




 下载:
下载:

-
香榧Torreya grandis‘Merrillii’是榧树Torreya grandis经人工嫁接繁殖而成的优良栽培类型[1],具有极高的经济价值、生态价值和文化价值,是集材用、药用、果用、油用和观赏于一体的多功能珍贵树种[2-3]。近几年香榧产业发展迅速,使得香榧产区有了很大的扩展。新老产区香榧林多为纯林,在相对单一的生态环境中易成片感染各种病虫害。香榧瘿螨Nalepella abiesis是香榧的主要虫害之一,其种名最初鉴定为冷杉大嘴瘿螨Rhyncaphytopus abiesis[4],2013年经重新分类后更名为Nalepella abiesis[5]。国外尚无该种瘿螨的相关报道。瘿螨是一类古老的高度微型化、形态简化和多样化的植食性寄生螨[6],大多数种类对寄主植物具有高度专一性[7]。瘿螨靠其螯肢演化的口针刺入寄主组织,以吸食细胞里的汁液和营养物质为生[8]。瘿螨可以为害植物的叶、芽、果实、茎干等部位,大部分瘿螨属于自由生活型,寄生在植物表面,少部分能引起寄主植物产生各种虫瘿、毛毡等症状[9],使寄主植物器官发生畸形,组织结构发生改变。香榧瘿螨属于寡食性昆虫,且自由生活在香榧树体地上部分的器官表面,以叶片为主要寄生场所。目前,关于香榧瘿螨的研究甚少,仅有一些关于香榧瘿螨生物学特性和发生规律[4, 10-11]及防治[11]的研究报道,但针对瘿螨吸食为害香榧叶片部位却存在着不同说法[12]。本研究从植物形态解剖学的角度探究香榧瘿螨为害对香榧叶片结构及叶绿素质量分数的影响,以期对香榧瘿螨的为害机理和防治提供理论基础。
-
香榧瘿螨采自浙江省嵊州市谷来镇(29°41'N,120°38'E),该镇为香榧老产区。香榧瘿螨带回实验室后在室内用新鲜香榧叶片继代饲养,并用体式显微镜对其进行观察。香榧苗来自浙江省诸暨市立勤香榧种植基地长势一致的2+3(2年生实生苗嫁接后再栽培3 a)香榧盆栽苗(盆径25 cm)。
-
试验于2018年9-10月在浙江农林大学国家林业和草原局林木良种繁育基地温室大棚(30°15'N,119°43'E)内进行。香榧叶条形,近对生于枝条的两侧,上表面光绿色,下表面有2条白色气孔带[1]。选择健康无病虫害且叶片大小、数量基本一致的香榧1年生成熟枝条,按30头·枝条-1的虫口密度,用0号带微钩的昆虫针轻轻地将香榧瘿螨接种到枝条叶表上,以未接种瘿螨的盆栽苗为空白对照组。每个处理重复3次,每重复5盆苗。每盆苗之间均保持一定的距离,对每个接螨的枝条于基部节处涂抹凡士林以防止瘿螨爬出供试枝条[13],并保持供试枝条隔离无交叉感染。依次在瘿螨侵染的第0天(即健康叶片)、第12天、第25天和第40天,分别剪取2个处理的香榧叶片,将每个重复的叶片各自混匀。采集的叶片用于形态解剖学的观察和总叶绿素质量分数的测定,使用前先用纸巾擦除叶表的瘿螨。
-
① 叶片组织结构。选取实验叶片靠中间部位切成1 mm × 2 mm大小的块状,将块状叶置于体积分数2.5%的戊二醛溶液中4 ℃条件下固定过夜;然后将样品用磷酸缓冲液(pH 7.0)漂洗3次,用质量分数1.0%的锇酸固定1.5 h,再用磷酸缓冲液漂洗3次;后用300、500、700、800、900、950 g·kg-1梯度的乙醇溶液逐级脱水,用Spurr包埋剂与丙酮混合液进行渗透过夜;用半薄切片机(瑞典LKB 11800)切片,经亚甲基蓝染色后在正置荧光显微镜(日本OLYMPUS BX 60)下观察、拍照。②叶片气孔带结构。样品双固定、脱水方法同叶片组织结构,经临界点干燥、镀膜后在扫描电镜(日本Hitacbi TM-1000)下观察、拍照。③叶绿体超微结构。样品双固定、脱水、渗透处理方法同叶片组织结构,包埋好的样品用超薄切片机(日本Leica EM UC7)切片,经柠檬酸铅溶液和乙酸双氧铀染液染色后,在透射电镜(日本JEM-1230)下观察、拍照。
-
采用乙醇浸泡法[14],用紫外-可见分光光度计(日本UV-2600)全光谱扫描测定波长665 nm处的吸光值D(665)和649 nm处的吸光值D(649),并按公式计算鲜叶叶绿素质量分数,叶绿素质量分数(mg·g-1)=0.1×[18.08×D(649)+6.63×D(665)]。数据采用Excel 2007与SPSS 17.0进行分析处理。
-
体式显微镜下观察螨害下的香榧叶片,发现香榧瘿螨可以吸食为害香榧植株的叶片上、下表面(图 1)。经长期观察发现,成螨多集中于香榧叶片上表面吸食(图 1A~B),并且雌成螨基本在香榧叶背2条白色气孔带上产卵,卵呈透明状的椭球形(图 1B~C)。

图 1 体式显微镜下观察香榧癭螨
Figure 1. Stereomicroscope observation of N. abiesis
-
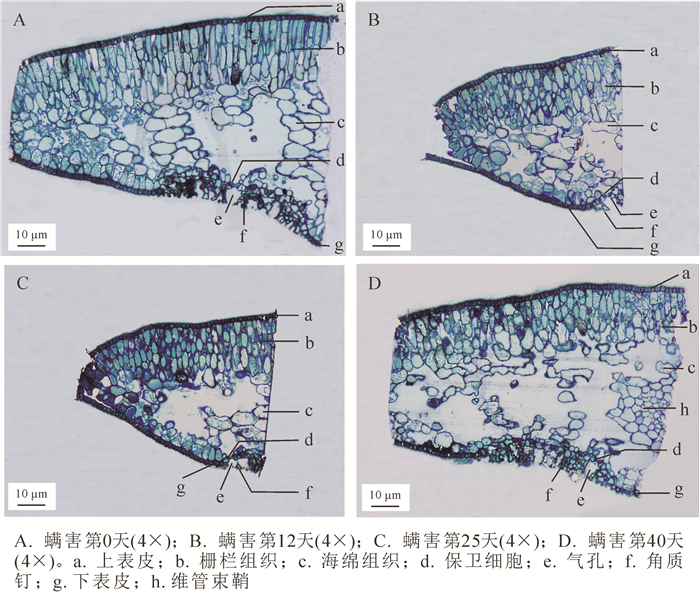
正置显微镜下观察瘿螨为害不同天数的香榧叶片组织结构,发现健康叶片上、下表皮均由一层紧密排列的细胞组成,不含叶绿体,表皮外覆着一层较薄的角质层;气孔器分布在下表皮,略内陷,由2个保卫细胞控制其开闭,周围密生分叉的角质钉(图 2A),这与前人有关榧树属Torreya植物解剖特征的描述[15-16]一致;叶肉组织发达且明显分化为栅栏组织和海绵组织,栅栏细胞呈长圆柱状,垂直于叶表皮,且排列较紧密,内含较多的叶绿体,海绵细胞呈不规则的椭圆形或圆形,排列较为疏松(图 2A)。随着螨害时间的持续,栅栏组织和海绵组织细胞形状发生改变,但上、下表皮细胞未见有明显破损(图 2B~D)。从螨害第40天的叶片结构可见,栅栏组织厚度明显减小,细胞数量减少,尤其是下表皮上方的栅栏细胞;海绵细胞亦缩水变小,且细胞间隙变大而更为松散;气孔角质钉结构有所破损,保卫细胞缩小(图 2D)。
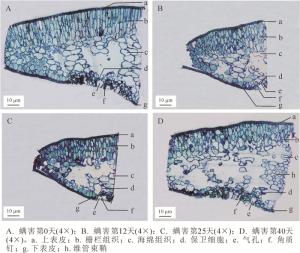
图 2 瘿蟎为害香榧叶片组织结构的动态变化
Figure 2. Dynamic changes in the leaf tissue structure on different days after inoculation with mites
-
扫描电镜下观察健康与螨害第40天的香榧叶片气孔结构,发现健康叶片的气孔呈狭长椭圆形(图 3A~B,示半闭合状态的气孔),被乳突状角质钉包围而内陷,而角质钉高度密集,呈无规则分布(图 3A)。瘿螨为害后的叶片气孔有明显的损伤和萎缩(图 3D)。气孔周围的角质钉上附着有香榧瘿螨的卵(图 3C),卵在样品制备过程中因漂洗等操作而破裂或脱落。完整的卵呈椭圆形,平均大小为49.87 μm×39.48 μm;相比健康叶片,螨害后的角质钉表面附着了更多的絮状绒毛状物(图 3D)。

图 3 健康与螨害40 d香榧叶片气孔的扫描结构的比较
Figure 3. A comparison in the leaf stomatal structure between healthy and 40-day mite-hurt leaves under the scanning electron microscope
-
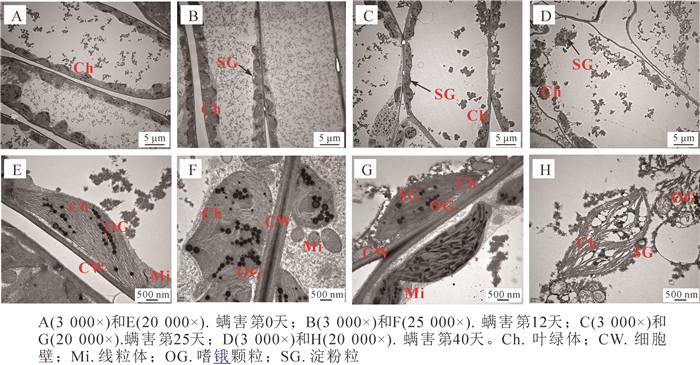
透射电镜下观察瘿螨为害不同天数香榧叶肉细胞超微结构,发现健康叶片的叶肉细胞间有一定的空隙,相邻细胞壁界限明显,细胞内叶绿体数量较多,且紧贴细胞壁分布,线粒体也比较丰富(图 4A)。单个叶绿体的基粒片层垛叠紧密,结构清晰良好,体内嗜锇颗粒不多(图 4E)。随着螨害时间的持续,叶肉细胞逐渐膨胀变形,细胞间隙变大;叶绿体数量逐渐减少,体内出现膨大的淀粉粒;线粒体数量亦减少(图 4B~D)。螨害第12天(图 4F),叶绿体内嗜锇颗粒数量明显增多,基粒片层排列逐渐散乱,结构不清晰,至螨害第25天变得模糊(图 4G)。螨害第40天,叶肉细胞内大部分叶绿体膜被破坏,体内有淀粉粒沉积,并脱离细胞壁散乱地分布在细胞质中,片层结构瓦解甚至消失(图 4H)。

图 4 瘿螨为害香榧叶肉细胞超微结构的动态变化
Figure 4. Dynamic changes in the ultrastructure of a mite-hurt mesophyll cell
-
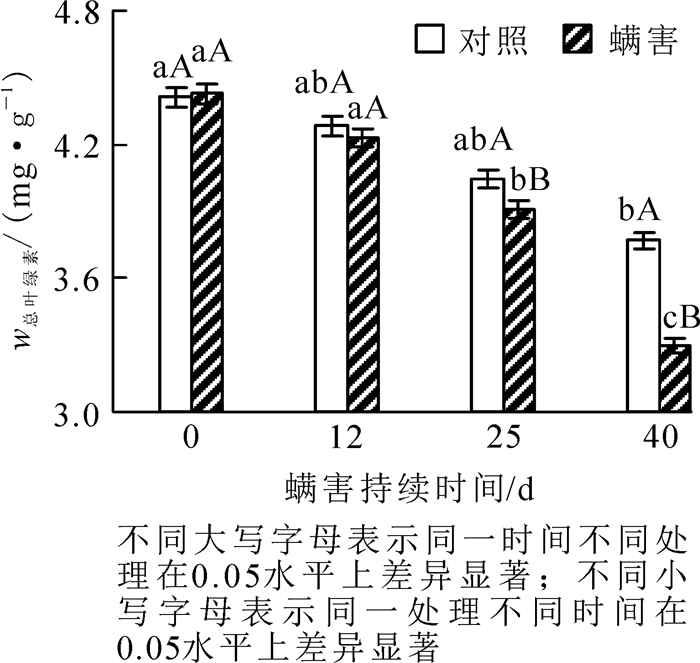
相同螨害天数下,0~12 d处理与对照的叶绿素质量分数无显著差异,但在接种瘿螨25~40 d处理与对照间叶绿素质量分数达差异显著水平,瘿螨处理叶片的叶绿素质量分数下降很快,说明瘿螨为害随着时间的推移影响逐渐增强。瘿螨为害期处理与对照香榧叶片叶绿素质量分数动态发现,对照与处理香榧叶片叶绿素质量分数均呈下降趋势,但对照组0~12 d有轻微的下降,12 d后不同时间点间差异不显著;而螨害处理0~12 d香榧叶片叶绿素质量分数无显著下降,12 d后不同时间点间叶片叶绿素质量分数下降明显,差异显著(P < 0.05),在瘿螨接种第25~40天,叶绿素质量分数从3.91 mg·g-1下降至3.3 mg·g-1;螨害第40天与第0天相比,叶绿素质量分数下降了25.34%(图 5)。另外,通过F检验发现:瘿螨为害持续天数对叶绿素质量分数变化有显著影响(F=25.331,P < 0.05)。说明螨害导致叶绿素减少,从而影响叶片进行正常的光合作用。
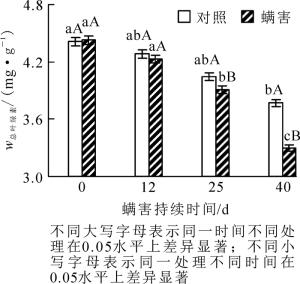
图 5 瘿螨为害香榧叶片叶绿素质量分数的动态变化
Figure 5. Dynamic changes in the leaf chlorophyll content with days after inoculatio with mites
-
逆境胁迫有多种,其中虫害胁迫是常见的一类生物胁迫[17]。不同胁迫会对植物造成不同程度的为害,同样螨害会引起植物叶片细胞结构[18]、生理[19]、生化[20]等方面的变化,这些变化反映植物体对逆境胁迫的响应。香榧叶片被香榧瘿螨刺吸式取食后,叶片组织细胞在形态结构、生理生化等方面产生了一系列变化。本研究表明:香榧叶片的下表皮上也有栅栏细胞,香榧瘿螨主要为害香榧叶片的叶肉组织,瘿螨为害后栅栏组织厚度明显减小,而栅栏组织是大量叶绿体存在的部位,进而影响叶绿素质量分数。结合体式显微镜下的观察,认为香榧瘿螨在香榧叶片上、下表面都有刺吸取食的行为。实验观察发现,成螨更喜欢在香榧叶片上表面取食,这可能跟与上表皮相连的栅栏组织厚且细胞内容物多有关。寄主植物的叶片下表皮是螨虫躲藏的主要场所,叶片气孔更是螨虫口针探入海绵组织的天然孔道[8],持续螨害使得气孔有明显的损伤和萎缩,严重影响了气孔的正常开闭,从而直接影响叶片的蒸腾作用和气体交换。由于保卫细胞内也含有少量叶绿体,推测其也可能是被香榧瘿螨刺吸后缩水变小[21]。另外,通过体式显微镜观察发现,香榧瘿螨偏好在叶片气孔带上集中产卵,一方面叶片乳突状的结构为卵提供了更好的附着点,另一方面红豆杉属Taxus叶片气孔带的角质乳突具有抵御低温侵袭的功能[22],加上植物通过气孔蒸腾失水,气孔处所保持的湿度也为卵提供了合适的孵化条件。此外,叶片气孔密度一直是抗螨性的研究热点,气孔密度与植株抗螨性呈负相关[21, 23],这为后续选育抗螨性较强的香榧种质提供了研究思路。
叶绿体是植物体内最主要的细胞器之一,它的主要功能是进行光合作用,其结构的完整性与其功能的发挥紧密关联,而叶绿体对环境胁迫又比较敏感[24]。超微结构研究表明:叶绿体对香榧瘿螨胁迫反应非常敏感。螨害后的叶绿体片层结构受损,膜系统被破坏,影响了光合产物的正常输出,导致叶绿体中有淀粉粒的沉积[18],无法进一步降解转化。叶绿体内嗜锇颗粒数量的增加,跟叶绿体的衰老退化有关[25],这具有一定的生理学意义。螨害使叶绿体数量减少,叶绿素质量分数减少,叶片逐渐失绿,继而枯萎脱落,直接导致香榧叶片光合能力下降,无法合成更多的养分,进而影响香榧的正常生长。
综上所述,香榧瘿螨吸食破坏了香榧叶片叶肉细胞和气孔器的结构,使得叶绿素质量分数减少,从而影响了香榧叶片正常的光合作用。
Leaf structure and chlorophyll content in Torreya grandis 'Merrillii' with Nalepella abiesis infestation
-
摘要:
目的 研究香榧瘿螨Nalepella abiesis对香榧Torreya grandis ‘Merrillii’叶片的为害机制,对叶片结构及叶绿素质量分数的影响,可为香榧瘿螨的防治提供理论依据。 方法 利用正置显微镜、扫描电镜和透射电镜研究螨害对香榧叶片组织结构、气孔结构以及叶绿体超微结构的动态变化,并用分光光度法测定叶绿素质量分数的动态变化。 结果 随着螨害时间的持续,香榧叶片叶肉栅栏细胞形状改变,细胞数量减少;螨害第40天时,栅栏组织厚度明显减小,海绵细胞亦缩水变小,海绵组织更为松散;气孔区域部分角质钉结构破损,保卫细胞缩小,气孔器有明显的损伤和萎缩。叶肉细胞逐渐膨胀变形,细胞间隙变大,叶绿体数量逐渐减少,并出现膨大的淀粉粒;螨害第12天时,叶绿体内嗜锇颗粒数量明显增多,基粒片层排列逐渐散乱,结构不清晰,螨害第25天时则变得模糊,到螨害第40天时片层结构瓦解消失。接种瘿螨的处理与未接种瘿螨的对照相比,0~12 d叶绿素质量分数无显著差异,但在25~40 d叶绿素质量分数差异显著(P < 0.05);随着螨害时间的持续,叶绿素质量分数下降更多。 结论 香榧瘿螨吸食破坏了香榧叶片叶肉细胞和气孔器的结构,使得叶绿素质量分数减少,从而影响了香榧叶片正常的光合作用。 Abstract:Objective The aim is to provide a theoretical basis for the prevention and control of Nalepella abiesis and to study the damage mechanism and chlorophyll content from N. abiesis on leaves of Torreya grandis 'Merrillii'. Method Dynamic changes in leaf tissue structure, stomatal structure, chloroplast ultrastructure, and chlorophyll content were studied using the fluorescence microscope, transmission electron microscope (TEM), scanning electron microscope (SEM), and spectrophotometer. Result The palisade cells changed in shape and decreased in number for an increase in days after inoculation (DAI) with mites. By the 40th day, thickness of the palisade tissue decreased considerably; spongy cells shrank; spongy tissue became looser; around stomata the cuticular peg was destroyed; guard cells shrank; and stomata shrank revealing obvious injury. Mesophyll cells gradually swelled and became deformed with the gap between cells becoming larger and larger. Also, the number of chloroplasts gradually decreased with the appearance of starch grains. By the 12th day, the number of osmiophilic granules within the chloroplast had increased with grana lamellae becoming more and more irregular in structure, especially on the 25th day, and finally disintegrating and disappearing on the 40th day. There was no significant difference in chlorophyll content between ck and the treatment with mite infection from 0 to 12 d, but a significant difference occurred from 25 to 40 d(P < 0.05). Compared to the ck, the decrease in chlorophyll content continued as time increased. Conclusion N. abiesis infestation could destroy the structure of the mesophyll cells and leaf stomata of T. grandis 'Merrillii' leading to a decreased chlorophyll content that could affect photosynthesis. -
Key words:
- forest protection /
- Torreya grandis 'Merrillii' /
- Nalepella abiesis /
- leaf structure /
- stomata /
- chloroplast /
- chlorophyll content
-
图 2 瘿蟎为害香榧叶片组织结构的动态变化
Figure 2 Dynamic changes in the leaf tissue structure on different days after inoculation with mites
图 3 健康与螨害40 d香榧叶片气孔的扫描结构的比较
Figure 3 A comparison in the leaf stomatal structure between healthy and 40-day mite-hurt leaves under the scanning electron microscope
图 4 瘿螨为害香榧叶肉细胞超微结构的动态变化
Figure 4 Dynamic changes in the ultrastructure of a mite-hurt mesophyll cell
-
[1] 黎章矩, 戴文圣.中国香榧[M].北京:科学出版社, 2007:26-55. [2] 戴文圣, 黎章矩, 程晓建, 等.杭州市香榧生产的发展前景与对策[J].浙江林学院学报, 2006, 23(3):334-337. DAI Wensheng, LI Zhangju, CHEN Xiaojian, et al. Development future and strategies of production of Torreya grandis 'Merrillii' in Hangzhou[J]. J Zhejiang For Coll, 2006, 23(3):334-337. [3] 陈力耕, 王辉, 童品璋.香榧的主要品种及其开发价值[J].中国南方果树, 2005, 34(5):36-37. CHEN Ligeng, WANG Hui, TONG Pinzhang. The varieties and values of Torreya grandis 'Merrillii'[J]. South China Fruits, 2005, 34(5):36-37. [4] 徐志宏, 吾中良.香榧病虫害防治彩色图谱[M].北京:中国农业科学技术出版社, 2004:49-50. [5] XUE Xiaofeng, GUO Jingfeng, HONG Xiaoyue. Eriophyoid mites from northeast China (Acar:Eriophyoidea)[J]. Zootaxa, 2013, 3689(1):9-12. [6] CHETVERIKOV P E, CVRKOVIC T, MAKUNIN A, et al. Basal divergence of Eriophyoidea (Acariformes, Eupodina) inferred from combined partial COI and 28S gene sequences and CLSM genital anatomy[J]. Exp Appl Acarol, 2015, 67:219-245. [7] 陈健文, 韦绥概.瘿螨与寄主植物关系的研究概况[J].广西农业生物科学, 2002(3):195-198. CHEN Jianwen, WEI Suigai. Advanced summarization of the relationships between Eriophyoid mites and host plants[J]. J Guangxi Agric Biol Sci, 2002(3):195-198. [8] 雍小菊, 丁伟.植物的抗螨机理[J].应用昆虫学报, 2011, 48(5):1495-1504. YONG Xiaoju, DING Wei. The resistant mechanisms of plants to mites[J]. Chin J Appl Entomol, 2011, 48(5):1495-1504. [9] 薛晓峰.中国古北界瘿螨总科(蜱螨亚纲: 前气门目)的分类研究[D].南京: 南京农业大学, 2007. XUE Xiaofeng. A Taxonomic Study of the Eriophoidea from the Palaearctic Region of China (Acari: Prostigmata)[D]. Nanjing: Nanjing Agricultural University, 2007. [10] 吾中良, 徐志宏, 陈秀龙, 等.香榧病虫害种类及主要病虫害综合控制技术[J].浙江林学院学报, 2005, 22(5):545-552. WU Zhongliang, XU Zhihong, CHEN Xiulong, et al. Species and controls of main diseases and insect pests of Torreya grandis 'Merrillii'[J]. J Zhejiang For Coll, 2005, 22(5):545-552. [11] 叶旭燕, 毛朝明.香榧瘿螨发生规律与防治方法[J].林业科技通讯, 2016(8):30-32. YE Xunyan, MAO Chaoming. The occurrence regularity and control methods of Eriophyes sp.[J]. For Sci Technol, 2016(8):30-32. [12] 高樟贵, 张敏, 厉锋, 等.香榧病虫害研究进展[J].浙江林业科技, 2018, 38(5):98-104. GAO Zhanggui, ZHANG Min, LI Feng, et al. Research advances on diseases and pests in Torreya grandis[J]. J Zhejiang For Sci Technol, 2018, 38(5):98-104. [13] 温娟, 郅军锐, 吕召云, 等.二斑叶螨为害对番茄叶片主要营养物质和防御酶活性的影响[J].环境昆虫学报, 2017, 39(1):172-181. WEN Juan, ZHI Junrui, LÜ Zhaoyun, et al. Effects of Tetranychus urticae feeding on the contents of main nutrient and defensive enzymes activities of tomato leaves[J]. J Environ Entomol, 2017, 39(1):172-181. [14] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社, 2000:130-132. [15] 廖海民, 叶华.香榧幼苗的形态学研究[J].贵州农学院学报, 1996, 15(1):40-45. LIAO Haimin, YE Hua. A study of seedling morphology of Torreya grandis Foxt et Lindl[J]. J Guizhou Agric Coll, 1996, 15(1):40-45. [16] 苏应娟.红豆杉科、三尖杉科和罗汉松科植物叶片结构的比较观察[J].武汉植物学研究, 1997, 15(4):307-316. SU Yingjuan. Comparative observation on leaf structure of Taxaceae, Cephalotaxaceae and Podocarpaceae[J]. J Wuhan Bot Res, 1997, 15(4):307-316. [17] 张正斌.植物对环境胁迫整体抗逆性研究若干问题[J].西北农业学报, 2000, 9(3):112-116. ZHANG Zhengbin. Discussion about studies of plant stress resistance to environment[J]. Acta Agric Boreali-Occident Sin, 2000, 9(3):112-116. [18] 张坤鹏, 孙瑞红, 杨洪强.害螨胁迫对富士苹果叶片细胞超微结构及游离脯氨酸含量的影响[J].山东农业科学, 2016, 48(6):36-41. ZHANG Kunpeng, SUN Ruihong, YANG Hongqiang. Influences of pest mite stress on cell ultrastructure and free proline content of fuji apple leaves[J]. Shandong Agric Sci, 2016, 48(6):36-41. [19] DAUD R D, CASSIA C E, FERES R J. Changes in leaf physiology caused by Calacarus heveae (Acari:Eriophyidae) on rubber tree[J]. Exp Appl Acarol, 2012, 57:127-137. [20] BEAULIEU F, CUERRIER A, FILION V J, et al. A gall mite, Aceria rhodiolae (Acari:Eriophyidae), altering the phytochemistry of a medicinal plant, Rhodiola rosea (Crassulaceae), in the Canadian Arctic[J]. Ann Mag Nat Hist, 2016, 50(19/20):1237-1263. [21] 桂连友, 龚信文, 孟国玲, 等.茄子叶片气孔密度与侧多食跗线螨发生数量的关系[J].园艺学报, 2001, 28(2):170-172. GUI Lianyou, GONG Xinwen, MENG Guoling, et al. The relationship between the stoma density of eggplant leaves and their resistance to Polyphagotarsonemus latus[J]. Acta Hortic Sin, 2001, 28(2):170-172. [22] 李凤英, 梁士楚.元宝山南方红豆杉的解剖结构及其环境适应性[J].广西植物, 2013, 33(2):219-224. LI Fengying, LIANG Shichu. Anatomical structure and environmental adaptability of Taxus wallichiana var. mairei in Yuanbaoshan[J]. Guihaia, 2013, 33(2):219-224. [23] 肖建辉, 周颖, 叶剑秋, 等.木薯叶片厚度、蜡质含量和气孔密度与抗朱砂叶螨的关系[J].热带作物学报, 2017, 38(3):541-544. XIAO Jianhui, ZHOU Ying, YE Jianqiu, et al. The relationship between the leaf thickness, waxiness content and stoma density and the cassava resistance to Tetranychus cinnabarinus[J]. Chin J Trop Crops, 2017, 38(3):541-544. [24] 任丽花, 黄敏敏, 江枝和, 等.干旱胁迫对圆叶决明叶片超微结构的影响[J].中国草地学报, 2008, 30(1):30-34. REN Lihua, HUANG Minmin, JIANG Zhihe, et al. Effect of drought stress on leaf ultrastructure of Chamaecrista rotundifolia[J]. Chin J Grassl, 2008, 30(1):30-34. [25] 陈玉银, 洪健, 童启庆.茶叶叶绿体的嗜锇颗粒含量和适制茶类的关系[J].广西植物, 1992, 12(4):345-348. CHEN Yuyin, HONG Jian, TONG Qiqing. A study on the relation of osmiophilic granules numbers in chlorophyll to the suitability for tea preparation of Camellia sinensis (L.) Kuntze[J]. Guihaia, 1992, 12(4): 345-348. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.02.014
 点击查看大图
点击查看大图

计量
- 文章访问数: 2779
- HTML全文浏览量: 928
- PDF下载量: 38
- 被引次数: 0
