




 下载:
下载:


-
萜(terpenoid)是以异戊二烯(isoprene)为基本单元的生物大分子,在人们日常生活、食品、医疗保健、化工材料及军事等领域蕴藏着巨大的商业价值。甲羟戊酸途径(mevalonate pathway,MVA pathway)和2-甲基-D-赤藓醇-4-磷酸途径(2-C-methyl-D-erythritol-4-phosphate,MEP pathway)是生物中萜类物质合成的2条基本途径,前者存在于几乎所有古生菌和真核生物,也存在于一些革兰氏阳性菌中;后者存在于大多数细菌和植物体内[1-2]。2-甲基-D-赤藓醇-2,4-环焦磷酸合酶(2-C-Methyl-D-erythritol-2,4-cyclodiphosphate synthase,MDS)是MEP途径第5个作用酶,催化4-(5′-焦磷酸胞苷)-2-C-甲基-D-赤藓醇-2-磷酸(4-diphosphocytidyl-2-C-methyl-D-erythritol-2-phosphate,CDP-MEP)生成2-甲基-D-赤藓醇-2,4-环焦磷酸(2-C-methyl-D-erythritol-2,4-cyclodiphosphate,MEcPP),该反应需要镁离子(Mg2+)或锰离子(Mn2+)协助,并释放出1分子胞苷磷酸[3-4]。一些研究认为MDS是MEP途径关键酶,在异戊烯焦磷酸(IPP)生物合成过程中起重要的调节作用。在悬浮培养长春花细胞中发现上调MDS基因可提高单萜类物质吲哚胆碱的含量,也有利于MEP途径代谢流向更下游的方向[5]。半定量RT-PCR结果显示红豆杉Taxus chinensis和银杏Ginkgo biloba MDS基因具有组织特异性,并都以叶中表达量最高[6-7]。杜仲Eucommia ulmoides是中国名贵的中药材和工业橡胶原料树种,适生于华中、华西、西南及西北各地,现广泛栽培[8]。以杜仲胶和环烯醚萜类为典型的杜仲萜类次生产物具有重要的应用与经济价值,其中杜仲胶属多萜化合物,具有优良的共混和加工性能,是材料领域重要的新型战略物质[9-10];杜仲环烯醚萜类属单萜化合物,具有利胆、镇痛、保肝、抗癌、抗炎、抗氧化以及抗骨质疏松等功能,是保健及医药领域重要的天然活性成分[11-12]。目前对杜仲萜类生物合成特别是MEP途径相关作用基因开展的研究不多,对杜仲MDS基因的克隆及序列分析尚无报道。本研究以杜仲叶片为材料,分离MDS同源基因全长cDNA,通过生物信息学方法对基因序列及推导的氨基酸序列进行分析,以期为研究杜仲MDS基因功能,阐释杜仲萜类生物合成机制和分子育种提供参考。
-
2011年4月下旬于中国林业科学研究院经济林研究开发中心院内采集杜仲良种‘华仲6号’叶片,清洗干净后投入液氮带回室内-80 ℃保存备用。
-
焦碳酸二乙酯(DEPC)(Sigma,德国),聚乙烯吡咯烷酮(PVP)(Amresco,美国),十二烷基磺酸钠(SDS)(上海生工,中国),氯化锂(Amresco,美国),十六烷基三甲基溴化铵(CTAB)(Amresco,美国),MightyAmp DNA Polymerase Ver.2(Takara,中国大连),琼脂糖凝胶DNA回收试剂盒(天根,中国北京),3′-Full RACE Core Set(Takara,中国大连),5′-Full RACE Kit(Takara,中国大连),M-MLV RTase cDNA Synthesis Kit(Takara,中国大连),pEASY-T1 Cloning Kit(全式金,中国北京),DH5α感受态细胞(天根,中国北京)。
-
采用改良的CTAB-LiCl法提取杜仲叶片RNA[13-14],对满足实验要求的RNA样品保存于-80 ℃冰箱备用。按M-MLV RTase cDNA Synthesis Kit说明书进行单链cDNA的合成。
-
根据一段已知杜仲MDS unigene序列,结合试剂盒锚定引物序列,设计适于3′RACE的巢式扩增的引物3P1:5′-CGTACACTCGGTTCTCGTT-3′,3P2:5′-CTCGACTCCGTCGAAGTCGCTC-3′以及5′RACE巢式扩增引物5P1:5′-TGGATCGGAATCTGGAAATATC-3′,5P2:5′-AGCAATACATCGCCGTCGGAGT-3′。
-
3′RACE及5′RACE 的聚合酶链式反应(PCR) 反应体系与反应条件参照Takara 3′-Full RACE Core Set Ver.2.0与Takara 5′-Full RACE Kit 说明书操作。
-
按TIANGEN通用型DNA回收试剂盒说明进行目的PCR 产物的回收;按pEAZY-T克隆试剂盒说明将扩增片段连接至克隆载体,鉴定后将阳性克隆送至南京金斯瑞公司测序。
-
利用美国生物技术信息中心NCBI(http://www.ncbi.nlm.nih.gov)Blast程序进行序列相似性检索,并用ORF Finder程序查找基因cDNA开放阅读框架;利用ExPASy(http://cn.expasy.org)ProtParam程序与ScanProsite程序分析氨基酸残基数目与组成、蛋白质相对分子量、理论等电点以及功能位点等;利用Predict Protein()以及PSIPRED方法(http://bioinf.cs.ucl.ac.uk/psipred)预测蛋白质的细胞定位及二级结构[15];利用ChloroP 1.1 Server()进行转运肽的预测[16];利用SWISS-MODEL程序(http://swissmodel.expasy.org/)进行蛋白质的同源建模[17];利用Lasergene软件进行蛋白质序列的多重比对并通过MEGA 5软件构建基因的系统进化树[18]。
-
分别利用3′RACE引物和5′RACE引物在逆转录的cDNA模板上扩增出1条约650 bp和1条约500 bp的特异条带(图 1a,图 1b),测序拼接后得到1条长976 bp的基因序列,与紫茎泽兰Ageratina adenophora(GU828010.1),甜菊Stevia rebaudiana(DQ631427.3),长春花Catharanthus roseus(EU034700.1),、葡萄Vitis vinifera(XM_002278370.1),芜菁Brassica rapa(AB300309.1)MDS序列的相似性分别为79%,78%,79%,77%,79%。通过ORF finder工具查找到1个长711 bp的开放阅读框,5′-UTR长119 bp,3′-UTR长146 bp,共编码236个氨基酸残基(图 2)。推导氨基酸序列与毛果杨Populus trichocarpa(XP_002304519.1),橡胶Hevea brasiliensis(AAS94122.1),啤酒花Humulus lupulus(AEV89963.1),丹参Salvia miltiorrhiza (AEZ55667.1),萝芙木Rauvolfia verticillata(ABV89583.1)MDS蛋白序列相似性分别为73%,73%,76%,74%,76%,确定取得杜仲MDS基因cDNA全长序列,将其命名为EuMDS。

图 1 EWlffiS基因RT-PCR扩增的琼脂糖凝胶电泳图
Figure 1. Agarose gel electrophoresis of RT-PCR amplification of EuMDS

图 2 EuMDS全长cDNA序列及推导的氨基酸序列
Figure 2. Full-length sequence of cDNA and deduced amino acid sequence of EuMDS
-
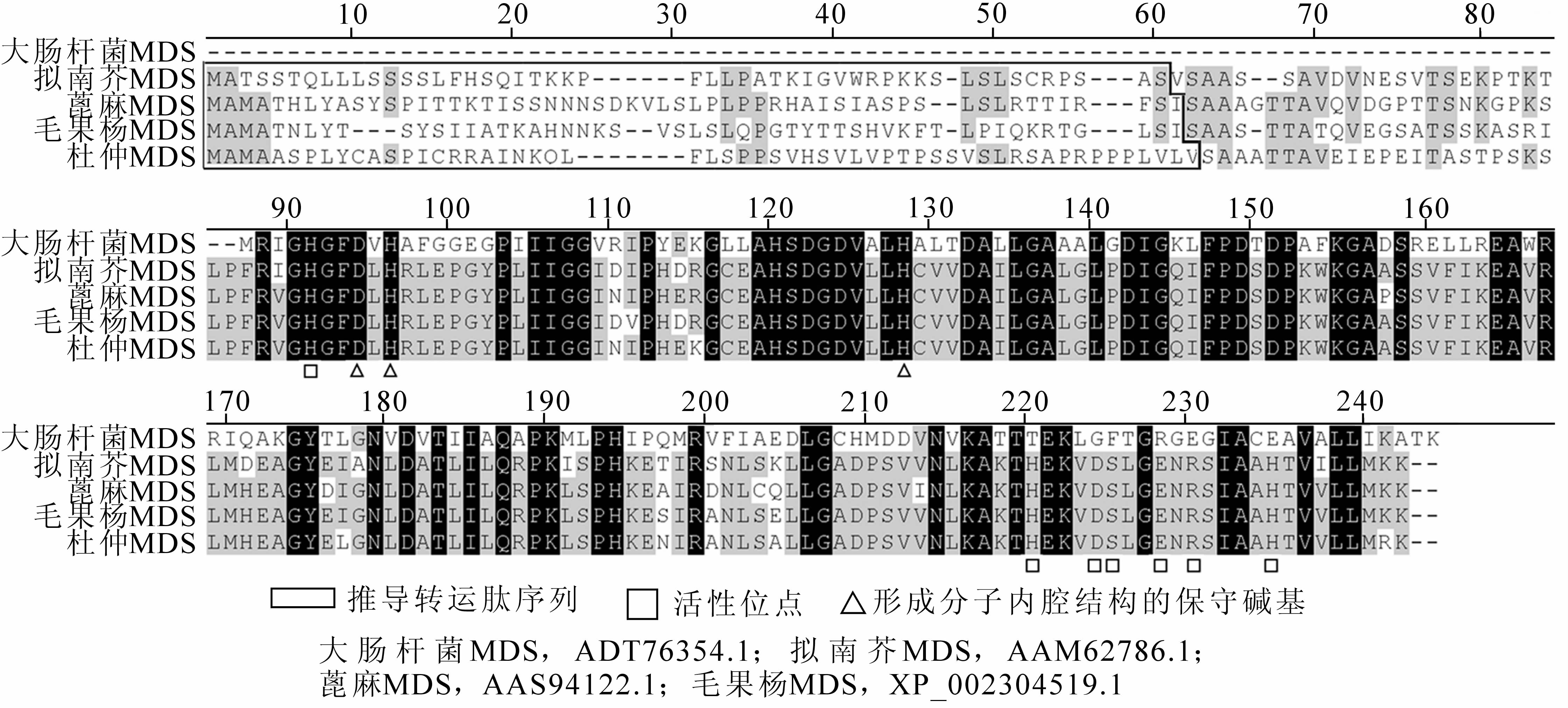
ExPaSy ProtParam程序预测EuMDS编码蛋白分子量为25.14 kD,理论等电点为7.77;氨基酸组成中以亮氨酸(12.3%),丙氨酸(10.6%),脯氨酸(9.3%),丝氨酸(9.3%)含量较高,蛋白不稳定系数50.02,属不稳定蛋白质。TargetP 1.1 Server预测EuMDS亚细胞定位于叶绿体上,预测分值为0.943,可靠性Ⅰ级。Expasy protscale程序推断EuMDS为疏水性蛋白。Vector NTI Advance 10多重比对显示EuMDS蛋白具有植物MDS蛋白典型的保守位点(图 3),包括构成蛋白分子内腔所需的天冬氨酸位点(A87)和2个组氨酸位点(A89,A121),以及其他保守的活性位点(A84,A213,A217,A218,A221,A223,A228)。5种同源MDS序列多重比对后相同的氨基酸位点达70个,几种植物MDS蛋白N端比大肠埃希菌Escherichia coli多出1段约80~90个氨基酸残基的蛋白序列,说明此段区域可能存在转运肽,ChloroP 1.1 Server程序推导EuMDS蛋白转运肽序列长为56个氨基酸残基,在去除转运肽序列后EuMDS成熟蛋白分子量为19.24 kD。

图 3 EuMDS氨基酸序列与同源序列的多重比对
Figure 3. Multi-alignment in deduced EuMDS amino acid sequence and homologous sequence
-
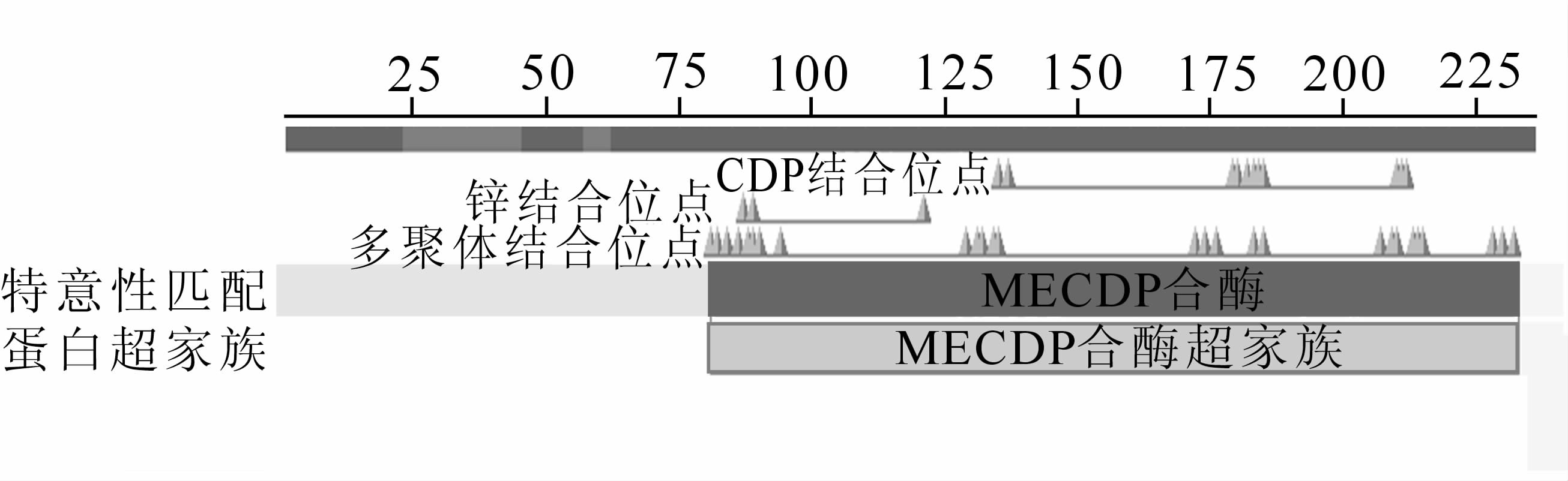
Predict Protein在线预测EuMDS蛋白二级结构中α-螺旋占40.25%,β-折叠占13.56%;螺环结构占46.19%,属于混合型结构(图 4)。保守结构域分析EuMDS结构域属MECDP合成酶蛋白家族,并包含锌离子(Zn2+)结合位点、CDP结合位点以及三聚体接触面等功能域。

图 4 EuMDS蛋白二级结构预测
Figure 4. Predicted secondary structure of the deduced protein of EuMDS

图 5 EuMDS蛋白保守结构域预测
Figure 5. Predicted conserved domains of deduced EuMDS protein
-
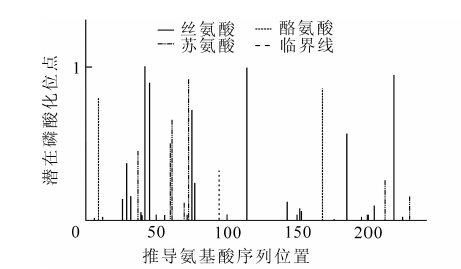
ExPaSy ScanProsite程序分析EuMDS基序类型分为5种,含有9个潜在的功能位点,包括1个蛋白激酶C磷酸化位点(SlR,42-44);3个酪蛋白激酶Ⅱ磷酸化位点(TavE,61-64;SdgD,114-117;SlgE,148-151);2个N端糖基化位点(NRSI,152-155;NLSA,189-192);2个N端豆蔻酰化位点(GAdpSV,195-200;GAasSV,214-219);1个酪氨酸激酶磷酸化位点(RlmhEag.Y,226-233)。NetPhos 2.0 server共预测出11个磷酸化位点,包括6个丝氨酸磷酸化位点(A42,A45,A75,A114,A185,A218),3个苏氨酸磷酸化位点(A60,A61,A73)以及2个酪氨酸磷酸化位点(A9,A168)(图 6)。

图 6 EuMDS蛋白翻译后磷酸化位点预测
Figure 6. Predicted phosphorylation sites of deduced protein of EuMDS
-
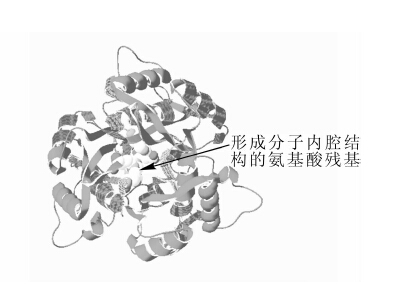
以拟南芥Arabidopsis thalana MDS蛋白(2 pmp)为模板对EuMDS蛋白同源建模,并利用Swiss Pdb Viewer 4.0.4对相应功能域进行标注,如图 7所示,EuMDS蛋白空间上由3个亚单位组成,三者相互围绕形成1个分子内腔结构,且结构内部有多个高度保守的氨基酸残基。ExPAsy structure assessment 程序评测推导的EuMDS蛋白模型QMEAN 6 得分为0.950,与模板蛋白序列的相似性为91.88%。

图 7 EuMDS三级结构预测
Figure 7. Predicted teriary structure of the deduced protein of EuMDS
-
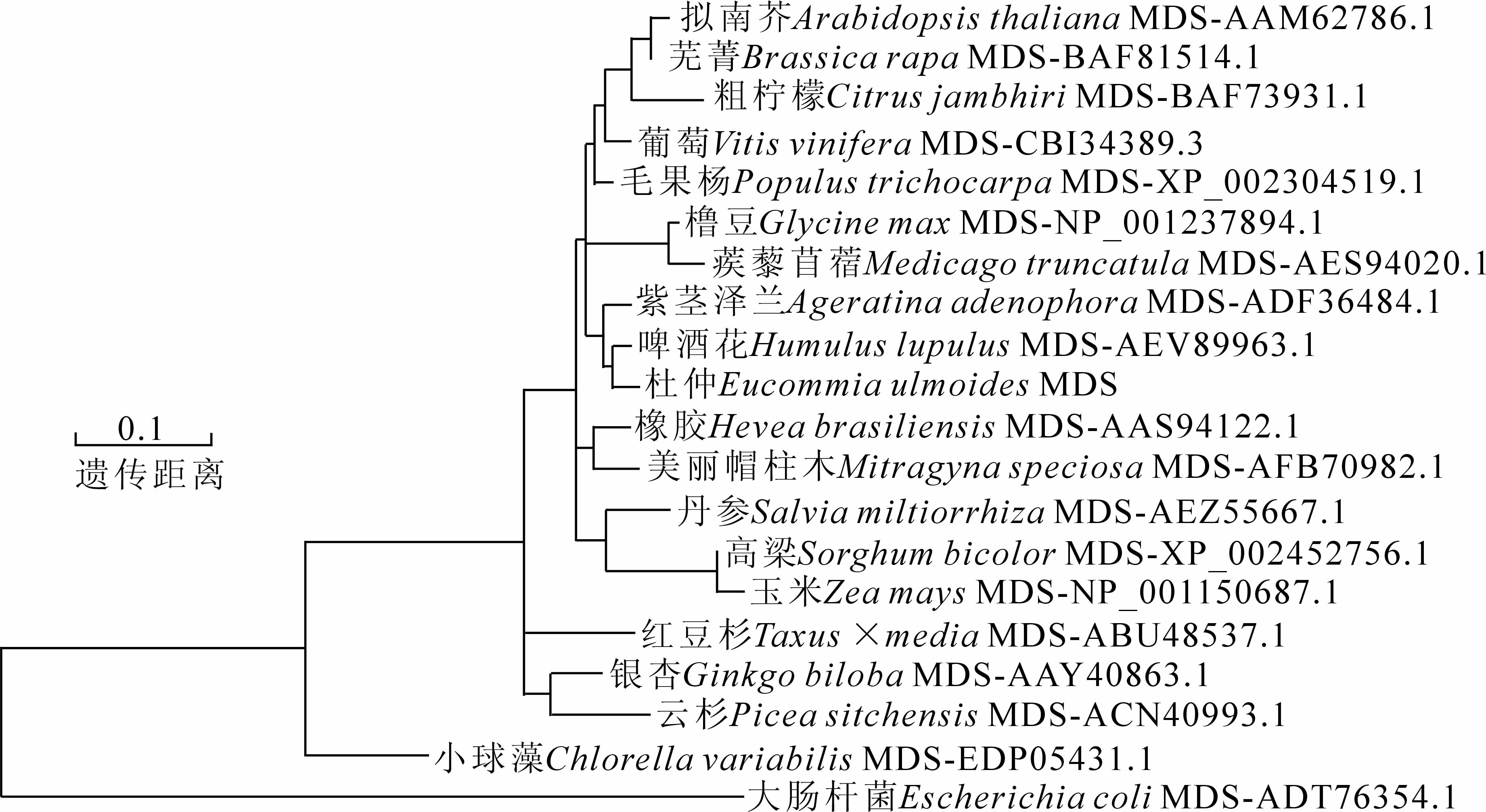
利用MEGA5中Clustal W法对20个物种MDS氨基酸序列进行比对,并用Neighbor-joining法构建了系统进化树(图 8)。结果表明:不同来源的MDS蛋白序列在进化上分属不同的分类群,植物MDS蛋白之间分类界限较为模糊,说明其进化关系较为复杂。EuMDS蛋白与啤酒花MDS蛋白间的亲缘关系最为接近,距离为0.032,其次为紫茎泽兰(0.039),美丽帽柱木Mitragyna speciosa(0.052),毛果杨(0.052),葡萄(0.052)和橡胶(0.066)。

图 8 MDS同源蛋白的系统进化树分析
Figure 8. Phylogenetic analysis in MDS homologous protein
-
从杜仲皮、叶中分离出十几种环烯醚萜类化合物,包括京尼平苷、京尼平苷酸、桃叶珊瑚苷、杜仲苷、筋骨草苷和车叶草酸等。这些活性物质具有丰富且独特的药理药效和保健功能[19-20],而高分子材料杜仲胶在海底电缆、器件装饰、绿色轮胎、医疗器械、消音减震、密封堵漏等领域发挥独特作用[21]。自杜仲萜类化合物开发利用以来,研究者们不断尝试各种方法以期提高产量。但是基于化学合成法需经一系列繁杂的催化反应,且成本高、得率低、毒性大,随基因工程方法不断发展,于分子水平上进行萜类合成的人工调控,对目的化合物定向生产和树种定向培育具有重要意义。MEP合成途径相对于MVA途径更广泛地存在于自然界中,MEP途径相关基因对萜类合成的生物调节机制日益引起萜类研究关注。基于生物信息学分析的结果表明EuMDS蛋白具有植物MDS典型功能位点、基序及结构域,意味着所克隆EuMDS是高等植物MDS基因家族新成员,可为杜仲萜类代谢工程候选基因筛选提供基础信息,但对其基因功能的判定有待深入研究。
植物MDS蛋白晶体结构首先从拟南芥中得到解析,发现与细菌类存在一定差别,主要在于由3个MDS分子亚单位相互围绕形成的分子内腔(molecular cavity)结构不同,细菌MDS蛋白分子内腔适合与IPP结合,并被推测发挥着负反馈调节功能,而拟南芥分子内腔中几个高度保守的氨基酸残基阻碍与二磷酸键结合,两者效应位点的差异意味着细菌和植物MDS蛋白在MEP合成途径中具有不同的调节机制。但也有一些实验推断与之相悖,缺失MDS的大肠埃希菌突变体转入拟南芥MDS会使其致死表形得到转变,因而推断细菌与植物MDS催化机制类似[22-23]。目前PDB数据库已注册53种MDS蛋白晶体模型,包括类鼻疽杆菌Burkholderia pseudomallei,栖热菌Thermus thermophilu,大肠埃希菌等51种细菌类型和2种真核生物类型。本研究通过Swiss-model同源建模法预测EuMDS蛋白三维结构与已解析拟南芥MDS蛋白空间特征吻合度很高,为蛋白功能预测以及进一步解析杜仲MDS催化机制提供初步参考。
Cloning and sequence of 2-C-methyl-D-erythritol-2,4-cyclodiphosphate synthase gene cDNA from Eucommia ulmoides
-
摘要: 2-甲基-D-赤藓糖醇-2,4-环焦磷酸合酶(MDS)基因曾被认为是调控植物2-甲基-D-赤藓糖醇4-磷酸(MEP)途径的一个关键节点。为解析杜仲MDS基因序列信息和预测基因功能,以叶片cDNA为模板,采用反转录-聚合酶链式反应(RT-PCR)及cDNA末端快速扩增(RACE)技术分离出杜仲MDS基因的cDNA克隆,并通过一系列生物信息学方法进行序列分析。结果表明:EuMDS基因cDNA全长976 bp,5'端非编码区长119 bp,3'端非编码区长146 bp,编码236个氨基酸。推导EuMDS氨基酸序列中包含转运肽序列(A1~A56)以及多个植物MDS蛋白保守的功能位点(A84,A87,A89,A121,A213,A217,A221,A223,A228)。推导EuMDS蛋白二级结构中α-螺旋占40.3%,β-折叠占13.6%,螺环结构占46.2%。推导EuMDS蛋白三级结构由3个亚单位组成,并相互围绕形成1个分子内腔。系统进化分析表明EuMDS蛋白与啤酒花MDS蛋白亲缘关系最为接近。预测所克隆的EuMDS基因在杜仲萜类生物合成中发挥重要功能。
-
关键词:
- 林木育种学 /
- 杜仲 /
- 2-甲基-D-赤藓醇-2,4-环焦磷酸合酶(MDS) /
- 基因 /
- 序列分析
Abstract: 2-C-methyl-D-erythritol-2,4-cyclodiphosphate synthase (MDS) gene had been regarded as a key regulating plot in plant 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway.To dissect the MDS gene sequence information and predict the gene function of Eucommia ulmoides,the homologous MDS gene cDNA was isolated from leaves by the reverse transcription polymerase chain reaction (RT-PCR) and rapid amplification of cDNA ends (RACE) techniques, and the sequence analysis was also conducted by series of bioinformatic methods.Results showed that and the full-length cDNA of EuMDS was 976 bp, including a 5'non-coding region of 119 bp and a 3'non-coding region of 146 bp with 236 amino acids encoded.The transit peptide sequence (A1-A56) and multiple conserved functional sites (A84, A87, A89, A121, A213, A217, A221, A223, and A228) of plant MDS enzyme were found in the deduced coding sequence of EuMDS.The secondary structure of the EuMDS protein was predicted with proportions of α-helix to 40.3%, β-sheet to 13.6%, and loop/coil to 46.2%.The calculated protein tertiary structure of EuMDS was exhibited as a molecular cavity formed by three subunits.Phylogenetic analysis revealed that the evolutionary relationship of EuMDS protein was closest to the Humulus lupulus MDS protein.It was suggested that the cloned EuMDS exert an important function in Eucommia ulmoides terpene biosysthesis.-
Key words:
- forest tree breeding /
- Eucommia ulmoides /
- MDS /
- gene /
- sequence analysis
-
图 1 EWlffiS基因RT-PCR扩增的琼脂糖凝胶电泳图
Figure 1 Agarose gel electrophoresis of RT-PCR amplification of EuMDS
图 2 EuMDS全长cDNA序列及推导的氨基酸序列
Figure 2 Full-length sequence of cDNA and deduced amino acid sequence of EuMDS
图 3 EuMDS氨基酸序列与同源序列的多重比对
Figure 3 Multi-alignment in deduced EuMDS amino acid sequence and homologous sequence
图 4 EuMDS蛋白二级结构预测
Figure 4 Predicted secondary structure of the deduced protein of EuMDS
图 6 EuMDS蛋白翻译后磷酸化位点预测
Figure 6 Predicted phosphorylation sites of deduced protein of EuMDS
-
[1] MICHEL R.The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria,algae and higher plants[J].Nat Prod Rep,1999,16:565-574. [2] HENRY M M.Enzymes of the mevalonate pathway of isoprenoid biosynthesis[J].Arch Biochem Biophys,2011,505(2):131-143. [3] LAURIS E K,CHARLES S B,WILLIAM N H.Structure of 2C-methyl-D-erythritol 2,4-cyclodiphosphate synthase:An essential enzyme for isoprenoid biosynthesis and target for antimicrobial drug development[J].PNAS,2002,99(10):6591-6596. [4] STEFAN S,JOHANNES K,JURAITHIP W,et al.Structure of 2C-methyl-D-erythritol-2,4-cyclodiphosphate synthase involved in mevalonate independent biosynthesis of isoprenoids[J].J Mol Biol,2002,316:79-88. [5] BERTRAND V,MARTINE C,AUDREY O,et al.Cloning and expression of cDNAs encoding two enzymes of the MEP pathway in Catharanthus roseus[J].Biochem Biophys Acta,2000,1517(1):159-163. [6] 刘万宏.紫杉醇前体合成途径2个关键酶基因克隆和分析[D].重庆:西南大学,2008. LIU Wanhong.Cloning and Characterization of Two Key Enzyme Genes Involved in the Biosynthesis Pathway of Taxol precursors[D].Chongqing:Southwest University, 2008. [7] 彭梅芳,阳义健,杨春贤,等.银杏IspF基因的克隆与功能分析[J].林业科学,2008,44(10):49-54. PENG Meifang, YANG Yijian,YANG Chunxian,et al.Cloning and functional analysis of a new IspF gene from Ginkgo biloba[J].Sci Silv Sin, 2008,44(10):49-54. [8] 李芳东,杜红岩.杜仲[M].北京:中国中医药出版社,2001. [9] 朱峰,岳红,祖恩峰,等.新型功能材料杜仲胶的研究与应用[J].安徽大学学报:自然科学版,2005,29(3):89-93. ZHU Feng, YUE Hong, ZHU Enfeng, et al.The study and appfication of Eucommia ulmoides gum[J].J Anhui Univ Nat Sci Ed, 2005, 29(3):89-93. [10] 李清泉,李志刚.杜仲胶与沥青共混性能试验[J].解放军理工大学学报:自然科学版,2007,8(2):176-179. LI Qingquan, LI Zhigang.Research on blending asphalt with Eucommia ulmoides gum[J].J PLA Univ Sci&Technol,2007,8(2):176-179. [11] 董娟娥,张靖.植物中环烯醚萜类化合物研究进展[J].西北林学院学报,2004,19(3):131-135. DONG Juan'e,ZHANG Jing.Advances in the research of iridoids occurring in plants[J].J Northwest For Univ, 2004, 19(3):131-135. [12] TAKAMURA C, HIRATA T, YAMAGUCHI Y, et al.Studies on the chemical constituents of green leaves of Eucommia ulmoides Oliv.[J].J Nat Med, 2007, 61:220-221. [13] 陈建.几种提取杜仲RNA方法的比较[J].林业科技开发,2007,21(5):19-21. CHEN Jian.Comparision of methods of RNA extraction with Eucommia ulmoides[J].China For Sci Technol, 2007, 21(5):19-21. [14] 周明兵,王红珍,赵德刚.杜仲叶和树皮总RNA的快速提取法[J].山地农业生物学报,2003,22(5):430-431. ZHOU Mingbing,WANG Hongzhen, ZHAO Degang.Fast extraction of RNA in barks and leaves of Eucommia ulmoides Oliver[J].J Mountain Agric Biol,2003,22(5):430-431. [15] JONES D T.Protein secondary structure prediction based on position-specific scoring matrices[J].J Mol Biol, 1999, 292(2):195-202. [16] EMANUELSSON O,NIELSEN H, von HEIJNE G.ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites[J].Protein Sci,1999,8:978-984. [17] ARNOLD K,BORDOLI L, KOPP J,et al.The SWISS-MODEL workspace:a web-based environment for protein structure homology modelling[J].Bioinformatics,2006,22:195-201. [18] TAMURA K,PETERSON D,PETERSON N,et al.MEGA5:Molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Mol Biol Evol,2011,28:2731-2739. [19] 王文明,庞晓萍,成军,等.杜仲化学成分研究概况(Ⅱ)[J].西北药学杂志,1998,13(2):60-62. WANG Wenming, PANG Xiaoping, CHENG Jun, et al.Studies on the lignans from Eucommia ulmoides(Ⅱ)[J].Northwest Pharm J,1998, 13(2):60-62. [20] 刘慧,刘仲华,张盛.杜仲中活性成分的研究进展[J].农产品加工学刊,2011(8):12-18. LIU Hui,LIU Zhonghua,ZHANG Sheng.Active composition of Eucommiae ulmoides Oliv.[J].Acad Period Farm Prod Process, 2011(8):12-18. [21] 张继川,薛兆弘,严瑞芳,等.天然高分子材料——杜仲胶的研究进展[J].高分子学报,2011(10):1105-1117. ZHANG Jichuan,XUE Zhaohong,YAN Ruifang,et al.Natural polymer material-recent studies on Eucommia ulmoides gum[J].Acta Polympus Sin,2011(10):1105-1117. [22] HSIEH M H, HOWARD M G.Functional evidence for the involvement of Arabidopsis IspF homolog in the nonmevalonate pathway of plastid isoprenoid biosynthesis[J].Planta,2006,223:779-784. [23] BARBARA M C,JORDI P G,MARIA B,et al.Biosynthesis of Isoprenoids in plants:Structure of the 2C-methyl-D-erithrytol-2,4-cyclodiphosphate synthase from Arabidopsis thaliana comparison with the bacterial enzymes[J].Protein Sci, 2007, 16:2082-2088. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.03.013
 点击查看大图
点击查看大图

计量
- 文章访问数: 4111
- HTML全文浏览量: 663
- PDF下载量: 430
- 被引次数: 0
