

A stress wave propagation velocity model of standing trees
-
摘要: 为了提高应力波技术在活立木无损检测应用领域的可行性,分析了应力波在健康树木中的传播规律,并建立了应力波传播速度数学模型。针对银杏Ginkgo biloba等不同树种的健康树木进行了多组实验,并利用树木断层成像技术对提出的应力波传播速度模型进行验证。结果表明:在健康树木中,方向角与传播方向速度vT和径向速度vR比值之间的关系为vT/vR-0.22+1,与所提出的理论模型吻合,方向角与应力波传播速度之间的线性回归模型拟合度较高,决定系数高于0.95。提出的应力波传播速度模型不受树种变化影响。图6表3参12Abstract: To improve the feasibility of nondestructive stress wave testing of standing trees, stress wave propagation in healthy trees was analyzed using linear regression analysis and a mathematical model of the stress wave propagation velocity was established. Several experiments were carried out for healthy trees of different species, and simultaneously the tomogram imaging technology was used to validate the proposed propagation velocity model of the stress wave. Experimental results of healthy trees showed that the linear regression relationship between the direction angle , the velocity (vT) for the propagation direction, and the radial velocity (vR) could be expressed as vT/vR-0.22 + 1, and the coefficient of determination R2>0.95, which demonstrated that the proposed mathematical model was correct. [Ch, 6 fig. 3 tab. 12 ref.]
-
Key words:
- wood science and technology /
- stress wave /
- velocity model /
- nondestructive testing
-
密码子是识别和传递生物体遗传信息、联系蛋白质与DNA之间的重要桥梁,在生物体遗传和变异中起着至关重要的作用[1]。编码同一氨基酸的不同密码子被称为同义密码子。由于基因突变和自然选择的影响,某些同义密码子在蛋白质翻译过程中往往被高频使用,被称为密码子的使用偏好性[2−3]。物种的生物学功能与密码子偏好性密切相关,密码子偏好性不仅可以影响生物编码基因的蛋白质合成速率和翻译速率[4],还会影响蛋白质结构、折叠程度和mRNA的合成[5]。研究表明:同一物种或亲缘关系相近的物种,具有相似的密码子偏好使用模式[6],通过分析物种的密码子偏好性可以衡量物种之间的基因表达量,进而探究物种之间亲属关系[7]。通过密码子偏好性的研究,能够更好地阐明物种进化过程中基因的表达规律[8],为利用基因工程技术改良物种目标基因提供参考依据[9]。
梁山慈竹Dendrocalamus farinosus属竹亚科Bambusoideae牡竹属Dendrocalamus,又名大叶竹和瓦灰竹,是中国西南地区重要的经济竹种[10],生长速度快,适应性强,竹笋效益高,属于优良的笋竹两用竹种,与硬头黄竹Bambusa rigida都属于竹编和制浆造纸的优质原料[11]。针对梁山慈竹叶绿体基因组密码子使用偏好性的研究鲜见报道。为了更好地挖掘和利用梁山慈竹的潜在经济价值,本研究以梁山慈竹叶绿体基因组序列为研究对象,分析其密码子偏好性使用模式,探究并总结其相关表达基因的密码子偏好性,以期分析影响梁山慈竹叶绿体基因组密码子偏好性的主要因素,并筛选出最优密码子,为后续梁山慈竹叶绿体基因工程改造等研究提供理论基础。
1. 材料与方法
1.1 叶绿体基因组序列的获取
根据GenBank登录号MZ681865.156在美国国家生物技术信息中心(NCBI)数据库中搜索并下载梁山慈竹叶绿体基因组序列,共有85条编码序列(CDS)。序列重复或小于300 bp会对密码子偏好性指标的测定产生影响[12]。对基因序列进行筛选,剔除序列长度小于300 bp且重复的序列,获取起始密码子为ATG,终止密码子为TAG、TGA和TAA的序列,最终获得51条CDS序列作为后续分析的样本序列。
1.2 密码子组成分析
运用CodonW1.4.2 (
http://sourceforge.net/projects/codonw )和EMBOSS (http://imed.med.ucm.es/EMBOSS/ )计算有效密码子数(ENC)、适应指数(CAI)、密码子偏性指数(CBI)、最优密码子频率(FOP)以及密码子第3位核苷酸A、T、C、G的含量(分别记为A3、T3、C3、G3)。利用ENC判断密码子偏好性程度,ENC>35说明密码子偏好性比较弱;反之,说明偏好性强[13]。通过CUSP软件分析并获得密码子鸟嘌呤(G)和胞嘧啶(C)所占的比率(GC比率)及GC平均比率(GCall),使用SPSS 25.0软件对梁山慈竹密码子各位置的GC比率与ENC进行相关分析。1.3 相对同义密码子使用度分析
运用CodonW 1.4.2对同义密码子相对使用度(RSCU)进行分析,即该密码子的实际使用频率与其理论使用频率的比值[14]。当RSCU大于1时,同义密码子中偏好使用该密码子,被称为高频密码子;当RSCU等于1时,密码子无偏好性;当RSCU小于1时,密码子使用偏好性较弱[15]。
1.4 中性绘图分析
中性绘图分析是对影响密码子使用偏好性的关键因素进行分析,X轴为GC3,Y轴为GC1和GC2的平均值,绘制二维散点图对GC3和GC12 (各基因 GC1和GC2的平均值)的相关性进行分析(GC1、GC2、GC3分别代表第1、2、3位密码子的GC比例)。若回归系数接近1,代表GC3和GC12显著相关,碱基组成没有差异,说明突变是决定密码子偏好性的主要因素;若回归系数接近0,则代表自然选择是主要因素。
1.5 ENC-plot绘图分析
ENC-plot绘图分析表现密码子的使用偏好性受到突变和自然选择的影响程度。使用Python 3.7进行ENC-plot绘图分析,构建散点图,横纵坐标分别为GC3、ENC,并绘制ENC的标准曲线。基因位点靠近或在标准曲线上,表明突变是决定密码子偏好性的主要因素,若基因位点和标准曲线距离很大,则说明偏好性主要由自然选择决定。
1.6 PR2-plot偏倚分析
PR2-plot分析表明基因中密码子的第3位碱基的构成情况。计算密码子碱基中第3位上4种碱基A、T、C、G比例,G3/(G3+C3)为X轴,A3/(A3+T3)为Y轴,绘制PR2-plot散点图,中心点为碱基比例A=T、C=G时的值,代表处于此区域的密码子并无使用偏好性[16]。
1.7 最优密码子分析
将51条基因升序排列后的ENC前后两端10%的基因建立高、低表达基因库。通过CodonW软件计算2个表达库中密码子的RSCU和ΔRSCU,同时满足高频密码子(RSCU>1)和高表达密码子(ΔRSCU≥0.08)的为最优密码子[17]。
1.8 梁山慈竹和其他几种生物密码子偏好性比较分析
在Codon Usage Database (
http://www.kazusa.or.jp/codon/ )下载异源表达宿主和植物代表类群,包括巨龙竹D. farinosus、粉麻竹D. sinicus、小叶龙竹D. pulverulentus、硬头黄竹、大肠埃希菌Escherichia coli、烟草Nicotiana tabacum、拟南芥Arabidopsis thaliana和酿酒酵母Saccharomyces cerevisiae等物种基因组密码子的使用频率,与梁山慈竹基因组密码子使用频率比值进行比较分析,当梁山慈竹密码子使用频率比其他生物的比值≥2.0或≤0.5时,说明该物种与梁山慈竹的同义密码子的使用偏好性差异较大,当比值不在上述范围内时,表明这2个物种对该密码子的偏好性较接近。1.9 对应分析
将叶绿体基因如表1所示进行功能分类,使用CodinW软件,选择对应分析计算样本中各个基因的RSCU,将分析结果分布在59维向量空间中,分析指标间的对应性。
表 1 梁山慈竹叶绿体基因结构分析Table 1 Structural analysis of the choroplast genome of D. farinosus基因分类 基因分组 基因名称 光合系统基因 光系统Ⅰ基因 psaA、psaB、psbA、psbC、psbD、psbB 光系统Ⅱ基因 petA、petB、petD 细胞色素b/f复合体基因 atpA、atpB、atpE、atpF、atpI 三磷酸腺苷合成酶基因 ndhA、ndhB、ndhC、ndhD、ndhE、ndhF、ndhG、ndhH、ndhI、ndhJ、ndhK 遗传系统基因 烟酰胺腺票吟二核甘酸氧化还原酶基因 rbcL 二磷酸核酮糖羧化酶大亚基基因 rpoA、rpoB、rpoC1、rpoC2 RNA聚合酶亚基基因 rps2、rps3、rps4、rps7、rps8、rps11、rps12、rps14、rps18 核糖体蛋白小亚基基因 rpl2、rpl14、rpl16、rpl20、rpl22 其他基因 成熟酶K基因 matK 膜蛋白基因 cemA 细胞色素合成基因 ccsA 酪蛋白分解蛋白酶基因 clpP 未知功能基因 假定叶绿体阅读框 ycf2、ycf3、infA 2. 结果与分析
2.1 密码子的碱基组成分析
分析梁山慈竹叶绿体基因组CDS序列的碱基组成:梁山慈竹的4种碱基所对应的同义密码子的第3位碱基比例 (T3s、A3s、C3s、G3s)分别为45.28%、42.07%、18.13%、17.96%,T3s和A3s远高于G3s和C3s,表明梁山慈竹叶绿体基因组密码子的第3位碱基以A/U结尾为主。梁山慈竹的ENC为50.40,CAI为16.6%,第3位同义密码子的GC比率 (GC3S)为28.1%,表明其叶绿体基因组密码子偏好性较弱。
梁山慈竹叶绿体基因组密码子的GC平均比率为39.48%,且GC1 (47.69%)>GC2 (39.70%)>GC3 (31.05%)。ENC为 39.04~61.00,均值为49.51,GC比率在基因密码子上并没有均匀分布(表2)。ENC和密码子3个位置GC比率的相关分析(表3)结果发现:ENC与GC3比率显著相关,与GC1、GC2不显著相关,说明密码子使用偏好性形成过程中GC3的影响作用大于GC1、GC2。
表 2 梁山慈竹叶绿体基因组各基因密码子相关参数统计Table 2 Statistics of codon related parameters of various genes in the chloroplast genome of D. farinosus基因 GC比率/% ENC CAI FOP 基因 GC比率/% ENC CAI FOP GC GC1 GC2 GC3 GC GC1 GC2 GC3 rps12 41.87 52.00 47.20 26.40 44.85 0.140 0.341 rps18 33.53 34.50 39.77 26.32 39.04 0.147 0.333 psbA 42.56 49.72 42.94 35.03 41.33 0.313 0.532 rpl20 36.11 38.33 40.83 29.17 50.97 0.112 0.298 matK 34.44 40.82 32.42 30.08 49.49 0.166 0.329 clpP 43.01 52.53 38.25 38.25 52.37 0.175 0.337 psbD 44.44 53.39 43.50 36.44 48.99 0.242 0.456 psbB 44.01 54.42 45.97 31.63 50.73 0.190 0.380 psbC 44.66 53.59 44.73 35.63 48.91 0.183 0.386 petB 41.06 48.93 41.20 33.05 47.31 0.191 0.333 rpoB 39.19 49.81 38.01 29.74 49.69 0.153 0.353 petD 40.37 50.93 39.13 31.06 49.46 0.161 0.305 rpoC1 39.87 49.93 38.07 31.63 52.77 0.156 0.347 rpoA 37.06 46.18 35.59 29.41 49.94 0.151 0.311 rpoC2 38.95 49.01 36.64 31.18 52.29 0.154 0.333 rps11 43.52 50.69 56.25 23.61 44.33 0.174 0.396 rps2 38.40 40.51 40.93 33.76 52.55 0.168 0.338 infA 40.35 43.86 35.96 41.23 61.00 0.181 0.409 atpI 38.84 47.58 36.29 32.66 50.55 0.163 0.353 rps8 36.50 41.61 41.61 26.28 46.62 0.122 0.374 atpF 38.27 47.62 35.45 31.75 53.17 0.147 0.353 rpl14 38.71 54.84 37.10 24.19 51.90 0.181 0.392 atpA 42.06 56.01 39.96 30.12 49.96 0.182 0.385 rpl16 44.76 52.14 53.57 28.57 39.41 0.115 0.354 rps14 39.42 39.42 46.15 32.69 41.73 0.135 0.384 rps3 33.47 43.75 31.67 25.00 48.03 0.193 0.402 psaB 41.81 48.71 43.13 33.61 49.34 0.172 0.350 rpl22 37.56 41.33 36.67 34.67 47.48 0.188 0.415 psaA 43.68 51.80 43.28 35.95 52.07 0.198 0.373 rpl2 44.56 51.77 48.58 33.33 53.33 0.143 0.361 ycf3 39.69 47.40 38.15 33.53 55.45 0.156 0.343 ndhB 38.16 42.07 39.33 33.07 46.71 0.156 0.348 rps4 37.13 47.52 37.13 26.73 49.59 0.169 0.386 rps7 39.49 49.68 45.22 23.57 48.31 0.164 0.373 ndhJ 39.38 49.38 36.88 31.88 51.48 0.176 0.356 ndhF 34.19 37.84 38.92 25.81 46.19 0.144 0.321 ndhK 38.60 41.70 43.72 30.36 51.91 0.159 0.329 ccsA 33.64 33.74 41.10 26.07 45.60 0.152 0.307 ndhC 39.67 50.41 36.36 32.33 48.75 0.177 0.345 ndhD 36.19 40.72 36.93 30.94 48.98 0.133 0.314 atpE 42.51 52.17 39.13 36.23 59.51 0.167 0.405 ndhE 33.33 41.18 32.35 26.47 59.06 0.144 0.316 atpB 42.62 53.91 41.68 32.26 47.43 0.192 0.381 ndhG 34.46 44.07 32.77 26.55 45.77 0.125 0.250 rbcL 44.14 57.11 43.93 31.38 50.19 0.271 0.454 ndhI 34.99 37.57 38.67 28.73 52.09 0.171 0.345 ycf4 41.22 48.39 39.78 35.48 47.14 0.162 0.385 ndhA 33.98 42.42 36.36 23.14 44.35 0.140 0.321 cemA 33.62 41.99 27.71 31.17 55.91 0.176 0.342 ndhH 37.82 50.76 34.77 27.92 49.95 0.155 0.322 petA 40.29 53.58 35.2 32.09 51.12 0.155 0.331 表 3 梁山慈竹叶绿体基因组中各基因参数的相关性分析Table 3 Correlation analysis of various gene parameters in the chloroplast genome of D. farinosus参数 GC1 GC2 GC3 ENC CAI CBI FOP GC3s GC GC1 1 GC2 0.300* 1 GC3 0.265 −0.009 1 ENC 0.142 −0.425** 0.389** 1 CAI 0.409** 0.076 0.370** 0.012 1 CBI 0.438** 0.272 0.322* −0.092 0.774** 1 FOP 0.402** 0.312* 0.341* −0.064 0.797** 0.965** 1 GC3s 0.271 −0.029 0.946** 0.445** 0.330* 0.330* 0.370** 1 GC 0.814** 0.673** 0.525** 0.010 0.407** 0.512** 0.518** 0.499** 1 说明: *表示显著相关 (P<0.05);**表示极显著相关 (P<0.01)。 2.2 相对同义密码子使用度分析
梁山慈竹叶绿体基因组中共包含18110个密码子(表4),总计编码20个氨基酸,密码子数为12~705个,其中密码子UGA共有12个,密码子含量最多的是编码谷氨酸的GAA,共有705个。梁山慈竹叶绿体基因组蛋白编码序列RSCU分析表明:氨基酸含量较高的有亮氨酸(Leu)和精氨酸(Arg),均为6个密码子编码,编码精氨酸的是UUA、UUG、CUU、CUC、CUA和CUG;编码亮氨酸的有AGA、AGG、CGU、CGC、CGA和CGG;除此之外,蛋氨酸(Met)和色氨酸(Trp)均只有1个密码子编码,分别是AUG和UGG,其余氨基酸密码子编码个数分别为2~4个。
表 4 梁山慈竹叶绿体基因组蛋白编码序列RSCU分析Table 4 RSCU of protein coding region in the chloroplast of D. farinosus氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU Phe UUU* 644 1.29 Tyr UAU* 532 1.59 Ser UCU* 343 1.58 Cys UGU* 151 1.53 Phe UUC 351 0.71 Tyr UAC 137 0.41 Ser UCC* 260 1.19 Cys UGC 47 0.47 Leu UUA* 634 1.94 TER UAA* 28 1.56 Ser UCA* 222 1.02 Arg AGA* 322 1.75 Leu UUG* 362 1.11 TER UAG 14 0.78 Ser UCG 119 0.55 Arg AGG 119 0.64 Leu CUU* 420 1.29 TER UGA 12 0.67 Ser AGU* 273 1.25 Arg CGU* 261 1.41 Leu CUC 138 0.42 Trp UGG* 328 1.00 Ser AGC 89 0.41 Arg CGC 95 0.51 Leu CUA 295 0.90 Gln CAA* 477 1.53 Thr ACU* 403 1.68 Arg CGA* 234 1.27 Leu CUG 107 0.33 Gln CAG 148 0.47 Thr ACC 181 0.75 Arg CGG 76 0.41 Ile AUU* 740 1.48 Glu GAA* 705 1.46 Thr ACA* 259 1.08 Gly GGU* 421 1.24 Ile AUC 295 0.59 Glu GAG 263 0.54 Thr ACG 116 0.48 Gly GGC 145 0.43 Ile AUA 461 0.92 Lys AAA* 647 1.44 Ala GCU* 493 1.73 Gly GGA* 538 1.58 Met AUG* 416 1.00 Lys AAG 253 0.56 Ala GCC 172 0.60 Gly GGG 259 0.76 Val GUU* 382 1.47 Asp GAU* 522 1.54 Ala GCA* 343 1.20 Pro CCU* 286 1.48 Val GUC 126 0.49 Asp GAC 155 0.46 Ala GCG 135 0.47 Pro CCC* 196 1.01 Val GUA* 390 1.50 His CAU* 311 1.47 Asn AAU* 528 1.48 Pro CCA* 209 1.08 Val GUG 139 0.54 His CAC 112 0.53 Asn AAC 187 0.52 Pro CCG 84 0.43 说明:*表示RSCU大于1的高频密码子。 梁山慈竹叶绿体基因组RSCU大于1的密码子数目为34个(分别为UUU、UUA、UUG、CUU、AUU、AUG、GUU、GUA、UCU、UCC、UCA、AGU、ACU、ACA、GCU、GCA、AAU、UAU、UAA、UGG、CAA、GAA、AAA、GAU、CAU、UGU、AGA、CGU、CGA、GGU、GGA、CCU、CCC和CCA),即筛选出了34个高频密码子,其中以A、U、C、G结尾的密码子分别有13、16、2和1个,这说明密码子偏好以A和U结尾,RSCU较高的3个密码子分别为UUU (1.94)、CUA (1.73)和UCU (1.75)。
2.3 中性绘图分析
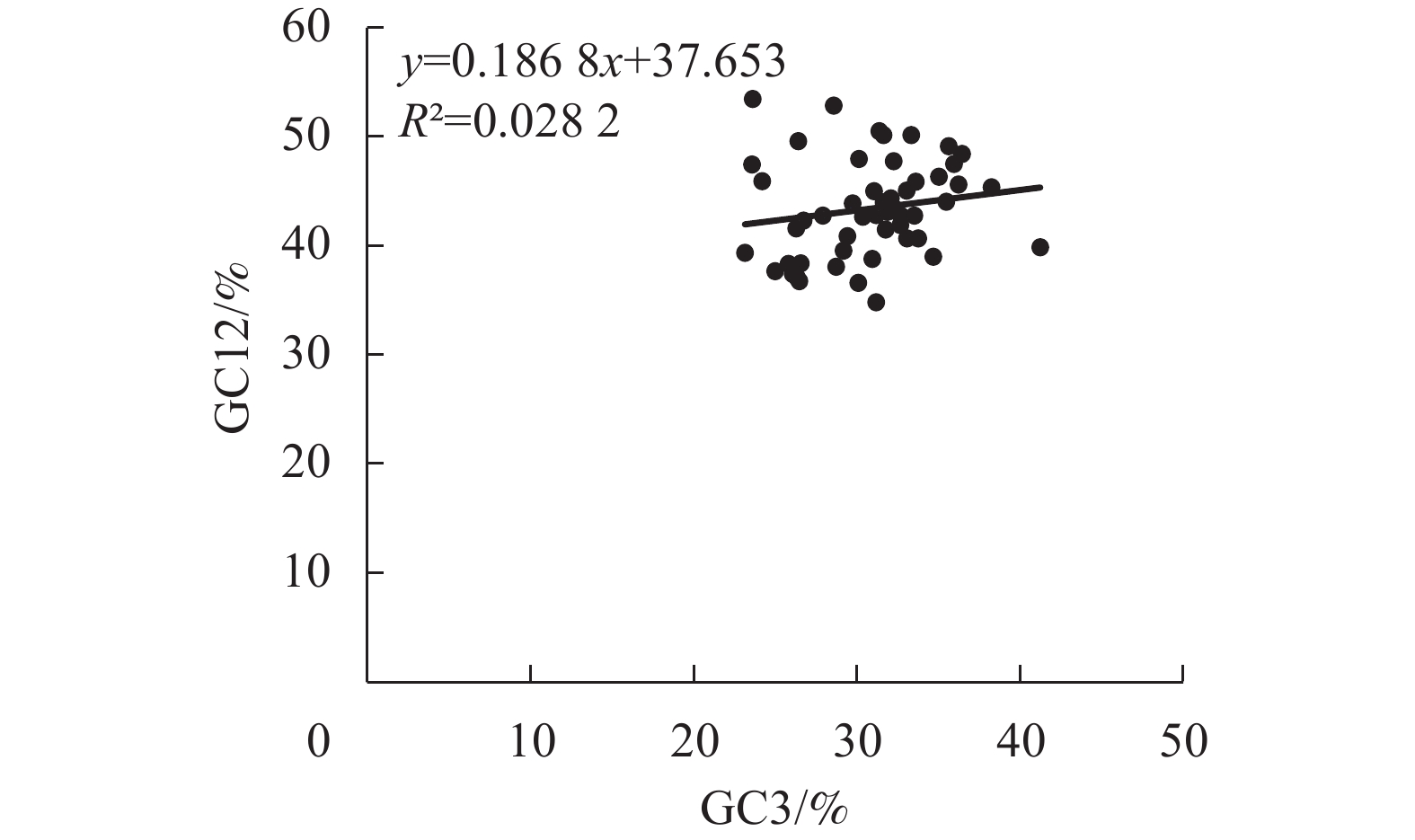
中性绘图分析量化自然选择和突变压力之间的关系,阐明3个密码子位置之间的联系。结果表明:横坐标GC3的数值为23.14%~41.23%,纵坐标GC12的数值为39.04%~61.00% (图1)。梁山慈竹的Pearson相关系数为0.17,呈正相关关系,数据拟合后的回归系数为0.1868,决定系数(R2)较小,为0.0282,GC12和GC3的相关性不显著,说明其叶绿体基因组密码子偏好性受自然选择影响较大。
2.4 ENC-plot绘图分析
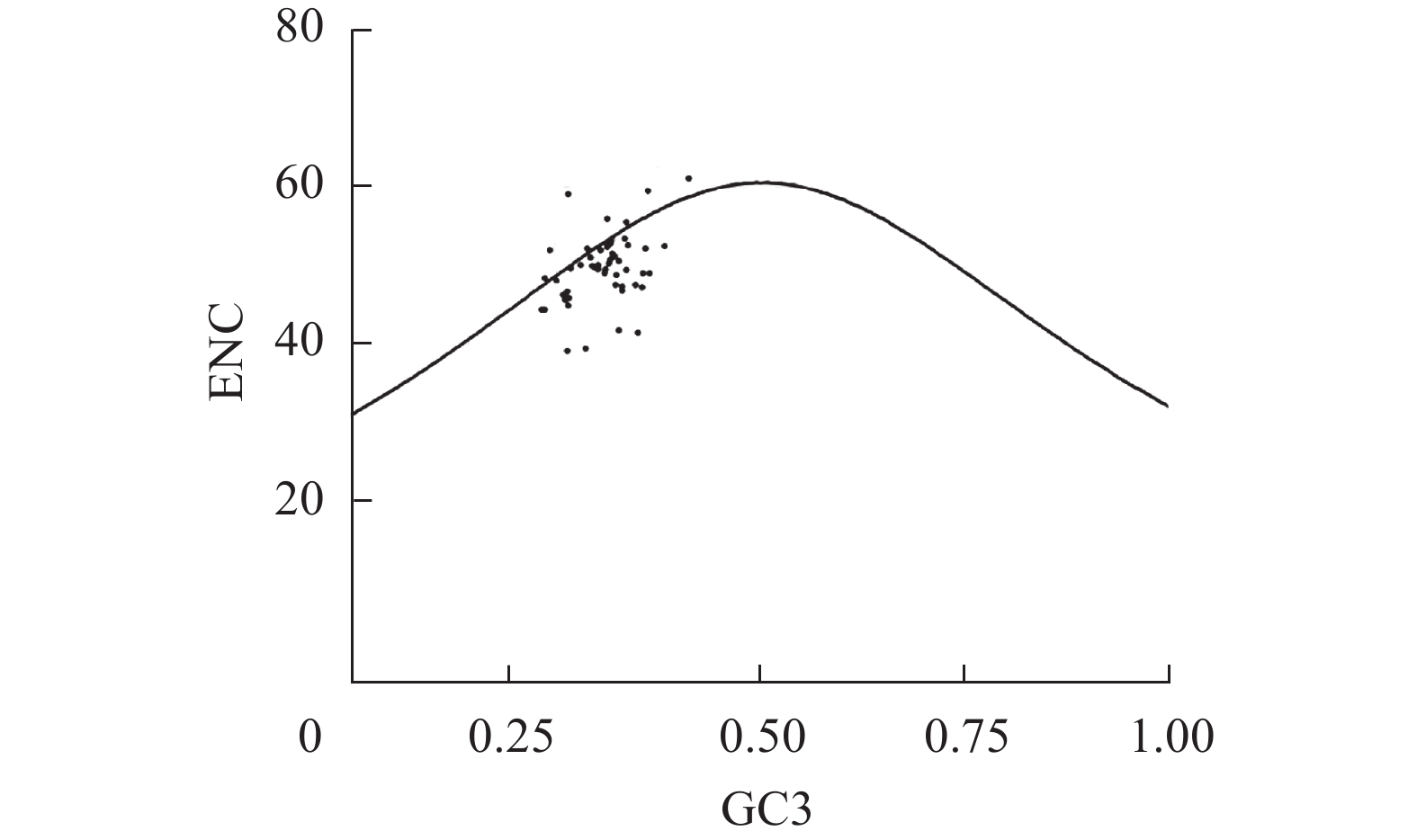
图2显示:ENC分布并不紧密,少量分布在标准曲线附近,还有个别分布在标准曲线上侧,位点的ENC均大于35,与预期ENC值有差距。说明梁山慈竹密码子偏好性较弱且自然选择和突变都对其偏好性有影响。由于落在标准曲线下方的基因点数量比较多,所以梁山慈竹基因组密码子使用偏好性主要受自然选择的影响。
2.5 PR2-plot绘图分析
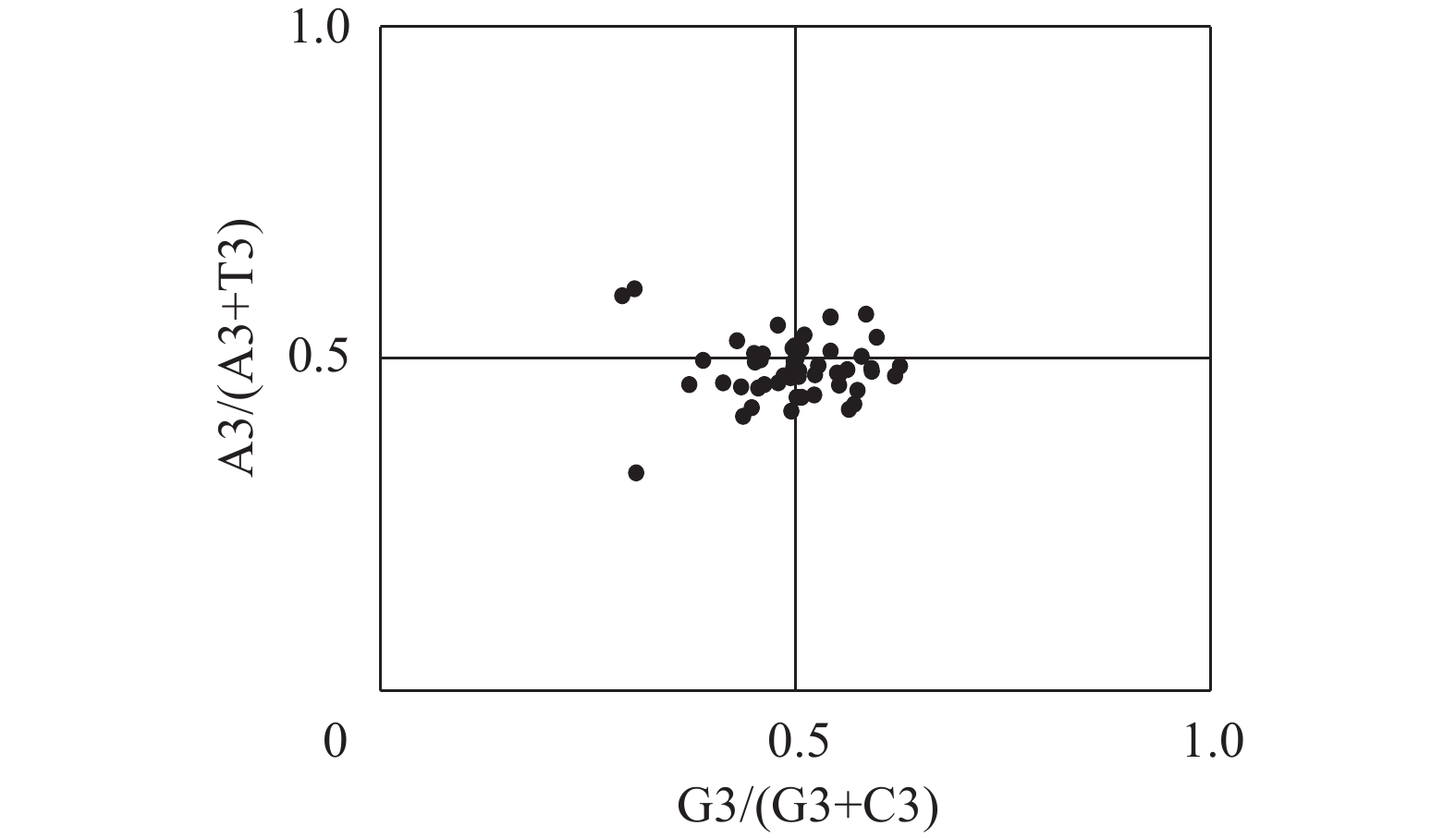
图3显示:基因位点在平面图4个区域内分布并不均匀,在A3/(A3+T3)<0.5和G3/(G3+C3)>0.5区域范围内分布最多。表明第3位碱基使用频率为:T>A、G>C,梁山慈竹叶绿体基因组密码子的第3位碱基在选择上具有偏好性,同时说明其密码子使用偏好性主要受自然选择的影响。
2.6 最优密码子的确定
对梁山慈竹的ENC进行升序排列,前10%为高表达基因,即rps18、rpl16、psbA、rps14、rps11,后10%为低表达基因,即 ycf3、cemA、ndhE、atpE、infA。梁山慈竹的RSCU和ΔRSCU表明(表5):梁山慈竹叶绿体基因组有32个高频密码子,筛选出GCA、GCU等25个高表达密码子,最终确定18个密码子作为梁山慈竹叶绿体基因组的最优密码子,分别为UAA、GCA、GCU、UUC、GGU、AAA、CUU、UUA、CCA、CCU、CAA、AGA、CGU、AGU、UCC、ACU、GUA、GUU。其中16个以A/U结尾,2个以C结尾。
表 5 梁山慈竹叶绿体基因组各氨基酸的RSCU分析及最优密码子分析Table 5 RSCU analysis and optimal codon analysis of amino acids in chloroplast genome of D. farinosus氨基酸 密码子 基因组
RSCU高表达
RSCU低表达
RSCUΔRSCU 氨基酸 密码子 基因组
RSCU高表达
RSCU低表达
RSCUΔRSCU Ter UAA*** 1.560 0 1.800 0 1.200 0 0.600 0 Met AUG 1.000 0 1.000 0 1.000 0 0 UAG 0.780 0 0.600 0 1.200 0 −0.600 0 Asn AAC* 0.520 0 0.893 6 0.625 0 0.268 6 UGA 0.670 0 0.600 0 0.600 0 0 AAU 1.480 0 1.106 4 1.375 0 −0.268 6 Ala GCA** 1.200 0 1.200 0 0.734 7 0.465 3 Pro CCA** 1.080 0 0.800 0 0.500 0 0.300 0 GCC 0.600 0 0.457 1 0.653 1 −0.196 0 CCC 1.010 0 0.800 0 1.166 7 −0.366 7 GCG 0.470 0 0.228 6 0.734 7 −0.506 1 CCG 0.430 0 0.444 4 1.000 0 −0.555 6 GCU* 1.730 0 2.114 3 1.877 6 0.236 7 CCU*** 1.480 0 1.955 6 1.333 3 0.622 3 Cys UGC** 0.470 0 0.400 0 0 0.400 0 Gln CAA* 1.530 0 1.500 0 1.368 4 0.131 6 UGU 1.530 0 1.600 0 2.000 0 −0.400 0 CAG 0.470 0 0.500 0 0.631 6 −0.131 6 Asp GAC* 0.460 0 0.500 0 0.411 8 0.088 2 Arg AGA* 1.750 0 1.723 4 1.534 9 0.188 5 GAU 1.540 0 1.500 0 1.588 2 -0.088 2 AGG 0.640 0 0.319 1 0.837 2 −0.518 1 Glu GAA 1.460 0 1.189 2 1.578 9 −0.389 7 CGA 1.270 0 1.276 6 1.395 3 −0.118 7 GAG** 0.540 0 0.810 8 0.421 1 0.389 7 CGC 0.510 0 0.319 1 0.837 2 −0.518 1 Phe UUC** 1.290 0 1.041 7 0.650 0 0.391 7 CGG 0.410 0 0.319 1 0.279 1 0.040 0 UUU 0.710 0 0.958 3 1.350 0 −0.391 7 CGU*** 1.410 0 2.042 6 1.116 3 0.926 3 Gly GGA 1.580 0 1.253 7 1.818 2 −0.564 5 Ser AGC 0.410 0 0.384 6 0.470 6 −0.086 0 GGC 0.430 0 0.417 9 0.484 8 −0.066 9 AGU** 1.250 0 1.846 2 1.411 8 0.434 4 GGG 0.760 0 0.119 4 0.363 6 −0.244 2 UCA 1.020 0 0.615 4 1.058 8 −0.443 4 GGU*** 1.240 0 2.209 0 1.333 3 0.875 7 UCC*** 1.190 0 1.769 2 0.941 2 0.828 0 His CAC** 0.530 0 0.941 2 0.571 4 0.369 8 UCG 0.550 0 0.153 8 0.705 9 −0.552 1 CAU 1.470 0 1.058 8 1.428 6 −0.369 8 UCU 1.580 0 1.230 8 1.411 8 −0.181 0 Ile AUA 0.920 0 0.850 7 0.949 4 −0.098 7 Thr ACA 1.080 0 1.181 8 1.176 5 0.005 3 AUC* 0.590 0 0.626 9 0.531 6 0.095 3 ACC 0.500 0 0.818 2 1.058 8 −0.240 6 AUU 1.480 0 1.522 4 1.519 0 0.003 4 ACG 0.480 0 0.363 6 0.588 2 −0.224 6 Lys AAA** 1.440 0 1.471 7 1.155 6 0.316 1 ACU** 1.680 0 1.636 4 1.176 5 0.459 9 AAG 0.560 0 0.528 3 0.844 4 −0.316 1 Val GUA*** 1.500 0 1.767 4 1.257 1 0.510 3 Leu CUA 0.900 0 0.833 3 1.295 5 −0.462 2 GUC 0.490 0 0 1.028 6 −1.028 6 CUC 0.420 0 0 0.545 5 −0.545 5 GUG 0.540 0 0.372 1 0.342 9 0.029 2 CUG 0.330 0 0.250 0 0.477 3 −0.227 3 GUU** 1.470 0 1.860 5 1.371 4 0.489 1 CUU* 1.290 0 1.333 3 1.227 3 0.106 0 Trp UGG 1.000 0 1.000 0 1.000 0 0 UUA*** 1.940 0 2.166 7 1.022 7 1.144 0 Tyr UAC** 0.410 0 0.521 7 0.166 7 0.355 0 UUG 1.110 0 1.416 7 1.431 8 −0.015 1 UAU 1.590 0 1.478 3 1.833 3 −0.355 0 说明: 高频密码子(RSCU>1.00)带下划线;*. ΔRSCU≥0.08;**. ΔRSCU≥0.3;***. ΔRSCU≥0.5; 加粗的密码子表示最优密码子。 2.7 梁山慈竹和其他几种生物密码子偏好性比较分析
将梁山慈竹基因组密码子使用频率与巨龙竹、粉麻竹、小叶龙竹、硬头黄竹、大肠埃希菌、烟草、拟南芥和酿酒酵母等物种的基因组密码子使用频率进行比较(图4)。结果显示:梁山慈竹与巨龙竹、粉麻竹、小叶龙竹和硬头黄竹的密码子使用频率为0.5~2.0,说明它们的密码子使用偏好性相似,推测具有亲缘关系的禾本科Gramineae牡竹属植物叶绿体基因组密码子偏好性相似;在大肠埃希菌、烟草、拟南芥和酿酒酵母的密码子使用比值中筛选≥2.0或≤0.5的密码子,分别有28和15、15、14个,表明梁山慈竹与这些物种在同义密码子的偏好性上有一定差异。
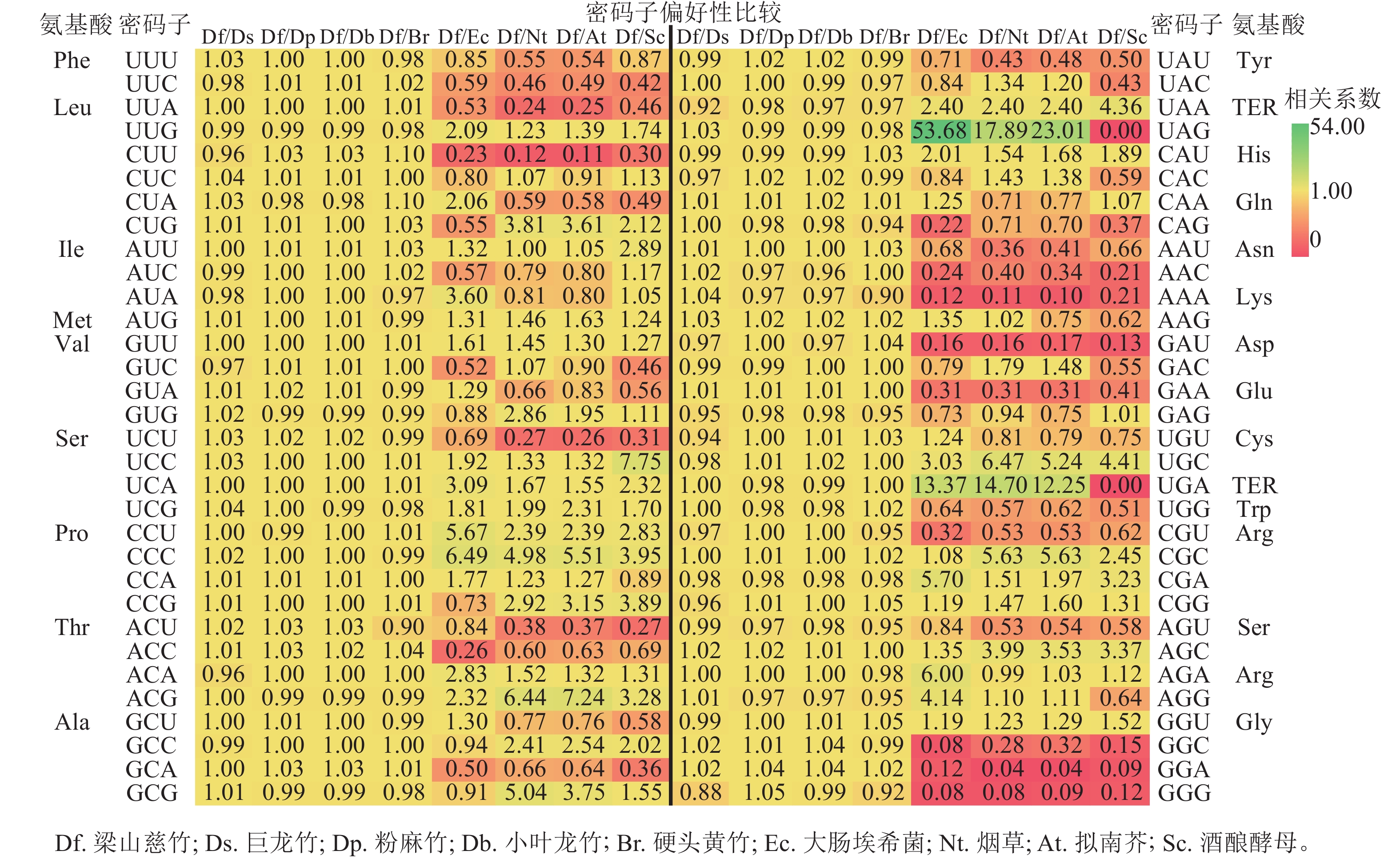 图 4 梁山慈竹与其他物种密码子偏好性比较Figure 4 Comparison of codon preference between D. farinosus and other species
图 4 梁山慈竹与其他物种密码子偏好性比较Figure 4 Comparison of codon preference between D. farinosus and other species2.8 对应分析
将梁山慈竹的51个叶绿体基因的基因功能分为光合系统基因、遗传系统基因、其他基因和未知功能基因四大类,在计算RSCU的基础上将各个基因分布到59维的向量空间。对应分析结果(图5)显示:前4个向量轴分别存在18.3%、16.8%、15.6%和15.4%的差异,前4向量轴累计差异为66.1%,4个轴对密码子均有不同程度的影响;第1轴的值大于其他轴,说明第1轴对梁山慈竹叶绿体基因组密码子偏好性的影响较大。对第1轴与CAI、CBI、FOP、ENC和GC3s等指数进行进一步的相关分析发现:梁山慈竹基因在第1轴上的坐标值与CAI (r=−0.001 7,P<0.01)、CBI (r=0.099 0,P<0.01)、FOP (r=0.083 0,P<0.01)、ENC (r=0.112 0,P<0.01)、GC3s (r=−0.145 0,P<0.01)间具有极显著的相关关系,其中CAI和GC3s第1轴具有负相关关系,表明基因组密码子的偏好性不止受单一因素的影响,自然选择、基因突变均有可能影响梁山慈竹基因组密码子使用偏好性[18]。
3. 讨论
本研究对梁山慈竹叶绿体基因组密码子进行使用偏好性分析,筛选出51条CDS序列,分析表明:GC1>GC2>GC3,密码子在3个位置上的分布并不均匀,密码子偏好使用以A或U结尾的碱基,且梁山慈竹叶绿体基因组的ENC均值为49.51,表明其叶绿体基因组密码子使用偏好性较弱。这与乳油木Vitellaria paradoxa[19]和二乔玉兰Magnolia soulangeana[20]等植物叶绿体基因组密码子偏好性相似。
对梁山慈竹叶绿体基因组密码子进行中性绘图、ENC-plot分析、PR2-plot分析和对应分析。在中性绘图分析中,回归系数为0.412 8,说明密码子偏好性更多受到自然选择的影响;在ENC-plot分析中,多数基因离标准曲线距离较远,实际ENC和预期ENC有差距,表明该部分基因的密码子偏好性主要受自然选择的影响;在PR2-plot绘图分析中,大部分基因位于平面图的右下方,即T>A、G>C,表明其密码子的使用更多受自然选择的影响。综上所述,影响梁山慈竹叶绿体基因组密码子偏好性的主要原因是自然选择。该研究结果与巨桉Eucalyptus grandi[21]、灰毛浆果楝Cipadessa cinerascens、酸枣Ziziphus jujuba var. spinosa[22]和云南油杉Keteleeria evelyniana[23]等叶绿体基因组密码子偏好性研究结果基本一致;但在对4种蔷薇科 Rosaceae果树[24]和银白杨Populus alba[25]的研究中发现:突变是影响密码子偏好性的主要因素。这说明密码子的使用偏好性受自然选择或基因突变因素影响。基于RSCU的对应分析表明:梁山慈竹的密码子使用变异原因除了突变和自然选择之外,还有其他的因素,这其中光合系统基因和遗传系统基因分布相对集中,各类基因密码子使用偏好性较为接近。该结论与木薯Manihot esculenta[26]和高山松Pinus densata[27]的研究结果一致。密码子使用频率比较结果显示:梁山慈竹与禾本科牡竹属的植物密码子偏好性相似,在基因选择外源系统表达时,可以选择密码子偏好性差异相对较小的酿酒酵母,在选择大肠埃希菌、烟草和拟南芥作为外源表达宿主时,需要根据密码子使用偏好性进行碱基优化,从而使基因在宿主体内更好地表达。
最优密码子分析表明:梁山慈竹叶绿体基因组有GCU、GAU以及GGU等18个最优密码子,最优密码子大部分以A或U结尾。该结果与抽筒竹Gelidocalamus tessellatus[28]和毛竹Phyllostachys edulis[29]叶绿体基因组最优密码子分析结果一致,这可能与亲缘关系相近,但不同物种之间叶绿体基因组进化过程中的相对保守性有关系[21]。通过筛选获取梁山慈竹偏好使用密码子,可进一步对目标基因进行密码子优化,提高梁山慈竹的竹笋产量和造纸纤维含量,以及利用新一代精准基因编辑工具CRISPR/Cas9优化梁山慈竹密码子,从而改造梁山慈竹基因组编辑的Cas9基因,提高该基因在梁山慈竹中的表达水平[30]。
4. 结论
本研究通过分析梁山慈竹叶绿体基因组的CDS序列,对梁山慈竹的叶绿体基因组进行生物信息学分析,筛选出梁山慈竹叶绿体基因组有GCU、GAU以及GGU等18个最优密码子。研究结果表明:影响梁山慈竹密码子偏好性的主要因素是自然选择。研究结果为后续在分子层面上利用基因工程开发梁山慈竹优良资源提供参考。
期刊类型引用(1)
1. 何刀山,阳小强,秦雅林,何海燕,谢维,罗治国,李鹏. ‘湘辣14号’叶绿体基因组密码子偏好性分析. 中国果菜. 2025(01): 47-54+79 .  百度学术
百度学术其他类型引用(0)
-

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2015.01.003
 下载:
下载:

 点击查看大图
点击查看大图
计量
- 文章访问数: 2425
- HTML全文浏览量: 343
- PDF下载量: 585
- 被引次数: 1
 下载:
下载:
