




-
结球甘蓝Brassica oleracea var. capitata是十字花科Brassicaceae芸苔属Brassica的1年生或2年生草本植物,为甘蓝B. oleracea的变种,又称卷心菜、洋白菜、圆白菜、疙瘩白、包菜、包心菜等。结球甘蓝具有耐寒、抗病,适应性强、易储耐运、产量高、品质好等特点,因此在中国各地普遍栽培,已成为中国的主要蔬菜品种之一,播种面积和产量在所有蔬菜中位居第3位,在周年供应中均占有重要地位[1]。现代营养学研究表明:甘蓝富含多种生物活性成分,尤以异硫氰酸盐[2]、叶酸[3]、酚酸[4]为突出。因此,甘蓝在人们饮食健康中占据着重要的地位[5]。
因为甘蓝是忌连作作物,所以下一茬必须种植非十字花科作物。然而,在山西旱地即使后茬种植不同种类的非十字花科作物,其生长也均会受到一定的抑制。以玉米Zea mays为例,在播种45 d后测定株高及叶片数,株高仅为其他茬口玉米的60%,且展开叶约少2片,茎基部最初展开叶发紫,严重影响玉米的前期生长,可造成减产[6]。这种植物间的相生相克被称为化感作用,该作用是活体植物通过根系分泌,地上部的挥发与雨雾淋溶或植物的残体分解等途径,向环境中释放次生代谢产物,从而影响邻近植物生长的化学生态现象[7-8],对栽培的前后茬作物生长和产量等方面的影响较大[9]。许多药用植物如丹参Salvia miltiorrhiza、三七Panax notoginseng、当归Angelica sinensis等均已证实存在明显的化感作用[10-11],多种蔬菜的化感作用也得到了系统评价[12-16],但对于结球甘蓝作为供体植物对下茬作物产生的化感作用研究鲜见报道。为此,本研究参照化感作用的研究方法,模拟雨雾淋溶与浸溶途径,研究结球甘蓝叶片营养生长期内对当地常见作物的种间化感作用,为建立合理的结球甘蓝轮作制度提供理论依据。
-
供体材料为结球甘蓝‘铁将军’B. oleracea var. capitata‘Tiejiangjun’叶片,2019年取样于山西农业大学山西有机旱作农业研究院河村示范试验基地。该地区年平均降水量为450 mm,集中于6—9月,水热同期。受体材料为糯玉米‘万糯2000’Zea mays ‘Wannuo 2000’和西葫芦‘绿霸先锋’Cucurbita pepo‘Lübaxianfeng’,种子均购于市场。
-
将结球甘蓝结球部位的叶片清洗后风干,经粉碎后分别称取30、40、50 g (预实验结果),分别置于洁净的锥形瓶中,加入500 mL蒸馏水后放入恒温摇床中浸泡振荡24 h (温度25 ℃,转速150 r·min−1),经双层滤纸过滤制得质量浓度为0.06、0.08、0.10 kg·L−1的水提液。
-
挑选颗粒饱满、质量接近的糯玉米和西葫芦种子播种在高8 cm, 上边长10 cm,下边长 7 cm的小方盒中,进行盆栽培养。每盒播种2粒,出苗后及时间苗,以保证每盆中仅留1株幼苗。装盆土壤均为菜豆茬口,种下土壤质量为250 g,种上土壤质量为200 g。添加的结球甘蓝叶片水提液质量浓度为0.10 (处理1)、0.08 (处理2)、0.06 kg·L−1(处理3),每盆均总计加入100 mL的水提液,分种上与种下2个部分等量加入,以蒸馏水为对照,置于恒温培养箱(温度为25 ℃,湿度为50%,12 h 4 000 lx光照;温度为20 ℃,湿度为60%,12 h黑暗)。培养过程中处理组与对照组分别定时补充适量水提液与蒸馏水,保持土壤湿润。每个处理设置4个重复,20 d后测定苗高、根长、地上部干质量与根干质量。
-
采用化感作用效应指数(IR,简称化感指数)度量化感作用的类型和强度:
$$ I_{\mathrm{R}}=\left\{\begin{array}{cc}1-C / T, & T {\text{≥}} C \\ T/C -1, & T<C\end{array}\right.。 $$ 其中:C为对照值,T为处理值。IR>0为促进作用,IR<0为抑制作用,绝对值的大小表示作用强度。处理组与对照组的差异显著性检验采用SPSS 18.0软件进行Duncan新复极差法分析。
采用综合效应指数对不同受体的化感效应进行综合评价。综合效应指数即为同一处理下受体作物的苗高、根长、地上部干质量与根系干质量的化感指数(IR)的算术平均值[17]。
-
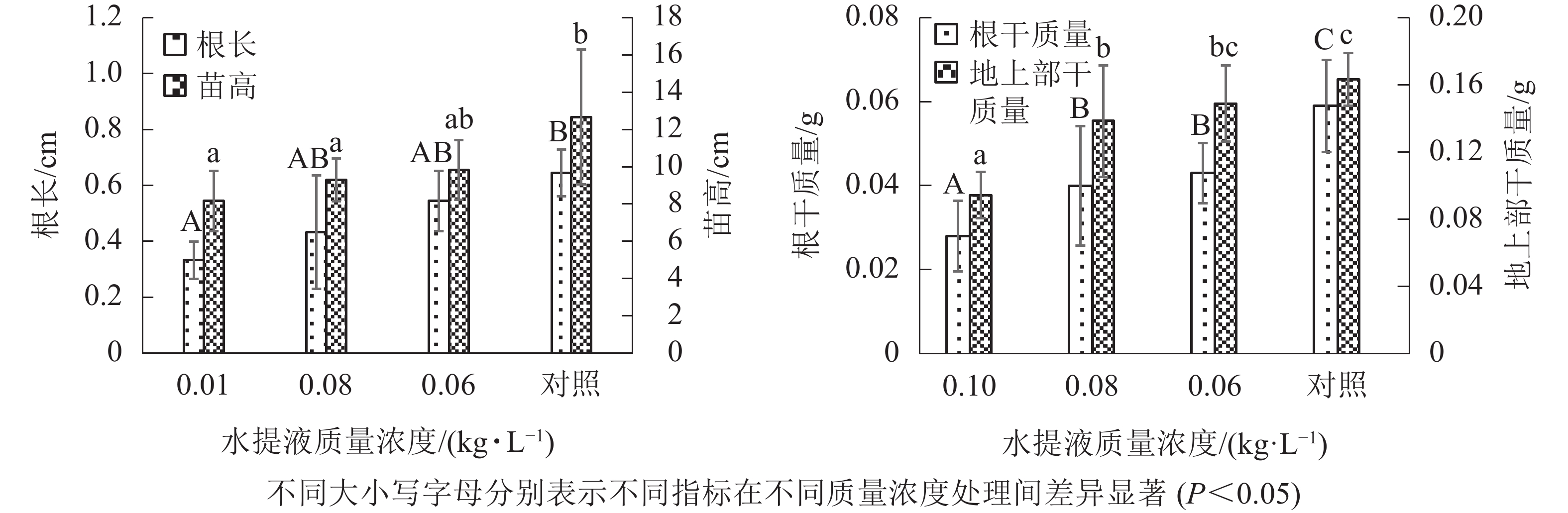
从图1可知:不同质量浓度结球甘蓝叶水提液均可显著(P<0.05)抑制盆栽糯玉米幼苗根系与地上部的伸长,且对两者的抑制程度随结球甘蓝叶水提液质量浓度的增大而增强,当水提液质量浓度增大至0.10 kg·L−1时,对苗高与根长的抑制效应最为明显,抑制率分别达到42.3%和57.8%,显著大于其他处理(P<0.05)。0.08与0.06 kg·L−1这2个质量浓度之间的差异未达到显著水平。

图 1 不同质量浓度结球甘蓝叶水提液对盆栽糯玉米苗高、根长、根干质量与地上部干质量的影响
Figure 1. Effects of different concentrations of cabbage-leaf water extracts on the seedling height, root length, root and shoot dry weight of waxy corn in pot
从图1可知:与对照相比,添加不同质量浓度的结球甘蓝叶水提液均可显著(P<0.05)降低盆栽20 d的糯玉米幼苗根系与地上部的生物量。当添加质量浓度最高(0.10 kg·L−1)时,对根系与地上部干质量均表现出最强的抑制效果,抑制率分别高达43.5%与55.2%;质量浓度降低至0.08 kg·L−1时,对地上部干质量的抑制作用明显减弱,而对根干质量的抑制效应仍未减轻;当质量浓度降至0.06 kg·L−1时,对地上部干质量的抑制程度反而又显著(P<0.05)增强,但对根系干质量的抑制作用明显减弱。综上可知,结球甘蓝叶水提液对糯玉米幼苗根干质量的影响表现出了“剂量效应”,即水提液质量浓度增大,抑制效果增强,抑制率为17.2%~43.5%。相比之下,结球甘蓝叶水提液对盆栽糯玉米的根系生长的抑制效果要强于地上部,可能与根系直接接触化感抑制物有关。
由表1可以看出:各质量浓度水提液处理对糯玉米苗高、根长等生物学检测指标的化感效应指数均为负值,即结球甘蓝叶水提液对盆栽糯玉米苗期生长的化感效应均表现为抑制作用。随着结球甘蓝叶水提液质量浓度的增加,对糯玉米苗高、根长与根干质量的抑制效应强度也随之增强。其中糯玉米苗期根长对最大处理质量浓度0.10 kg·L−1的敏感性响应最强(IR=−0.573 6),且显著(P<0.05)强于其他2个处理;根干质量对最小处理质量浓度0.06 kg· L−1的敏感性响应最弱(IR =−0.171 8)。
表 1 不同质量浓度结球甘蓝叶水提液对盆栽糯玉米幼苗生长的化感指数
Table 1. Allelopathic index of different concentrations of cabbage-leaf water extracts on the seedling growth of waxy corn in pot
水提液质量浓
度/(kg·L−1)化感指数 苗高 根长 地上部干质量 根干质量 0.10 −0.422 7 a −0.576 3 a −0.552 4 a −0.435 1 a 0.08 −0.323 0 b −0.487 8 b −0.347 9 b −0.272 9 ab 0.06 −0.308 5 b −0.472 7 b −0.470 0 a −0.171 8 b 说明:同列不同小写字母表示差异显著(P<0.05) -
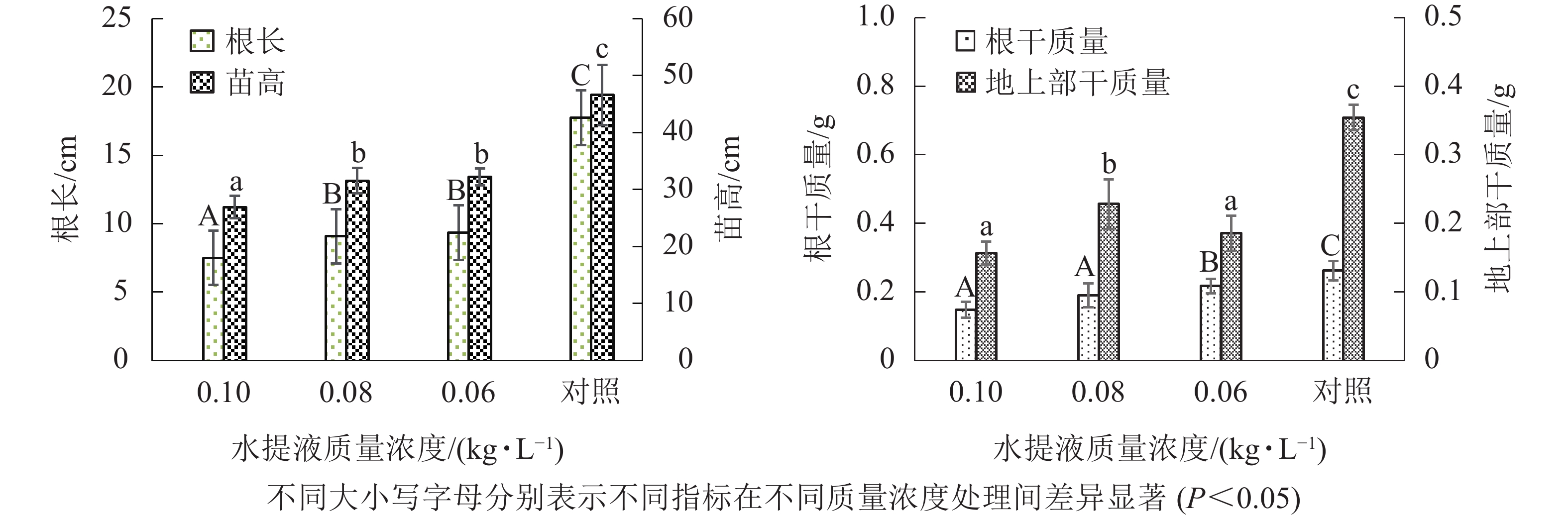
从图2可知:结球甘蓝叶水提液可以抑制盆栽西葫芦地上与地下部的伸长,其抑制程度随水提液质量浓度的降低而逐渐减弱,仅当添加最高质量浓度(0.10 kg·L−1)的结球甘蓝叶水提液显著(P<0.05)降低西葫芦的苗高与根长,降幅分别为40.0%与50.0%。与对盆栽玉米生长的影响类似,结球甘蓝叶高质量浓度水提液对西葫芦根系伸长的抑制程度大于地上部。但0.08与0.06 kg·L−1结球甘蓝叶水提液均未显著抑制盆栽西葫芦的苗高与根长,表明西葫芦对结球甘蓝叶水提液有更强的耐受力。

图 2 不同质量浓度结球甘蓝叶水提液对盆栽西葫芦幼苗高、根长、根干质量和地上部干质量的影响
Figure 2. Effects of different concentrations of cabbage-leaf water extracts on the seedling height, root length, root and shoot dry weight of cocozelle in pot
图2结果显示:结球甘蓝叶水提液均能显著(P<0.05)抑制西葫芦幼苗根干质量的增加。不同质量浓度水提液的抑制作用由强到弱依次为0.10、0.08、0.06 kg·L−1。较对照而言,3个处理可使西葫芦幼苗根干质量降低27.1%~52.5%。与对根干质量的影响类似,各处理亦可抑制地上部干质量,但仅0.10与0.08 kg·L−1能显著(P<0.05)降低西葫芦苗期的地上部干质量,降幅分别为42.2%与15.0%。最低质量浓度处理(0.06 kg·L−1)较对照仅下降了8.6%。
由表2可以得出:西葫芦苗期根干质量对结球甘蓝叶最大水提液质量浓度(0.10 kg·L−1)的处理敏感性响应最强,其化感指数为−0.5254,且显著(P<0.05)强于其他2个质量浓度处理;地上部干质量对最小处理质量浓度(0.06 kg·L−1)的敏感性响应最弱,其化感指数仅为−0.0859。检测的各项生物学指标所对应的化感指数均为负值,表明供试质量浓度的结球甘蓝叶水提液均可抑制西葫芦苗期根系与地上部的生长。随着处理质量浓度的降低,各项指标的化感抑制负效应逐渐减弱,其中对根长的抑制效应呈显著(P<0.05)减弱的趋势。
表 2 不同质量浓度结球甘蓝叶水提液对盆栽西葫芦幼苗生长的化感指数
Table 2. Allelopathic index of different concentrations of cabbage-leaf water extracts on cocozelle seedling growth in pot
水提液质量浓
度/(kg·L−1)化感指数 苗高 根长 地上部干质量 根干质量 0.10 −0.357 0 a −0.484 5 a −0.420 8 a −0.525 4 a 0.08 −0.267 7 b −0.329 9 b −0.149 9 b −0.322 0 b 0.06 −0.225 7 b −0.158 1 c −0.085 9 b −0.271 2 b 说明:同列不同小写字母表示差异显著(P<0.05) -
如表3所示:各供试质量浓度的结球甘蓝叶水提液对糯玉米与西葫芦苗期生长均表现化感抑制作用,其中最高质量浓度的结球甘蓝叶水提液对2种受体作物的化感综合效应均高于低质量浓度,分别为−0.496 6与−0.446 9。随着水提液质量浓度的降低,对2种作物的综合化感抑制效应均逐渐减弱,其中西葫芦的减幅要大于糯玉米。结球甘蓝叶水提液对糯玉米的综合化感抑制作用要强于对西葫芦。
表 3 不同质量浓度结球甘蓝叶水提液对2种作物幼苗生长的化感综合效应
Table 3. Allelopathic comprehensive effects of different concentrations of cabbage-leaf water extract on the seedling growth of two crops
受体作物 不同质量浓度水提液的化感综合效应 0.10 0.08 0.06 kg·L−1 糯玉米 −0.496 6 −0.357 9 −0.355 7 西葫芦 −0.446 9 −0.267 4 −0.185 2 -
在蔬菜栽培中化感作用是常见的现象,是作物与土壤相互作用的结果。在山西旱地,6—8月雨水较丰富,结球甘蓝营养生长旺盛,体内代谢速度快,次生产物丰富,通过淋溶和浸溶的途径能使次生代谢物进入土壤环境并积累。当自然界中的化感物质在土壤中累积到一定程度后,就会影响植物种子萌发与幼苗生长,进而影响植物对地上和地下资源的竞争[18]。本研究所设置的结球甘蓝叶水提液质量浓度是在预试验的基础上选取的相对较高的质量浓度水平,故对糯玉米与西葫芦均表现出了较为明显的化感效应。其中0.06 kg·L−1处理与田间结球甘蓝茬对西葫芦的影响程度较为接近,与对照(菜豆茬)相比并无直观差异。需要补充说明的是,预试验的结果表明:当添加结球甘蓝叶水提液的质量浓度≤0.03 kg·L−1时,对盆栽玉米苗期生长几乎没有抑制作用;当质量浓度降至0.01 kg·L−1时,还能促进盆栽西葫芦苗期根长,苗高及地上部干质量等指标显著增加。同一化感物质在不同质量浓度时会对同一植物出现抑制或促进2种截然不同的结果[19],因此建议在采收结球甘蓝的过程中,应尽量将地上部完全去除,避免其累积残留于土壤中影响其他作物生长。
有研究表明:当归水浸提物对受体作物油菜Brassica campestris、蚕豆Vicia faba和小麦Triticum aestivum种子萌发,胚芽和胚根生长均具有较强的化感作用,且多表现为抑制作用,其中尤以油菜最为敏感[20]。与该结果类似,本研究发现:糯玉米和西葫芦对结球甘蓝叶水提液的敏感性也不同。尽管对糯玉米和西葫芦苗期生长均表现出了抑制效应,但其中糯玉米的敏感性要强于西葫芦。与文献[20]研究结果有所不同的是,结球甘蓝叶水提液需要在较高质量浓度时才能表现出抑制作用,而当归水提液在相对较低的质量浓度时即可产生抑制作用。还有研究证明:药用植物如三七、人参Panax ginseng、地黄Rehmannia glutinosa、黄芪Astragalus membranaceus、苍耳Xanthium sibiricum、西洋参Panax quinquefolius等都具有显著的化感效应[21-26]。推测药材中的有效成分是作用很强的化感物质,而蔬菜中的化感物质活性与含量均不及药用植物。
也有相当多的研究结果表明:许多蔬菜如番茄Lycopersicon esculentum、大蒜Allium sativum、大葱Allium fistulosum、黄瓜Cucumis sativus及苋菜Amaranthus tricolor等对其他蔬菜具有显著的抑制作用,且不同作物对化感物质的反应有明显差别[12-16, 27-28]。因此,在农业生产中无论是作物的单植,还是间作、轮作、覆盖、翻埋、重茬种植等,都需要考虑化感作用的影响[28]。本研究结果发现:结球甘蓝叶水提液对西葫芦的综合化感抑制作用弱于糯玉米,这与观察到田间种植情况相吻合,建立西葫芦与甘蓝的轮作制度可能是解决结球甘蓝栽培中化感问题的对策之一。有关结球甘蓝根系及其分泌物中的化感物质的作用尚待进一步研究。
Allelopathic effect of water extract of Brassica oleracea var. capitata leaves on seedling growth of Zea mays and Cucurbita pepo
-
摘要:
目的 探讨结球甘蓝Brassica oleracea var. apitata化感作用与茬口障碍的关系,为建立合理轮作制度提供理论依据。 方法 以结球甘蓝风干叶片为供试材料,以适宜山西寒旱区栽培且经济收益相对较高的2种作物:糯玉米Zea mays和西葫芦Cucurbita pepo作为受体植物,采用室内生物测定法,比较了不同质量浓度结球甘蓝叶水提液对上述2种受体作物幼苗营养生长的影响,并计算相应的化感效应指数和综合效应指数。 结果 当结球甘蓝叶水提液质量浓度≥0.06 kg·L−1时均显著(P<0.05)抑制盆栽糯玉米苗期根系与地上部的生长。仅当添加的结球甘蓝叶水提液质量浓度为0.10 kg·L−1时,显著降低(P<0.05)西葫芦的苗高与根长。在水提液同一质量浓度下,对糯玉米根长的化感抑制作用始终大于苗高。对西葫芦则表现为当结球甘蓝叶水提液质量浓度≥0.08 kg·L−1时,对根长的化感抑制作用大于苗高;当水提液质量浓度降至0.06 kg·L−1时,对根长的化感抑制作用小于苗高。由综合效应指数可知,对西葫芦的化感综合抑制作用要小于糯玉米。 结论 西葫芦可用于结球甘蓝轮作体系,以减轻化感作用而引起的障碍。图2表3参28 Abstract:Objective The objective is to probe the relationship between toxicity of Brassica oleracea var. capitata and stubble obstacles, so as to provide a theoretical basis for establishing a reasonable crop rotation system. Method Taking the air-dried leaves of B. oleracea var. capitata as test materials, two crops species (Zea mays and Cucurbita pepo) with relatively high economic benefits and suitable for cultivation in cold and arid area of Shanxi were used as receptor. The effects of water extract from B. oleracea var. capitata leaves with four mass concentrations 0, 0.06, 0.08 and 0.10 kg·L−1 on the nutritional growth of the above two receptor crops were compared by indoor bioassay method, and the corresponding allelopathic effect index and comprehensive effect index were calculated. Result When the water extract concentration of B. oleracea var. capitata leaves was ≥0.06 kg·L−1, the growth of roots and shoots of potted Z. mays seedling could be significantly inhibited (P<0.05). The seedling height and root length of C. pepo were significantly reduced (P<0.05) only when the mass concentration of water extract of B. oleracea var. capitata leaves was 0.10 kg·L−1. At the same concentration, the allelopathic inhibition on root length of Z. mays was always greater than that on seedling height. For C. pepo, the allelopathic inhibition on root length was greater than that on seedling height when the water extract concentration of B. oleracea var. capitata leaves was ≥0.08 kg·L−1. When the mass concentration decreased to 0.06 kg·L−1, the allelopathic inhibition on root length was less than that on seedling height. According to the results of allelopathic comprehensive effect index, the allelopathic comprehensive inhibition on C. pepo was less than that on Z. mays. Conclusion C. pepo can be used in the rotation system of B. oleracea var. capitata to alleviate the obstacles caused by other toxicity. [Ch, 2 fig. 3 tab. 28 ref.] -
Key words:
- Brassica oleracea var. apitata /
- water extract /
- allelopathic effect /
- Zea mays /
- Cucurbita pepo
-
图 1 不同质量浓度结球甘蓝叶水提液对盆栽糯玉米苗高、根长、根干质量与地上部干质量的影响
Figure 1 Effects of different concentrations of cabbage-leaf water extracts on the seedling height, root length, root and shoot dry weight of waxy corn in pot
图 2 不同质量浓度结球甘蓝叶水提液对盆栽西葫芦幼苗高、根长、根干质量和地上部干质量的影响
Figure 2 Effects of different concentrations of cabbage-leaf water extracts on the seedling height, root length, root and shoot dry weight of cocozelle in pot
表 1 不同质量浓度结球甘蓝叶水提液对盆栽糯玉米幼苗生长的化感指数
Table 1. Allelopathic index of different concentrations of cabbage-leaf water extracts on the seedling growth of waxy corn in pot
水提液质量浓
度/(kg·L−1)化感指数 苗高 根长 地上部干质量 根干质量 0.10 −0.422 7 a −0.576 3 a −0.552 4 a −0.435 1 a 0.08 −0.323 0 b −0.487 8 b −0.347 9 b −0.272 9 ab 0.06 −0.308 5 b −0.472 7 b −0.470 0 a −0.171 8 b 说明:同列不同小写字母表示差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同质量浓度结球甘蓝叶水提液对盆栽西葫芦幼苗生长的化感指数
Table 2. Allelopathic index of different concentrations of cabbage-leaf water extracts on cocozelle seedling growth in pot
水提液质量浓
度/(kg·L−1)化感指数 苗高 根长 地上部干质量 根干质量 0.10 −0.357 0 a −0.484 5 a −0.420 8 a −0.525 4 a 0.08 −0.267 7 b −0.329 9 b −0.149 9 b −0.322 0 b 0.06 −0.225 7 b −0.158 1 c −0.085 9 b −0.271 2 b 说明:同列不同小写字母表示差异显著(P<0.05)
下载: 导出CSV
表 3 不同质量浓度结球甘蓝叶水提液对2种作物幼苗生长的化感综合效应
Table 3. Allelopathic comprehensive effects of different concentrations of cabbage-leaf water extract on the seedling growth of two crops
受体作物 不同质量浓度水提液的化感综合效应 0.10 0.08 0.06 kg·L−1 糯玉米 −0.496 6 −0.357 9 −0.355 7 西葫芦 −0.446 9 −0.267 4 −0.185 2
下载: 导出CSV
-
[1] 马爱民, 王峰, 潘国云, 等. 结球甘蓝主要加工利用途径分析[J]. 中国果菜, 2018, 38(5): 5 − 8. MA Aimin, WANG Feng, PAN Guoyun, et al. Analysis on the main processing and utilization ways of cabbage [J]. China Fruit Veg, 2018, 38(5): 5 − 8. [2] TANG Li, PAONESSA J D, ZHANG Yuesheng, et al. Total isothiocyanate yield from raw cruciferous vegetables commonly consumed in the United States [J]. J Funct Foods, 2013, 5(4): 1996 − 2001. [3] DELCHIER N, HERBIG A, RYCHLIK M, et al. Folates in fruits and vegetables: contents, processingand stability [J]. Compr Rev Food Sci Food Saf, 2016, 15(3): 506 − 528. [4] CARTEA M E, FRANCISCO M, SOENGAS P, et al. Phenolic compounds in Brassica vegetables [J]. Molecules, 2010, 16(1): 251 − 280. [5] NOOYENS A C J, BUENO-de-MESQUITA H B, van BOXTEL M P J, et al. Fruit and vegetable intake and cognitive decline in middle-aged men and women: the doetinchem cohort study [J]. Br J Nutr, 2011, 106(5): 752 − 761. [6] 张伟, 张冬梅, 韩彦龙, 等. 不同前茬下旱地玉米的水分动态及产量效应[J]. 山西农业科学, 2017, 45(5): 749 − 752, 781. ZHANG Wei, ZHANG Dongmei, HAN Yanlong, et al. Effects of different previous crops on soil moisture and crop yield of maize in dryland [J]. J Shanxi Agric Sci, 2017, 45(5): 749 − 752, 781. [7] 张重义, 林文雅. 药用植物的化感自毒作用与连作障碍[J]. 中国生态农业学报, 2009, 17(1): 189 − 196. ZHANG Zhongyi, LIN Wenya. Continuous cropping obstacle and allelopathic autotoxicity of medicinal plants [J]. Chin J Eco-Agric, 2009, 17(1): 189 − 196. [8] 曹璞, 沈益新. 狗牙根对5种禾本科杂草化感作用的研究[J]. 草地学报, 2010, 18(3): 452 − 455. CAO Pu, SHEN Yixin. Study on allelopathic effects of bermudagrass on five gramineous weeds [J]. Acta Agrestia Sin, 2010, 18(3): 452 − 455. [9] 孙敏, 姚海燕, 任旭琴, 等. 凹土对甘蓝的大蒜鳞茎浸提液化感胁迫的缓解效应[J]. 北方园艺, 2016(24): 31 − 34. SUN Min, YAO Haiyan, REN Xuqin, et al. Relieved allelopathy effect of attapulgite for garlic bulb aqueous extracts on cabbage [J]. Northern Hortic, 2016(24): 31 − 34. [10] 张亚琴, 陈雨, 雷飞益, 等. 药用植物化感自毒作用研究进展[J]. 中草药, 2018, 49(8): 1964 − 1953. ZHANG Yaqin, CHEN Yu, LEI Feiyi, et al. Advances in research on allelopathic autotoxicity effects of medicinal plants [J]. Chin Tradit Herbal Drugs, 2018, 49(8): 1964 − 1953. [11] 马瑞君, 惠继瑞, 朱慧, 等. 当归营养期的化感作用[J]. 中国生态农业学报, 2008, 16(6): 1483 − 1488. MA Ruijun, HUI Jirui, ZHU Hui, et al. Allelopathy of Angelica sinensis at vegetative stage [J]. Chin J Eco-Agric, 2008, 16(6): 1483 − 1488. [12] 徐勇峰, 黄斌, 朱陈名, 等. 堆制番茄秸秆浸提液对黄瓜和大白菜的化感作用[J]. 浙江农林大学学报, 2017, 34(2): 276 − 282. XU Yongfeng, HUANG Bin, ZHU Chenming, et al. Allelopathic effects of anaqueous extract from composted tomato residues on the growth of cucumber and Chinese cabbage [J]. J Zhejiang A&F Univ, 2017, 34(2): 276 − 282. [13] 蒋红云, 张燕宁, 冯平章, 等. 石蒜对萝卜、黄瓜番茄和油菜幼苗的化感效应[J]. 应用生态学报, 2006, 17(9): 1655 − 1659. JIANG Hongyun, ZHANG Yanning, FENG Pingzhang, et al. Allelopathic effects of Lycoris radiate on radish, cucumber, tomato and rape seedlings [J]. Chin J Appl Ecol, 2006, 17(9): 1655 − 1659. [14] 赵红, 王婷, 余李. 大蒜鳞茎浸提液对大豆种子萌发和幼苗生长的化感效应[J]. 大豆科学, 2019, 38(4): 548 − 553. ZHAO Hong, WANG Ting, YU Li. Allelopathic effects of garlic bulb aqueous extracts on seed germination and seedling growth of soybean [J]. Soybean Sci, 2019, 38(4): 548 − 553. [15] 姚岭柏, 韩海霞, 李倩雯, 等. 大葱水浸液对萝卜的化感效应及其生理机制研究[J]. 河南农业科学, 2017, 46(8): 115 − 120. YAO Lingbai, HAN Haixia, LI Qianwen, et al. Research on allelopathy of Allium fistulosum L. aqueous extracts and its physiological mechanism on Raphanus sativus L. [J]. J Henan Agric Sci, 2017, 46(8): 115 − 120. [16] 由海霞. 设施黄瓜不同种植模式的环境效应及其化感作用研究[D]. 杨凌: 西北农林科技大学, 2007. YOU Haixia. Study on the Environmental Effects of Different Planting Patterns and the Allelopathy Mechanism of Cucumber in Facility Condition[D]. Yangling: Northwest A&F Univercity, 2007. [17] 刘学东, 陈林, 杨新国, 等. 中间锦鸡儿花水浸提液对3种作物种子萌发及幼苗生长的化感作用[J]. 北方园艺, 2016(12): 65 − 70. LIU Xuedong, CHEN Lin, YANG Xinguo, et al. Allelopathic effects of aqueous extracts from Caragana intermadia’s flower on seed germination and seeding growth of three kinds of crop [J]. Northern Hortic, 2016(12): 65 − 70. [18] 王玉芝, 张汝民, 高岩. 冷蒿浸提液对几种饲用植物的化感作用[J]. 中国草地学报, 2008, 30(2): 47 − 53. WANG Yuzhi, ZHANG Rumin, GAO Yan. Allelopathy effect of extracts from Artemisia frigida Willd. on some feeding plants [J]. Chin J Grassland, 2008, 30(2): 47 − 53. [19] 和丽忠, 陈锦玉, 董宝生, 等. 国内植物化感作用研究概况[J]. 云南农业科技, 2001(1): 37 − 41. HE Lizhong, CHEN Jinyu, DONG Baosheng, et al. Survey of allelopathy research on plant in China [J]. J Yunnan Agric Sci Technol, 2001(1): 37 − 41. [20] 王田涛, 王琦, 王惠珍, 等. 当归自毒作用和其对不同作物的化感效应[J]. 草地学报, 2012, 20(6): 1132 − 1138. WANG Tiantao, WANG Qi, WANG Huizhen, et al. Autotoxicity of Angelica sinens and allelopathy on tested plants [J]. Acta Agrectia Sin, 2012, 20(6): 1132 − 1138. [21] 陈长宝. 人参化感作用及其忌连作机制研究[D]. 长春: 吉林农业大学, 2006. CHEN Changbao. Study on Ginseng Allelopathy and Elements of Unable Successive Cultivation[D]. Changchun: Jilin Agricultural University, 2006. [22] 张子龙, 王文全, 杨建忠, 等. 三七连作土壤对其种子萌发及幼苗生长的影响[J]. 土壤, 2010, 42(6): 1009 − 1014. ZHANG Zilong, WANG Wenquan, YANG Jianzhong, et al. Effects of continuous Panax notoginseng cropping soil on P. notoginseng seed germination and seedling growth [J]. Soils, 2010, 42(6): 1009 − 1014. [23] 朱广军. 地黄连作障碍中化感物质的鉴定[D]. 郑州: 河南农业大学, 2007. ZHU Guangjun. Identify the Phytotoxic Substances of Continuously Cropping Rehmannia glutinosa Libosch Obstacle[D]. Zhengzhou: Henan Agricultural University, 2007. [24] 赵晓萌. 连作对西洋参生长发育的影响及根际化感物质的研究[D]. 北京: 北京协和医学院, 2009. ZHAO Xiaomeng. The Effect of Continous Cropping on the Growth of American Ginseng and Allelochemicals[D]. Beijing: Peking Union Medical College, 2009. [25] 高兴祥, 李美, 高宗军, 等. 苍耳对不同植物幼苗的化感作用研究[J]. 草业学报, 2009, 18(2): 95 − 101. GAO Xingxiang, LI Mei, GAO Zongjun, et al. Allelopathic potential of Xanthium sibiricum on seeds germination and seedling growth of different plants [J]. Acta Prataculturae Sin, 2009, 18(2): 95 − 101. [26] 赵培强. 黄芪(Astragalus membranaceus)连作障碍的研究[D]. 兰州: 西北师范大学, 2009. ZHAO Peiqiang. Study on Continuous Cropping Obstacle of Astragalus membranaceus[D]. Lanzhou: Northwest Normal University, 2009. [27] 管安琴. 苋菜化感作用的初步研究[D]. 南京: 南京农业大学, 2015. GUAN Anqin. Preliminary Study on Allelopathy of Amaranth (Amaranthus Tricolor L. )[D]. Nanjing: Nanjing Agricultural University, 2015. [28] 佟飞, 程智慧, 金瑞, 等. 大蒜植株水浸液醇溶成分的化感作用[J]. 西北农林科技大学学报(自然科学版), 2007, 35(6): 119 − 124. TONG Fei, CHENG Zhihui, JIN Rui, et al. Allelopathy of methanol dissolved ingredient from garlic plant aqueous extracts [J]. J Northwest A&F Univ Nat Sci Ed, 2007, 35(6): 119 − 124. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210683
 点击查看大图
点击查看大图
计量
- 文章访问数: 1545
- HTML全文浏览量: 320
- PDF下载量: 29
- 被引次数: 0

