





-
肥料是植物生长发育所必需的矿物质营养来源,直接决定着作物产量。为了获得作物高产,施肥是农业生产的必需措施,其中最普遍的是施加氮肥。氮作为生命元素,作物缺乏后其叶面积减小[1-2]、光合速率降低[2-4]、植株发育受阻[2, 5],最终导致减产。在实际生产中,为了获得作物高产,往往会施加过量的氮肥,这不仅增加了生产成本,同时也会污染生态环境,尤其是水体环境[6-8]。研究表明:中国肥料的使用量持续增长,而氮肥的平均利用率却只有32%~45%,即使在有效的农田管理下也不超过50%[9]。由此可见,合理施肥对降低农业投入和保护生态环境均具有重要意义。毛竹Phyllostachys edulis是中国分布面积最大、范围最广、开发利用程度最高,对竹产区地方经济和竹农收入影响最为深远的集经济、生态和社会效益于一体的笋材两用竹种。施肥是集约经营毛竹林提高经济效益的重要手段之一。施肥后毛竹出笋提前,出笋期延长[10],出笋数和活笋数提高[11],同时胸径和林分密度增加[12]。在毛竹展叶期到绿叶期进行施肥处理,其中展叶期施肥更有利于提高新竹的光合能力,促进新竹生长[13]。宋艳冬等[14]通过研究5月和8月施肥对毛竹叶片光合色素含量以及光合速率的影响表明,5月施肥效果较好,然而8月施肥能延长叶片的光合能力,延缓叶片衰老。目前,关于毛竹林施肥的研究主要集中于提高毛竹林生产力以及施肥时期的确定,然而在毛竹林经营过程中如何进行合理施肥尚未见研究报道。因此,为了确定毛竹林准确的氮素施肥量以促进其集约经营,本研究通过长期调查氮素施肥对竹笋和竹材产量的影响以及氮素施肥对毛竹光合作用的影响,以期从实践和理论角度对毛竹林的合理施肥予以揭示。
-
试验地位于浙江省遂昌县妙高镇,39°35′51.75″N,116°35′19.51″E。所用试验毛竹林为笋用林,坡向南坡,坡度22°~25°,土壤为山地红壤,土层厚度60 cm以上。土壤pH 5.3~6.0,有机质为4.10 g·kg-1,全氮、速效磷、速效钾分别为0.151 g·kg-1,5.40 mg·kg-1,80.20 mg·kg-1。竹林均采用中度经营,立竹密度为2 400~2 700株·hm-2,年龄结构1度:2度:3度竹为1:1:1,立竹分布、大小均匀,平均胸径为9~10 cm。
-
选择立地条件相近的毛竹笋用林设置试验样地,每块样地面积为20 m×30 m。试验于2004-2011年进行,共设置4个氮素施肥水平,分别为对照(不施氮素, ck), N1(施氮素量100 kg·hm-2),N2(施氮素量250 kg·hm-2)和N3(施氮素量400 kg·hm-2),设样地3块·处理-1,共计样地12块,完全随机排列。施肥样地(包含对照)配施五氧化二磷(P2O5)125 kg·hm-2和氧化钾(K2O)40 kg·hm-2。肥料品种:氮肥为尿素(含氮46%),磷肥为过磷酸钙(含P2O512.1%),钾肥为氧化钾(含K2O 63%),施肥时间为每年的5月上旬,施肥方法采用竹蔸施肥法。
2006-2011年连续3个生长周期内,在2006年、2008年和2010年11月至翌年2月记录冬笋产量;在2007年、2009年和2011年3-4月记录春笋产量,7月下旬测量新生竹胸径,10月上旬测定竹材产量。2011年7月下旬-8月上旬,在每块样地选取生长良好、高度(12~15 m)及胸径(9~10 cm)一致的1年生和3年生毛竹各3株作为测试样竹,取其阳面枝条顶端成熟、健康的叶片3片测量其光合色素质量分数和光合性能,测定9次·样地-1,取其平均值作为该样地的测量值。
-
新生毛竹的胸径采用围径尺进行测量,冬笋、春笋和竹材产量采用称量法测量。
-
毛竹叶片除去主脉后剪碎并混匀,称取0.1 g置于具塞试管中,加入体积分数为80%的丙酮10 mL,置于黑暗处室温下萃取24 h后(样品完全变白),分别测定663,645和470 nm处的吸光度D(λ)值,按Lichtenthaler等[15]修订的Arnon法计算叶绿素a,叶绿素b和类胡萝卜素质量分数。m叶绿素a=12.72D663-2.59D645, m叶绿素b=12.88D645-4.67D663, m类胡罗卜素=(10 000 D470-3.27 m叶绿素a-104 m叶绿素b)229。
-
采用LI-6400便携式光合仪(美国Li-COR公司)测定毛竹的光响应曲线,光强设为0,20,50,100,150,200,250,300,400,600,800,1 000,1 200,1 400,1 600,1 800和2 000 μmol·m-2·s-1,数据采集时间为200 s,并计算光补偿点、光饱和点和最大光合速率。
-
试验数据采用SPSS 13.0进行Duncan检验,采用Origin 8.0进行绘图。
-
3 a的调查结果表明:氮素施肥能极显著提高毛竹冬笋和春笋产量。在N1, N2和N3施肥水平下,冬笋3 a产量的平均值分别比对照提高了3.7倍(P<0.01),6.8倍(P<0.01)和7.1倍(P<0.01);春笋3 a产量的平均值分别比对照提高了1.5倍(P<0.01),2.0倍(P<0.01)和1.9倍(P<0.01),其中N2和N3施肥水平对冬笋和春笋产量的影响没有明显差异(表 1, 表 2)。
表 1 不同氮素施肥水平条件下毛竹林冬笋产量
Table 1. Yield of winter shoots at different nitrogen fertilization levels
处理 冬笋产量/ (kg·hm-2) 2006年 2008年 2010年 平均值 ck 397.5±55.8Cc 468.0±47.1Cd 526.5±63.8Cc 464.0±64.6Cc N1 2 305.5±87.3Bb 2 104.5±88.1Bc 2 182.5±73.9Bb 2 197.5±101.3Bb N2 3 672.0±99.4Aa 3 451.5±156.2Ab 3 681.0±78.2Aa 3 601.5±130.0Aa N3 3 723.0±156.9Aa 3 949.5±210.3Aa 3 547.5±96.8Aa 3 740.0±201.5Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 表 2 不同氮素施肥水平条件下毛竹林春笋产量
Table 2. Yield of spring shoots at different nitrogen fertilization levels
处理 春笋产量/ (kg·hm-2) 2007年 2009年 2011年 平均值 ck 2 959.5±82.1 Cc 3 159.0±102.3Cc 3 046.5±79.8Cc 3 055.0±100.0Cc N1 7 657.3±254.1 Bb 7 448.7±213.8Bb 8 001.2±195.2Bb 7 702.4±279.0 Bb N2 8 980.5±246.2 Aa 9 094.5±301.5Aa 9 648.0±257.8 Aa 9 241.0±357.1 Aa N3 8 605.5±269.4 Aa 9 463.5±309.4 Aa 8 953.0±400.2Aa 9 007.3±431.6Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 -
2007年、2009年和2011年7月下旬对试验样地保留新竹进行调查表明,氮素施肥后新竹胸径显著增加。在N1,N2和N3施肥水平下,3 a调查结果的胸径平均值分别比对照增加了9.5%(P<0.05),33.3%(P<0.01)和34.5%(P<0.01),其中N2和N3施肥水平间没有明显差异(表 3)。
表 3 不同氮素施肥水平条件下毛竹林新竹胸径
Table 3. Diameter at breast height in new Phllostachys edulis at different nitrogen fertilization levels
处理 胸径/cm 2007年 2009年 2011年 平均值 ck 8.71±1.32 Bb 8.16±1.61 Bb 8.43±1.31 Bc 8.43±1.51 Bc N1 9.23±1.12 Bb 8.83±1.34 Bb 9.52±1.13 Bb 9.23±1.21 Bb N2 11.23±0.92 Aa 10.76±1.14 Aa 11.63±1.22 Aa 11.24±1.23 Aa N3 11.55±1.02 Aa 11.11±1.32 Aa 11.47±1.41 Aa 11.34±1.43 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 -
2007年、2009年和2011年的调查结果均表明,氮素施肥能显著提高毛竹的竹材产量。在N1,N2和N3施肥水平下,竹材3 a的平均产量分别比对照增加了0.7倍(P<0.01),1.1倍(P<0.01)和1.2倍(P<0.01),其中后2种施肥水平间无显著差异(表 4)。
表 4 不同氮素施肥水平条件下毛竹林竹材产量
Table 4. Yield of bamboo timber at different nitrogen fertilization levels
处理 竹材产量/ (kg·hm-2) 2007年 2009年 2011年 平均值 ck 11 700±602 Cc 10 200±719 Cd 10 800±873 Cc 10 900±755 Cc N1 18 563±568 Bb 17 738±610 Bc 18 975±710 Bb 18 425±630 Bb N2 22 688±1 115 Aa 21 450±1 007 Ab 23 513±1 074 Aa 22 550±1 038 Aa N3 23 513±416 Aa 24 383±429 Aa 23 925±402 Aa 23 940±413 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 -
氮素施肥后,1年生和3年生毛竹光合色素质量分数均显著增加,而叶绿素a/b则降低。在N2施肥水平下,光合色素增加最多,其中1年生毛竹叶绿素a,叶绿素b,叶绿素(a+b)和类胡萝卜素质量分数分别比对照增加了19.6%(P<0.01),27.7%(P<0.01),21.7%(P<0.01)和12.9%(P<0.05);3年生毛竹分别比对照增加了14.0%(P<0.05),29.7%(P<0.01),18.2%(P<0.01)和9.3%(P<0.05)(表 5)。
表 5 不同氮素施肥水平条件下毛竹叶片光合色素质量分数
Table 5. Photosynthetie pigment content in leaves of Ph. edulis at different nitrogen fertilization levels
竹龄 处理 叶绿素a/(mg·g-1) 叶绿素b/(mg·g-1) 叶绿素总量/(mg·g-1) 叶绿素a/b 类胡萝卜素/(mg·g-1) ck 2.91±0.10 Bc 1.01±0.09 Bc 3.92±0.15 Bc 2.91±0.29 Aa 0.85±0.07 Ab N1 3.24±0.29 Ab 1.13±0.26 ABb 4.37±0.53 ABb 3.03±0.68 Aa 0.87±0.03 Ab 1年生 N2 3.48±0.17 Aa 1.29±0.09 Aa 4.77±0.24 Aa 2.68±0.15 Ab 0.96±0.06 Aa N3 3.34±0.22 Aab 1.30±0.09 Aa 4.64±0.30 Aab 2.57±0.07 Ab 0.81±0.03 Ab ck 3.01±0.33 Abc 1.01±0.11 Bc 4.01±0.60 Bc 3.02±0.56 Aa 0.86±0.20 Ab N1 3.20±0.30 Ab 1.12±0.11 ABab 4.32±0.68 ABab 2.86±0.21 Aab 0.87±0.10 Ab 3年生 N2 3.43±0.27 Aa 1.31±0.12 Aa 4.74±0.38 Aa 2.62±0.05 Ab 0.94±0.11 Aa N3 3.11±0.24 Abc 1.20±0.07 Aab 4.31±0.38 ABab 2.59±0.14 Ab 0.86±0.11 Ab 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 -
氮素施肥后,1年生和3年生毛竹叶片的净光合速率明显高于对照,并且在N2施肥水平下净光合速率最高(图 1)。

图 1 不同氮素施肥水平条件下毛竹叶片净光合速率
Figure 1. Net photosynthetie rate in leaves of Ph. edulis at different nitrogen fertilization levels
在N1,N2和N3施肥水平下,1年生和3年生毛竹的光补偿点明显降低,而光饱和点和最大光合速率则显著明显。在N2施肥水平下,各指标的变化最为明显,其中1年生毛竹的光补偿点降低了49.5%(P<0.01),光饱和点和最大光合速率则分别增加了5.1%(P<0.05)和11.6%(P<0.05);3年生毛竹的光补偿点降低了20.4%(P<0.01),光饱和点和最大光合速率分别增加了9.0%(P<0.05)和14.3%(P<0.05)(表 6)。
表 6 不同氮素施肥水平条件下毛竹叶片光合参数
Table 6. Photosynthesis parameters in leaves of Ph. edulis at different nitrogen fertilization levels
竹龄 处理 光补偿点/ (μmol · m-2· s-1) 光饱和点/ (μmol · m-2· s-1) 最大光合速率/ (μmol · m-2· s-1) ck 53.3±3.2 Aa 580.3±10.7 Ab 17.3±1.1 Ab N1 46.2±3.1 Ab 590.9±7.8 Aab 18.6±1.3 Aab 1年生 N2 26.9±1.3 Cd 610.0±8.9 Aa 19.3±1.2 Aa N3 34.3±2.7 Bc 597.4±10.1 Aab 18.5±0.9 Aab ck 44.2±2.3 Aa 590.1±9.7 Abc 15.4±0.5 Ab 3年生 N1 37.2±1.9 ABb 621.0±10.3 Aa 16.3±0.2 Aab N2 35.2±3.1 Bb 643.5±9.3 Aa 17.6±0.5 Aa N3 41.2±3.9 ABa 616.4±11.5 Aab 17.2±0.6 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。 -
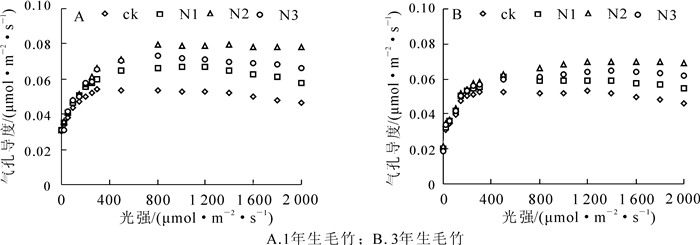
对1年生和3年生毛竹而言,不同氮素施肥水平均能使其叶片气孔导度明显增加,其中在N2施肥水平下气孔导度增至最大;当施氮素增至N3水平时,气孔导度有所降低,但是仍明显高于对照(图 2)。

图 2 不同氮素施肥水平条件下毛竹叶片气孔导度
Figure 2. Cond in leaves of Ph. edulis at different nitrogen fertilization levels
-
氮素施肥后,1年生和3年生毛竹叶片的胞间二氧化碳摩尔分数均明显降低。随着施肥量增加,毛竹叶片胞间二氧化碳摩尔分数逐渐降低,当施肥量在N2水平时胞间二氧化碳摩尔分数最低。当施肥量继续增加,胞间二氧化碳摩尔分数又有所升高,但仍明显低于对照(图 3)。

图 3 不同氮素施肥水平条件下毛竹叶片胞间二氧化碳摩尔分数
Figure 3. CO2 concentration in leaves of Ph. edulis at different nitrogen fertilization levels
-
施肥是毛竹林集约经营,提高经济效益的重要手段之一,对毛竹进行深翻再加施稀土复合肥后,其出笋提前6 d,出笋期延长10 d,出笋总量增加2.2倍[10]。李睿等[11]研究亦有相似发现,毛竹林施肥可使其出笋数和活笋数提高3倍以上。除笋产量提高外,施肥还能使毛竹胸径和林分密度明显增加,同时林分水平的长期生物产量增长[12]。本研究亦表明,氮素施肥处理能明显提高竹笋和竹材产量,促进毛竹胸径生长,并且250 kg·hm-2与400 kg·hm-2施肥水平之间的促进效果无显著性差异。
对1年生和3年生毛竹的测定结果表明:施肥能明显促进其光合色素合成,从而使其质量分数增加,其中以250 kg·hm-2的施肥效果最好(表 5)。这与金晓春等[13]和宋艳冬等[14]施肥处理后毛竹光合色素含量明显升高的研究结果相一致。氮是叶绿素卟啉环与镁离子结合的关键元素,因此施加氮肥有利于植物叶绿素合成,从而使其含量提高。Anderson等[16]研究表明:叶绿素b含量增加对叶绿体基粒以及基粒片层的增加具有重要作用,从而有利于叶片光能捕获与传递能力的提高。叶绿素a/b与叶绿素a的状态有关,其比值升高对光合作用具有不利影响[17]。施氮肥处理能不同程度地降低毛竹叶片叶绿素a/b,其中以250 kg·hm-2施肥水平处理后降低量最大,这可能是此施肥水平下毛竹光合能力较高的重要原因。
施加氮肥后1年生和3年生毛竹叶片的光合能力明显增强,然而随着氮素施肥量增加,光合能力先增强后降低。这与适宜施肥后桑树Morus alba光合能力提高而过量施肥则降低的研究结果相类似,其原因可能与过量施肥对植物所造成的胁迫有关[18]。氮素影响植物叶片的光合作用主要有气孔因素和非气孔因素,其中气孔因素是指氮素通过影响气孔开度而影响光合作用,非气孔因素是指氮素作为光合机构的构架元素而影响光合过程。Heithold等[19]研究表明,气孔因素是影响高氮小麦Triticum aestivum光合作用的主要原因,并且这种影响明显高于低氮小麦。在缺氮条件下,植物叶片的1, 5-二磷酸核酮糖羧化/加氧酶(Rubisco)含量和活性明显降低,因此光合能力减弱[20]。增加氮素供应后,植物类囊体膜蛋白的磷酸化和Rubisco活性显著增强,因此光合速率提高[21]。施加氮肥后,毛竹叶片的气孔导度和叶绿素质量分数明显增加,而胞间二氧化碳摩尔分数降低则表明Rubisco对二氧化碳的同化能力增强,因此,施氮肥后毛竹叶片光合能力明显提高与气孔和非气孔因素均有关。
光合作用是植物体内最基本的代谢过程,为植物生长发育提供必需的能量和营养物质。氮素施肥后毛竹的光合能力明显提高,从而有利于光合产物积累,进而促进竹笋和竹材产量增长。因此,综合竹笋与竹材产量与光合性能指标分析,毛竹林氮素施肥水平以250 kg·hm-2为宜。
Productivity and photosynthetic ability of Phyllostachys edulis with nitrogen fertilization
-
摘要: 竹林施肥是集约经营毛竹Phyllostachys edulis林提高经济效益的重要手段之一。为了确定毛竹林氮素的合理施肥量, 从2006-2011年持续调查了氮素施肥对毛竹竹笋和竹材产量的影响, 在2011年测定了施肥对毛竹光合色素和光合作用的影响。结果表明:100, 250和400 kg·hm-2施肥水平均能显著促进毛竹冬笋、春笋和竹材产量提高以及新竹胸径增大, 其中在250 kg·hm-2施肥水平时, 各平均值分别比对照提高了6.8倍(P < 0.01), 2.0倍(P < 0.01), 1.1倍(P < 0.01)和33.3%(P < 0.01), 但是250和400 kg·hm-2施肥水平之间无明显差异。氮素施肥后, 1年生和3年生毛竹叶片的光合色素质量分数、光饱和点、净光合速率均明显提高, 而光补偿点和胞间二氧化碳摩尔分数则明显降低。这表明植株的光合能力明显提高, 其中以250 kg·hm-2施肥效果最明显。综合分析试验结果, 毛竹林施肥量以250 kg·hm-2为宜。Abstract: Fertilization is an important intensive operation for improving economic benefits of a Phyllostachys edulis stand. To determine the amount of N fertilizer needed from practical and theoretical aspects, the effects of N fertilization (at 100, 250, and 400 kg·hm-2) on the production of winter shoots, spring shoots, bamboo stems, and diameter at breast height (DBH) of a Ph. edulis stand from 2006-2011 were measured using complete randomized design (CRD). The Ph. edulis stand without addition of N fertilizer was the control. Each treatment and the control had 3 replicates, and Duncan test was used to analyze the difference. The photosynthetic pigment content and photosynthesis of bamboo leaves were also calculated. Results showed that 250 kg·hm-2 N fertilization significantly increased (P < 0.01) new bamboo production of winter shoots, 6.8 fold; spring shoots, 2.0 fold; bamboo stems, 1.1 fold; and DBH 33.3%. However, no significant differences were found for the 100 and 400 kg·hm-2 treatments. After fertilization, the photosynthetic pigment content, light saturation point, and net photosynthetic rate increased; whereas, the light compensation point and intercellular CO2 concentrations (Ci) decreased, indicating that N fertilization raised photosynthetic ability. Among the three fertilization level, 250 kg·hm-2 had the best effect on promoting Ph. edulis photosynthesis. Therefore, 250 kg·hm-2 was the best fertilization rate for Ph. edulis stands.
-
Key words:
- botany /
- Phyllostachys edulis /
- nitrogen fertilization /
- productivity /
- photosynthetic pigment /
- photosynthetic ability
-
图 1 不同氮素施肥水平条件下毛竹叶片净光合速率
Figure 1 Net photosynthetie rate in leaves of Ph. edulis at different nitrogen fertilization levels
图 2 不同氮素施肥水平条件下毛竹叶片气孔导度
Figure 2 Cond in leaves of Ph. edulis at different nitrogen fertilization levels
图 3 不同氮素施肥水平条件下毛竹叶片胞间二氧化碳摩尔分数
Figure 3 CO2 concentration in leaves of Ph. edulis at different nitrogen fertilization levels
表 1 不同氮素施肥水平条件下毛竹林冬笋产量
Table 1. Yield of winter shoots at different nitrogen fertilization levels
处理 冬笋产量/ (kg·hm-2) 2006年 2008年 2010年 平均值 ck 397.5±55.8Cc 468.0±47.1Cd 526.5±63.8Cc 464.0±64.6Cc N1 2 305.5±87.3Bb 2 104.5±88.1Bc 2 182.5±73.9Bb 2 197.5±101.3Bb N2 3 672.0±99.4Aa 3 451.5±156.2Ab 3 681.0±78.2Aa 3 601.5±130.0Aa N3 3 723.0±156.9Aa 3 949.5±210.3Aa 3 547.5±96.8Aa 3 740.0±201.5Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。  下载: 导出CSV
下载: 导出CSV
表 2 不同氮素施肥水平条件下毛竹林春笋产量
Table 2. Yield of spring shoots at different nitrogen fertilization levels
处理 春笋产量/ (kg·hm-2) 2007年 2009年 2011年 平均值 ck 2 959.5±82.1 Cc 3 159.0±102.3Cc 3 046.5±79.8Cc 3 055.0±100.0Cc N1 7 657.3±254.1 Bb 7 448.7±213.8Bb 8 001.2±195.2Bb 7 702.4±279.0 Bb N2 8 980.5±246.2 Aa 9 094.5±301.5Aa 9 648.0±257.8 Aa 9 241.0±357.1 Aa N3 8 605.5±269.4 Aa 9 463.5±309.4 Aa 8 953.0±400.2Aa 9 007.3±431.6Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。
下载: 导出CSV
表 3 不同氮素施肥水平条件下毛竹林新竹胸径
Table 3. Diameter at breast height in new Phllostachys edulis at different nitrogen fertilization levels
处理 胸径/cm 2007年 2009年 2011年 平均值 ck 8.71±1.32 Bb 8.16±1.61 Bb 8.43±1.31 Bc 8.43±1.51 Bc N1 9.23±1.12 Bb 8.83±1.34 Bb 9.52±1.13 Bb 9.23±1.21 Bb N2 11.23±0.92 Aa 10.76±1.14 Aa 11.63±1.22 Aa 11.24±1.23 Aa N3 11.55±1.02 Aa 11.11±1.32 Aa 11.47±1.41 Aa 11.34±1.43 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。
下载: 导出CSV
表 4 不同氮素施肥水平条件下毛竹林竹材产量
Table 4. Yield of bamboo timber at different nitrogen fertilization levels
处理 竹材产量/ (kg·hm-2) 2007年 2009年 2011年 平均值 ck 11 700±602 Cc 10 200±719 Cd 10 800±873 Cc 10 900±755 Cc N1 18 563±568 Bb 17 738±610 Bc 18 975±710 Bb 18 425±630 Bb N2 22 688±1 115 Aa 21 450±1 007 Ab 23 513±1 074 Aa 22 550±1 038 Aa N3 23 513±416 Aa 24 383±429 Aa 23 925±402 Aa 23 940±413 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。
下载: 导出CSV
表 5 不同氮素施肥水平条件下毛竹叶片光合色素质量分数
Table 5. Photosynthetie pigment content in leaves of Ph. edulis at different nitrogen fertilization levels
竹龄 处理 叶绿素a/(mg·g-1) 叶绿素b/(mg·g-1) 叶绿素总量/(mg·g-1) 叶绿素a/b 类胡萝卜素/(mg·g-1) ck 2.91±0.10 Bc 1.01±0.09 Bc 3.92±0.15 Bc 2.91±0.29 Aa 0.85±0.07 Ab N1 3.24±0.29 Ab 1.13±0.26 ABb 4.37±0.53 ABb 3.03±0.68 Aa 0.87±0.03 Ab 1年生 N2 3.48±0.17 Aa 1.29±0.09 Aa 4.77±0.24 Aa 2.68±0.15 Ab 0.96±0.06 Aa N3 3.34±0.22 Aab 1.30±0.09 Aa 4.64±0.30 Aab 2.57±0.07 Ab 0.81±0.03 Ab ck 3.01±0.33 Abc 1.01±0.11 Bc 4.01±0.60 Bc 3.02±0.56 Aa 0.86±0.20 Ab N1 3.20±0.30 Ab 1.12±0.11 ABab 4.32±0.68 ABab 2.86±0.21 Aab 0.87±0.10 Ab 3年生 N2 3.43±0.27 Aa 1.31±0.12 Aa 4.74±0.38 Aa 2.62±0.05 Ab 0.94±0.11 Aa N3 3.11±0.24 Abc 1.20±0.07 Aab 4.31±0.38 ABab 2.59±0.14 Ab 0.86±0.11 Ab 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。
下载: 导出CSV
表 6 不同氮素施肥水平条件下毛竹叶片光合参数
Table 6. Photosynthesis parameters in leaves of Ph. edulis at different nitrogen fertilization levels
竹龄 处理 光补偿点/ (μmol · m-2· s-1) 光饱和点/ (μmol · m-2· s-1) 最大光合速率/ (μmol · m-2· s-1) ck 53.3±3.2 Aa 580.3±10.7 Ab 17.3±1.1 Ab N1 46.2±3.1 Ab 590.9±7.8 Aab 18.6±1.3 Aab 1年生 N2 26.9±1.3 Cd 610.0±8.9 Aa 19.3±1.2 Aa N3 34.3±2.7 Bc 597.4±10.1 Aab 18.5±0.9 Aab ck 44.2±2.3 Aa 590.1±9.7 Abc 15.4±0.5 Ab 3年生 N1 37.2±1.9 ABb 621.0±10.3 Aa 16.3±0.2 Aab N2 35.2±3.1 Bb 643.5±9.3 Aa 17.6±0.5 Aa N3 41.2±3.9 ABa 616.4±11.5 Aab 17.2±0.6 Aa 说明:小写字母表示差异在P < 0.05水平下差异显著;大写字母表示差异在P < 0.01水平下差异极显著。
下载: 导出CSV
-
[1] van DELDEN A. Yield and growth components of potato and wheat under organic nitrogen management[J]. Agron J, 2001, 93(6):1370-1385. [2] ZHAO Duli, REDDY K R, KAKANI V G, et al. Corn (Zea mays L.) growth, leaf pigment concentration, photosynthesis and leaf hyperspectral reflectance properties as affected by nitrogen supply[J]. Plant Soil, 2003, 257(1):205-217. [3] da SILVA A F, LOURENCE S O, CHALOUB R M. Effects of nitrogen starvation on the photosynthetic physiology of a tropical marine microalga Rhodomonas sp. (Cryptophyceae)[J]. Aquat Bot, 2009, 91(4):291-297. [4] LONG Jirui, MA Guohui, WAN Yizheng, et al. Effects of nitrogen fertilizer level on chlorophyll fluorescence characteristics in flag leaf of super hybrid rice at late growth stage[J]. Rice Sci, 2013, 20(3):220-228. [5] DEV S P, BHARDWAJ K K R. Effect of crop wastes and nitrogen levels of biomass production and nitrogen uptake in wheat-maize sequence[J]. Ann Agric Res, 1995, 16:264-267. [6] DOMAGALSKI J, LIN Chao, LUO Yang, et al. Eutrophication study at the Panjiakou-Daheiting Reservoir system, northern Hebei Province, People's Republic of China:Chlorophyll-a model and sources of phosphorus and nitrogen[J]. Agr Water Manage, 2007, 94(1):43-53. [7] XIE Yingxing, XIONG Zhengqin, XING Guangxi, et al. An assessment of nitrogen pollutant sources in surface water in Taihu region[J]. Pedosphere, 2007, 17(2):200-208. [8] BOWESA M J, SMITHB J T, JARVIEA H P, et al. Modelling of phosphorus inputs to rivers from diffuse and point sources[J]. Sci Total Environ, 2008, 395(2/3):125-138. [9] JU Xiaotang, KOU C L, CHRISTIE P, et al. Changes in the soil environment from excessive application of fertilizers and manures to two contrasting intensive cropping systems on the North China Plain[J]. Environ Pollut, 2007, 45(2):497-506. [10] 胡集瑞.施肥对毛竹笋产量和出笋规律的影响[J].福建林业科技, 2000, 27(1):34-35. HU Jirui. Effects of the fertilizer application on the bamboo shoot yield and bamboo shooting law of Phyllostachys pubescens[J]. J Fujian For Sci Technol, 2000, 27(1):34-35. [11] 李睿, 维尔格M J A, 钟章成.施肥对毛竹(Phyllostachys pubescens)竹笋生长的影响[J].植物生态学报, 1997, 21(1):19-26. LI Rui, WERGER M J A, ZHONG Zhangcheng. Influence of fertilization on the clonal growth of bamboo shoots in Phyllostachys pubescens[J]. Acta Phytoecol Sin, 1997, 21(1):19-26. [12] 王宏, 金晓春, 金爱武, 等.施肥对毛竹生长量和秆形的影响[J].浙江农林大学学报, 2011, 28(5):741-746. WANG Hong, JIN Xiaochun, JIN Aaiwu, et al. Growth and culm form of Phyllostachys pubescens with fertilization[J]. J Zhejiang A & F Univ, 2011, 28(5):741-746. [13] 金晓春, 金爱武, 宋艳冬, 等.施肥对毛竹林换叶期冠层形成及光合能力的影响[J].浙江林学院学报, 2010, 27(1):57-62. JIN Xiaochun, JIN Aiwu, SONG Yandong, et al. Canopy development and photosynthesis with fertilization during leaf expansion period of Phyllostachys pubescens[J]. J Zhejiang For Coll, 2010, 27(1):57-62. [14] 宋艳冬, 金爱武, 金晓春, 等.施肥对毛竹叶片光合生理的影响[J].浙江林学院学报, 2010, 27(3):334-339. SONG Yandong, JIN Aiwu, JIN Xiaomei, et al. Physiology of leaf photosynthesis with fertilization in Phyllostachys pubescens[J]. J Zhejiang For Coll, 2010, 27(3):334-339. [15] LICHTENTHALER H K, WELBURN A R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents[J]. Biochem Soc Transac, 1983, 603:142-196. [16] ANDERSON J M, GOODCHILD D J, BOARDMAN N V. Composition of the photosystems and chloroplast structure in extreme shade plants[J]. Biochem Biophys Acta, 1973, 325(3):573-585. [17] 冯玉龙, 刘利刚, 王文章, 等.长白落叶松水曲柳混交林增产机理的研究(Ⅱ)落叶松水曲柳混交林生理特性的研究[J].东北林业大学学报, 1996, 24(3):1-7. FENG Yulong, LIU Ligang, WANG Wenzhang, et al. Study on the mechanism of production increasing of mixed Larix olgensis Henry and Fraxinus mandshurica stand (Ⅱ) Study on physiological characteristics of mixed L. olgensis Henry and F. mandshurica stand[J]. J Northeast For Univ, 1996, 24(3):1-7. [18] HUANG Gaiqun, WEI Ling, TONG Wanhong, et al. Effects of N, P and K fertilizers on photosynthetic characteristics of mulberry[J]. Agric Sci Technol, 2012, 13(12):2562-2568. [19] HEITHOLT J J, JOHNSON R C, FERRIS D M. Stomatal limitation to carbon dioxide assimilation in nitrogen and drought stressed wheat[J]. Crop Sci, 1991, 31(1):135-139. [20] ELRILI I R, HOLMES J J, WEGER H G, et al. RuBP limitation of photosynthetic carbon fixation during NH3 assimilation. Interactions between photosynthesis, respiration and ammonium assimilation in N-limited green algae[J]. Plant Physiol, 1988, 87(2):395-401. [21] FOYER C H, NOCTOR G, LELANDAIS M, et al. Short-term effects of nitrate, nitrite and ammonium assimilation on chlorophyll a fluorescence, thylakoid protein phosphorylation, net CO2 assimilation and amino acid biosynthesis in maize[J]. Planta, 1994, 192(21):211-220. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.05.006
 点击查看大图
点击查看大图

计量
- 文章访问数: 8688
- HTML全文浏览量: 804
- PDF下载量: 677
- 被引次数: 0
