





-
光合作用是植物最主要的生理活动,是植物生长发育的基础[1],除了直接测定叶片净光合速率外,叶绿素荧光也往往被用来反映其光合能力的强弱[2]。杜仲Eucommia ulmoides是中国特有的经济树种,也是分布范围最广的胶原植物[3],‘红叶’杜仲Eucommia ulmoides ‘Hongye’和‘小叶’杜仲Eucommia ulmoides ‘Xiaoye’是中国林业科学研究院经济林研究开发中心选育的优良杜仲无性系。‘小叶’杜仲叶片形态优美,‘红叶’杜仲颜色鲜红,且此2个杜仲无性系叶片有效活性成分均远超其他无性系,观赏和药用价值均较高,发展前景广阔[4]。近年来,前人[5-9]对杜仲叶片的光合作用及叶绿素荧光参数进行初步研究,但采用的研究材料均为实生苗,试验误差控制不严格,以至于研究的重复性及可比性均较差。本研究以‘红叶’杜仲和‘小叶’杜仲2个无性系苗期成熟叶片为研究材料,通过对其叶片传统光响应曲线、快速光曲线及不同温度条件下‘红叶’杜仲叶片的传统光响应曲线的测定,探寻2个杜仲无性系叶片光合能力的差异,以期为其高效栽培技术提供理论依据。
-
实验材料为‘红叶’杜仲和‘小叶’杜仲无性系幼苗,2011年播种,当年7月下旬嫁接,2012年12月平茬,测定时苗高121.0~157.0 cm,地径1.0~1.3 cm。每个无性系随机选择3个生长状况良好的单株,每个单株选择1片朝向东南方位的叶片,位于从顶芽开始第15~20片叶(约生长30 d,位于4/5苗高处)。将‘红叶’杜仲依次编号为1,3,5;‘小叶’杜仲依次编号为2,4,6,灌透水后进行叶片光合特性和叶绿素荧光特性测定。测定期间大气中二氧化碳摩尔分数为364.000~391.000 μmol· mol-1,气温32~38 ℃。
-
2013年8月24日-26日,每天早上9:00-11:00进行杜仲叶片传统光响应曲线测定。测定方法为:用Li-6400光合分析仪在0~2 000 μmol·m-2·s-1光合有效辐射(PPAR)范围内,利用人工光源设定光合有效辐射分别为2 000,1 800,1 600,1 400,1 200,1 000,800,600,400,200,100,50,30,0 μmol·m-2·s-1,从强光到弱光依次测定各叶片的净光合速率(Pn),测定1次·d-1,连续测定3 d。第1天按照1—2—3—4—5—6顺序测定,第2天按照6—5—4—3—2—1顺序测定,第3天按照4—5—6—1—2—3顺序测定。分别计算各条传统光响应曲线的光合作用参数,以每个无性系的光合有效辐射及对应的净光合速率的平均值绘制其传统光响应曲线。
-
2013年8月24上午9:00-11:00,用便携式调制荧光仪PAM-2500(德国)按照6—5—4—3—2—1的顺序进行杜仲2个无性系快速光响应曲线(EETR-PPAR)测定。EETR为光合电子传递的相对速率,光合有效辐射设定为2,52,133,236,407,659,998,1 386,1 973,2 845 μmol·m-2·s-1。
-
2013年8月27-29日上午9:00-11:30,以‘红叶’杜仲叶片为研究对象,用Li-6400光合分析仪测定20,23,25,28,30,32和35 ℃温度下的传统光响应曲线,光合有效辐射依次为0,20,50,100,200,400,600,800,1 000,1 500,1 800,2 000,2 500 μmol·m-2·s-1,叶片按照1—3—5测定,测定1片·d-1。
-
使用Spss 16.0和Excel 2007软件进数据处理。杜仲叶片Pn对PPAR的响应关系采用直角双曲线模型[10-11]进行拟合:
$$ {P_{\rm{n}}}=\left({\alpha {\rm{ \times }}{P_{{\rm{PAR}}}}{\rm{ \times }}{P_{\max }}} \right)/\left({\alpha {\rm{ \times }}{P_{{\rm{PAR}}}}{\rm{+}}{P_{\max }}} \right)- R. $$ (1) 其中:Pn为特定PPAR条件下的净光合速率。PPAR为光合有效辐射,α为初始量子斜率,即植物光合作用对传统光响应曲线在PPAR=0时的斜率。Pmax代表最大净光合速率,R为暗呼吸速率。
令式(1)中Pn=0,可得光补偿点(PLCP)为:
$$ {P_{{\rm{LCP}}}}=\frac{{R \cdot {P_{\max }}}}{{\alpha \left({{P_{\max }} - R} \right)}}. $$ (2) 利用光能能力(Pa)为最大净光合速率与暗呼吸速率之和,即:
$$ {P_{\rm{a}}}={P_{\max }}+R. $$ (3) 根据经验公式,光饱和点计算公式为:
$$ {P_{{\rm{LSP}}}}=\left({{P_{\max }}+R} \right)/{P_{{\rm{AQE}}}}={P_{\rm{a}}}/{P_{{\rm{AQE}}}}. $$ (4) 式(4)中PAQE为弱光合有效辐射条件下(≤200 μmol·m-2·s-1)用直线方程拟合光响应数据得到表观量子效率。
-
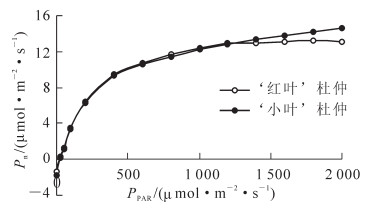
杜仲2个无性系的传统光响应曲线如图 1所示,净光合速率随着光合有效辐射的增加而升高,不过前期增幅较大,后期增幅较小。在0~1 200 μmol·m-2·s-1的光合有效辐射范围内,‘红叶’杜仲的净光合速率略高于‘小叶’杜仲,而在1 400~2 000 μmol·m-2·s-1的范围内,‘红叶’杜仲净光合速率随着光合有效辐射的增加没有显著变化,而‘小叶’杜仲的净光合速率则随着光合有效辐射的增加继续升高,且超过‘红叶’杜仲。2个杜仲无性系的光合作用参数如表 1所示,‘小叶’杜仲的光补偿点和最大净光合速率分别高于‘红叶’杜仲10.55%和9.47%,2个无性系的暗呼吸速率几乎相同,而初始斜率则是‘红叶’杜仲高于‘小叶’杜仲11.43%。2个杜仲无性系的传统光响应曲线及光合参数值均说明‘红叶’杜仲利用弱光至中强光能力较强,而‘小叶’杜仲利用强光的能力较强。

图 1 杜仲2个无性系的传统光响应曲线
Figure 1. Light-response curve of 2 Eucommia ulmoides clones
表 1 杜仲2个无性系光合作用参数值
Table 1. Photosynthesis parameter of 2 Eucommia ulmoides clones (mean±sd)
无性系 叶片编号 PLCP /(μmol·m-2·s-1) PLSP /(μmol·m-2·s-1) Pmax /(μmol·m-2·s-1) PAQE R /(μmol·m-2·s-1) 初始斜率 1 18.000±5.000 453.000±19.000 15.350±0.940 0.037±0.001 1.290±0.330 0.080±0.003 3 25.000±6.000 536.000±53.000 18.420±1.110 0.038±0.002 1.700±0.390 0.075±0.004 ‘红叶’杜仲 5 29.000±7.000 499.000±9.000 16.950±0.880 0.038±0.003 2.010±0.500 0.078±0.005 平均 24.000±6.000 496.000±41.000 16.910±1.540 0.038±0.001 1.670±0.360 0.078±0.003 2 28.000±15.000 593.000±57.000 21.780±2.490 0.039±0.002 1.530±1.040 0.055±0.015 ‘小叶’杜仲 4 30.000±13.000 521.000±35.000 18.430±1.650 0.039±0.003 1.940±0.990 0.072±0.012 6 21.000±2.000 451.000±15.000 15.300±0.670 0.037±0.001 1.560±0.050 0.082±0.007 平均 26.000±5.000 522.000±7.000 18.500±3.240 0.039±0.001 1.670±0.230 0.070±0.014 -
杜仲2个无性系的快速光曲线如图 2所示,在250~1 400 μmol·m-2·s-1,‘红叶’杜仲EETR高于‘小叶’杜仲,而2 000 μmol·m-2·s-1以上,‘小叶’杜仲EETR则超过‘红叶’杜仲,说明‘红叶’杜仲利用中强光的能力高于‘小叶’杜仲,而‘小叶’杜仲利用强光的能力优于‘红叶’杜仲。该结果与通过Li-6400得到的结果类似。

图 2 杜仲2个无性系叶绿素焚光快速响应曲线
Figure 2. Chlorophyll fluorescence rapid light-response curve of 2 Eucommia ulmoides clones
-
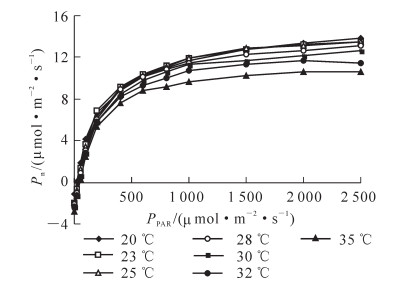
温度对‘红叶’杜仲光合作用的影响如图 3和表 2所示。由图 3可知:不同温度条件下的净光合速率在0~2 500.000 μmol·m-2·s-1的范围内均随着光合有效辐射的增加而增加,但在0~1 000.000 μmol·m-2·s-1范围内,增幅较大,而在1 000.000~2 500.000 μmol·m-2·s-1范围内,增幅明显降低。在35 ℃条件下,任何光照下的净光合速率均低于其他温度水平。20 ℃时的净光合速率在0~100 μmol·m-2·s-1光合有效辐射的范围内高于其他水平,在200~1 000 μmol·m-2·s-1居中,在1 500 μmol·m-2·s-1以上又达到最高。

图 3 '红叶'杜仲在不同温度下的传统光响应曲线
Figure 3. Light-response curve of Eucommia ulmoide 'Hongye' under different tempreatures
表 2 ‘红叶’杜仲在不同温度下的光合作用参数
Table 2. Photosynthesis parameter of Eucommia ulmoides 'Hongye' under different tempuretures (mean±sd)

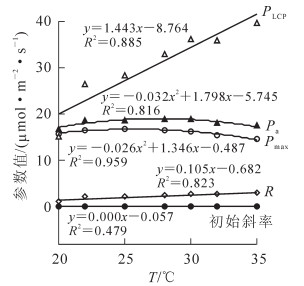
‘红叶’杜仲的光合作用参数与温度的关系如图 4所示。温度与各光合作用参数间存在较强的线性回归关系,回归方程的决定系数为0.790~0.959,‘红叶’杜仲的光合效率、暗呼吸速率及光补偿点均随着温度的上升而升高。‘红叶’杜仲叶片利用光能能力在随着温度的升高出现先升后降的趋势。由于光合参数与温度存在显著的关系,描述光合参数时有必要同时描述其特定的温度条件。

图 4 ‘红叶’杜仲温度与光合作用参数关系
Figure 4. R elationship between tempreatures and photosynthesis parameter of Eucommia ulmoides 'Hongye'
-
本研究在前人研究的基础上,选用了2个杜仲无性系为研究材料,由于排除了以前研究中实生苗之间的遗传差异,测定时又通过固定叶片及2个无性系交替测量的方法,有效地控制试验误差,准确地反映2个无性系的光合能力。研究得到‘红叶’杜仲的光补偿点为24.000 μmol·m-2·s-1,光饱和点为496.000 μmol·m-2·s-1,最大净光合速率为16.910 μmol·m-2·s-1,暗呼吸速率1.670 μmol·m-2·s-1,初始斜率为0.078;‘小叶’杜仲的光补偿点为26.000 μmol·m-2·s-1,光饱和点为521.000 μmol·m-2·s-1,最大净光合速率为18.500 μmol·m-2·s-1,暗呼吸速率为1.670 μmol·m-2·s-1,初始斜率为0.070;‘小叶’杜仲和‘红叶’杜仲的光补偿点、光饱和点和暗呼吸速率高于黄秦军[12]选择的3个杨树无性系。最大净光合速率高于杨树L3无性系,却均低于杨树H4无性系。说明了杜仲是强阳性树种,‘小叶’杜仲在强光环境下生长状况优于‘红叶’杜仲。
本研究以Li-6400光合仪和PAM-2500荧光仪研究2个杜仲无性系的光合能力,通过对2个杜仲无性系传统光响应曲线和快速光曲线异同之处的剖析,解释了不同之处的根本原因。研究表明杜仲2个无性系快速光曲线虽然和传统光响应曲线相似,但仍有不同。传统光响应曲线在0~1 000 μmol·m-2·s-1的光合有效辐射范围内,净光合速率随光照增加增幅明显,但1 000 μmol·m-2·s-1以后增幅逐渐减小。而快速光曲线则表现为,0~1 000 μmol·m-2·s-1范围内光合电子传递相对速率(EETR)随光合有效辐射增加而增加,但其增加幅度小于净光合速率增幅,但光合有效辐射在1 000 μmol·m-2·s-1之后,光合电子传递相对速率随着光合有效辐射的增加,仍保持相对较高的增幅。该结果与2种仪器的测定原理有关,Li-6400测定的是实际固定的二氧化碳量,受植物光合能力和呼吸作用的双重影响,叶绿素荧光测定的是叶片利用光能的能力,由于在1 000 μmol·m-2·s-1之后,光合有效辐射的增加会引起叶片温度的迅速上升,而叶片温度的上升除了能提高光能利用能力,同时也导致暗呼吸速率的升高,两者有一定的抵消,所以才产生了在较强光照条件下,净光合速率增加幅度降低,最终致使2种方法得到的光曲线形状略有不同。
本研究以‘红叶’杜仲为对象,研究了不同温度对杜仲光合参数的影响,结果表明,‘红叶’杜仲不同温度水平下的传统光响应曲线特征也略有不同,在35 ℃条件下,任何光照下的净光合速率均低于其他温度水平,这是因为温度与暗呼吸速率存在显著的正相关。该温度条件下,暗呼吸速率高于其他水平,甚至达到20 ℃下暗呼吸速率的298.5%,较高的暗呼吸速率使得净光合速率低于其他温度水平。20 ℃温度条件下的净光合速率在0~100 μmol·m-2·s-1光合有效辐射的范围内高于其他水平,在200~1 000 μmol·m-2·s-1居中,在1 500 μmol·m-2·s-1以上又达到最高。该现象的产生可能是在0~100 μmol·m-2·s-1的弱光条件下,碳同化效率均较低。此时,对净光合速率影响最大的因素为暗呼吸速率,温度越低,暗呼吸速率越小,所以此时20 ℃的净光合速率高于其他水平;当光合有效辐射超过1 500 μmol·m-2·s-1时,较强的光合有效辐射使得叶片温度迅速上升,有可能超过叶片的最适温度而使得碳同化能力有所下降,最主要的是增强暗呼吸速率,而将叶室温度控制在较低温度时,又可以一定程度上的降低叶片温度,使得暗呼吸速率降低,所以20 ℃最终净光合速率在强光下达到最大。
开展的‘红叶’杜仲光合参数随温度变化规律的研究,得到温度和‘红叶’杜仲各光合参数间的线性方程的决定系数为0.479~0.959。随着温度的增加,‘红叶’杜仲的光合能力和净光合速率呈现先升高后降低的趋势,而暗呼吸速率、光补偿点、初始斜率则是呈直线上升,上升幅度在33.8%~198.5%。温度对光合作用的影响主要是因为影响了酶促反应速度。温度低时,光合酶活性低,二氧化碳同化慢,淀粉与蔗糖的合成减少,对磷酸丙糖的需求量下降,进入叶绿体的无机磷酸减少,同化力的合成受到限制[1]。温度高时,酶钝化,光合机构受损,而导致光合速率迅速下降。同时由于暗呼吸速率的最适温度比光合作用的高,当光合作用不再升高时,暗呼吸速率继续升高,而使净光合速率下降。温度对‘红叶’杜仲叶片净光合速率的影响主要为在弱光和强光条件下影响大,在中等强度光照条件下影响小,该研究结论与王玉涛[13]等的研究结果相似。根据研究结果,在光热可控条件下栽培杜仲,可将温度控制在23~25℃,‘红叶’杜仲光照条件控制在为1 000~1 500 μmol·m-2·s-1,‘小叶’杜仲的光合有效辐射可适当提高至1 200~1 800 μmol·m-2·s-1,此时既能达到较高的净光合速率,也不会浪费大量能源。研究结论为杜仲光合特性的深入研究提供了理论依据,但不足之处是仅针对‘红叶’杜仲1个无性系开展研究,如果能由多个无性系共同研究,则结论会更具有说服性。
Photosynthetic capacity of two Eucommia ulmoides clones
-
摘要: 为了揭示杜仲Eucommia ulmoides不同无性系光合特性变异规律, 以杜仲2个观赏型无性系的苗期叶片为材料, 分别采用Li-6400和PAM2500测定其传统光响应曲线、快速光曲线及不同温度条件下的传统光响应曲线, 通过直角双曲线模型计算其光合作用参数。结果表明:‘红叶’杜仲Eucommia elmoides ‘Hongye’的光补偿点为23.960 μmol·m-2·s-1, 光饱和点为495.940 μmol·m-2·s-1, 最大净光合速率为16.907 μmol·m-2·s-1, 暗呼吸速率为1.666 μmol·m-2·s-1, 初始斜率为0.078;‘小叶’杜仲Eucommia elmoides ‘Xiaoye’的光补偿点为26.487 μmol·m-2·s-1, 光饱和点为521.920 μmol·m-2·s-1, 最大净光合速率为18.502 μmol·m-2·s-1, 暗呼吸速率为1.674 μmol·m-2·s-1, 初始斜率为0.070;‘红叶’杜仲温度和各光合参数间存在显著的线性回归关系, 回归方程的决定系数为0.479~0.959。杜仲2个无性系的光合作用参数表明, ‘红叶’杜仲更适宜在中强光环境中生长, ‘小叶’杜仲则在强光环境下生长状况优于‘红叶’杜仲。研究揭示的2个杜仲无性系的光合作用参数及变异规律为杜仲后期生理生化特性及生态适应性研究提供了依据。Abstract: To provide a theoretical basis with Eucommia ulmoides for ecological adaptability and on breeding of new varieties, leaves of two ornamental E. ulmoides clone seedlings ('Hongye' and 'Xiaoye') were selected and compared. For each clone, three leaves (i.e. three replicates) were selected and measured alternatively. Light response curves for both clones were measured using a Li-6400 and a rapid light curve found with a PAM2500. Light response curves under different temperatures for 'Hongye' were measured too. Photosynthetic parameters were computed by rectangular hyperbola model. Equations of linear regression between photosynthetic parameters and the temperatures were fitted. Results for 'Hongye' leaves (in μmol·m-2·s-1) showed a light compensation point of 23.96, a light saturation point of 495.94, a maximum net photosynthetic rate of 16.907, and a respiration rate of 1.666; the initial slope was 0.078.'Xiaoye'leaves(in μmol·m-2·s-1) revealed a light compensation point of 26.487, a light saturation point of 521.920, a maximum net photosynthetic rate of 18.502, and a respiration rate of 1.674; the initial slope was 0.070. There were significant regression relationship between temperatures and photosynthetic parameters of 'Hongye'(R2=0.479-0.959). These results suggest that that E. ulmoides is a strong heliphilous species and temperature has significant effects on E. ulmoides photosynthetic parameters.
-
Key words:
- cash forestry /
- Eucommia ulmoides /
- clones /
- photosynthesis /
- rapid light curve /
- temperature
-
图 2 杜仲2个无性系叶绿素焚光快速响应曲线
Figure 2 Chlorophyll fluorescence rapid light-response curve of 2 Eucommia ulmoides clones
图 3 '红叶'杜仲在不同温度下的传统光响应曲线
Figure 3 Light-response curve of Eucommia ulmoide 'Hongye' under different tempreatures
图 4 ‘红叶’杜仲温度与光合作用参数关系
Figure 4 R elationship between tempreatures and photosynthesis parameter of Eucommia ulmoides 'Hongye'
表 1 杜仲2个无性系光合作用参数值
Table 1. Photosynthesis parameter of 2 Eucommia ulmoides clones (mean±sd)
无性系 叶片编号 PLCP /(μmol·m-2·s-1) PLSP /(μmol·m-2·s-1) Pmax /(μmol·m-2·s-1) PAQE R /(μmol·m-2·s-1) 初始斜率 1 18.000±5.000 453.000±19.000 15.350±0.940 0.037±0.001 1.290±0.330 0.080±0.003 3 25.000±6.000 536.000±53.000 18.420±1.110 0.038±0.002 1.700±0.390 0.075±0.004 ‘红叶’杜仲 5 29.000±7.000 499.000±9.000 16.950±0.880 0.038±0.003 2.010±0.500 0.078±0.005 平均 24.000±6.000 496.000±41.000 16.910±1.540 0.038±0.001 1.670±0.360 0.078±0.003 2 28.000±15.000 593.000±57.000 21.780±2.490 0.039±0.002 1.530±1.040 0.055±0.015 ‘小叶’杜仲 4 30.000±13.000 521.000±35.000 18.430±1.650 0.039±0.003 1.940±0.990 0.072±0.012 6 21.000±2.000 451.000±15.000 15.300±0.670 0.037±0.001 1.560±0.050 0.082±0.007 平均 26.000±5.000 522.000±7.000 18.500±3.240 0.039±0.001 1.670±0.230 0.070±0.014  下载: 导出CSV
下载: 导出CSV
表 2 ‘红叶’杜仲在不同温度下的光合作用参数
Table 2. Photosynthesis parameter of Eucommia ulmoides 'Hongye' under different tempuretures (mean±sd)
-
[1] 武维华.植物生理学[M]. 2版.北京:科学出版社, 2008:130-190. [2] 李鹏民, 高辉远, STRASSER R J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理学与分子生物学学报, 2005, 31(6):559-566. LI Pengmin, GAO Huiyuan, STRASSER R J. Application of the fast chlorophyll fluorescence Induction dynamics analysis in photosynthesis study[J]. J Plant Physiol Mol Biol, 2005, 31(6):559-566. [3] 李芳东, 杜红岩.杜仲[M].北京:中国中医药出版社, 2001:232-256. [4] 杜红岩, 胡文臻, 俞锐.杜仲产业绿皮书:中国杜仲橡胶资源与产业发展报告(2013)[M].北京:社会科学文献出版社, 2013:114-132. [5] 李淑容, 谈锋.杜仲对不同光强度的适应性研究[J].西南师范大学学报:自然科学版, 1995, 20(3):293-296. LI Shurong, TAN Feng. Studies on the adaptation of Eucommia ulmoides to light intensity[J]. J Southwest China Norm Univ Nat Sci, 1995, 20(3):293-296. [6] 郭连旺, 沈允钢, 武海, 等.杜仲光合特性的研究[J].植物学报, 1996, 38(4):283-286. GUO Lianwang, SHEN Yungang, WU Hai, et al. Study on the photosynthetic characteristics of Eucommia ulmoides leaves[J]. Acta Bot Sin, 1996, 38(4):283-286. [7] 刘淑明, 梁宗锁, 董娟娥.水分对杜仲截干萌条光合特性及生长的影响[J].西北农林科技大学学报:自然科学版, 2008, 36(4):89-93. LIU Shuming, LIANG Zongsuo, DONG Juane. Photosynthetic characteristics and growth of sprout of Eucommia ulmoides under different water conditions[J]. J Northwest A & F Univ Nat Sci Ed, 2008, 36(4):89-93. [8] 高超, 闫文德, 田大伦, 等.杜仲光合速率日变化及其与环境因子的关系[J].中南林业科技大学学报, 2011, 31(5):100-104. GAO Chao, YAN Wende, TIAN Dalun, et al. Diurnal change in photosynthetic rate of Eucommia ulmoides and its relationship with environmental factors[J]. J Cent South Univ For & Technol, 2011, 31(5):100-104. [9] 李蕾.四倍体杜仲光合特性的研究[D].河北农业大学, 2012. LI Lei. Study on Photosynthetic Characteristics of Tetraploid Eucommia ulmoides Oliv.[D]. Baoding:Agricultural University of Hebei, 2012. [10] LEAKEY A D B, URIBELARREA M, AINSWORTH E A, et al. Photosynthesis, productivity, and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought[J]. Plant Physiol, 2006, 140(2):779-790. [11] 叶子飘, 于强.光合作用光响应模型的比较[J].植物生态学报, 2008, 32(6):1356-1361. YE Zipiao, YU Qiang. Compatision of new and several classical models of photosynthesis in response to irradiance[J]. J Plant Ecol, 2008, 32(6):1356-1361. [12] 黄秦军, 黄国伟, 丁昌俊, 等.美洲黑杨杂种不同生长势无性系光合特征[J].林业科学, 2013, 49(3):56-62. HUANG Qinjun, HUANG Guowei, DING Changjun, et al. Comparative analysis of photosynthetic characteristics of Populus deltoides clones with different growth vigor[J]. Sci Silv Sin, 2013, 49(3):56-62. [13] 王玉涛, 马志波, 马钦彦, 等.北京地区4种阔叶树光合作用对CO2浓度及温度变化的响应[J].河北农业大学学报, 2006, 29(6):39-43. WANG Yutao, MA Zhibo, MA Qinyan, et al. Photosynthesis response to variation of CO2 concentration and temperature of four broad-leaved trees in Beijing region[J]. J Agric Univ Hebei, 2006, 29(6):39-43. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.05.007
 点击查看大图
点击查看大图
计量
- 文章访问数: 3585
- HTML全文浏览量: 645
- PDF下载量: 547
- 被引次数: 0
