






-
农业面源污染已成为全球许多国家流域水污染和水体富营养化的首要污染来源[1-2]。近年来,国内诸多流域尺度的污染源调查解析表明:中国许多河湖、水库等地表水体中的氮磷输入主要来自农田种植、畜禽水产养殖等农业生产以及农村生活所产生的非点源污染,其中种植业所引起的农田退水污染正逐步成为污染的主要贡献者[3-5]。农田退水污染具有排放时间瞬时多变,迁移途径复杂多样,污染负荷时空变化幅度大等特点,治理难度相对更大[1-2]。目前基于生态工程原理的生态沟渠、人工湿地等技术受到格外重视[6-8]。此类通常建立在培植大型水生植物基础上的生态拦截工程具有建设成本低且可控、技术难度低、生态友好等优点,近年来在中国许多地区特别是生态敏感区域得到了较大规模的推广应用。2018年以来,浙江省在全省范围内实施了300项农田面源污染氮磷生态拦截沟渠示范推广项目[9]。研究已表明:水生植物对包括氮磷营养物质在内的众多污染物的净化效率及其稳定性具有重要的影响[10-11]。沉水植物占据了根系-基质(根际)、水-基质和植物茎叶-水3个关键微界面,由其介导形成的微环境结构更为复杂,往往能够表现出更加稳定和持续的水质净化潜力[12-14]。近几年以沉水植物为先锋物种的“水下植被”技术拓展应用到农业面源污染的拦截处理[15-17]。目前,国内外对挺水植物型人工湿地去污动力学已开展了比较深入的研究[18-20],但对“水下植被”型净化系统污染物去除动力学特征还十分欠缺,特别是对拦截净化农田退水、池塘养殖尾水等具有瞬间排放特点的非点源污染的除磷动力学研究至今还未见报道。在动力学模型应用研究方面,一级动力学、Grau二级动力学和Monod动力学因具有模型构式可线性化处理、参数求解过程简单、适用范围广等优点而被广泛采用[19-22];近年来,修正的Gompertz模型作为一种多参数非线性模型在污染物去除动力学研究领域逐步被重视,并且已被成功应用于新型生物处理系统的工艺设计和污染物去除效率的预测[23-25]。本研究以苦草Vallisneria natans和金鱼藻Ceratophyllum demersum等2种典型沉水植物为供试材料,研究苦草型和金鱼藻型2种“水下植被”型净化系统对农田退水中磷污染的净化效率。在此基础上,采用一级动力学、Grau二级动力学、Monod 动力学和修正的Gompertz模型模拟总磷的去除过程并进行统计验证,获取相关动力学参数,进而探讨2种沉水植物净化系统除磷效率的差异,以期为“水下植被”型生态净化系统的工艺调控及其设计应用提供理论依据和技术参考。
-
选择不同生长型的沉水植物苦草和金鱼藻作为供试植物,这2种植物均为长江中下游流域常见的优势沉水植物,偏好于富营养化水体,在溪沟、河流、池塘、湖泊等水域中均有分布。2种供试植物均取自浙江农林大学乌镇大学生实践基地。采集时选取生长状态良好且株高相近的植株,运至实验基地培养10 d,统一修剪至株高25 cm,去离子水清洗干净后用作试验材料。
受降雨强度、降雨历时、施肥量、土壤类型、土地利用类型等因素影响,农田退水中总磷质量浓度波动极大,低值时可低于地表水Ⅳ类标准,而峰值时高达5.00 mg·L−1以上[26-27]。本研究人工配置中,农田退水总磷质量浓度处于相对较高水平,试验原水取自浙江农林大学平山实验基地内的池塘,原水中总磷质量浓度为 0.17 mg·L−1,铵态氮(NH3-N)质量浓度为0.39 mg·L−1,重铬酸盐指数(CODcr)为8.56 mg·L−1。经配置后的试验用水总磷质量浓度为4.17 mg·L−1。
-
考虑到农田退水排放的瞬间性、间歇性等特征以及池塘、相对封闭河道等受纳水体的水动力特点,本研究采用批式试验考察2种供试植物净化系统对农田退水总磷的去除效果及其动力学特征。试验共设置3组,分别为苦草组、金鱼藻组和无植物对照组,每组均重复3次。试验于浙江农林大学平山实验基地温室大棚内进行,试验装置采用白色聚乙烯塑料箱(长×宽×高为40 cm×80 cm×60 cm),各塑料箱内注入试验用水,试验水深50 cm,计为160 L。每个塑料箱底部铺设1层厚度为10 cm的火山石基质(过10目筛)。为避免植物直接定植在基质上导致基质分布不均,先将经预培养的供试植物(鲜质量300 g)定植于渗排水网垫网上(厚度为5 mm),然后再铺设于基质层上。
试验周期共计49 d (2018年7—8月),试验期间平均最高气温为34.0 ℃,平均最低气温为25.0 ℃,隔1周采样1次。采用虹吸管抽取方式采集水样,分别于水面以下15和25 cm处采集等体积水样组成混合样(100 mL)。每次取样后添加去离子水以弥补蒸发、蒸腾以及取样所带来的水损耗。水样及试验用水总磷采用过硫酸钾氧化-钼锑抗分光光度法测定。
-
分别利用一级动力学[20]、Grau二级动力学[18]、Monod动力学[20]和修正的Gompertz模型[28]对批式试验数据进行拟合分析。
-
总磷去除负荷表达式为:
$$ \sum\limits_{t = 0}^t {C'} = \frac{{\left( {{C_0} - {C_t}} \right)V}}{{L W}}{\text{。}} $$ (1) 式(1)中:t为反应时间(d);C0为总磷初始质量浓度(mg·L−1);Ct为反应时间t时的总磷质量浓度(mg·L−1);C′为总磷累计去除负荷(mg·m−2);V为试验用水体积(L);L为试验装置长度(m);W为试验装置宽度(m)。
-
一级动力学模型通用表达式在边界条件下积分处理为:
$$\ln \left( {\frac{{{C_0}}}{{{C_t}}}} \right) = {F_{\rm{a}}}t{\text{。}}$$ (2) 式(2)中:Fa为总磷一级动力学速率常数(d−1)。
-
Grau二级动力学通用表达式边界条件下积分并线性化处理为:
$$\frac{{{C_0}t}}{{{C_0} - {C_t}}} = t + \frac{{{C_0}}}{{{S_{\rm{a}}}X}}{\text{。}}$$ (3) 令:m=C0/SaX,则式(3)可整理成:
$$\frac{{{C_0}t}}{{{C_0} - {C_t}}} = m + nt{\text{。}}$$ (4) 式(4)中:m和n为Grau二级动力学常数。其中:n为接近1的无量纲常数;m代表总磷去除效率(d),其值越大,去除效率越低。
-
Monod动力学通用表达式经积分和线性化处理为:
$${C_0} - {C_t} + {C_{\rm{h}}}\ln \frac{{{C_0}}}{{{C_t}}} = {M_{\max }}t{\text{。}}$$ (5) 式(5)中:Mmax为Monod动力学总磷最大体积去除速率常数(mg·m−3·d−1);Ch为Monod动力学总磷半饱和速率常数(mg·L−1),取0.2 mg·L−1[22]。
-
Gompertz模型经修正可用于描述污染物质量浓度与处理时间的定量关系。表达式为[28]:
$${C_t} = {C_0} - {C_0}\exp \left\{ { - \exp \left[ {\frac{{{\rm{e}} {G_{{\rm{max}}}}}}{{{C_0}}}\left( {{t_{\rm{a}}} - t} \right) + 1} \right]} \right\}{\text{。}}$$ (6) 式(6)可转化为:
$$R_{\rm{E}} = 100 \times \exp \left\{ { - \exp \left[ {\frac{{{\rm{e}} {G_{{\rm{max}}}}}}{{{C_0}}}\left( {{t_{\rm{a}}} - t} \right) + 1} \right]} \right\}{\text{。}}$$ (7) 式(7)中:RE为总磷去除率(%);Gmax为Gompertz模型总磷最大体积去除速率常数(mg·m−3·d−1);ta为迟滞时间(d);e为数学常数。
-
采用Excel和SPSS进行数据统计与分析(显著水平设为P<0.05),采用Origin 9.1软件进行模型创建、拟合和图形绘制。图表中所列数据均为3个重复的平均值。引入判定系数(R2)和相对均方根误差(RRMSE)评价动力学模型的准确性和适用性。其中:R2反映2组数据线性回归拟合优度,该值接近于1(取值为0~1)表明两者之间高度线性相关;RRMSE反映模型预测值与试验实测值之间的整体偏差,RRMSE越小(取值为0~∞),表明预测值与实测值间越接近。各统计参数计算公式为:
$${R^2} = \frac{{\displaystyle \sum\limits_{i = 1}^N {{{\left[ {\left( {{X_i} - \overline X} \right)\left( {{Y_i} - \overline Y} \right)} \right]}^2}} }}{{\displaystyle \sum\limits_{i = 1}^N {{{\left( {{X_i} - \overline X} \right)}^2}} \displaystyle \sum\limits_{i = 1}^N {{{\left( {{Y_i} - \overline Y} \right)}^2}} }};$$ (8) $$R_{\rm{RMSE}} = \frac{{\sqrt {\dfrac{1}{N}\displaystyle \sum\limits_{i = 1}^N {{{\left( {{Y_i} - {{\hat Y}_i}} \right)}^2}} } }}{{\overline Y}}{\text{。}}$$ (9) 式(8)~(9)中:Xi和Yi分别为2组数据(反应时间和试验实测值),
$\overline X$ 和$\overline Y$ 分别为2组数据的平均值;${\hat Y_i}$ 为动力学模型预测值;N为样本数。 -
随着处理时间的延长,2组植物净化系统总磷累计去除负荷呈逐渐增加趋势(图1)。至试验结束时(第49 天),苦草组和金鱼藻组净化系统的累计去除负荷分别为1 151.17和1 167.50 mg·m−2,对应总磷去除率分别为82.8%和84.0%。方差分析表明:试验结束时2组植物系统总磷去除效率无显著差异(P>0.05),但均显著好于无植物的对照组(P<0.05)。进一步对各周总磷去除负荷分析发现(图2):2组植物系统各周之间的总磷去除负荷均存在较大波动,并且均在第3周出现峰值,单周贡献了整个试验期间(共7周)总磷累计去负荷的28.5%(苦草组)和16.7%(金鱼藻组)。无植物的对照组总磷去除负荷在第1周出现峰值。
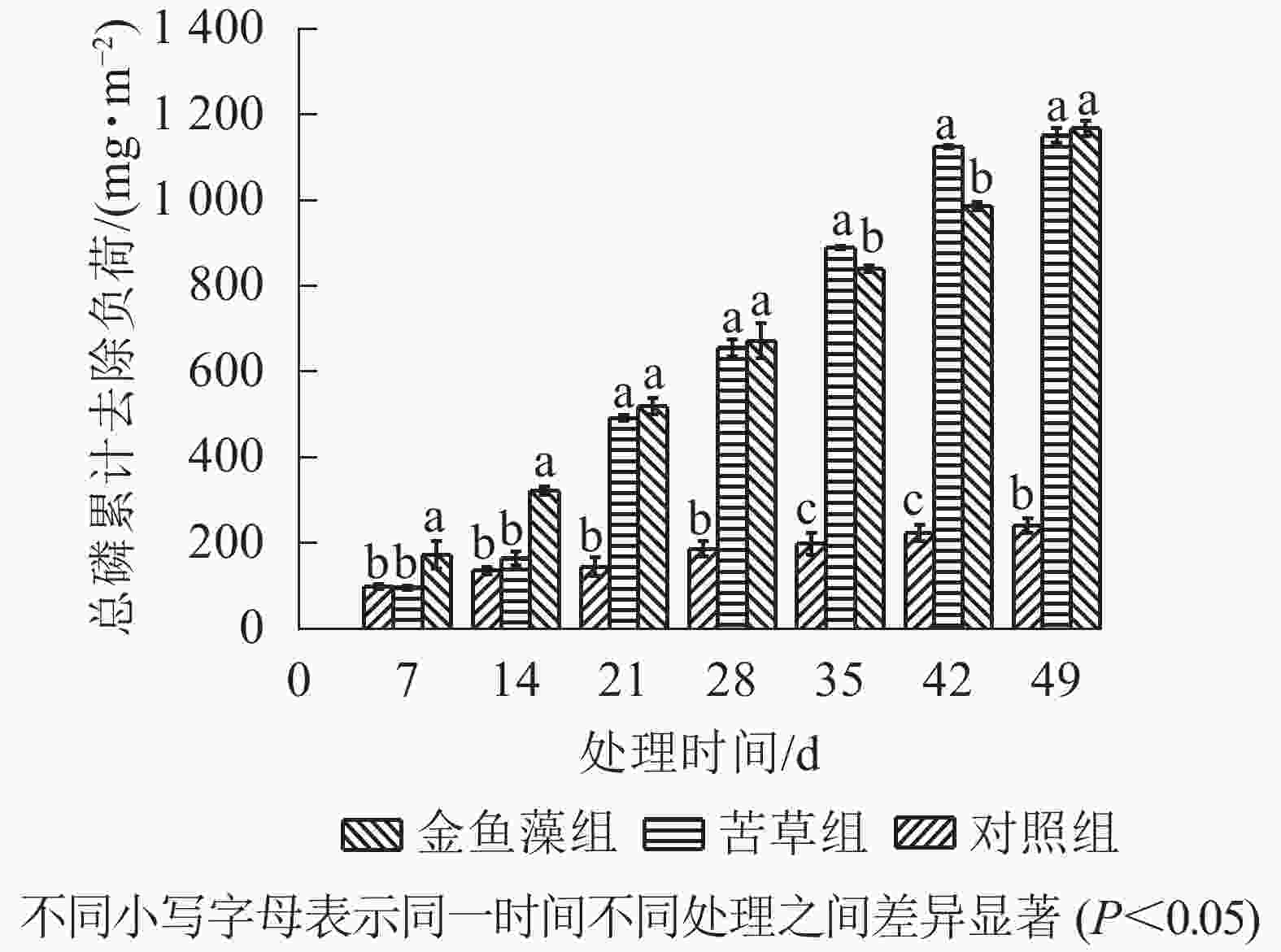
图 1 沉水植物净化系统总磷累计去除负荷随处理时间的变化
Figure 1. Total phosphorus removal loading over treatment time in the submerged plant treatment systems

图 2 沉水植物净化系统各周总磷去除负荷
Figure 2. Total phosphorus weekly removal loading for the submerged plant treatment systems
在不同时间尺度上比较2组植物净化系统总磷去除效率发现(图1):尽管在试验结束时(第49 天)和试验中期(第21 天和第28 天)两者差异不显著,但在试验初期(第7 天和第14 天)和中后期(第35 天和第42 天)2组植物净化系统总磷去除效率存在差异显著(P<0.05)。其中试验初期,金鱼藻组净化系统总磷去除效率显著高于苦草组净化系统;而当处理时间增至35和42 d时,苦草组又显著高于金鱼藻组。这种不同沉水植物系统除磷效率的差异在不同时间尺度上出现偏离甚至逆转的现象可能具有一定的普通性,如文献[29]、文献[30]和文献[31]的研究中均存在这种现象。值得注意的是,忽视植物效应差异的不确性,可能会导致水生植物净化系统工艺优化,特别是水生植物的筛选结果出现偏离甚至失真,因此,有必要借助污染物去除动力学模拟分析获取用于指示和预测污染物去除效率的相关特征参数。
-
分别运用一级动力学[公式(2)]和Grau二级动力学[公式(4)]模型对2组沉水植物净化系统试验数据进行拟合,并进行模型统计验证分析,结果见图3和表1。一级动力学模型具有较高的拟合度(判定系数R2>0.930),模型预测值与实测值之间吻合程度较好(相对均方根误差RRMSE<0.200),表明一级动力学模型可用来描述沉水植物净化系统批式运行周期内总磷的去除过程。2组净化系统回归求得的总磷去除一级动力学常数(Fa)分别为0.039 d−1(苦草组)和0.035 d−1(金鱼藻组)。不同于一级动力学,Grau二级动力学模型拟合度较差(R2<0.600),表明该模型不适用于模拟本试验沉水植物净化系统总磷的去除过程,获取的动力学参数(n和m)也不具有现实意义。

图 3 2种沉水植物净化系统总磷去除一级动力学模型拟合曲线
Figure 3. Regression of first-order kinetic model for TP removal over time in the submerged plant treatment systems
表 1 2种沉水植物净化系统总磷去除一级动力学和Grau二级动力学拟合结果
Table 1. Fitting result of First-order and Grau second-order kinetic models for total phosphorus removal in the submerged plant treatment systems
沉水植物型生态净化系统 一级动力学模型 Grau二级动力学模型 Fa/d−1 R2 RRMSE m/d n R2 RRMSE 苦草组 0.039 0.931 0.196 109.90 −1.35 0.588 0.324 金鱼藻组 0.035 0.937 0.169 57.25 0.03 0.081 0.127 一级动力学模型已被广泛应用于挺水植物型人工湿地系统中有机物、营养物、悬浮物等污染物去除效果的预测[19-22],但该模型是否适用于模拟沉水植物型净化系统的污染物去除动力学至今还未得到证实。本研究发现:一级动力学模型可用于描述沉水植物型净化系统在批式模式下的总磷去除过程。
一级动力学常数(Fa)代表着反应器系统在相对稳定运行状态下的去污能力,该参数不仅可用于反应器放大设计所需有效体积或面积的测算,还可以用于处理系统的工艺调控和比较优化。如在同等污染物质量浓度水平下,Fa越大,达到同一出水质量浓度所需的处理时间越短,即污染物的去除效率越高。方差分析结果表明:2组沉水植物系统的总磷去除一级动力学常数Fa存在显著差异(P<0.05),说明植物种类对总磷的净化效率产生显著影响。基于Fa及其物理意义可知:试验条件下苦草组净化系统的总磷去除效率显著好于金鱼藻组净化系统。
-
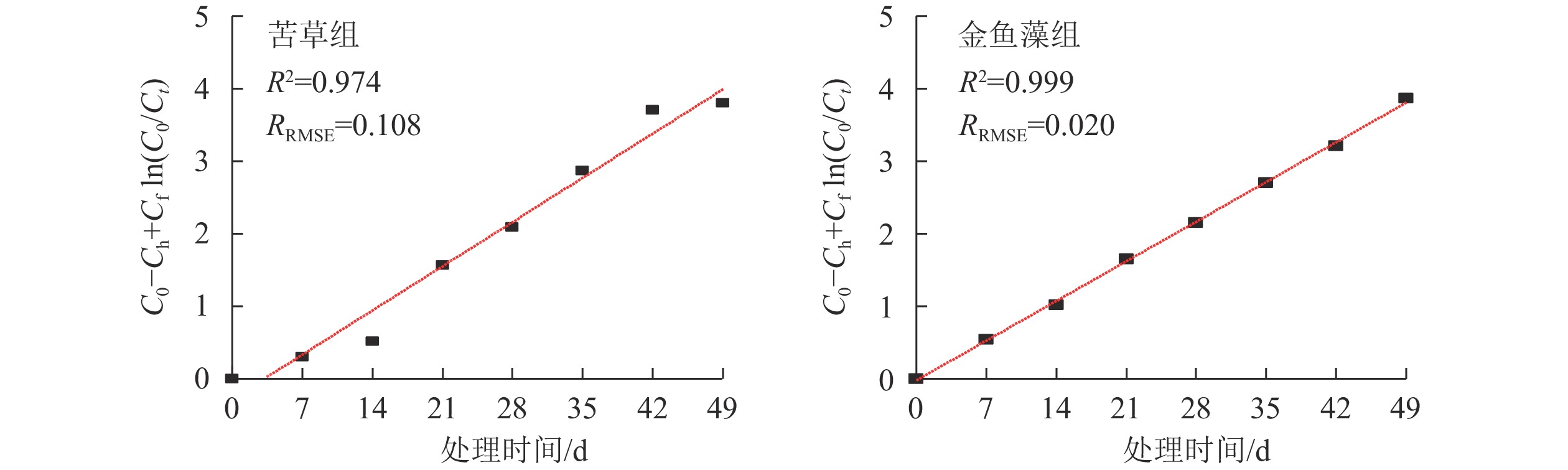
图4为Monod动力学模型f(C0, Ct, Ch)与净化处理时间(t)之间的回归曲线,表2列出了由回归系数求得的总磷最大体积去除速率常数(Mmax)以及统计验证参数。结果表明:Monod动力学模型可以较好地拟合2组沉水植物系统总磷的去除过程(R2>0.970,RRMSE<0.110)。2组植物系统拟合得到的总磷最大体积去除速率分别为87.14 mg·m−3·d−1(苦草组)和78.15 mg·m−3·d−1(金鱼藻组),并且差异达到显著性水平(P<0.05)。
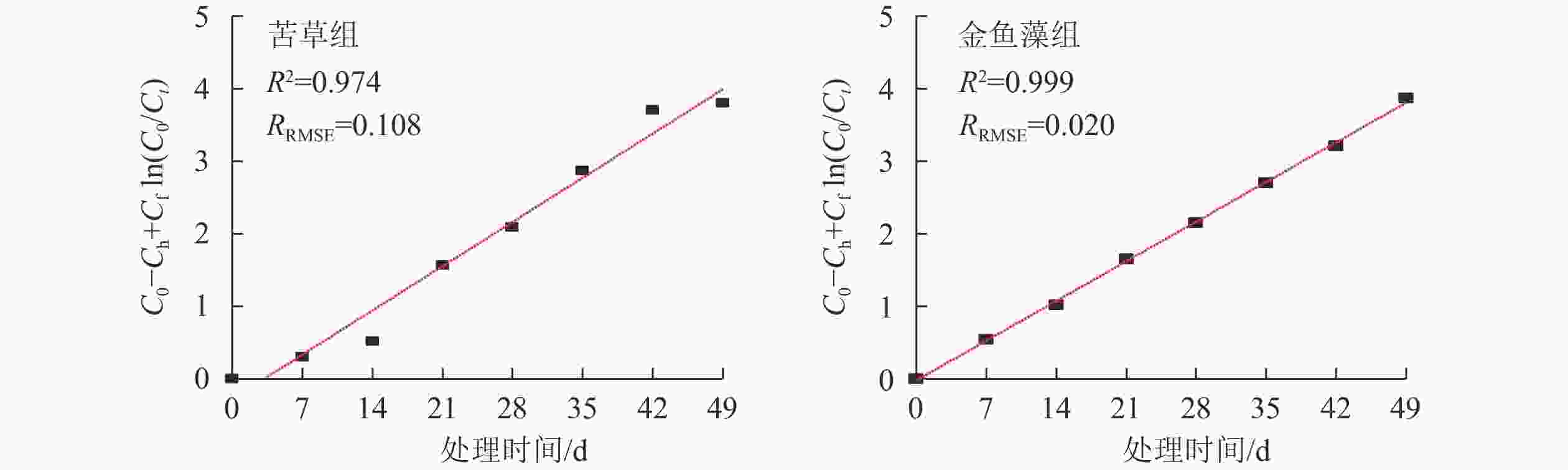
图 4 2种沉水植物净化系统总磷去除Monod动力学拟合曲线
Figure 4. Regression of Monod kinetic model for total phosphorus removal over time in the submerged plant treatment systems
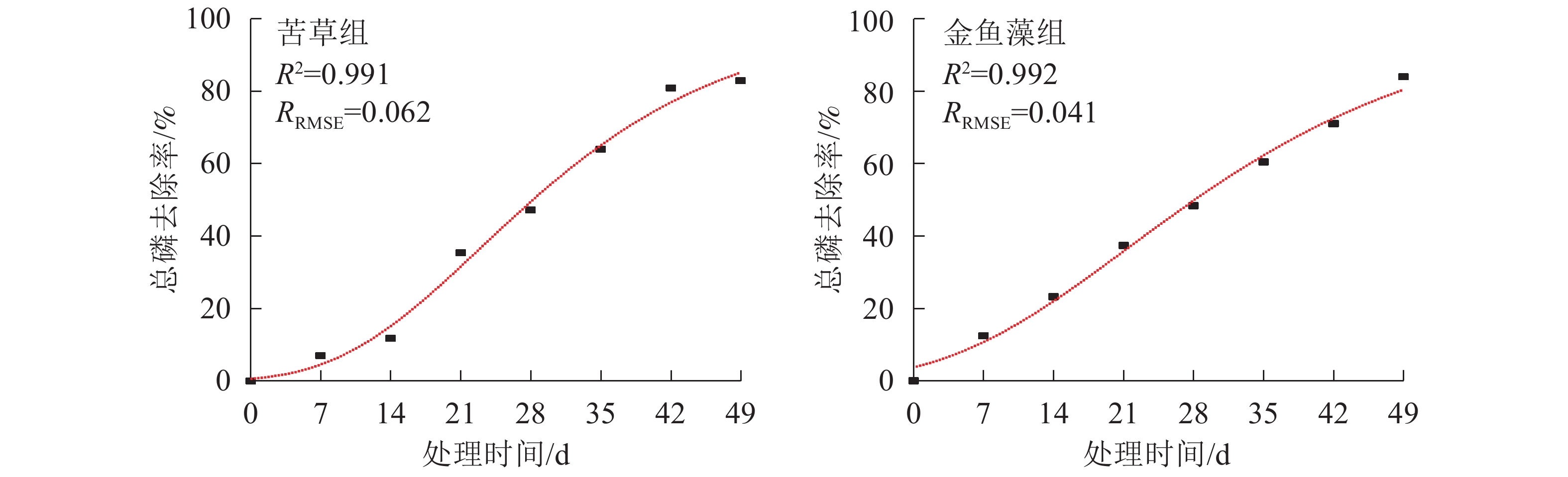
表 2 2种沉水植物净化系统总磷去除Monod动力学和修正的Gompertz模型拟合结果
Table 2. Fitting result of Monod kinetic and modified Gompertz model for total phosphorus removal in the submerged plant treatment systems
沉水植物型生态净化系统 Monod动力学模型 修正的Gompertz模型 Mmax/(mg·m−3·d−1) R2 RRMSE Gmax/(mg·m−3·d−1) ta/d R2 RRMSE 苦草组 87.14 a 0.974 0.108 107.96 a 8.70 a 0.991 0.062 金鱼藻组 78.15 b 0.999 0.020 84.82 b 3.37 b 0.992 0.041 说明:不同字母表示差异显著(P<0.05) Monod动力学模型表征了污染物去除速率与其质量浓度之间的关系由一级向零级反应过渡的状态,因而可以避免单纯使用一级动力学模型时可能造成的设施尺寸过大问题。作为一种在经典的米-门方程基础上建立起来的半经验模型,Monod动力学模型起初主要用于辅助活性污泥法、生物接触氧化法等污水生物处理工程设计。近年来Monod动力学模型越来越多地被应用于挺水植物型人工湿地系统有机物和氮素污染物去除能力的预测[19-22]。本研究证实:Monod动力学模型可用于沉水植物型净化系统批式运行模式下总磷去除过程的模拟及其去除效果的预测 (表2)。
-
自FAN等[28]于2004年首次对Gompertz模型进行修正,并将其用于描述好氧生化处理过程中邻苯二甲酸降解动力学特征以来,这种可以同时输出用于指示污染物去除效率和初始延迟时间的多参数模型越来越多地应用于厌氧反应器、好氧反应器等工艺系统污染物去除过程的模拟[23-25]。本研究采用修正的Gompertz模型[公式(7)]对2组沉水植物型净化系统总磷去除过程进行非线性拟合,结果见图5和表2。
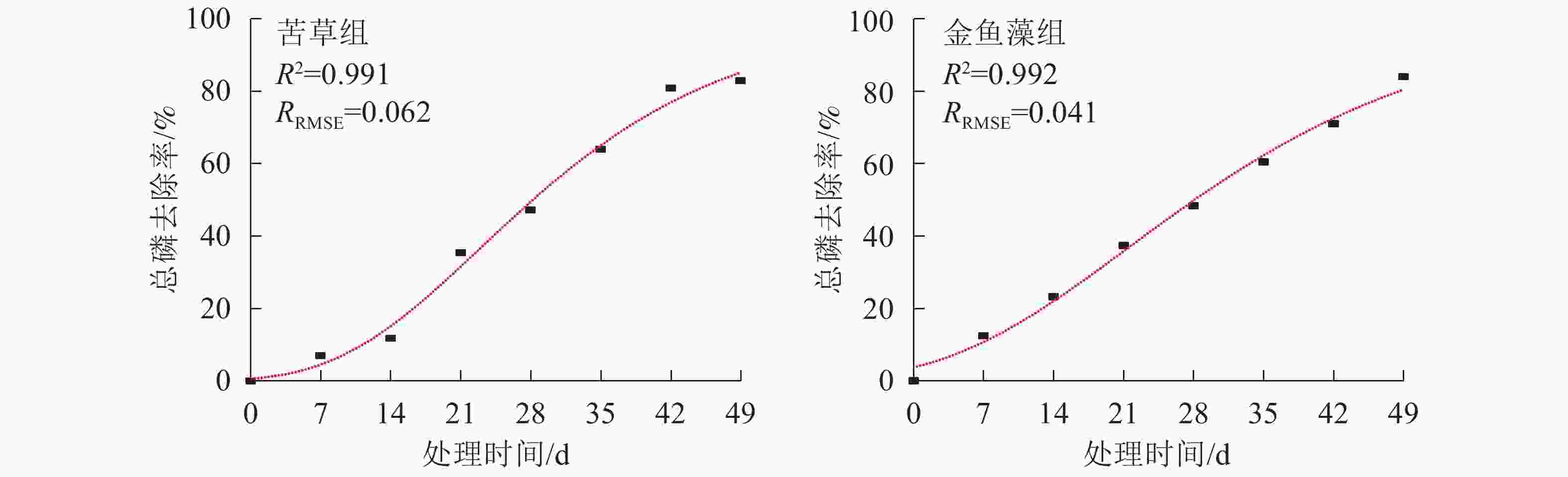
图 5 2种沉水植物净化系统总磷去除Gompertz模型动力学拟合曲线
Figure 5. Regression of modified Gompertz model for total phosphorus removal over time in the submerged plant treatment systems
结合图5和表2可以看出:修正的Gompertz模型可以很好地拟合本试验条件下沉水植物净化系统总磷去除效率与处理时间之间的关系(R2>0.990,RRMSE<0.110)。2组植物净化系统拟合获得的总磷最大体积去除速率和除磷延迟时间均有所不同(P<0.05),其中苦草组净化系统总磷最大体积去除速率为107.96 mg·m−3·d−1,显著高于金鱼藻组净化系统(84.82 mg·m−3·d−1) (P<0.05),表明苦草组净化系统总磷去除效率显著高于金鱼藻组净化系统。但苦草组净化系统拟合获得的除磷延迟时间(8.70 d)显著长于金鱼藻组净化系统(3.37 d),表明苦草型沉水植物净化系统相较于金鱼藻型需要更长的生境适应周期。
-
污染物去除动力学是指导工程设计的重要基础。许多动力学模型已用于模拟污水生物处理和人工湿地系统污染物的去除过程,其中一级动力学、Grau二级动力学和Monod 动力学模型简化了污染物去除过程中所涉及的生物、化学、物理及其相互影响等复杂过程,模型构式可线性化处理而使得参数求解十分简单,拟合求得的动力学参数又可以较好地代表处理系统污染物的去除能力。这些有利因素促使此类模型越来越多地应用到包括水平潜流[32]、垂直流[33]、潮汐流[34]、静止状态[35]及其组合工艺[19]等多种人工湿地系统中污染物的去除动力学特征的研究。但近年来研究发现:此类建立在众多假定条件基础上的表观动力学模型的适用性,不仅与湿地工艺类型(水平潜流和垂直流)及其水动力模式(推流或完全混合态)有关,还与所描述的污染物类型有关[19-20]。本研究结果表明:一级动力学和Monod 动力学模型均可以用来描述沉水植物型净化系统批式试验周期内的总磷去除过程,而Grau二级动力学却不适用;相较于一级动力学,Monod 动力学模型具有更高的拟合优度。
近年来,修正的Gompertz模型作为一种多参数非线性模型,在污染物去除动力学研究领域逐步被重视。众多研究已发现:这种非线性预测模型描述异养硝化菌、厌氧氨氧化等新型好氧或厌氧生物处理系统的脱氮除磷效果具有较好的拟合精确度[23-25]。但该模型能否用于模拟人工湿地、植物塘等生态处理系统中污染物的去除过程至今还未见报道。本研究发现:修正的Gompertz模型可以很好地模拟沉水植物净化系统总磷的去除动力学过程。相较于一级动力学和Monod动力学模型,修正的Gompertz模型的拟合除了可以输出代表着污染物去除能力的关键性能参数(如最大体积去除速率),还可以输出可用于指示前期阶段污染物去除的迟缓时间。这些参数的获取有助于对污染物去除机制的理解,也可用于协助工艺系统的优化设计和过程调控。
-
本研究发现:2组植物系统总磷去除效率的差异在时间尺度上存在明显的不确定性;这种不同种类植物净化效率的差异,在时间尺度上出现偏离甚至逆转的现象极有可能普遍存在。如李琳等[31]针对不同种类沉水植物氮磷净化效率的对比研究发现:当处理时间为7和21 d时金鱼藻组总磷净化效率显著好于苦草组(P<0.05),但持续处理至第35 天时,两者总磷的净化效率无显著差异。究其原因,一方面可能是由于不同种类的沉水植物对磷的吸收量存在时间变异,但更为重要的原因可能是不同植物所介导的间接增效作用也存在时间尺度上的差异。
众多研究已表明:在由沉水植物、基质(自然沉积物或人工填料)、微生物和水体四大基本要素构成的水生态系统或由其衍生而来的“水下植被”型生态拦截净化系统中,沉水植物、基质和微生物以及三者相互之间通过一系列物理(沉积、挥发等)、化学(基质吸附、沉淀等)和生物(动植物吸收、微生物降解转化等)作用,实现对污染物的有效去除,其中沉水植物的作用被证实是磷去除的主要机制[6, 30]。一直以来,植物的直接吸收作用被认为是“水下植被”净化系统除磷的主要途径,但近年来越来越多的证据显示:因沉水植物所介导的间接增效作用(如根际效应、茎叶生物膜效应等)对总磷去除的贡献强度远超过植物的直接吸收作用,并且该作用途径对总磷去除的贡献强度也与植物的种类密切相关[30-31]。这种尚未引起重视的现象给水生植物的筛选带来了不确定性,有必要借助诸如动力学模型分析等手段加以分析。本研究通过动力学分析发现:2组沉水植物系统总磷去除一级动力学速率常数Fa、Monod动力学速率常数Mmax以及修正的Gompertz模型拟合常数Gmax均存在显著差异,并且一致地表现为苦草组指标大于金鱼藻组指标。据此可以推断,苦草型净化系统对总磷的去除效率好于金鱼藻型。
-
苦草型和金鱼藻型2种植物净化系统总磷去除效率无显著差异,总磷去除率分别为82.8%和84.0%,但两者总磷去除效率在不同时间尺度上的差异存在不确定性。4种动力学模型模拟与验证结果表明:除Grau二级动力学外,一级动力学、Monod 动力学模型和修正的Gompertz模型均可以有效描述2种沉水植物型净化系统试验条件下总磷的去除过程,其中Monod动力学和Gompertz模型的模拟具有更高的拟合优度,模型预测值与实测值之间吻合程度更好。3种有效模型拟合获得的动力学常数在植物种类之间均存在显著差异(P<0.05),其中指示除磷效率的动力学速率常数一致表现为苦草组大于金鱼藻组,表明苦草型净化系统对总磷的去除效率好于金鱼藻型。但苦草组除磷延迟时间显著长于鱼藻组(P<0.05),表明其生境适应周期相对较长。
Kinetic modeling of total phosphorus removal from farmland drainage with submerged macrophyte-type ecological purification system
-
摘要:
目的 探讨以沉水植物为先锋物种的生态净化系统处理农田退水磷污染的动力学性能。 方法 通过批式试验研究苦草Vallisneria natans和金鱼藻Ceratophyllum demersum等2种沉水植物型净化系统对农田退水总磷的净化效率及其动力学特征。 结果 至试验结束时(第49天),2种植物净化系统总磷去除效率无显著差异(P>0.05),总磷去除率分别为82.8%(苦草型)和84.0%(金鱼藻型),但两者总磷去除效率的差异在时间尺度上存在不确定性;动力学模拟分析发现:除Grau二级动力学外,一级动力学、Monod 动力学和修正的Gompertz模型均可以描述试验条件下总磷的去除过程[判定系数(R2)>0.930,相对均方根误差(RRMSE)<0.200],其中Monod动力学和Gompertz模型具有更高的拟合度(R2>0.970),预测值与实验观测值之间吻合程度更好(RRMSE<0.110)。 结论 3种有效模型拟合获得的动力学常数在植物种类水平上均存在显著差异(P<0.05),其中指示总磷去除效率的动力学常数值一致表现为苦草型>金鱼藻型,表明苦草型净化系统除磷效率好于金鱼藻型。图5表2参35 Abstract:Objective The objective of this study is to investigate the dynamic performance of ecological purification system with submerged macrophytes as pioneer species for treatment of phosphorus pollution in farmland drainage. Method A batch-mode mesocosm experiment was performed to study total phosphorus (TP) purification efficiency and kinetic parameters of two submerged macrophyte systems: Vallisneria natans and Ceratophyllum demersum. Four kinetic models including first-order kinetic, Grau second-order kinetic, Monod kinetic and modified Gompertz model were used to evaluate TP removal efficiency. Result At the end of the experiment (Day 49), there was no significant difference in TP removal efficiency between the two mesocosm systems (P>0.05), and the TP removal rates were 82.8% for V. natans and 84.0% for C. demersum, but the difference in TP removal efficiency was uncertain in time scale. Kinetic simulation analysis showed that except Grau second-order kinetic, first-order kinetic, Monod kinetic and modified Gompertz model were proven capable of predicting TP removal process of the two submerged plant systems under experimental conditions, with R2 (coefficient of determination)>0.930 and RRMSE (relative root mean square error)<0.200. Among them, Monod kinetic and modified Gompertz model showed a higher fitting degree (R2>0.970), and a better agreement between the predicted value and the observed value (RRMSE<0.110). Conclusion The kinetic parameters obtained by the three effective model fitting show significant differences at the plant species level (P<0.05), and V. natans has higher kinetic parameter of TP removal efficiency compared with C. demersum (P<0.05), indicating that TP removal efficiency of V. natans is better than that of C. demersum. [Ch, 5 fig. 2 tab. 35 ref.] -
图 1 沉水植物净化系统总磷累计去除负荷随处理时间的变化
Figure 1 Total phosphorus removal loading over treatment time in the submerged plant treatment systems
图 2 沉水植物净化系统各周总磷去除负荷
Figure 2 Total phosphorus weekly removal loading for the submerged plant treatment systems
图 3 2种沉水植物净化系统总磷去除一级动力学模型拟合曲线
Figure 3 Regression of first-order kinetic model for TP removal over time in the submerged plant treatment systems
图 4 2种沉水植物净化系统总磷去除Monod动力学拟合曲线
Figure 4 Regression of Monod kinetic model for total phosphorus removal over time in the submerged plant treatment systems
图 5 2种沉水植物净化系统总磷去除Gompertz模型动力学拟合曲线
Figure 5 Regression of modified Gompertz model for total phosphorus removal over time in the submerged plant treatment systems
表 1 2种沉水植物净化系统总磷去除一级动力学和Grau二级动力学拟合结果
Table 1. Fitting result of First-order and Grau second-order kinetic models for total phosphorus removal in the submerged plant treatment systems
沉水植物型生态净化系统 一级动力学模型 Grau二级动力学模型 Fa/d−1 R2 RRMSE m/d n R2 RRMSE 苦草组 0.039 0.931 0.196 109.90 −1.35 0.588 0.324 金鱼藻组 0.035 0.937 0.169 57.25 0.03 0.081 0.127  下载: 导出CSV
下载: 导出CSV
表 2 2种沉水植物净化系统总磷去除Monod动力学和修正的Gompertz模型拟合结果
Table 2. Fitting result of Monod kinetic and modified Gompertz model for total phosphorus removal in the submerged plant treatment systems
沉水植物型生态净化系统 Monod动力学模型 修正的Gompertz模型 Mmax/(mg·m−3·d−1) R2 RRMSE Gmax/(mg·m−3·d−1) ta/d R2 RRMSE 苦草组 87.14 a 0.974 0.108 107.96 a 8.70 a 0.991 0.062 金鱼藻组 78.15 b 0.999 0.020 84.82 b 3.37 b 0.992 0.041 说明:不同字母表示差异显著(P<0.05)
下载: 导出CSV
-
[1] 杨林章, 施卫明, 薛利红, 等. 农村面源污染治理的“4R”理论与工程实践——总体思路与“4R”治理技术[J]. 农业环境科学学报, 2013, 32(1): 1 − 8. YANG Linzhang, SHI Weiming, XUE Lihong, et al. Reduce-retain-reuse-restore technology for the controlling the agricultural non-point source pollution in countryside in China: general countermeasures and technologies [J]. J Agro-Environ Sci, 2013, 32(1): 1 − 8. [2] 武淑霞, 刘宏斌, 刘申, 等. 农业面源污染现状及防控技术[J]. 中国工程科学, 2018, 20(5): 23 − 30. WU Shuxia, LIU Hongbin, LIU Shen, et al. Review of current situation of agricultural non-point source pollution and its preventionand control technologies [J]. Eng Sci, 2018, 20(5): 23 − 30. [3] 翟敏婷, 辛卓航, 韩建旭, 等. 河流水质模拟及污染源归因分析[J]. 中国环境科学, 2019, 39(8): 3457 − 3464. ZHAI Minting, XIN Zhuohang, HAN Jianxu, et al. Water quality simulation and multi-source attribution analysis [J]. China Environ Sci, 2019, 39(8): 3457 − 3464. [4] 韦晓雪, 李晓琳, 郑毅. 基于输出系数模型的1998—2016年洱海流域磷素时空变化特征分析[J]. 农业环境科学学报, 2020, 39(1): 171 − 181. WEI Xiaoxue, LI Xiaolin, ZHENG Yi. Analysis of temporal and spatial variation characteristics of phosphorus in Erhai Lake basin from 1998 to 2016 based on export coefficient model [J]. J Agro-Environ Sci, 2020, 39(1): 171 − 181. [5] 彭亚辉, 周科平, 蒋俊伟. 湘江流域长株潭段水污染负荷时空分布规律及成因[J]. 中国农业大学学报, 2018, 23(9): 108 − 116. PENG Yahui, ZHOU Keping, JIANG Junwei. The spatial-temporal distribution and causes of water pollution loads on Xiangjiang River Basin in Changzhutan [J]. J China Agric Univ, 2018, 23(9): 108 − 116. [6] 刘福兴, 王俊力, 付子轼. 不同规格生态沟渠对排水污染物处理能力的研究[J]. 土壤学报, 2019, 56(3): 561 − 570. LIU Fuxing, WANG Junli, FU Zishi. Comparative research on effects of ecological ditches different in specification treating pollutants in drainage [J]. Acta Pedol Sin, 2019, 56(3): 561 − 570. [7] LI Dan, ZHENG Binghui, CHU Zhaosheng, et al. Seasonal variations of performance and operation in field-scale storing multipond constructed wetlands for nonpoint source pollution mitigation in a plateau lake basin [J]. Bioresour Technol, 2019, 280: 295 − 302. [8] KUMWIMBA M N, MENG Fangang, ISEYEMI O I, et al. Removal of non-point source pollutants from domestic sewage and agricultural runoff by vegetated drainage ditches (VDDs): design, mechanism, management strategies, and future directions [J]. Sci Total Environ, 2018, 639: 742 − 759. [9] 浙江省农业农村厅. 浙江省农业绿色发展试点先行区3年行动计划(2018—2020)[EB/OL]. (2018-07-02)[2020-02-20]. http://www.zjagri.gov.cn/art/2018/7/4/art1589297_30567167.html. Department of Agriculture and Rural Affairs of Zhejiang Province. Zhejiang Province Agricultural Green Development Pilot Three-Year (2018−2020) Plan [EB/OL]. (2018-07-02)[2020-02-20]. http://www.zjagri.gov.cn/art/2018/7/4/art1589297_30567167.html. [10] 岑璐瑶, 陈滢, 张进, 等. 种植不同植物的人工湿地深度处理城镇污水处理厂尾水的中试研究[J]. 湖泊科学, 2019, 31(2): 365 − 374. CEN Luyao, CHEN Ying, ZHANG Jin, et al. Pilot-scale study on advanced treatment of tail water of urban sewage treatment plant by constructed wetlands with different plants [J]. J Lake Sci, 2019, 31(2): 365 − 374. [11] 李晓东, 孙铁珩, 李海波, 等. 人工湿地除磷研究进展[J]. 生态学报, 2007, 27(3): 1226 − 1232. LI Xiaodong, SUN Tieheng, LI Haibo, et al. Current researches and prospects of phosphorus removal in constructed wetland [J]. Acta Ecol Sin, 2007, 27(3): 1226 − 1232. [12] 王文林, 刘波, 韩睿明, 等. 沉水植物茎叶微界面及其对水体氮循环影响研究进展[J]. 生态学报, 2014, 34(22): 6409 − 6416. WANG Wenlin, LIU Bo, HAN Ruiming, et al. Research advancements and perspectives on leaf and stem micro-interfaces in submerged macrophytes and its effect on water nitrogen cycling [J]. Acta Ecol Sin, 2014, 34(22): 6409 − 6416. [13] LIN Qinshuo, GU Binhe, HONG Jianming. Tracking uptake of submerged macrophytes (Ceratophyllum demersum): derived nitrogen by cattail (Typha angustifolia) using nitrogen stable isotope enrichments [J]. Ecol Eng, 2017, 99: 114 − 118. [14] LI Qi, GU Peng, JI Xiyan, et al. Response of submerged macrophytes and periphyton biofilm to water flow in eutrophic environment: plant structural, physicochemical and microbial properties [J]. Ecotoxicol Environ Saf, 2020, 189: 1 − 8. [15] ZHANG Shunan, LIU Feng, HUANG Zhenrong, et al. Are vegetated drainage ditches effective for nitrogen removal under cold temperatures [J]. Bioresour Technol, 2020, 301: 1 − 6. [16] OLESEN A, JENSON S M, ALNOEE A B, et al. Nutrient kinetics in submerged plant beds: a mesocosm study simulating constructed drainage wetlands [J]. Ecol Eng, 2018, 122: 263 − 270. [17] 刘淼, 陈开宁. 植物配置与进水碳氮比对沉水植物塘水质净化效果的影响[J]. 环境科学, 2018, 39(6): 2706 − 2714. LIU Miao, CHEN Kaining. Purification effect of submerged macrophyte system with different plants combinations and C/N ratios [J]. Environ Sci, 2018, 39(6): 2706 − 2714. [18] SAMAL K, DASH R R, BHUNIA P. Design and development of a hybrid macrophyte assisted vermifilter for the treatment of dairy wastewater: a statistical and kinetic modelling approach [J]. Sci Total Environ, 2018, 645: 156 − 169. [19] NGUYEN X C, CHANG S W, NGUYEN T L, et al. A hybrid constructed wetland for organic-material and nutrient removal from sewage: process performance and multi-kinetic models [J]. J Environ Manage, 2018, 222: 378 − 384. [20] SAEED T, SUN Guangzhi. Kinetic modelling of nitrogen and organics removal in vertical and horizontal flow wetlands [J]. Water Res, 2011, 45(10): 3137 − 3152. [21] 殷志平, 吴义锋, 吕锡武. 基于一级动力学模型的水培蔬菜滤床氮磷去除模拟[J]. 东南大学学报(自然科学版), 2016, 46(4): 812 − 817. YIN Zhiping, WU Yifeng, LU Xiwu. Simulation of nitrogen and phosphorus removal in hydroponic vegetable filter bed based on first-order kinetics model [J]. J Southeast Univ Nat Sci Ed, 2016, 46(4): 812 − 817. [22] 殷志平, 吴义锋, 吕锡武. 景观型与蔬菜型水平潜流湿地除磷动力学模型[J]. 化工学报, 2016, 67(5): 2048 − 2055. YIN Zhiping, WU Yifeng, LÜ Xiwu. Kinetic modelling of total phosphorus removal in landscape type and vegetable type horizontal subsurface-flow constructed wetlands [J]. CIESC J, 2016, 67(5): 2048 − 2055. [23] 黄硕, 于德爽, 陈光辉, 等. 氧化石墨烯强化厌氧氨氧化菌的脱氮性能[J]. 中国环境科学, 2019, 39(5): 1945 − 1953. HUANG Shuo, YU Deshuang, CHEN Guanghui, et al. Improvement of the activity of anammox bacteria using graphene oxide [J]. China Environ Sci, 2019, 39(5): 1945 − 1953. [24] 杨垒, 陈宁, 任勇翔, 等. 异养硝化细菌Acinetobacter junii NP1的同步脱氮除磷特性及动力学分析[J]. 环境科学, 2019, 40(8): 3713 − 3721. YANG Lei, CHEN Ning, REN Yongxiang, et al. Simultaneous nitrogen and phosphorus removal and kinetics by the heterotrophic nitrifying bacterium Acinetobacter junii NP1 [J]. Environ Sci, 2019, 40(8): 3713 − 3721. [25] MU Yang, YU Hanqing, WANG Gang. A kinetic approach to anaerobic hydrogen-producing process [J]. Water Res, 2007, 41(5): 1152 − 1160. [26] 王涛, 张维理, 张怀志. 滇池流域人工模拟降雨条件下农田施用有机肥对磷素流失的影响[J]. 植物营养与肥料学报, 2008, 14(6): 1092 − 1097. WANG Tao, ZHANG Weili, ZHANG Huaizhi. Effects of swine manure application on P losses from different farmlands under simulated rainfall in Dianchi watershed of Yunnan Province [J]. Plant Nutr Fert Sci, 2008, 14(6): 1092 − 1097. [27] 申东, 唐家良, 章熙峰, 等. 紫色土丘陵区农业小流域暴雨事件磷素多尺度流失特征[J]. 水土保持学报, 2017, 31(5): 56 − 63. SHEN Dong, TANG Jialiang, ZHANG Xifeng, et al. Characteristics of phosphorus loss of small agricultural watershed during rainstorm events in hilly area of purple soil [J]. J Soil Water Conserv, 2017, 31(5): 56 − 63. [28] FAN Yanzhen, WANG Yingying, QIAN Peiyuan, et al. Optimization of phthalic acid batch biodegradation and the use of modified Richards model for modelling degradation [J]. Int Biodeterior Biodegradation, 2004, 53(1): 57 − 63. [29] 吴旻, 赵群芬. 3种沉水植物在不同污染水体中的生长及其对水质的影响[J]. 生物学杂志, 2015, 32(4): 43 − 47. WU Min, ZHAO Qunfen. The growth of three submerged plants in different polluted water and its impact on water quality [J]. J Biol, 2015, 32(4): 43 − 47. [30] 金树权, 周金波, 包薇红, 等. 5种沉水植物的氮、磷吸收和水质净化能力比较[J]. 环境科学, 2017, 38(1): 156 − 161. JIN Shuquan, ZHOU Jinbo, BAO Weihong, et al. Comparison of nitrogen and phosphorus uptake and water purification ability of five submerged macrophytes [J]. Environ Sci, 2017, 38(1): 156 − 161. [31] 李琳, 岳春雷, 张华, 等. 不同沉水植物净水能力与植株体细菌群落组成相关性[J]. 环境科学, 2019, 40(11): 4962 − 4970. LI Lin, YUE Chunlei, ZHANG Hua, et al. Correlation between water purification capacity and bacterial community composition of different submerged macrophytes [J]. Environ Sci, 2019, 40(11): 4962 − 4970. [32] SUN Guangzhi, SAEED T. Kinetic modelling of organic matter removal in 80 horizontal flow reed beds for domestic sewage treatment [J]. Process Biochem, 2009, 44(7): 717 − 722. [33] SAEED T, SUN Guangzhi. The removal of nitrogen and organics in vertical flow wetland reactors: predictive models [J]. Bioresour Technol, 2011, 102(2): 1205 − 1213. [34] 叶捷, 彭剑峰, 高红杰, 等. 潮汐流人工湿地低温下NH4 +-N去除模型的比较和优化[J]. 环境科学学报, 2011, 31(7): 1456 − 1463. YE Jie, PENG Jianfeng, GAO Hongjie, et al. Comparison and optimization of a NH4 +-N removal model of a tidal-flow constructed wetland in low temperature [J]. Acta Sci Circumstantiae, 2011, 31(7): 1456 − 1463. [35] 臧维玲, 刘永士, 戴习林, 等. 低频率运转下人工湿地对养虾水的去氮作用及其动力学[J]. 农业工程学报, 2013, 29(18): 210 − 217. ZANG Weiling, LIU Yongshi, DAI Xilin, et al. Performance and dynamics of nitrogen removal in constructed wetlands at low frequency for shrimp culture [J]. Trans Chin Soc Agric Eng, 2013, 29(18): 210 − 217. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210260
 点击查看大图
点击查看大图

计量
- 文章访问数: 2416
- HTML全文浏览量: 386
- PDF下载量: 54
- 被引次数: 0












