




 下载:
下载:


-
近年来,分布格局研究成为生态学研究领域的热点问题[1-3]。自然状态下,各类植物种群通常并非均匀分布,而是依不同年龄、不同研究尺度呈斑块状分布,形成复杂的种群空间分布特征。这些分布特征是对种群自身生物学特性和外在生态环境的一种表达,并且反过来影响着种群的发展趋势[4]。种群的空间分布格局可以反映物种在群落中不同尺度下的分布类型或配置方式,同一物种在不同的生活史阶段、不同的生境条件也可能表现出完全不同的分布类型[5-7]。而且分布格局具有尺度依赖性,在一个尺度表现为聚集分布,在其他尺度则可能表现为均匀分布或随机分布[8]。一般来说,如果竞争对种群起着主要制约因素,种群趋向于均匀分布;如果生物学特性起主要作用,则大多为聚集分布;如果以上两者都不起决定性作用,种群一般表现为随机分布[4]。种内关联性可以分析种群不同生长阶段个体间的功能关系和相互作用[9]。空间正关联表明相互促进及互利的生态关系,空间负关联表明相互排斥及不利的生态关系,空间无关联则意味着个体间无明确的生态关系[10-11]。狭果秤锤树Sinojackia rehderiana为安息香科Styracaceae秤锤树属Sinojackia植物,仅间断分布于华南和华东,分布范围狭窄且个体数量少。狭果秤锤树性喜湿润的沟谷常绿阔叶林内,分布海拔较低,易受到人为干扰。生境破坏和自身种子萌发率低等原因导致分布范围逐渐变窄[12],有标本采集记录的有些产地已找不到该种的分布。目前,已有一些学者对其群落区系[13]、年龄结构和分布格局[14]、群落结构及更新[15]、种间联结性[16]、遗传印记[17]等方面做过研究,而关于种群空间格局和关联性研究则鲜有报道。本研究旨在为该群落和狭果秤锤树种群就地保护提供理论支持,为迁地保护种群空间配置提供建议。
-
研究区地处江西省九江市永修县青山村村前一处风水林(29°07′55.8″~29°08′15.5″N,115°42′39.8″~115°42′52.7″E),海拔高度约23~25 m。林地面积约7 hm2,地势平坦,内有一条狭长的河沟,外围由田地和乡村小道环绕,部分地段人为活动频繁。该地属亚热带湿润季风气候区,年平均气温17.5 ℃,最冷月1月平均气温5.0 ℃,最热月7月平均气温29.1 ℃,年均降水量1 560.9 mm,年均相对湿度78%。土壤为沙质壤土,pH 6.0~6.5。该风水林为亚热带常绿阔叶林,垂直结构明显,乔木上层以樟树Cinnamomum camphora、紫弹树Celtis biondii占绝对优势,乔木下层以狭果秤锤树、尾叶冬青Ilex wilsonii和瓜木Alangium platanifolium为优势种;灌木层分布最多的为狭果秤锤树和瓜木、大青Clerodendrum cyrtophyllum等;草本层主要物种为江南短肠蕨Allantodia metteniana、接骨草Sambucus javanica、三脉紫菀Aster trinervius subsp. ageratoides等。
-
采用相邻格子样方法,在人为活动相对较少,狭果秤锤树集中分布地段,设置1个面积50 m × 100 m的固定监测大样方,调查时将大样方分成50个10 m × 10 m的小样方,小样方四角用罗盘仪定位,安装PVC-U管,距离用皮尺测量。采用每木调查法对小样方内所有狭果秤锤树进行统计,记录每株植物的胸径(地径)、树高、冠幅以及相对位置坐标、生长状况等信息。在1.3 m高度以上有萌枝的个体只记录主干信息,若萌枝在1.3 m以下分别记录各枝信息。以大样方西南角为坐标原点,东西向为坐标横轴,南北向为坐标纵轴,记录植株坐标位置。
-
在相同的环境下,种群的年龄结构与径级结构对环境具有一致的反应规律[18]。本研究采用以径级代替年龄级的方法对狭果秤锤树进行径级的划分,胸径(DBH)以1 cm为一级,即0<DBH<1 cm为第1级,1≤DBH<2 cm为第2级,依此类推。对狭果秤锤树高度级(H)的划分,以1 m为一级,方法同径级划分。将狭果秤锤树按径级和高度级,分别统计株数,采用SigmaPlot 14.0软件制表,绘制种群结构柱状图。
根据狭果秤锤树生活史以及种群实际年龄大小情况,将狭果秤锤树按不同的大小级分成4个生活史阶段,定义0<DBH<1.0 cm的植株为幼苗、1.0≤DBH<2.5 cm的植株为幼树、2.5≤DBH<6.0 cm植株为中树、DBH≥6.0 cm植株为成树。
-
分别对狭果秤锤树不同生活史阶段进行分布格局和类间关联性点格局分析。对某一生活史阶段进行空间分布格局分析时,采用单变量完全空间随机模型(CSR),将空间尺度限定在半径小于10 m内。根据Monte Carlo模拟结果,单变量O(r)值在置信区间之上为该尺度上的聚集分布,O(r)值大小代表聚集强度大小;O(r)值在置信区间之内则为该尺度下的随机分布;O(r)值在置信区间之下为该尺度下的均匀分布。不同生活史阶段进行点格局双变量分析,如果双变量O(r)值分别在置信区间上、置信区间内、置信区间下时,分别表明两者在该尺度空间正相关、相互独立、负相关[19-22]。
点格局分析采用生态学软件Programita(2014),研究尺度0~25 m,设定圆环宽度为1 m。运用Monte Carlo循环199次,95%置信区间。
-
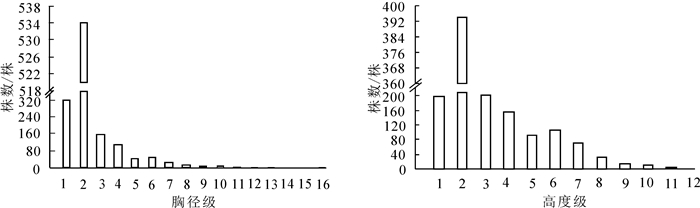
狭果秤锤树径级结构见图 1。样地内共调查到狭果秤锤树1 275株,胸径最大为15.7 cm。径级1~16的株数分别为321、534、159、106、45、46、27、13、9、6、2、2、2、1、0、2株。种群径级结构完整,小径级个体多,幼苗、幼树充足,表明种群更新能力较强;大径级个体相对较少,体现了狭果秤锤树灌木或小乔木的生物学特征。种群高度级结构见图 1。高度级可表现种群在垂直空间上的配置状况[23],与径级结构结合可更好地表现种群的大小状态。狭果秤锤树高度级完整无缺失,与径级结构类似,低矮个体较多,其中1~2 m高个体数量最多;5 m以上个体数234株,最高个体11 m。径级结构和高度级结构表明狭果秤锤树种群发展稳定,正处于增长阶段。

图 1 狭果秤锤树种群径级结构和高度级结构
Figure 1. DBH classes and height classes of S. rehderiana population
-
狭果秤锤树幼苗321株、幼树639株、中树251株、成树64株。各生活史阶段个体均散生在样地中(图 2),其中样地正西北中线附近个体较少,样地东北角分布有较多的幼苗和幼树,中树和成树分布的区域均有幼苗和幼树分布。
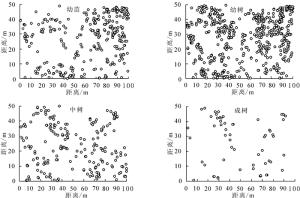
图 2 狭果秤锤树不同发育阶段个体在样地中的分布点图
Figure 2. Distribution of individuals of different development stages of S. rehderiana population
-
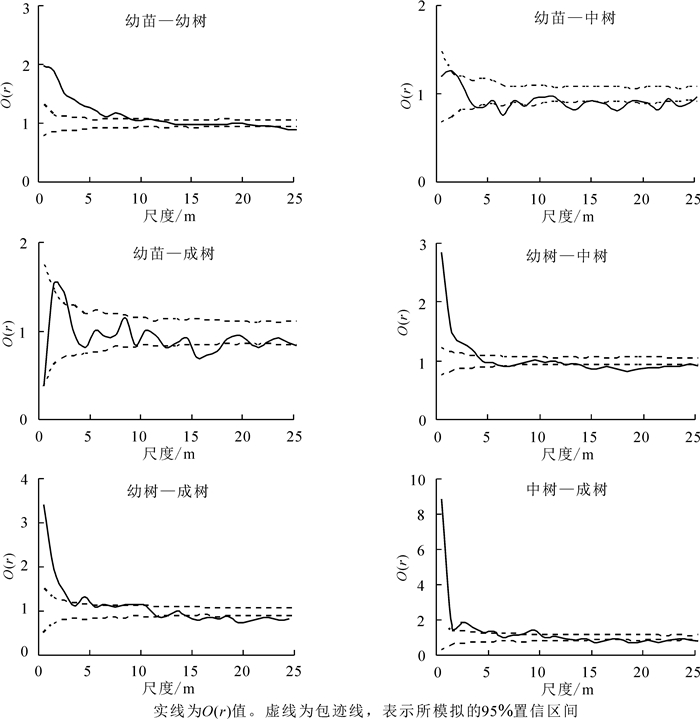
根据图 3可初步判断,狭果秤锤树幼苗在0~21 m的尺度上呈显著聚集分布,21~25 m的较大尺度上呈随机分布;幼树在0~11 m的尺度上聚集分布,18~20 m的尺度上均匀分布,其他尺度上随机分布;中树在0~4 m和7~8 m的较小尺度上聚集分布,其他尺度上随机分布;成树在0~1 m和2~4 m的较小尺度上聚集分布,其他更多尺度上表现为随机分布。总之,随着龄级的增加,狭果秤锤树在更多的尺度上表现出随机分布,分布格局表现为聚集分布向随机分布过渡的趋势。
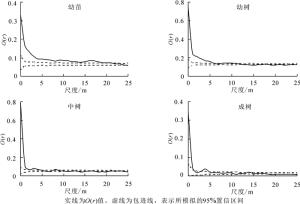
图 3 狭果秤锤树不同生活史阶段个体点格局分析
Figure 3. Spatial distribution pattern of individuals at different life stages of S. rehderiana population
-
幼苗与幼树小尺度上空间正相关,在其他更大更多的尺度上无明显空间相关性;幼苗与中树在全部尺度上表现出空间负相关与无明显相关性间断出现的趋势;幼苗和成树之间在大部分尺度上无明显空间相关性,仅在14.5~17.5 m尺度上空间负相关;幼树与中树在0~4.5 m尺度上表现为正相关,13.5~25.0 m尺度上负相关,其他尺度无明显相关性;幼树与成树在较小的尺度0~14.5 m空间正相关或不相关,14.5~25.0 m尺度上空间负相关;中树与成树在大部分尺度上空间正相关或不相关,仅在17.5~19.5、20.5~21.5 m尺度上表现出负的相关性(图 4)。总之,种群负相关性主要表现在幼苗与中树、幼树与中树、幼树与成树间,且都为较大尺度上。
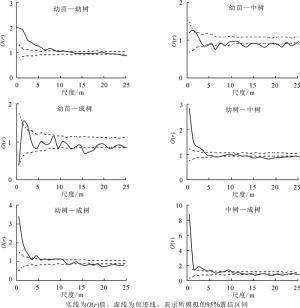
图 4 狭果秤锤树不同生活史阶段个体空间关联性
Figure 4. Spatial relationship of individuals at different life stages of S. rehderiana population
-
种群的径级结构和高度级结构不仅能反应种群的更新类型,而且能推测种群未来的发展趋势[23]。狭果秤锤树种群径级结构和高度级结构表现出一致的规律,小径级或较矮个体多,大径级或较高个体少,种群表现出稳定增长的趋势。这可能与种群所处的自然环境密不可分。调查区域地带性植被为常绿阔叶林,上层高大乔木枝繁叶茂,林内一条很长的河沟,造成林内小环境比较阴湿,而且土壤为砂性壤土,透气性很好且不积水,不会造成土壤板结和水分过饱和,天然适合于果皮坚硬且具有深休眠特性[16]的狭果秤锤树种子沙藏,导致样地内狭果秤锤树实生苗丰富。另外,样地内萌孽产生大量的萌生苗,种群表现为小个体较多,更新状态良好。
种群不同生活史阶段表现出不同的分布格局类型,这样不同年龄级的个体可以交错利用有限的环境资源[22-23]。许多研究表明:物种自身的生物学特性、其他物种的影响或者环境因素对种群空间格局均会造成影响[24-25]。狭果秤锤树不同生活史阶段个体均在小尺度上表现为聚集分布,且随着尺度的增大,聚集程度逐渐减弱;随着龄级的增加,逐渐表现出更多尺度上的均匀分布或随机分布。其原因可能是:狭果秤锤树结实量大,且种子较重,扩散能力差,掉落后主要分布在母株周围;研究区域环境适合狭果秤锤树种子萌发,产生大量的实生苗,且物种本身萌孽能力较强,产生大量的萌生植株,致使种群在较小的尺度上聚集分布;二是聚集分布更有利于形成适合种群自身生长的小环境,可以更好地发挥群体效应,以增强种间竞争力,抵御其他物种入侵,更好地维持种群的长远发展[7];三是随着龄级的增加,个体对环境中有限的资源竞争加剧,导致种内出现激烈的竞争,自疏和它疏作用增强,部分林木死亡,成熟个体向均匀分布或随机分布方向发展。
许多研究表明种群各龄级之间的相关性受龄级个体大小的影响,龄级差异越大,个体间正相关性越弱,更多的表现为不相关或负相关性[10, 19, 26-27]。形体较大的个体可能对形体较小的个体产生不利的影响,可以理解为与种内资源竞争有关,与本研究的结论相一致。狭果秤锤树空间负相关性主要表现在幼苗与中树间、幼树与中树间、幼树与成树间,反映了大径级个体对其幼苗幼树的生长有强烈的抑制作用;狭果秤锤树幼苗与幼树因形体较小,对土壤养分、光照等环境资源的竞争力较弱,在长期的生长过程中相互庇护,表现出明显的正相关关系;随着幼龄个体的不断生长,个体对资源的需求量加大,个体间相互有利的关系被打破,表现出相互胁迫的竞争关系,从而出现中树或成树对幼苗和幼树的不相关或负相关关系。
自然状态下,狭果秤锤树的结构和空间分布格局受其生物学特性和环境因素的综合影响,种群的空间格局和关联性研究仅是分析群落空间格局与过程的第一步[20],群落内狭果秤锤树种群与其他种群的空间关系,样地内各种环境因子与种群空间分布格局的相互作用如何尚不明确,均有待于深入研究。狭果秤锤树在华东和华南分布区域的缩减反应了适合的生态环境和植被的减少,在人类干扰频繁地区,如何减少对森林植被的影响和破坏,可持续的营林措施和管理决策显得至关重要。自然分布于永修县的这片狭果秤锤树林天然更新良好,在就地保护过程中要尽量避免人为干扰破坏,杜绝进林中取土挖坑、砍伐等活动;在迁地保护过程中努力营造类似原生境的立地条件,特别是土壤条件,提高苗木成活率和生长速度,促进狭果秤锤树种群的天然更新。
Natural population distribution pattern of Sinojackia rehderiana in an evergreen broadleaf forest
-
摘要:
目的 揭示狭果秤锤树Sinojackia rehderiana自然种群不同生活史个体分布格局,为该种群的保护提供科学依据。 方法 以自然分布于江西省永修县的狭果秤锤树种群为对象,采用相邻格子样方调查法、空间代替时间和O-ring完全随机零模型法,对狭果秤锤树自然种群不同生活史阶段个体的空间分布格局和空间关联性进行分析,探讨其空间分布格局形成原因。 结果 狭果秤锤树径级结构和高度级结构完整,幼苗和幼树的数量在整个种群中占绝对优势,种群更新状态良好,属增长型种群。狭果秤锤树种群在从幼苗发育到成树的过程中,在小尺度上聚集分布,大尺度上为随机分布至均匀分布;随着龄级的增加,聚集分布趋势减弱。在较小的尺度上,不同生活史阶段个体表现出正相关或不相关,在较大的尺度上则表现为不相关或负相关;负相关性主要表现在幼苗与中树、幼树与中树、幼树与成树间(P < 0.05)。 结论 在自然状态下,狭果秤锤树分布格局受自身生物学特性和种内、种间竞争影响,幼苗和幼树生长受限但数量较多,种群目前尚未达到稳定状态。 Abstract:Objective To reveal the spatial distribution pattern in different life cycle stages, and to provide a scientific basis for protection of Sinojackia rehderiana, which was naturally distributed in Yongxiu County, Jiangxi Province. Method To probe causes of the spatial distribution pattern formation, the adjacent lattice sampling method, the spatial sequence method instead of time sequence, and the O-ring Completely Random Zero Model method were used to analyze the spatial distribution pattern and spatial correlation of individuals in different life cycle stages of a natural population of S. rehderiana. Result The DBH class structure and height class structure of S. rehderiana were complete, and the number of seedlings and saplings occupied an absolute advantage for the entire population. Population renewal, which belonged to the growth population, was in a favorable condition. In the process of development from seedling to adult tree, the population of S. rehderiana had an aggregated distribution on a small scale, and had a random to uniform distribution on a large scale. As the age class increased, the aggregated distribution receded. On a small scale, at different life cycle stages of S. rehderiana, there was a significantly positive correlation or no correlation(P=0.05); however, on a large scale there was a negative association or no correlation. The spatial association between seedlings and middle-aged trees, saplings and middle-aged trees, and saplings and adult trees was significantly negative (P=0.05). Conclusion In a natural state, the distribution pattern of S. rehderiana was influenced by its own biological factors, intraspecific competition, and interspecific competition; seedling and sapling growth was limited but plentiful; and the population was not currently in a stable state. -
Key words:
- forest ecology /
- Sinojackia rehderiana /
- point pattern analysis /
- spatial association /
- age class
-
图 1 狭果秤锤树种群径级结构和高度级结构
Figure 1 DBH classes and height classes of S. rehderiana population
图 2 狭果秤锤树不同发育阶段个体在样地中的分布点图
Figure 2 Distribution of individuals of different development stages of S. rehderiana population
图 3 狭果秤锤树不同生活史阶段个体点格局分析
Figure 3 Spatial distribution pattern of individuals at different life stages of S. rehderiana population
-
[1] 杨乃坤, 邹天才, 刘海燕, 等.贵州特有植物长柱红山茶种群年龄结构及空间分布格局研究[J].热带亚热带植物学报, 2015, 23(2): 205-210. YANG Naikun, ZOU Tiancai, LIU Haiyan, et al. Age structure and spatial distribution pattern of endemic plant Camellia mairei var. lapidea population in Guizhou Province, China[J]. J Trop Subtrop Bot, 2015, 23(2): 205-210. [2] LÜ Xizhi, ZUO Zhongguo, SUN Juan, et al. Spatial patterns of dominant species and their implication for natural secondary forest ecosystem dynamics in a reserved forest of north China[J]. Ecol Eng, 2019, 127: 460-467. [3] 梁爽, 许涵, 林家怡, 等.尖峰岭热带山地雨林优势树种白颜树空间分布格局[J].植物生态学报, 2014, 38(12): 1273-1282. LIANG Shuang, XU Han, LIN Jiayi, et al. Spatial distribution pattern of the dominant species Gironniera subaequalis in tropical montane rainforest of Jianfengling, Hainan Island, China[J]. Chin J Plant Ecol, 2014, 38(12): 1273-1282. [4] CARRER M, CASTAGNERI D, POPA I, et al. Tree spatial patterns andstand attributes in temperate forests: the importance of plot size, sampling design, and null model[J]. For Ecol Manage, 2018, 407: 123-134. [5] COMITA L S, CONDIT R, HUBBELL S P. Developmental changes in habitat associations of tropical trees[J]. J Ecol, 2007, 95: 482-492. [6] COMDIT R, ASHTON P S, BAKER P, et al. Spatial patterns in the distribution of tropical tree species[J]. Science, 2000, 288(5470): 1414-1418. [7] 陈小荣, 李乐, 夏家天, 等.百山祖亮叶水青冈种群结构和分布格局[J].浙江农林大学学报, 2012, 29(5): 647-654. CHEN Xiaorong, LI Le, XIA Jiatian, et al. Population structure and distribution of Fagus lucida in the Baishanzu forest[J]. J Zhejiang A & F Univ, 2012, 29(5): 647-654. [8] 杨华, 李艳丽, 沈林, 等.长白山云冷杉林针阔混交林主要树种空间分布及其关联性[J].生态学报, 2014, 34(16): 4698-4706. YANG Hua, LI Yanli, SHEN Lin, et al. Spatial distributions and associations of main tree species in a spruce-fir forest in the Changbai Mountain area in northeastern China[J]. Acta Ecol Sin, 2014, 34(16): 4698-4706. [9] 王世彤, 吴浩, 刘梦婷, 等.极小种群野生植物黄梅秤锤树群落结构与动态[J].生物多样性, 2018, 26(7): 749-759. WANG Shitong, WU Hao, LIU Mengting, et al. Community structure and dynamics of a remnant forest dominated by a plant species with extremely small population (Sinojackia huangmeiensis) in central China[J]. Biodiver Sci, 2018, 26(7): 749-759. [10] 杨洪晓, 张金屯, 吴波, 等.毛乌素沙地油蒿种群点格局分析[J].植物生态学报, 2006, 30(4): 563-570. YANG Hongxiao, ZHNAG Jintun, WU Bo, et al. Point pattern analysis of Artemisia ordosica population in the Mu Us sandy land[J]. Chin J Plant Ecol, 2006, 30(4): 563-570. [11] 王燕, 毕润成, 许强.山西太岳山南部针阔混交林群落特征及空间分布格局[J].广西植物, 2017, 37(7): 901-911. WANG Yan, BI Runcheng, XU Qiang. Community characteristic s and spatial distribution of mixed conifer and broad-leaved forest in South Taiyue Mountain, Shanxi Province of China[J]. Guihaia, 2017, 37(7): 901-911. [12] 姚小洪, 叶其刚, 康明, 等.秤锤树属与长果安息香属植物的地理分布及其濒危现状[J].生物多样性, 2005, 13(4): 339-346. YAO Xiaohong, YE Qigang, KANG Ming, et al. Geographic distribution and current status of the endangered genera Sinojackia and Changiostyrax[J]. Biodiver Sci, 2005, 13(4): 339-346. [13] 谢国文, 王惟荣, 何静欣, 等.濒危植物狭果秤锤树所在群落的区系特征[J].广州大学学报(自然科学版), 2012, 11(4):18-24. XIE Guowen, WANG Weirong, HE Jingxin, et al. Floristic characteristics of a community with endangered plant Sinojackia rehderiana in the north of Jiangxi[J]. J Guangzhou Univ Nat Sci Ed, 2012, 11(4):18-24. [14] 徐惠明, 谢国文, 王业磷, 等.狭果秤锤树种群年龄结构和空间分布格局研究[J].广东农业科学, 2016, 43(8):51-57. XU Huiming, XIE Guowen, WANG Yelin, et al. Study on age structure and spatial distribution pattern of Sinojackia rehderiana population[J]. Guangdong Agric Sci, 2016, 43(8): 51-57. [15] 周赛霞, 彭焱松, 高浦新, 等.狭果秤锤树群落结构与更新特征[J].植物资源与环境学报, 2019, 28(1): 96-104. ZHOU Saixia, PENG Yansong, GAO Puxin, et al. Structure and regeneration characteristics of Sinojackia rehderiana community[J]. J Plant Resour Environ, 2019, 28(1): 96-104. [16] 周赛霞, 彭焱松, 丁剑敏, 等.珍稀植物狭果秤锤树群落木本植物种间联结性及群落稳定性研究[J].广西植物, 2017, 37(4): 442-448. ZHOU Saixia, PENG Yansong, DING Jianmin, et al. Analysis on community stability and inter-specific correlations among dominant woody populations of the endangered plant Sinojackia rehderiana communities[J]. Guihaia, 2017, 37(4): 442-448. [17] YAO X, ZHANG J, YE Q, et al. Fine-scale spatial genetic structure and gene flow in a small, fragmented population of Sinojackia rehderiana (Styracaceae), an endangered tree species endemic to China[J]. Plant Biol, 2011, 13(2):401-410. [18] FROST I, RYDIN H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests[J]. Ecoscience, 2000, 7(1): 38-44. [19] 陈科屹, 张会儒, 雷相东.天然次生林蒙古栎种群空间格局[J].生态学报, 2018, 38(10): 3462-3470. CHEN Keyi, ZHANG Huiru, LEI Xiangdong. Spatial pattern of Quercus mongolica in natural secondary forest[J]. Acta Ecol Sin, 2018, 38(10): 3462-3470. [20] 张健, 郝占庆, 宋波, 等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报, 2007, 18(8): 1681-1687. ZHANG Jian, HAO Zhanqing, SONG Bo, et al. Spatial distribution patterns and associations of Pinus koraiensis and Tilia amurensis in broad-leaved Korean pine mixed forest in Changbai Mountains[J]. Chin J Appl Ecol, 2007, 18(8): 1681-1687. [21] 缪宁, 刘世荣, 史作民, 等.青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局[J].生态学报, 2011, 31(1): 1-9. MIAO Ning, LIU Shirong, SHI Zuomin, et al. Spatial pattern analysis of a Rhododendron-Abies virginal forest near timberline on the eastern edge of Qinghai-Tibetan Plateau, China[J]. Acta Ecol Sin, 2011, 31(1): 1-9. [22] 郭垚鑫, 胡有宁, 李刚, 等.太白山红桦种群不同发育阶段的空间格局与关联性[J].林业科学, 2014, 50(1): 9-14. GUO Yaoxin, HU Youning, LI Gang, et al. Spatial pattern and spatial association of Betula albosinensis at different developmental stages at Taibai Mountain[J]. Sci Silv Sin, 2014, 50(1): 9-14. [23] 黄小波, 李帅锋, 苏建荣, 等.云龙天池云南松自然种群分布格局分析[J].林业科学研究, 2018, 31(4): 47-52. HUANG Xiaobo, LI Shuaifeng, SU Jianrong, et al. Distribution of Pinus yunnanensis natural population in Yunlong Tianchi National Nature Reserve[J]. For Res, 2018, 31(4): 47-52. [24] 沈志强, 华敏, 丹曲, 等.藏东南川滇高山栎种群不同生长阶段的空间格局与关联性[J].应用生态学报, 2016, 27(2): 387-394. SHEN Zhiqiang, HUA Min, DAN Qu, et al. Spatial pattern analysis and associations of Quercus aquifolioides population at different growth stages in Southeast Tibet China[J]. Chin J Appl Ecol, 2016, 27(2): 387-394. [25] CHENG Xiaoqing, HAN Hairong, KANG Fengfeng, et al. Point pattern analysis of different life stages of Quercus liaotungensis in Lingkong Mountain, Shanxi Province, China[J]. J Plant Interact, 2014, 9: 233-240. [26] 尤海舟, 刘兴良, 缪宁, 等.川滇高山栎种群不同海拔空间格局的尺度效应及个体间空间关联[J].生态学报, 2010, 30(15): 4004-4011. YOU Haizhou, LIU Xingliang, MIAO Ning, et al. Individual association and scale effect of spatial pattern of Quercus aquifolioides populations along the elevation gradients[J]. Acta Ecol Sin, 2010, 30(15): 4004-4011. [27] 牛丽丽, 余新晓, 岳永杰.北京松山自然保护区天然油松林不同龄级立木的空间点格局[J].应用生态学报, 2008, 19(7): 1414-1418. NIU Lili, YU Xinxiao, YUE Yongjie. Spatial patterns of different age-class individuals in Pinus tabulaeformis forest in Songshan Nature Reserve of Beijing, China[J]. Chin J Appl Ecol, 2008, 19(7): 1414-1418. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.02.004
 点击查看大图
点击查看大图
计量
- 文章访问数: 2269
- HTML全文浏览量: 660
- PDF下载量: 62
- 被引次数: 0
