





-
经过长期植树造林,近年来四川省东部地区植被得以恢复,不同林分构成的植被和土壤生态系统功能受到广泛关注[1]。华蓥市的人工林多为同龄纯林,结构和功能单一,易退化成脆弱和不稳定的生态系统。同时,因土层薄,肥力差,水土资源空间不匹配,当地土地退化,生态防护效果不够显著[2]。土壤团聚体作为土壤颗粒的聚合体和土壤肥力的物质基础,对提高林地土壤抗蚀能力具有重要作用[3]。良好的团聚体能充分协调土壤水、气、热和养分的循环,为植被生长创造良好的条件[4]。土壤团聚体的组成和稳定性是衡量土壤结构和品质的主要指标[5],水稳定性团聚体的数量则是判定土壤品质好坏的重要指标之一[6]。通过测定土壤团聚体的稳定性及粒径组成可以间接量化土壤的可蚀性能力[7]。目前,关于不同林分团聚体稳定性的影响因素方面已经有大量研究,LARS等[8]和CHUKWUEBUKA等[9]指出:土壤有机碳能够将直径较小的土壤颗粒粘结成较大的土壤颗粒。有研究[10-12]表明:土壤总孔隙度、毛管孔隙度、非毛管孔隙度的减少会显著增加土壤团聚体的稳定性,土壤有机碳含量能够促进土壤团聚过程及其稳定性。而谢贤健等[1]研究发现:土壤容重增加会恶化土壤团聚体稳定性,减弱土壤抗蚀能力,土壤中的速效钾含量对团聚体水稳性的保持、供应和转化能力具有重要作用。总体来说,土壤理化性质与团聚体稳定性密切相关,不同土壤类型决定其成土母质和土壤质地有所差异,使得这些研究结果不尽相同。此外,有关于渠江东岸川东平行岭谷区传统的人工经济林与近自然经营模式林分下土壤理化性质差异、土壤团聚体稳定性和土壤抗蚀性的关系的研究相对较少,无法满足提升该地区植被恢复和土壤保持服务功能的需要。研究林地水稳性团聚体稳定性及其影响因素对于调节林地土壤抗蚀能力,提升土壤服务功能具有重要价值。本研究选取该地区5种典型林分(柏木Cupressus funebris纯林、杉木Cunninghamia lanceolata纯林、马尾松Pinus massoniana -檵木Loropetalum chinense混交林、马尾松-杉木混交林、近自然经营杉木纯林),探究不同林分下土壤理化性质对团聚体稳定性和抗蚀能力的影响,有助于了解该地区土壤理化性质和土壤团聚体稳定性规律,为当地植被合理利用和水土流失治理提供参考依据。
-
研究区处于四川省华蓥市(30°25′21″N,106°50′02″E),地处四川盆地东缘,属于亚热带湿润性季风气候,四季分明,夏长雨多,年降水量1 200.0 mm,热量充足,年平均气温15.0~23.0 ℃,市内海拔为206.7~1 704.0 m。华蓥市西部地势偏低,多为丘陵,又以低丘为主,深丘较少,其间散布着一些平坝、台地。土壤肥沃,灌溉便利,是主要的农耕区。土壤类型以紫色土为主,土壤层发育完整,人为干扰较少。植被资源丰富,主要植被类型有亚热带常绿阔叶林、常绿落叶阔叶混交林、针阔叶混交林、常绿针叶林、针叶混交林、竹林、灌丛及人工植被。以马尾松、杉木、柏木、湿地松Pinus elliottii、柳杉Cryptomeria fortunei等树种组成的针叶林占华蓥市森林面积的96.9%,成为华蓥市典型的人工林。
-
2019年7−9月,在华蓥市天池国有林场狮子垴管护区,选取具有代表性的5种典型林分:柏木纯林(BM)、马尾松-杉木混交林(MS)、马尾松-檵木混交林(MJ)、杉木纯林(SC)、近自然经营杉木纯林(SCZ)。近自然经营是以森林的整个生命周期为时间设计单元,以目标树的标记和择伐,以及天然更新为主要技术特征,以永久性森林覆盖、多功能经营和多种产品生产为目标的森林经营体系。近自然经营的具体经营措施包括①间伐:至少2次间伐,第1次间伐年龄为10 a,强度为10~15 m3·hm−2;第2次间伐强度为25~40 m3·hm−2。②择伐:选择大约 400~600株,平均间距4~5 m,包括潜在的阔叶树。③皆伐和补植:皆伐 20 m×20 m 区域,补植枫香Liquidambar formosana、樟树Cinnamomum camphora、盐肤木Rhus chinensis等阔叶树,保留下层和中间层,促进阔叶树生长。本研究中除近自然经营杉木纯林外,其他4种林分均为以人工更新为主中国传统的森林抚育经营模式。样地调查和土壤样品采集时,每种林分设置3个重复样地(20 m×20 m),每个样地内按“S”形选择5个土壤采集样点,除去土壤表层覆盖的枯枝落叶层,挖取0~15、15~30 cm层土壤,装入塑料饭盒中,带回实验室,挑除其中的杂草、石块,风干后用于测量土壤团聚体和化学性质。使用环刀采取原状土壤,每层沿土壤剖面均匀采集3份,取其平均值作为该土壤层指标测定值,共选取90份土样,带回实验室称量烘干,测定土壤孔隙度、含水量等物理指标。环刀的容积为100 cm3,高为5.0 cm,直径为5.5 cm。样地基本情况见表1。
表 1 样地基本情况
Table 1. Basic situation of the sample plot
林分类型 纬度(N) 经度(E) 海拔/m 坡度/(°) 坡向/(°) 树高/m 胸径/cm 郁闭度 柏木纯林(BM) 30°25′17″ 106°49′59″ 518 25 西偏北10 1.80 1.98 未郁闭 马尾松-杉木混交林(MS) 30°26′15″ 106°50′43″ 1 100 31 东偏南10 15.60 31.38 0.76 近自然经营杉木纯林(SCZ) 30°18′06″ 106°47′45″ 1 409 33 北偏西10 14.30 20.51 0.85 马尾松-檵木混交林(MJ) 30°26′12″ 106°50′47″ 575 33 东偏南10 16.40 19.21 0.83 杉木纯林(SC) 30°18′28″ 106°47′47″ 1 133 33 东偏南70 15.80 32.37 0.72 -
土壤有机碳采用重铬酸钾外加热氧化法测定,土壤速效磷采用0.5 mol·L−1碳酸氢钠浸提、钼锑抗比色法测定,土壤速效钾采用四苯硼钠比浊法测定;土壤容重、毛管孔隙度、非毛管孔隙度等采用环刀法测定[13]。土壤大团聚体质量百分比(WSA,粒径>0.25 mm)、平均质量直径(MWD,单位为mm,用湿筛团聚体粒级计算)和土壤可蚀性因子K值参照文献[14-15]计算。
不同的平均质量直径对应不同的团聚体稳定性等级。平均质量直径大于2.00 mm代表团聚体非常稳定,为1.30~2.00 mm代表团聚体稳定,为0.80~1.30 mm代表团聚体中度稳定,为0.40~0.80 m代表不稳定,为0~0.40 mm代表团聚体非常不稳定[16]。
-
利用Excel 2010和SPSS 24.0进行数据统计分析,采用Origin 9.0软件作图。冗余分析(RDA)采用的是Canoco 5.0软件。采用单因素方差分析(one-way ANOVA)及多重比较LSD(least-significant difference)进行差异性分析。显著性水平均为0.05。
-
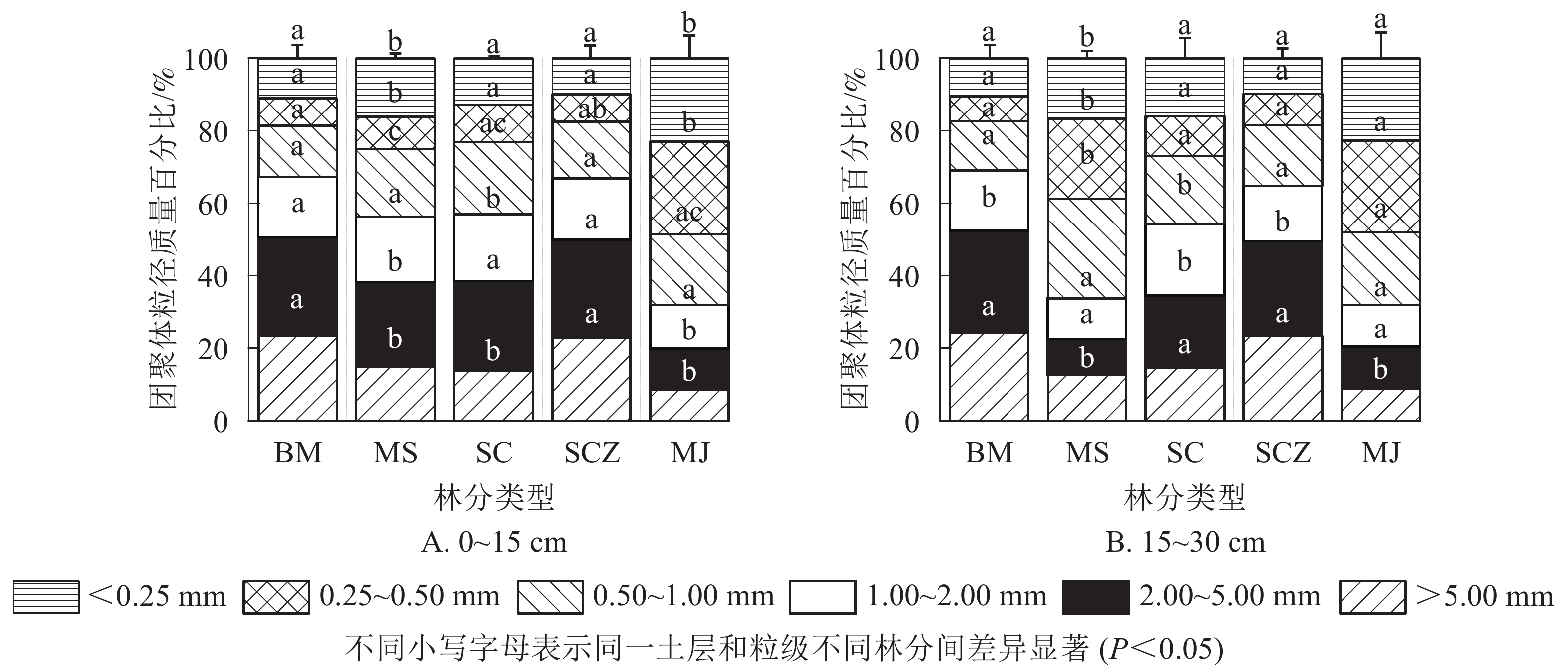
图1A表明:在0~15 cm土层,柏木纯林和近自然经营杉木纯林中粒径大于5.00 mm的团聚体质量百分比最大,分别为23.56%和22.80%,马尾松-檵木混交林中最低,为8.49%。马尾松-杉木混交林和杉木纯林粒径大于5.00 mm团聚体质量百分比分别为14.94%和13.67%。粒径大于5.00、2.00~1.00、1.00~0.50 mm的团聚体质量百分比,不同林分之间无显著差异。柏木纯林、马尾松-杉木混交林和近自然经营杉木纯林的团聚体质量百分比,随粒径减小总体变化趋势为先增加后减少再增加,马尾松-檵木混交林变化趋势为先增加后减少。在15~30 cm土层中(图1B),5种林分团聚体质量百分比随粒径减小总体变化趋势与0~15 cm土层基本相同。柏木纯林和近自然经营杉木纯林水稳性团聚体质量百分比显著高于其他3种林分(P<0.05)。2个土层中,柏木纯林、杉木纯林和近自然经营杉木纯林团聚体质量百分比最大值均在2.00~5.00 mm粒级内,最小值出现在0.25~0.50 mm。马尾松-檵木混交林最大值则出现在0.25~0.50 mm粒级内,最小值出现在大于5.00 mm。相关研究表明:直径小于0.25 mm的微团聚体是构成大团聚体的基础,在很大程度上决定了土壤团聚体的数量,土壤微团聚体的质量百分比和分布对土壤物理性质具有重要影响。由图1可知:在0~15 cm土层中,马尾松-檵木混交林小于0.25 mm团聚体粒级的质量百分比大于其他4种林分,为22.93%,近自然经营杉木纯林的质量百分比最低,为9.88%。15~30 cm土层中,小于0.25 mm的团聚体质量百分比马尾松-檵木混交林最高,为22.70%,柏木纯林最低,为9.77%。说明相比于柏木纯林、杉木纯林和马尾松-杉木混交林,马尾松-檵木混交林不利于土壤水稳性大团聚体的形成。
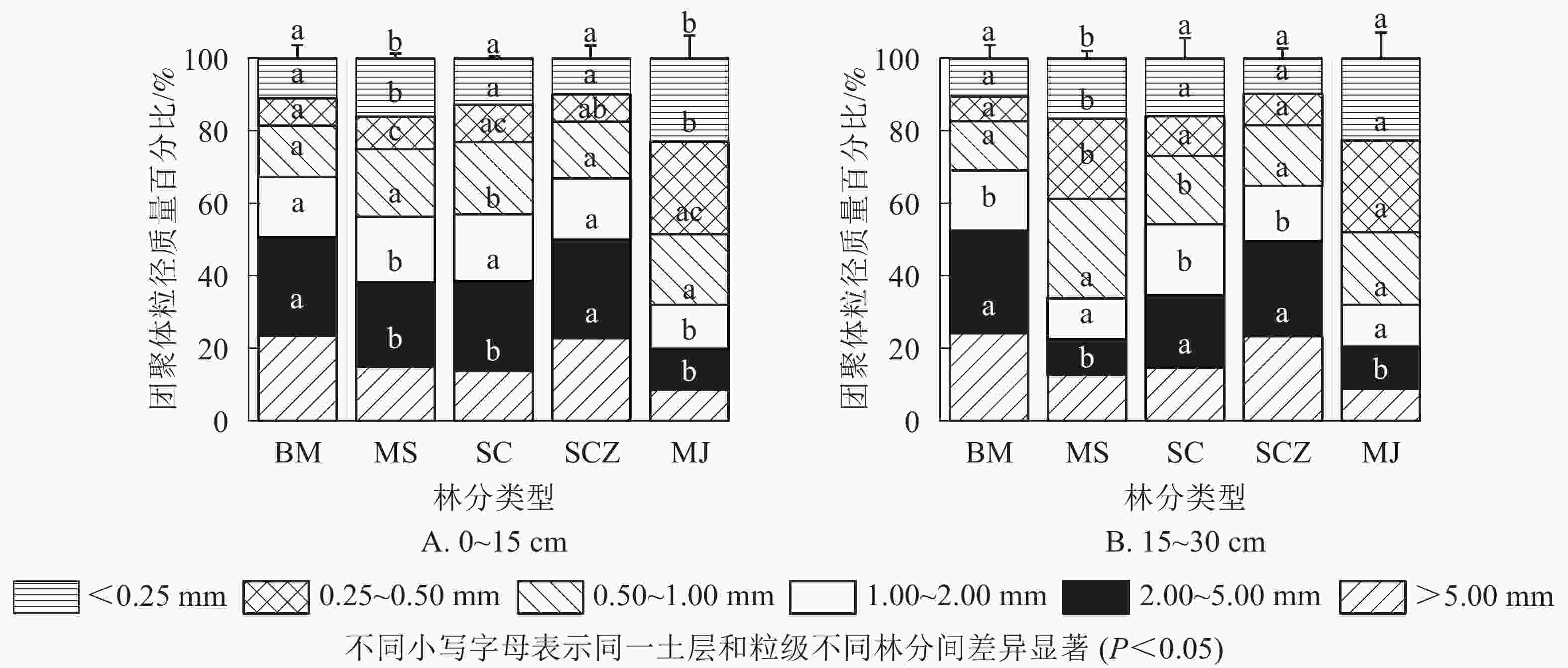
图 1 不同林分类型不同土层各粒径团聚体质量百分比
Figure 1. Mass percentage of aggregates in different soil layers of different forest types
-
由表2可知:2个土层中,大团聚体质量百分比基本在77%以上,在0~15 cm土层的平均质量直径(MWD)分布状况表现为柏木纯林最大,马尾松-檵木混交林最小。根据团聚体稳定性分级,柏木纯林和近自然经营杉木纯林为非常稳定,马尾松-檵木混交林为稳定,马尾松-杉木混交林、马尾松-檵木混交林和杉木纯林三者之间差异不显著。15~30 cm土层内平均质量直径表现为杉木纯林最大,表明杉木纯林土壤团聚体非常稳定,马尾松-檵木混交林土壤团聚体最小,表明马尾松-檵木混交林团聚体稳定。综合2个土层,马尾松-檵木混交林土壤水稳性团聚体稳定性最差,柏木纯林土壤水稳性团聚体最稳定,各林分表层和亚表层土壤水稳性团聚体平均质量直径变化规律不明显。
表 2 不同林分类型土壤水稳性团聚体稳定性
Table 2. Stability of soil water-stable aggregates of different forest types
土层深
度/cm林分类型 平均质量
直径/mm大团聚体
质量百分比/%土层深
度/cm林分类型 平均质量
直径/mm大团聚体
质量百分比/%0~15 柏木纯林 2.60±0.41 a 89.43±3.63 b 15~30 柏木纯林 2.06±0.12 a 88.14±4.65 a 马尾松-杉木混交林 1.43±0.56 b 80.75±7.98 a 马尾松-杉木混交林 2.37±0.13 b 85.19±3.78 a 马尾松-檵木混交林 1.29±0.09 b 77.09±4.28 a 马尾松-檵木混交林 1.77±0.11 a 81.24±1.40 ab 杉木纯林 1.70±0.32 b 82.93±7.27 ab 杉木纯林 2.49±0.19 b 93.72±1.44 a 近自然经营杉木纯林 2.51±0.68 a 90.12±3.51 b 近自然经营杉木纯林 1.81±0.12 a 87.54±5.34 a 说明:不同小写字母表示同一土层不同林分间差异显著(P<0.05) -
土壤可蚀性(K值)是土壤抗蚀能力大小的一个相对综合的指标。土壤可蚀性越大,土壤抗蚀能力越弱;土壤可蚀性越小,抗蚀能力越强。由图2可知:表层土壤的抗蚀能力均比亚表层强,且不同林分上下层土壤的变化趋势基本相同。2个土层中近自然经营杉木纯林和马尾松-杉木混交林的土壤可蚀性显著高于其他林分(P<0.05)。0~15 cm土层中,近自然经营杉木纯林土壤抗蚀能力最强,土壤可蚀性为0.023,马尾松-檵木混交林土壤抗蚀能力最弱,为0.048;15~30 cm土层中,近自然经营杉木纯林土壤可蚀性最小,为0.026,近自然经营杉木纯林土壤可蚀性最大,为0.050。综合2个土层可知:土壤抗蚀能力从大到小依次为近自然经营杉木纯林、柏木纯林、杉木纯林、马尾松-杉木混交林、马尾松-檵木混交林,这说明近自然经营模式能够提高林地土壤抗蚀性能。调查发现,柏木纯林林下灌草和苔藓植物丰富,密集的植被根系产生的胶结物质能够把小颗粒的土壤粘结成较大的土壤颗粒,从而使得柏木纯林比其他林分更有利于提高土壤抗蚀能力。
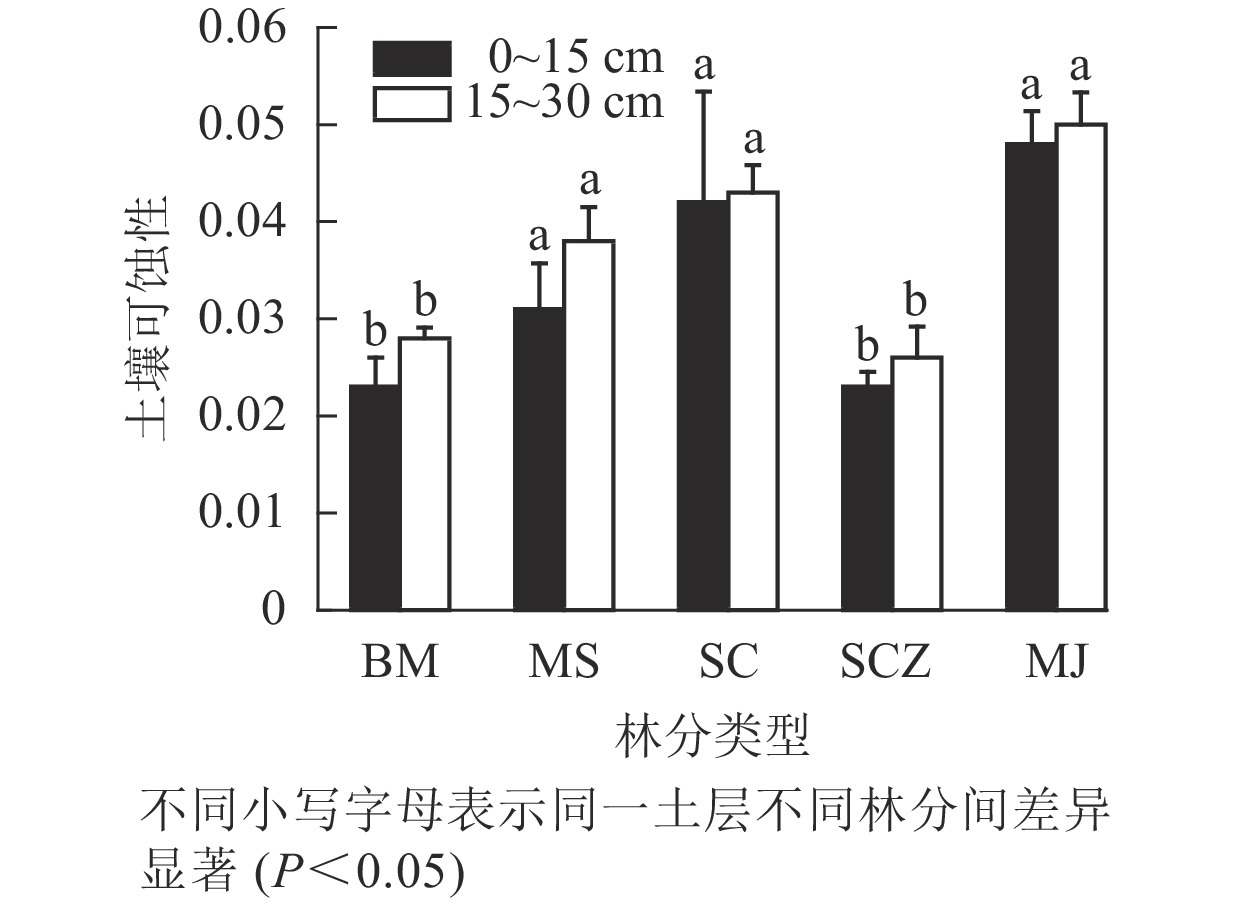
图 2 不同林分不同土层土壤可蚀性变化
Figure 2. Changes in K values of different forest types and different soil layers
-
表3可见:在0~15 cm土层不同林分土壤容重表现为近自然经营杉木纯林显著小于其他4种林分(P<0.05),而近自然经营杉木纯林的非毛管孔隙度显著大于另外4种林分,土壤有机碳质量分数在不同林分间表现为近自然经营杉木纯林显著大于柏木纯林、马尾松-杉木混交林和马尾松-檵木混交林(P<0.05),而近自然经营杉木纯林和杉木纯林之间无明显差异;不同林分速效钾质量分数从大到小依次为近自然经营杉木纯林、柏木纯林、杉木纯林、马尾松-檵木混交林、马尾松-杉木混交林,其中近自然经营杉木纯林和马尾松-杉木混交林差异显著(P<0.05)。15~30 cm土层内,近自然经营杉木纯林和杉木纯林的非毛管孔隙度显著大于其他3种林分(P<0.05),毛管孔隙度和总孔隙度表现为近自然经营杉木纯林显著大于柏木纯林(P<0.05),土壤有机碳质量分数表现为近自然经营杉木纯林显著大于马尾松-杉木混交林和马尾松-檵木混交林(P<0.05)。马尾松-杉木混交林的速效钾质量分数显著小于其他林分类型。毛管持水量在2个土层中表现为近自然经营杉木纯林显著大于其他4种林分类型,而速效磷质量分数在不同林分间差异不显著。
表 3 土壤基本理化性质
Table 3. Basic physical and chemical properties of soil
土层/
cm林分 容重/
(g·cm−3)非毛管孔
隙度/%毛管孔
隙度/%总孔
隙度/%毛管持水量/
(g·kg−1)土壤有机碳/
(g·kg−1)速效磷/
(mg·kg−1)速效钾/
(mg·kg−1)0~15 柏木纯林 1.30 a 3.00 b 44.89 a 47.89 a 345.98 b 32.38 b 31.30 a 97.56 ac 马尾松-杉木混交林 1.44 ab 3.76 b 34.48 a 38.23 a 255.31 b 29.76 b 35.01 a 70.94 b 马尾松-檵木混交林 1.46 a 4.96 b 33.42 a 38.38 a 230.80 b 30.17 b 29.88 a 76.84 ab 杉木纯林 1.20 a 4.37 b 38.93 a 43.31 a 313.08 b 40.74 ab 37.56 a 91.55 abc 近自然经营杉木纯林 0.93 b 7.61 a 47.25 a 54.85 a 505.79 a 52.74 a 25.37 a 117.65 a 15~30 柏木纯林 1.44 a 2.60 b 42.87 b 45.47 b 302.63 a 37.16 ab 29.00 a 92.92 a 马尾松-杉木混交林 1.45 a 3.69 b 38.30 ab 42.01 b 282.42 a 31.39 b 30.13 a 77.81 b 马尾松-檵木混交林 1.40 a 4.05 b 40.63 ab 44.68 b 290.84 a 33.20 b 28.51 a 89.58 a 杉木纯林 1.26 a 6.31 a 40.93 ab 47.24 b 331.51 a 43.13 ab 36.22 a 94.89 a 近自然经营杉木纯林 0.99 b 7.59 a 57.78 a 55.38 a 601.41 b 54.18 a 41.41 a 109.81 a 说明:不同小写字母表示同一土层不同林分间差异显著(P<0.05) -
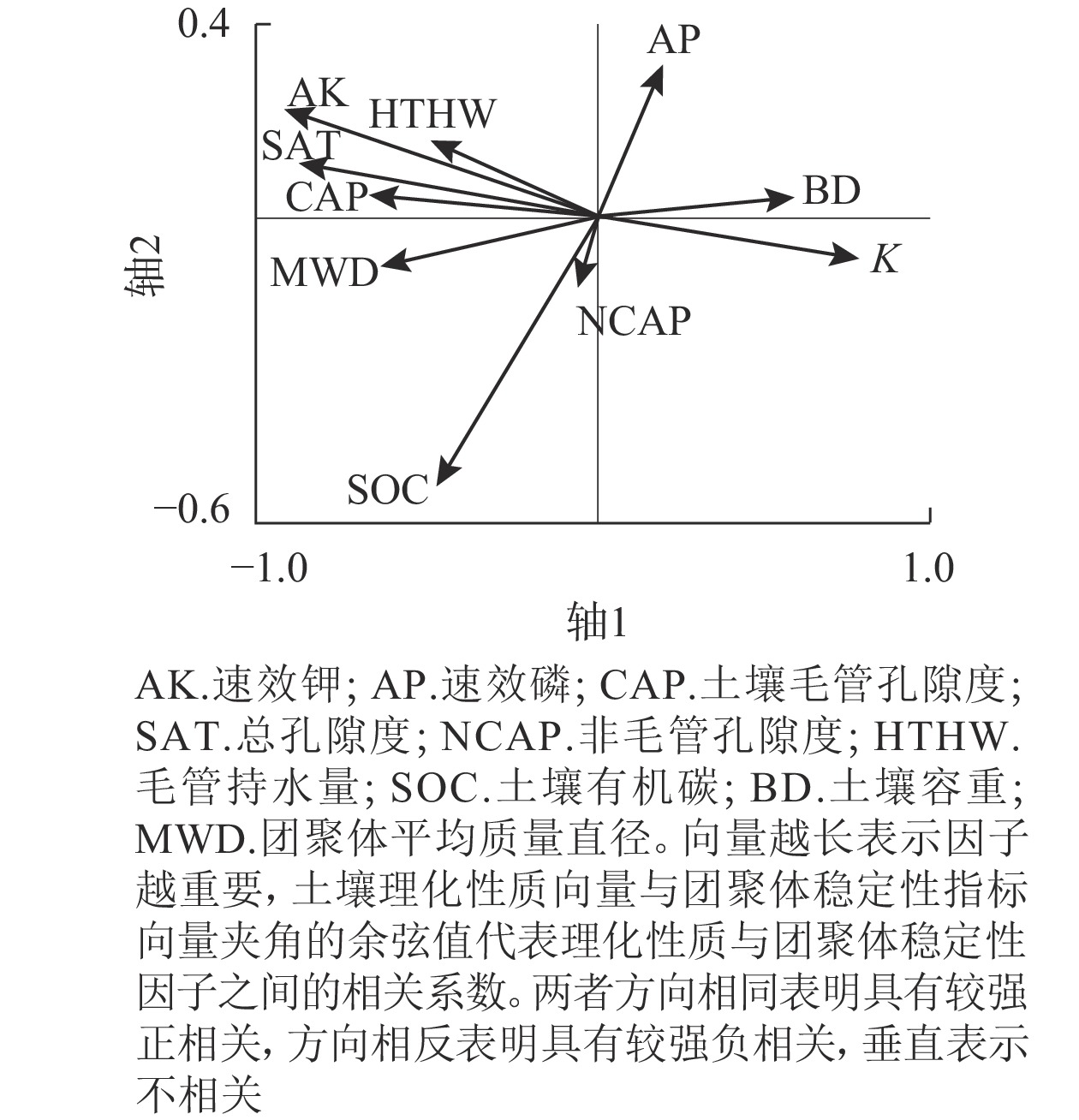
图3冗余分析(RDA)结果表明:研究区土壤理化性质对团聚体稳定性的累积解释量达95.64%。轴1土壤理化性质对团聚体稳定性的累积解释量为52.97%,前2个轴(轴1和轴2)土壤理化性质对团聚体稳定性的累积解释量为83.85%。土壤理化性质与团聚体稳定性的相关度累积解释量为99.89%。土壤有机碳、毛管孔隙度、总孔隙度、毛管持水量和速效钾与团聚体平均质量直径(MWD)之间呈显著正相关(P<0.05)。土壤容重与平均质量直径之间呈显著负相关(P<0.05)。土壤可蚀性因子与土壤容重呈显著正相关(P<0.05),与毛管孔隙度、总孔隙度、毛管持水量和速效钾显著负相关(P<0.05)。土壤有机碳、总孔隙度和速效磷相比其他土壤理化性质对土壤团聚体的影响更重要。水稳性团聚体稳定性指标平均质量直径与土壤可蚀性因子显著负相关(P<0.05)。土壤有机碳、总孔隙度和速效钾越大,土壤容重越小,土壤水稳性团聚体越稳定,土壤抗蚀能力越强。
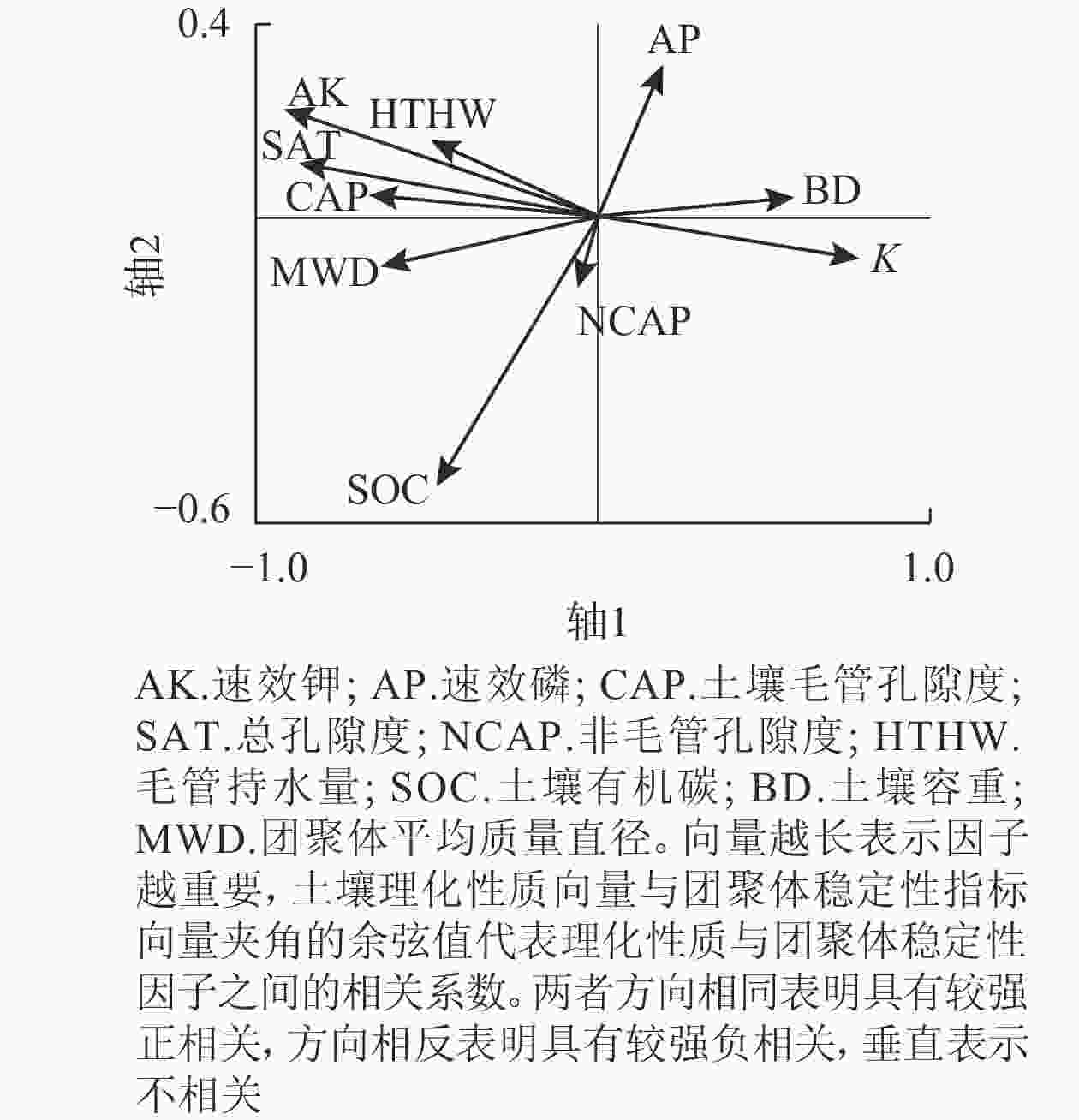
图 3 土壤理化性质与团聚体稳定性的RDA排序
Figure 3. RDA ranking of soil physical and chemical properties and aggregate stability
-
土壤团聚体的水稳定性与土壤侵蚀关系密切[17],水稳定性团聚体含量越大,表明土壤结构稳定性越好[18]。本研究表明:柏木纯林和近自然经营杉木纯林水稳性大团聚体质量百分比最高,马尾松-檵木混交林最低,说明马尾松-檵木混交不利于水稳性大团聚体的形成。土壤水稳性团聚体含量,特别是≥0.25 mm 团聚体含量是反映土壤抗蚀性的最佳指标之一[19]。本研究发现:5种林分的≥0.25 mm水稳性团聚体质量百分比均在77%以上,说明5种林分土壤结构较好,抗蚀能力强。直径<0.25 mm的微团聚体是构成团聚体的基础,土壤微团聚体质量百分比和分布对土壤物理性质具有重要影响。张文旭等[20]研究表明:近熟林转变为成熟林后,土壤团聚体中土壤大粒径微团聚体含量先增加后减少,小粒径微团聚体含量先减少后增加。本研究中,柏木纯林多为未郁闭的幼龄林,这可能是造成柏木纯林土壤团聚体粒级分布及其稳定性显著区别于其他林分的原因。此外,坡向与海拔通过分配降水和太阳辐射,影响植被分布和土壤理化性质,从而导致土壤抗蚀性的异质性[21]。本研究2个土层中马尾松-檵木混交林土壤微团聚体的质量百分比最大,这与吕宸等[22]对川西高寒草甸土壤团聚状况随海拔增加而降低的研究结论有所不同。这可能与植被类型和马尾松-檵木混交林在低海拔、半阳坡处分配降水和太阳辐射的差异,从而造成了植被分布和土壤理化性质的差异,进而导致土壤团聚状况的异质性有关。
-
土壤团聚体平均质量直径越大,表明土壤团聚体的团聚度高,稳定性强,土壤结构与质量越好[23]。团聚体的平均质量直径分级结果表明:柏木纯林和近自然经营杉木纯林土壤团聚体结构最稳定。本研究发现:土壤团聚体平均质量直径呈现近自然经营杉木纯林显著大于马尾松-檵木混交林,表明近自然经营杉木纯林土壤水稳性团聚体较其他4种人工林稳定,土壤抗侵蚀能力也较强。这是由于近自然经营杉木纯林与其他林分相比,郁闭度较高,凋落物较多。凋落物是产生有机质的重要来源,有机质能促进水稳定性团聚体含量和稳定性的增加[24]。而凋落物分解为微生物生存和繁殖提供了养分,这些微生物本身或生理活动的分泌物对土壤有胶结作用,可将较小的土壤颗粒粘聚成较大的团聚体,从而提高团聚体的稳定性[25]。此外,不同林地海拔高度和坡向的差异也会造成土壤团聚体稳定性和抗蚀性的差异,且阳坡由于强烈的冷热交替作用,土壤结构特征相较于阴坡随海拔高度的变化具有更加明显的分异特征[21]。本研究中,土壤团聚体稳定性和抗蚀性较强的近自然经营杉木纯林海拔高度最高,柏木纯林海拔最低,这说明海拔对土壤团聚体稳定性和抗蚀性的影响可能存在阈值。王进等[26]研究表明:原生灌木林地大团聚体(>0.25 mm)含量显著高于荒草地。这解释了柏木纯林郁闭度虽然不高,但是由于与其他林地相比柏木林地内原生灌木和苔藓丰富,植被覆盖度高,林地土壤水分较为充足,使得微生物群落大量繁殖。微生物通过分解作用形成多糖等胶结物质,从而促进了微小团聚体形成较大的团聚体[27]。丰富的林下灌草植被等会在土壤中产生大量的根系,根系所产生的分泌物可作为土粒团聚的胶结剂,配合须根的穿插和缠绕,提高了水稳性大团聚体(>1.00 mm)比例,从而增加水稳性团聚体稳定性[28]。
-
土壤团聚体平均质量直径和土壤可蚀性是表明团聚体稳定性的2个指标。土壤团聚体稳定性的大小除与土壤有机质、土壤含水量和土壤孔隙度有关外,还受到立地条件、植被类型和人类活动等环境的影响[29]。土壤有机碳质量分数与土壤团聚体平均质量直径呈显著正相关。该结果与聂富育等[30]的研究结论一致。土壤中的有机碳主要来源于凋落物和动植物残体的分解,分解产生的大量腐殖质对土壤颗粒具有粘结作用,而亚表层的土壤有机碳主要来源于表层有机碳的淋溶和迁移[31]。凋落物的数量与分解速率是不同林分间土壤有机碳存在差异的主要原因。王清奎等[32]研究表明:土壤容重及非毛管孔隙度相比其他土壤理化性质对土壤团聚体的影响更为重要。这与本研究的土壤容重、速效钾和总孔隙度更重要且土壤团聚体平均质量直径与土壤容重呈显著负相关关系,与速效钾和总孔隙度呈显著正相关的结论有所差异。原因可能是土壤理化性质对粒径较小的微团聚体影响更大,改变了团聚体的稳定性。土壤团粒和团粒之间孔隙较大有利于空气流通,团粒内部以持水毛管孔隙占绝对优势,这种孔隙状况为土壤水、肥、气、热的协调创造了良好的条件。土壤总孔隙度越大,容纳水分和空气的能力就会越强,有利于根系发育。速效钾是包括土壤中水溶态钾和交换性钾的总和,它的特点是能够直接被作物吸收利用,是作物获得高产的保证。此外,植被自身生长过程也会提高土壤中速效钾质量分数。速效钾质量分数的增加与促进团聚体的形成有很大的关系。总的来说,速效钾、土壤有机碳和土壤总孔隙度越大,土壤容重越小,水稳性团聚体稳定性越高,土壤抗蚀性能越好。
本研究通过分析传统经营模式林分与近自然经营模式林分下土壤理化性质变化对土壤水稳性团聚体和土壤抗蚀能力的影响,结合土壤可蚀性指标K值和土壤团聚体平均质量直径对不同林分土壤结构稳定性和抗蚀性进行整体评价,但忽略了地形和气候等立地条件因子的影响。此外,林下微生境和林分密度等也会对研究结果产生一定的影响。因此,后续研究可以结合生境类型和植被状况分析土壤理化性质对土壤团聚体稳定性和抗蚀性的影响,以期更全面地探究在近自然经营与传统经营模式下该地区森林土壤理化性质对团聚体稳定性和抗蚀性的作用机制。
-
5种林分间土壤水稳性团聚体稳定性差异显著。柏木纯林和近自然经营杉木纯林土壤团聚体的平均质量直径显著高于其他林分,且5种林分2个土层的土壤大团聚体质量百分比均为77%~93%,总体表明近自然经营杉木纯林和柏木纯林团聚体最稳定,而马尾松-檵木混交林团聚体稳定性最差。说明相对于杉木纯林和马尾松-杉木混交林,近自然经营林分比传统经营模式的林分土壤团聚体更稳定,而传统经营模式林地中柏木纯林土壤团聚体最稳定。
土壤理化性质对团聚体稳定性的累积解释量达95.64%。速效钾、土壤有机碳和土壤总孔隙度与土壤团聚体平均质量直径显著正相关,与土壤可蚀性呈负相关,土壤容重与可蚀性呈正相关,与土壤团聚体平均质量直径呈显著负相关。2个土层中,近自然经营杉木纯林的速效钾、土壤有机碳和土壤总孔隙度均高于其他林分,土壤容重均小于其他林分。说明近自然经营杉木纯林土壤结构较其他林分更好,土壤水稳性团聚体更加稳定,土壤抗蚀能力较强。
Soil aggregate stability and anti-erodibility of typical forest stands in Huaying mountain area
-
摘要:
目的 探究四川省华蓥市典型林分下土壤团聚体稳定性变化特征及其影响因素,为阐明该地区团聚体组成及其稳定性机制提供理论依据。 方法 选取华蓥市山区5种典型林分(柏木Cupressus funebris纯林、杉木Cunninghamia lanceolata纯林、马尾松Pinus massoniana-檵木Loropetalum chinense混交林、马尾松-杉木混交林、近自然经营杉木纯林)为研究对象,通过测定林分表层(0~15 cm)和亚表层(15~30 cm)土壤理化性质,分析土壤理化性质对团聚体变化特征和土壤抗蚀性的影响。 结果 ①5种典型林分土壤大团聚体质量百分比为77%~93%,且不同林分间平均质量直径(MWD)和土壤可蚀性存在显著差异(P<0.05),其平均质量直径柏木林最大(2.33 mm),马尾松-檵木混交林最小(1.53 mm)。土壤可蚀性从大到小依次为近自然经营杉木纯林、柏木林、杉木纯林、马尾松-杉木混交林、马尾松-檵木混交林。②冗余分析表明:速效钾、土壤总孔隙度和土壤有机碳与平均质量直径呈显著正相关(P<0.05),与土壤可蚀性呈显著负相关(P<0.05);土壤容重与平均质量直径呈显著负相关性(P<0.05),与土壤可蚀性呈显著正相关(P<0.05)。 结论 5种典型林分中,柏木林和近自然经营杉木纯林土壤水稳性团聚体最稳定、抗侵蚀性强,马尾松-檵木混交林土壤水稳性团聚体稳定性最低。传统经营模式下柏木林地土壤团聚体更稳定,土壤抗蚀性更强;近自然经营模式有利于土壤团聚体稳定性和土壤抗蚀性的提高。图3表3参32 Abstract:Objective The objective of this study is to explore the characteristics and influencing factors of soil aggregate stability under typical forest stands in Huaying City, Sichuan Province, so as to provide theoretical basis for elucidating the composition and stability mechanism of soil aggregates in this area. Method Five typical stands (Cupressus funebris pure forest, Cunninghamia lanceolata pure forest, Pinus massoniana-Loropetalum chinense mixed forest, P. massoniana-C. lanceolata mixed forest, and near natural C. lanceolata pure forest) were selected as the research objects. The soil physical and chemical properties of the surface layer (0−15 cm) and subsurface layer (15−30 cm) of the stands were measured to analyze the effects on aggregate change characteristics and soil erosion resistance. Result (1) The contents of soil macroaggregates in five typical stands were 77%−93%, with significant differences in mean weight diameter (MWD) and soil erodibility (K) among different stands (P<0.05). The MWD value of C. funebris forest was the largest (2.33 mm, and that of P. massoniana-L. chinense mixed forest was the smallest(1.53 mm). The K values of soil erodibility from large to small followed the order of near natural C. lanceolata pure forest, C. funebris forest, C. lanceolata pure forest, P. massoniana-C. lanceolata mixed forest, and P. massoniana-L. chinense mixed forest. (2) Redundancy analysis (RDA) showed that available potassium, total porosity and organic carbon were significantly positively correlated with MWD and negatively correlated with K value, and soil bulk density was significantly negatively correlated with MWD and positively correlated with K value. Conclusion Among the five typical stands, C. funebris and near natural C. lanceolata pure forest have the highest stability of soil water stable aggregates and strong erosion resistance, while P. massoniana-L. chinense mixed forest displays the lowest stability of soil water stable aggregates. Under the traditional management mode, the soil aggregate of C. funebris is more stable and soil anti-erodibility is stronger. The near natural management mode is conducive to the improvement of soil aggregate stability and soil anti-erodibility. [Ch, 3 fig. 3 tab. 32 ref.] -
Key words:
- stand /
- aggregate /
- soil physical and chemical properties /
- erosion resistance /
- average mass diameter
-
图 1 不同林分类型不同土层各粒径团聚体质量百分比
Figure 1 Mass percentage of aggregates in different soil layers of different forest types
图 2 不同林分不同土层土壤可蚀性变化
Figure 2 Changes in K values of different forest types and different soil layers
图 3 土壤理化性质与团聚体稳定性的RDA排序
Figure 3 RDA ranking of soil physical and chemical properties and aggregate stability
表 1 样地基本情况
Table 1. Basic situation of the sample plot
林分类型 纬度(N) 经度(E) 海拔/m 坡度/(°) 坡向/(°) 树高/m 胸径/cm 郁闭度 柏木纯林(BM) 30°25′17″ 106°49′59″ 518 25 西偏北10 1.80 1.98 未郁闭 马尾松-杉木混交林(MS) 30°26′15″ 106°50′43″ 1 100 31 东偏南10 15.60 31.38 0.76 近自然经营杉木纯林(SCZ) 30°18′06″ 106°47′45″ 1 409 33 北偏西10 14.30 20.51 0.85 马尾松-檵木混交林(MJ) 30°26′12″ 106°50′47″ 575 33 东偏南10 16.40 19.21 0.83 杉木纯林(SC) 30°18′28″ 106°47′47″ 1 133 33 东偏南70 15.80 32.37 0.72  下载: 导出CSV
下载: 导出CSV
表 2 不同林分类型土壤水稳性团聚体稳定性
Table 2. Stability of soil water-stable aggregates of different forest types
土层深
度/cm林分类型 平均质量
直径/mm大团聚体
质量百分比/%土层深
度/cm林分类型 平均质量
直径/mm大团聚体
质量百分比/%0~15 柏木纯林 2.60±0.41 a 89.43±3.63 b 15~30 柏木纯林 2.06±0.12 a 88.14±4.65 a 马尾松-杉木混交林 1.43±0.56 b 80.75±7.98 a 马尾松-杉木混交林 2.37±0.13 b 85.19±3.78 a 马尾松-檵木混交林 1.29±0.09 b 77.09±4.28 a 马尾松-檵木混交林 1.77±0.11 a 81.24±1.40 ab 杉木纯林 1.70±0.32 b 82.93±7.27 ab 杉木纯林 2.49±0.19 b 93.72±1.44 a 近自然经营杉木纯林 2.51±0.68 a 90.12±3.51 b 近自然经营杉木纯林 1.81±0.12 a 87.54±5.34 a 说明:不同小写字母表示同一土层不同林分间差异显著(P<0.05)
下载: 导出CSV
表 3 土壤基本理化性质
Table 3. Basic physical and chemical properties of soil
土层/
cm林分 容重/
(g·cm−3)非毛管孔
隙度/%毛管孔
隙度/%总孔
隙度/%毛管持水量/
(g·kg−1)土壤有机碳/
(g·kg−1)速效磷/
(mg·kg−1)速效钾/
(mg·kg−1)0~15 柏木纯林 1.30 a 3.00 b 44.89 a 47.89 a 345.98 b 32.38 b 31.30 a 97.56 ac 马尾松-杉木混交林 1.44 ab 3.76 b 34.48 a 38.23 a 255.31 b 29.76 b 35.01 a 70.94 b 马尾松-檵木混交林 1.46 a 4.96 b 33.42 a 38.38 a 230.80 b 30.17 b 29.88 a 76.84 ab 杉木纯林 1.20 a 4.37 b 38.93 a 43.31 a 313.08 b 40.74 ab 37.56 a 91.55 abc 近自然经营杉木纯林 0.93 b 7.61 a 47.25 a 54.85 a 505.79 a 52.74 a 25.37 a 117.65 a 15~30 柏木纯林 1.44 a 2.60 b 42.87 b 45.47 b 302.63 a 37.16 ab 29.00 a 92.92 a 马尾松-杉木混交林 1.45 a 3.69 b 38.30 ab 42.01 b 282.42 a 31.39 b 30.13 a 77.81 b 马尾松-檵木混交林 1.40 a 4.05 b 40.63 ab 44.68 b 290.84 a 33.20 b 28.51 a 89.58 a 杉木纯林 1.26 a 6.31 a 40.93 ab 47.24 b 331.51 a 43.13 ab 36.22 a 94.89 a 近自然经营杉木纯林 0.99 b 7.59 a 57.78 a 55.38 a 601.41 b 54.18 a 41.41 a 109.81 a 说明:不同小写字母表示同一土层不同林分间差异显著(P<0.05)
下载: 导出CSV
-
[1] 谢贤健, 张彬. 基于耦合关联分析的护岸植被恢复土壤抗蚀性综合评价[J]. 土壤, 2019, 51(3): 609 − 616. XIE Xianjian, ZHANG Bin. Comprehensive evaluation on recovery of soil anti-erodibility by revetment vegetation based on coupling relationship analysis [J]. Soil, 2019, 51(3): 609 − 616. [2] 雷贤斌, 蒋钵, 黄卫东, 等. 浅析华蓥市人工林近自然森林经营[J]. 四川林勘设计, 2015(2): 104 − 105. LEI Xianbin, JIANG Bo, HUANG Weidong, et al. Analysis of near natural forest management of plantation in Huaying City [J]. Sichuan For Explor Des, 2015(2): 104 − 105. [3] WANG Qing, WANG Dan, WEN Xuefa, et al. Differences in SOM decomposition and temperature sensitivity among soil aggregate size classes in a temperate grasslands[J]. PLoS One, 2015, 10(2). doi: 10.1371/journal.pone.0117033. [4] 黄晓强, 信忠保, 赵云杰, 等. 北京山区典型人工林土壤团聚体组成及其有机碳分布特征[J]. 水土保持学报, 2016, 30 (1): 236 − 243. HUANG Xiaoqiang, XIN Zhongbao, ZHAO Yunjie, et al. Soil aggregate composition and distribution characteristics of soil organic carbon of typical plantations in mountainous area of Beijing [J]. J Soil Water Conserv, 2016, 30 (1): 236 − 243. [5] DAS B, CHAKRABORTY D, SINGH V K, et al. Effect of integrated nutrient management practice on soil aggregate properties, its stability and aggregate-associated carbon content in an intensive rice-wheat system [J]. Soil Till Res, 2014, 136(C): 9 − 18. [6] 李嘉, 刘楠, 韩进斌, 等. 冻融对五台山典型林线植被土壤水稳性大团聚体的影响[J]. 干旱区资源与环境, 2020, 34(5): 140 − 146. LI Jia, LIU Nan, HAN Jinbin, et al. Effects of freeze-thaw cycles on soil water-stable macroaggregates in typical timberline vegetation of Wutai Mountain [J]. J Arid Land Resour Environ, 2020, 34(5): 140 − 146. [7] ZUO Fenglin, LI Xiaoyan, YANG Xiaofan, et al. Soil particle-size distribution and aggregate stability of new reconstructed purple soil affected by soil erosion in overland flow [J]. J Soil Sediment, 2020, 20(5): 272 − 283. [8] KRAUSE L, KLUMPP E, NOFZ I, et al. Colloidal iron and organic carbon control soil aggregate formation and stability in arable Luvisols[J]. Geoderma, 2020, 374. doi: 10.1016/j.geoderma.2020.114421. [9] OKOLO C C, GEBRESAMUEL G, AMANUEL A Z, et al. Accumulation of organic carbon in various soil aggregate sizes under different land use systems in a semi-arid environment[J]. Agric Ecosyst Environ, 2020, 297. doi: 10.1016/j. agee. 2020.106924. [10] 谢钧宇, 曹寒冰, 孟会生, 等. 不同施肥措施及施肥年限下土壤团聚体的大小分布及其稳定性[J]. 水土保持学报, 2020, 34(3): 274 − 281, 290. XIE Junyu, CAO Hanbing, MENG Huisheng, et al. Effects of different fertilization regimes and fertilization ages on size distribution and stability of soil aggregates [J]. J Soil Water Conserv, 2020, 34(3): 274 − 281, 290. [11] 赵友朋, 孟苗婧, 张金池, 等. 凤阳山主要林分类型土壤团聚体及其稳定性研究[J]. 南京林业大学学报(自然科学版), 2018, 42(5): 84 − 90. ZHAO Youpeng, MENG Miaojing, ZHANG Jinchi, et al. Study on the composition and stability of soil aggregates of the main forest stands in Fengyang Mountain, Zhejiang Province [J]. J Nanjing For Univ Nat Sci Ed, 2018, 42(5): 84 − 90. [12] 程欢, 宫渊波, 付雨欣, 等. 四川盆地西南缘不同林分类型土壤团聚体稳定性及有机碳组分特征[J]. 水土保持学报, 2018, 32 (5): 109 − 115. CHENG Huan, GONG Yuanbo, FU Yuxin, et al. Soil aggregate stability and characteristics of organic carbon components in three forests of the southwest edge of Sichuan Basin [J]. J Soil Water Conserv, 2018, 32 (5): 109 − 115. [13] 张万儒, 杨光滢, 屠星南. 森林土壤分析方法[M]. 北京: 中国标准出版社, 2000. [14] BISSONNAIS Y, PRIETO I, ROUMET C, et al. Soil aggregate stability in Mediterranean and tropical agro-ecosystems: effect of plant roots and soil characteristics [J]. Plant Soil, 424(1/2). doi: 10.1007/s11104-017-3423-6. [15] 张科利, 彭文英, 杨红丽. 中国土壤可蚀性值及其估算[J]. 土壤学报, 2007, 44(1): 7 − 13. ZHANG Keli, PENG Wenying, YANG Hongli. Soil erodibility and its estimation for agricultural soil in China [J]. Acta Pedol Sin, 2007, 44(1): 7 − 13. [16] LE BISSONNAIS Y. Aggregate stability and assessment of soil crustability and erodibility (Ⅰ) Theory and methodology [J]. Eur J Soil Sci, 1996, 47(1): 425 − 437. [17] 卢金伟. 土壤团聚体水稳定性及其与土壤可蚀性之间关系研究[D]. 杨凌: 西北农林科技大学, 2002. LU Jinwei. Water Stability of Soil Aggregates and Its Relationship with Soil Erodibility[D]. Yangling: Northwest A&F University, 2002. [18] CHAPLOT V, COOPER M. Soil aggregate stability to predict organic carbon outputs from soils [J]. Geoderma, 2015, 243: 205 − 213. [19] 谈正鑫. 盱眙月亮山典型人工林水土保持功能研究[D]. 南京: 南京林业大学, 2015. TAN Zhengxin. Research on Soil and Water Conservation Function of Artificial Forest in Xuyi Moon Mountains[D]. Nanjing: Nanjing Forestry University, 2015. [20] 张文旭, 李建红, 郭灵辉, 等. 太行山油松人工林土壤微团聚体变化特征及其影响因素[J]. 山地学报, 2019, 37(6): 797 − 807. ZHANG Wenxu, LI Jianhong, GUO Linghui, et al. Change features of soil micro-aggregates of Pinus tabulaeformis plantation at different developmental stages in the southern taihang mountain and its relationship with soil nutrients [J]. J Mt Sci, 2019, 37(6): 797 − 807. [21] 黄瑞灵, 周华坤, 刘泽华, 等. 坡向与海拔对青海省拉鸡山不同植被土壤种子库的影响[J]. 生态学杂志, 2013, 32(10): 2679 − 2686. HUANG Ruiling, ZHOU Huakun, LIU Zehua, et al. Effects of slope aspect and altitude on the soil seed bank under different vegetations in Laji Mountains of Qinghai [J]. Chin J Ecol, 2013, 32(10): 2679 − 2686. [22] 吕宸, 宫渊波, 车明轩, 等. 川西高寒山地灌丛草甸土壤抗蚀性研究[J]. 水土保持学报, 2020, 34(2): 9 − 17. LÜ Chen, GONG Yuanbo, CHE Mingxuan, et al. Study on the soil erosion resistance of alpine shrub meadow in western Sichuan [J]. J Soil Water Conserv, 2020, 34(2): 9 − 17. [23] 郑子成, 李延轩, 张锡洲, 等. 不同土地利用方式下土壤团聚体的组成及稳定性研究[J]. 水土保持学报, 2009, 23(5): 228 − 231. ZHENG Zicheng, LI Yanxuan, ZHANG Xizhou, et al. Study on the composition and stability of soil aggregates under different land use [J]. J Soil Water Conserv, 2009, 23(5): 228 − 231. [24] 冯嘉仪, 储双双, 王婧, 等. 华南地区几种典型人工林土壤有机碳密度及其与土壤物理性质的关系[J]. 华南农业大学学报, 2018, 39(1): 83 − 90. FENG Jiayi, CHU Shuangshuang, WANG Jing, et al. Soil organic carbon density and its relationship with soil physical properties of typical plantations in South China [J]. J South China Agric Univ, 2018, 39(1): 83 − 90. [25] 龙健, 赵畅, 张明江, 等. 不同坡向凋落物分解对土壤微生物群落的影响[J]. 生态学报, 2019, 39(8): 2696 − 2704. LONG Jian, ZHAO Chang, ZHANG Mingjiang, et al. Effect of litter decomposition on soil microbes on different slopes [J]. Acta Ecol Sin, 2019, 39(8): 2696 − 2704. [26] 王进, 刘子琦, 鲍恩俣, 等. 喀斯特石漠化区林草恢复对土壤团聚体及其有机碳含量的影响[J]. 水土保持学报, 2019, 33(6): 249 − 256. WANG Jin, LIU Ziqi, BAO Enyu, et al. Effects of forest and grass restoration on soil aggregates and its organic carbon in Karst Rocky Desertification Areas [J]. J Soil Water Conserv, 2019, 33(6): 249 − 256. [27] OADES J M. Soil organic matter and structural stability mechanisms and implications for management [J]. Plant Soil., 1984, 76(1/3): 319 − 337. [28] 宋日, 刘利, 马丽艳, 等. 作物根系分泌物对土壤团聚体大小及其稳定性的影响[J]. 南京农业大学学报, 2009, 32(3): 93 − 97. SONG Ri, LIU Li, MA Liyan, et al. Effects of crop root exudates on the size and stability of soil aggregate [J]. J Nanjing Agric Univ, 2009, 32(3): 93 − 97. [29] 王小云. 土壤团聚体与土壤侵蚀关系研究进展[J]. 安徽农业科学, 2016, 44(23): 106 − 108. WANG Xiaoyun. Research progress of the relationship between soil aggregates and soil erosion [J]. Anhui Agric Sci, 2016, 44(23): 106 − 108. [30] 聂富育. 四川盆地西缘4种人工林土壤团聚体及氮动态特征[D]. 成都: 四川农业大学, 2017. NIE Fuyu. Soil Aggregates and Labile Nitrogen Dynamics in 4 Plantations in the Western Edge of Sichuan Basin [D]. Chengdu: Sichuan Agricultural University, 2017. [31] 张琪, 方海兰, 史志华, 等. 侵蚀条件下土壤性质对团聚体稳定性影响的研究进展[J]. 林业科学, 2007, 43(增刊 1): 77 − 82. ZHANG Qi, FANG Hailan, SHI Zhihua, et al. Advances in influence factors of aggregate stability under erosion [J]. Sci Silv Sin, 2007, 43(suppl 1): 77 − 82. [32] 王清奎, 汪思龙, 高洪, 等. 土地利用方式对土壤有机质的影响[J]. 生态学杂志, 2005, 24(4): 360 − 363. WANG Qingkui, WANG Silong, GAO Hong, et al. Influence of land use on soil organic matter [J]. Chin J Ecol, 2005, 24(4): 360 − 363. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210142
 点击查看大图
点击查看大图
计量
- 文章访问数: 2455
- HTML全文浏览量: 664
- PDF下载量: 53
- 被引次数: 0
