Ecol Manage, 2008, 255(11): 3827 − 3838. [10] 姜生伟. 落叶松人工林林木动态模型的研究[D]. 哈尔滨: 东北林业大学, 2009. JIANG Shengwei. Studyon Forest Dynamic Model for Larixin Plantation[D]. Harbin: Northeast Forest University, 2009. [11] WEST P W, RATKOWSKY D A, SMITH R. Factors controlling individual branch development during early growth of an experimental plantation of Eucalyptus pilularis in sub-tropical Australia [J]. Trees, 2020, 35: 395 − 405. [12] HEIN S. Knot attributes and occlusion of naturally pruned branches of Fagus sylvatica [J]. For Ecol Manage, 2008, 256(12): 2046 − 2057. [13] WANG Chunsheng, ZHAO Zhigang, HEIN S, et al. Effect of planting density on knot attributes and branch occlusion of Betula alnoides under natural pruning in southern China [J]. Forests, 2015, 6(12): 1343 − 1361. [14] HEIN S, SPIECKER H. Comparative analysis of occluded branch characteristics for Fraxinus excelsior and Acer pseudoplatanus with natural and artificial pruning [J]. Can J For Res, 2007, 37(8): 1414 − 1426. [15] 陈东升, 李凤日, 孙晓梅, 等. 基于线性混合模型的落叶松人工林节子大小预测模型[J]. 林业科学, 2011, 47(11): 121 − 128. CHEN Dongsheng, LI Fengri, SUN Xiaomei, et al. Models to predict knot size for larch plantation using linear mixed model [J]. Sci Silv Sin, 2011, 47(11): 121 − 128. [16] 贾炜玮, 洪燕虎, 李凤日. 人工红松树干内部节子体积预测模型[J]. 应用生态学报, 2020, 31(9): 2943 − 2954. JIA Weiwei, HONG Yanhu, LI Fengri. Prediction models of knot volume inside the stem for Korean pine plantation [J]. Chin J Appl Ecol, 2020, 31(9): 2943 − 2954. [17] 郭文福, 蔡道雄, 贾宏炎, 等. 米老排人工林生长规律的研究[J]. 林业科学研究, 2006, 19(5): 585 − 589. GUO Wenfu, CAI Daoxiong, JIA Hongyan, et al. Growth laws of Mytilaria laosensis plantation [J]. For Res, 2006, 19(5): 585 − 589. [18] KARASZEWSKI Z, BEMBENEK M, MEDERSKI P S, et al. Identifying beech round wood quality-distribution of beech timber qualities and influencing defects on grading [J]. Drewno, 2013, 56(189): 39 − 54. [19] QIN Guoming, LI Rongsheng, ZOU Wentao, et al. Estimating biomass partition in Mytilaria laosensis using additive models [J]. Aust J For Sci, 2020, 137(2): 85 − 108. [20] CHEN Lin, ZENG Jie, JIA Hongyan, et al. Growth and nutrient uptake dynamics of Mytilaria laosensis seedlings under exponential and conventional fertilizations [J]. Soil Sci Plant Nutr, 2012, 58: 618 − 626. [21] 明安刚, 刘世荣, 农友, 等. 南亚热带3种阔叶树种人工幼龄纯林及其混交林碳贮量比较[J]. 生态学报, 2015, 35(1): 180 − 188. MING Angang, LIU Shirong, NONG You, et al. Comparison of carbon storage in juvenile monoculture and mixed plantation stands of three common broadleaved tree species in subtropical China [J]. Acta Ecol Sin, 2015, 35(1): 180 − 188. [22] 王志海, 尹光天, 杨锦昌, 等. 不同造林密度对米老排人工林枝条发育的影响[J]. 林业科学研究, 2019, 32(2): 78 − 86. WANG Zhihai, YIN Guangtian, YANG Jinchang, et al. Effects of planting density on branch development of Mytilaria laosensis plantations [J]. For Res, 2019, 32(2): 78 − 86. [23] 张阳锋, 尹光天, 杨锦昌, 等. 造林密度对米老排凋落物量及动态的影响[J]. 植物研究, 2017, 37(5): 768 − 777. ZHANG Yangfeng, YIN Guangtian, YANG Jinchang, et al. Effects of planting densities on production and dynamics of litter of Mytilaria laosensis plantation bulletin of botanical research [J]. Bull Bot Res, 2017, 37(5): 768 − 777. [24] 关追追, 张彦东. 水曲柳节子时空分布特征与变色规律研究[J]. 北京林业大学学报, 2020, 42(8): 53 − 60. GUAN Zhuizhui, ZHANG Yandong. Spatial and temporal distribution characteristics and discoloration law of Fraxinus mandshurica knot [J]. J Beijing For Univ, 2020, 42(8): 53 − 60. [25] 郝建, 蒙明君, 黄德卫, 等. 格木人工林节子的分布特征及预测模型[J]. 南京林业大学学报(自然科学版), 2017, 41(3): 100 − 104. HAO Jian, MENG Mingjun, HUANG Dewei, et al. Distribution and statistical analysis of knots in Erythrophleum fordii plantations [J]. J Nanjing For Univ Nat Sci Ed, 2017, 41(3): 100 − 104. [26] SANDI M, SANDI W, NICOLESCU V N. Wood discoloration in relation to wound size in northern red oak (Quercus rubra L. ) trees subject to artificial pruning [J]. Span J Rural Dev, 2012, 11(3): 53 − 60. [27] SCHWARZEF F, GRUNER J, SCHUBERT M, et al. Defence reactions and fungal colonisation in Fraxinus excelsior and Tilia platyphyllos after stem wounding [J]. Arbori J, 2007, 30(1): 61 − 82. [28] NICOLESCU V N, SANDI M, PAUN M. Occlusion of pruning wounds on northern red oak (Quercus rubra) trees in Romania [J]. Scand J For Res, 2013, 28(4): 340 − 345. [29] 关追追, 卢奇锋, 何双玉, 等. 楸树树干节子的空间分布特征及其面积预测模型[J]. 中南林业科技大学学报, 2021, 41(10): 20 − 28. GUAN Zhuizhui, LU Qifeng, HE Shuangyu, et al. Spatial distribution characteristics and area prediction model of stem knots in Catalpa bungei [J]. J Cent South Univ For Technol, 2021, 41(10): 20 − 28. [30] 关追追, 冯晨辛, 张彦东. 水曲柳人工林节子愈合与变色特征[J]. 东北林业大学学报, 2021, 49(1): 69 − 73. GUAN Zhuizhui, FENG Chenxin, ZHANG Yandong. Knot occlusion and discoloration of Fraxinus mandshurica plantation [J]. J Northeast For Univ, 2021, 49(1): 69 − 73. [31] DĂNESCU A, EHRING A, BAUHUS J, et al. Modelling discoloration and duration of branch occlusion following green pruning in Acer pseudoplatanus and Fraxinus excelsior [J]. For Ecol Manage, 2015, 335: 87 − 98. [32] WANG Chunsheng, HEIN S, ZHAO Zhigang, et al. Branch occlusion and discoloration of Betula alnoides under artificial and natural pruning [J]. For Ecol Manage, 2016, 375: 200 − 210. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210517
 点击查看大图
点击查看大图
计量
- 文章访问数: 1844
- HTML全文浏览量: 434
- PDF下载量: 32
- 被引次数: 0
