




 下载:
下载:
-
森林生态系统是陆地生态系统的主体,在减缓全球变暖方面具有不可替代的作用[1]。净生态系统生产力(net ecosystem productivity, NEP)作为净初级生产力和异养呼吸的差值,是表征生态系统碳收支的重要指标,被广泛应用于定量描述森林生态系统碳源/汇的能力。然而,大气温室气体浓度的增加使大气持水能力不断增强,显著增加了暴雨发生的频率和强度[2−3],已成为影响森林NEP最大的因子之一[4]。中国位于北半球中高纬度地区,跨越不同的气候带,不同地区气候条件、植被生长异质[5],使得中国极端降水及森林碳汇具有明显的区域特征,并且NEP对极端降水的响应也不同。因此,开展区域碳源/汇分布及其对极端降水响应研究,对气候灾害预防和生态保护具有重要意义。
准确估算极端降水时空格局是研究森林NEP响应极端降水的前提。相对阈值法是评判极端降水的常用方法,充分考虑了降水的地区间差异和季节因素[6],具有极端性弱、噪声低、显著性强等优点,可以更客观地分析极端降水的气候特征和变化趋势[7]。遥感观测协同生态过程模型不仅能估算森林NEP时空动态,更能帮助分析NEP对气候的响应机制,在区域NEP时空动态研究中具有显著优势[8]。InTEC模型是CHEN等[9]开发的融入森林年龄及干扰影响的生态系统模型,能更准确地模拟长期森林碳循环动态变化,在省域[10]和全国[11]尺度均得到了广泛应用。结构方程模型综合了方差、回归、路径和因子等统计分析方法,能模拟多因子的内在逻辑关系,综合评估因果联系和潜变量关系,已被广泛应用于生态学领域[12]。本研究以浙江省丽水市为研究区域,通过百分位阈值法分析1979—2079年极端降水时空演变特征,采用InTEC模型模拟不同情景下丽水市森林NEP时空格局,并构建结构方程定量分析丽水市森林NEP对极端降水的响应。研究结果将为丽水森林生态系统响应与适应全球气候变化提供理论支持,为维持和提升其植被碳汇功能提供依据。
-
丽水市位于浙江省西南部(27°25′~28°57′N,118°41′~120°26′E),森林面积为142.14 万hm2,森林覆盖率达82.27%,蓄积量为0.96 亿m 3,森林类型主要为亚热带针叶林、阔叶林和竹林等。林地的净初级生产力年总量与浙江省平均相比,所占比例较高[13]。丽水市地势西南高东北低,西南部以中山为主,东部以低山为主,中间有中山以及河谷盆地。全市主要气候类型为亚热带湿润季风气候,光照充足、四季分明、雨量丰沛。年均降水量为1 598.9 mm,多集中在3—9月,6月最多,是降水较为丰富的地区,也是极端气候事件多发的地区[14]。
-
本研究利用百分位法描述极端降水时空特征,将月尺度的降水量从小到大排序,取其第95百分位值作为极端降水量阈值(mm·月−1)[15]。如果某月降水量超过这一阈值,则称该月发生了极端降水事件,累积该月降水量即为年极端降水量(mm·a−1),并定义该年发生极端降水月数为极端降水频度(次·a−1)。最后,将极端降水量除以极端降水频度为极端降水强度(I)。
-
本研究使用InTEC模型模拟丽水市1979—2079年森林生态系统NEP,主要包括气象、土壤、森林年龄、参考年净初级生产力、氮沉降、二氧化碳(CO2)浓度及森林分布信息等时空分布数据。
-
1979—2015年丽水市气象数据来源于中国气象局国家气象中心丽水市地面日值气象数据,包括最高气温、最低气温、降水量、太阳辐射及相对湿度。通过反距离加权插值法获得空间分辨率为1 km的日尺度栅格气象数据,然后通过累计获得月尺度气象数据。2016—2079年气象数据则采用第5次国际耦合比较计划(Coupled Model Intercomparison Project Phase 5, CMIP5)的3种气候情景RCP 2.6、RCP 4.5和RCP 8.5获得,并通过最近邻法重采样到1 km分辨率。
-
利用最大似然法从Landsat5 TM数据中提取2004年丽水市针叶林、阔叶林、竹林等30 m分辨率森林分布信息。为了保持数据空间分辨率的一致性,采用局部平均法获取1 km森林丰度信息。具体步骤见文献[16]。
-
土壤数据包括土壤粉粒百分比、砂粒百分比、黏粒百分比、土壤深度、土壤有效持水力、土壤容重以及土壤萎蔫点。其中前3个数据来源于联合国粮食和农业组织及国际应用分析学会共同开发的全球土壤数据库(HWSD 1.2,
)。土壤容重使用Saxton改进的Brooks-Corey模型,结合土壤粉粒、砂粒、黏粒百分比来计算,土壤萎蔫点也根据粉粒、砂粒、黏粒百分比计算所得[17]。 -
根据浙江省森林资源清查小班数据提供的森林年龄信息,在对2004年浙江省森林资源样地清查年龄信息统计的基础上,利用克里金插值法得到2004年浙江省丽水市1 km分辨率的森林年龄空间分布数据。
-
1979—1999年的CO2浓度数据来源于全球监测地球系统研究实验室(
https://www.esrl.noaa.gov/gmd/ccgg/trends );2000—2079年RCP 2.6、RCP 4.5和RCP 8.5等3种情景下模拟的CO2浓度数据来自典型浓度路径(representative concentration pathways,RCPs)数据库(https://tntcat.iiasa.ac.at/RcpDb ),使用线性插值将10 a尺度的CO2浓度数据插值成年尺度。 -
氮沉降数据来源于全球格点大气总估计的无机氮、氮氢化合物和氮氧化物沉降量。1860、1993和2050年的氮沉降数据利用全球三维化传输模型(TM 3)模拟得到。本研究采用线性插值得到丽水市1979—2079年1 km空间分辨率的氮沉降数据。
-
本研究将2004年作为参考年校正初始的净初级生力值,使用BEPS模型模拟丽水市1 km分辨率净初级生力。具体模拟过程参考文献[18]。
-
本研究利用最小二乘法计算线性回归模型的斜率来分析变量的时空演变特征(θ)。计算公式为:
$$ \theta = \frac{{n\displaystyle \sum\limits_{i = 1}^n {{x_i}{y_i} - \displaystyle \sum\limits_{i = 1}^n {{x_i}\displaystyle \sum\limits_{i = 1}^n {{y_i}} } } }}{{n\displaystyle \sum\limits_{i = 1}^n {{x_i}^2 - {{\left(\displaystyle \sum\limits_{i = 1}^n {{x_i}} \right)}^2}} }} 。 $$ (1) 式(1)中:n为年的总数;xi为年($ i $=1,2,…,50);yi为第i年的变量值。θ>0表示上升趋势,<0表示下降趋势。
-
偏最小二乘(partial least square, PLS)通径分析是结构方程模型估计方法,可用以分析多变量之间的线性统计关系,可以有效解决多变量复共线问题,适用于变量多重相关或样本容量较小的情况,能克服变量数据分布复杂和变量之间相关的问题[12]。本研究采用该方法定量分析气候因子对NEP影响的相对重要程度及NEP驱动因子的直接作用和间接作用,在SmartPLS 3.3.9软件中进行。
-
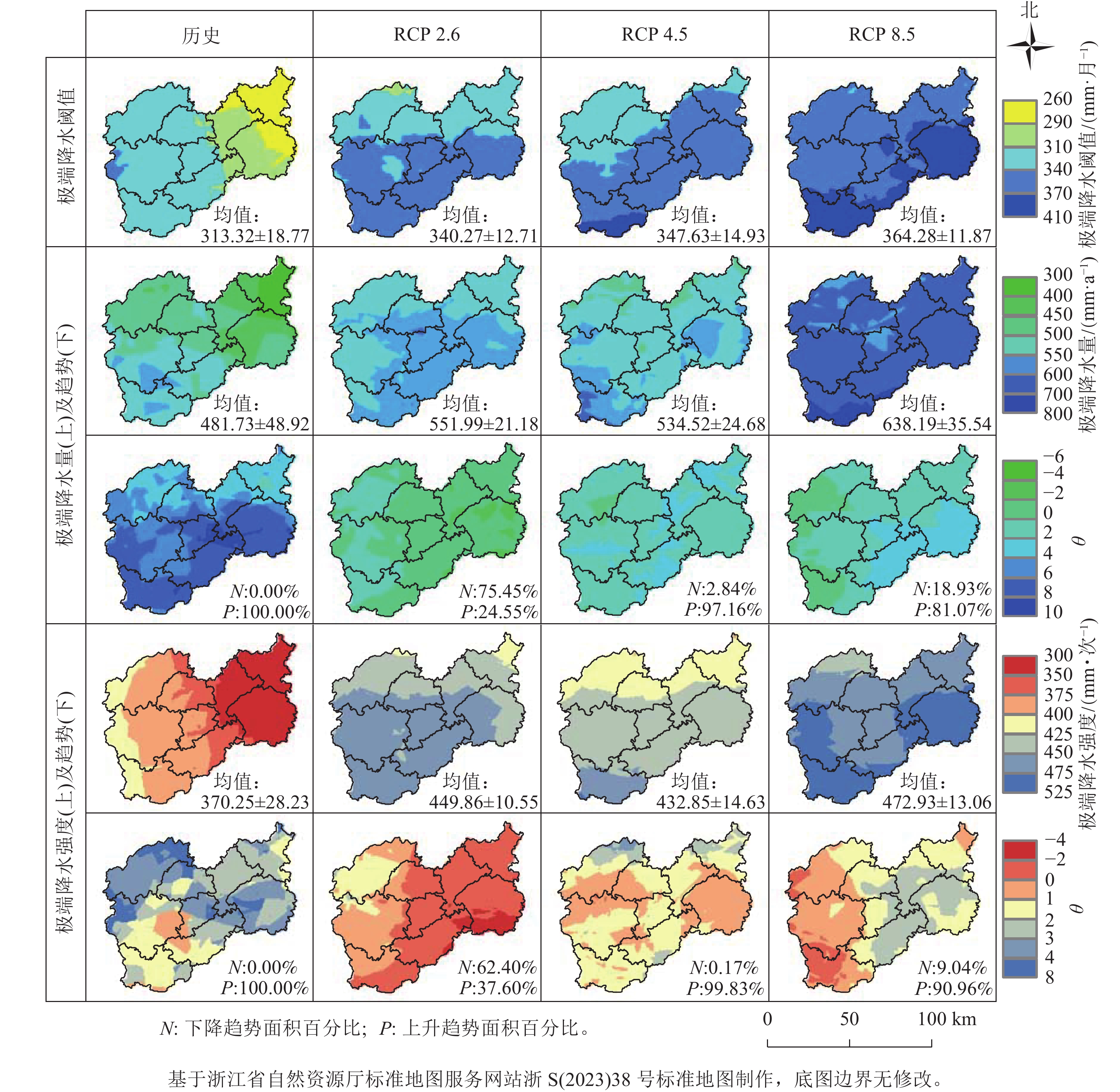
通过百分位阈值法计算月尺度的极端降水和阈值的空间分布结果,以及极端降水强度的空间分布结果如图1所示。总体上,未来丽水市极端降水量及强度除RCP 2.6情景外,其他2种情景下均呈上升趋势,并且RCP 4.5情景下2个指标上升趋势面积达90%以上,但上升趋势不显著。丽水市历史极端降水阈值为(313.32±18.77) mm·月−1,随着辐射胁迫增加极端降水阈值逐步增加,最大达(364.28±11.87) mm·月−1,在空间上呈现由西南向东北递减的趋势。2016年之前丽水市所有地区极端降水量及强度均呈上升趋势,极端降水量平均值为481.73 mm·a−1。RCP 8.5情景下未来变化最明显,极端降水量均值达638.19 mm·a−1,比历史的年均极端降水量增加了156.46 mm·a−1,强度是历史的1.28倍。
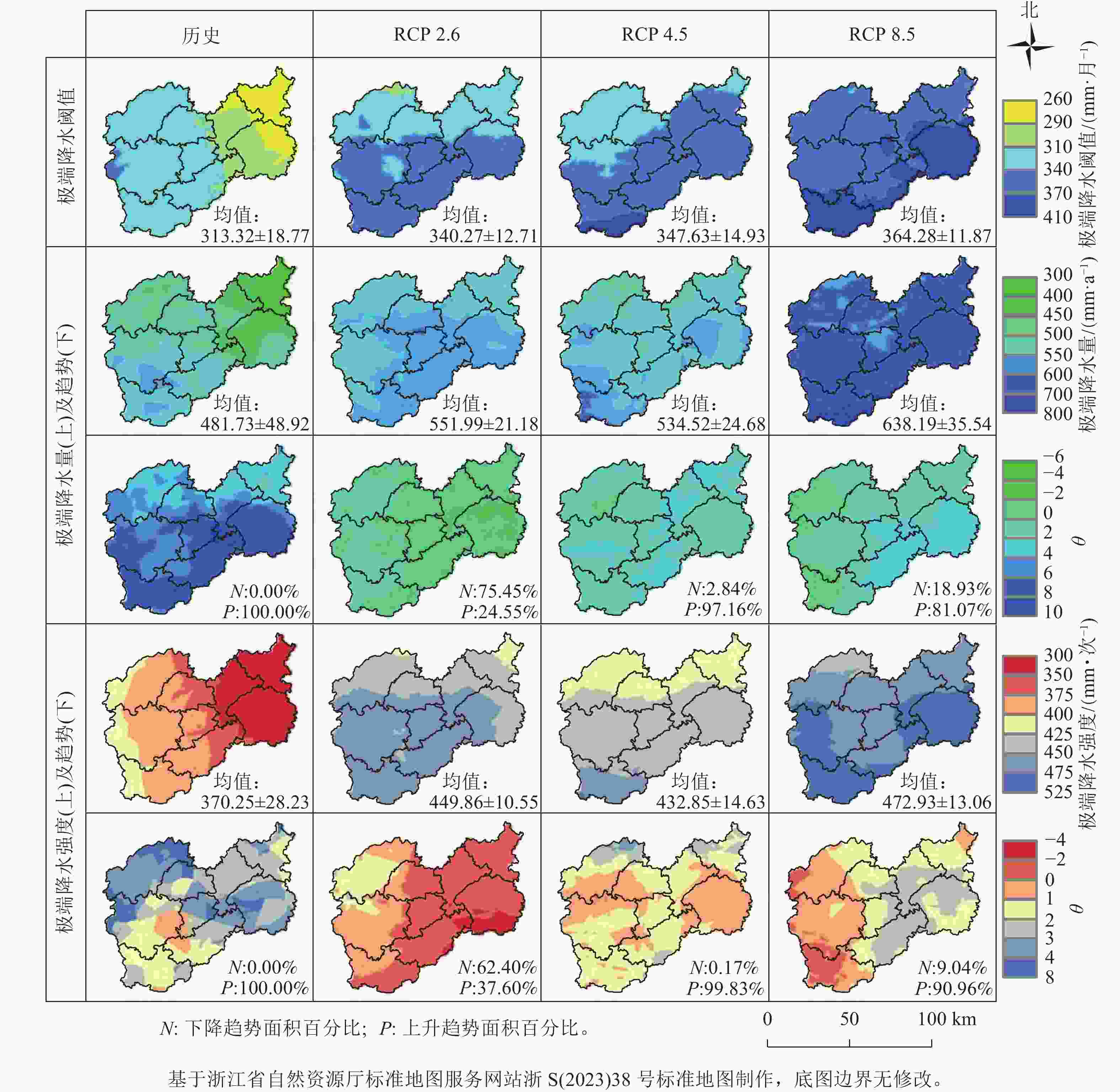
图 1 历史(1979—2015年)及不同气候情景下未来(2016—2079年)丽水市极端降水的空间分布
Figure 1. Spatial distribution of extreme precipitation in Lishui City under historical (1979-2015) and future (2016-2079) climate scenarios
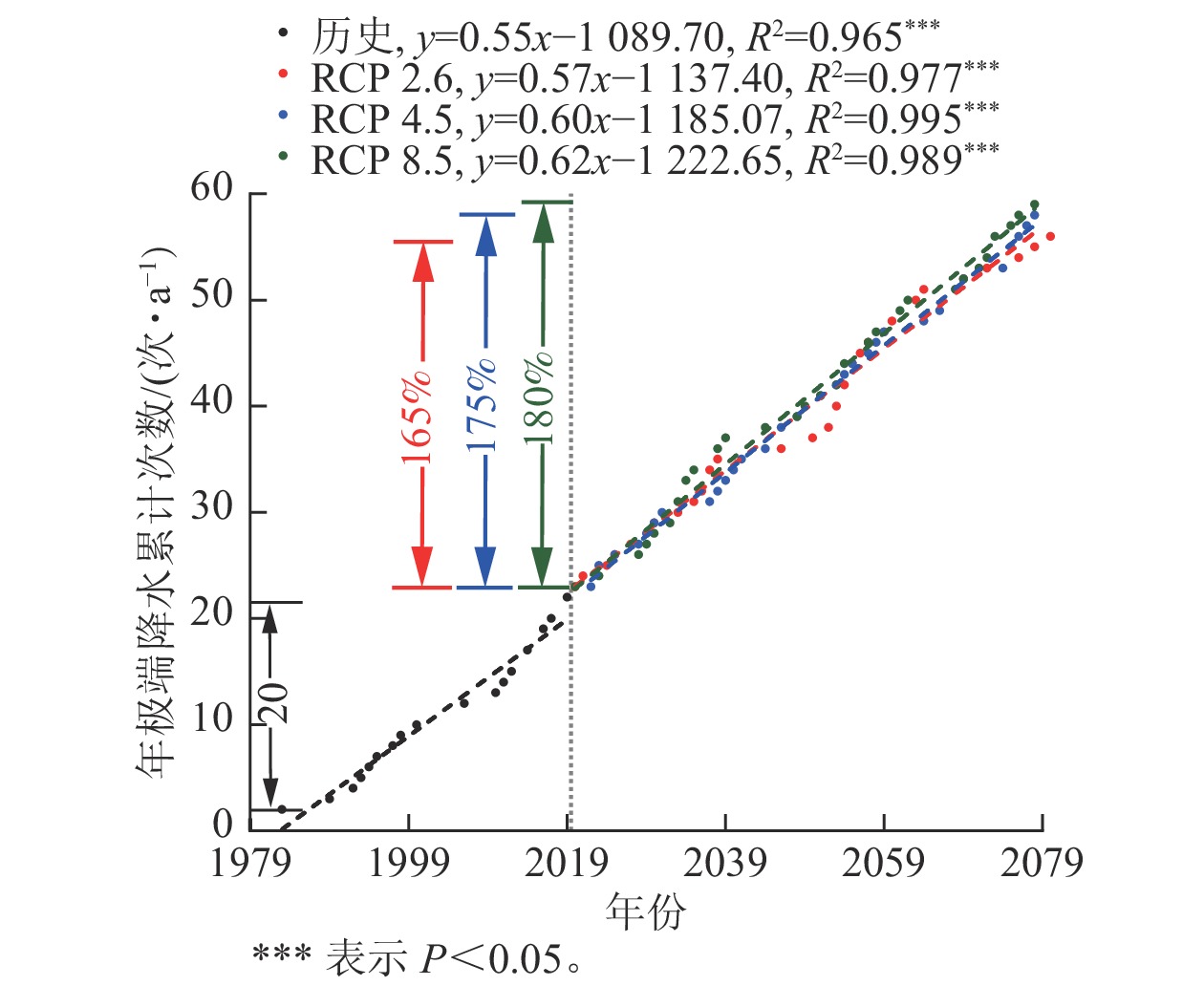
1979—2079年丽水市年极端降水累计次数的年际变化如图2所示。总体上,历史和未来极端降水增加趋势相近,分别为0.55与0.60,历史年极端降水次数增加量为20次·a−1,未来气候情景下年极端降水次数随辐射胁迫的增加而增加,至2079年累计发生次数分别较2016年增加33、35和36次·a−1,分别比历史次数增加了165%、175%、180%。

图 2 历史(1979—2019年)及不同气候情景下未来(2019—2079年)丽水市年极端降水累计次数的年际变化
Figure 2. Interannual changes of the cumulative number of extreme precipitation in Lishui City under historical (1979-2019) and different climate scenarios (2019-2079)
-
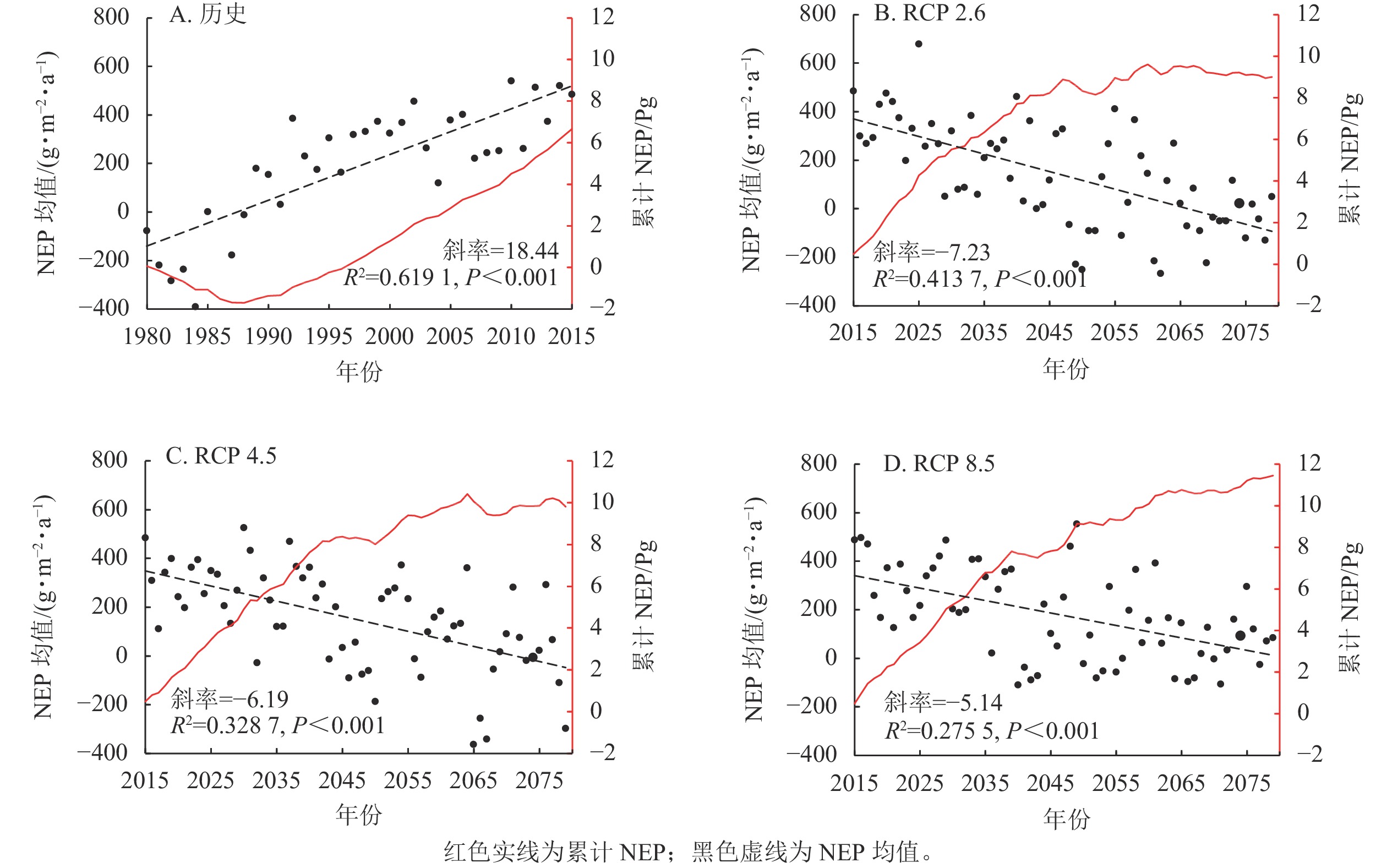
1979—2079年丽水市森林NEP的年际变化如图3所示。由图3A可见:1979—2015年丽水市森林NEP整体呈显著增长趋势。1979—1988年表现为碳源,1986年NEP值历史最低,为−439.24 g·m−2·a−1,1989年开始从碳源转为碳汇,累计NEP增至2015年的6.8 Pg。由图3B~C可见:3种情景下2016—2079年丽水市森林NEP碳汇量呈下降趋势,RCP 8.5气候情景对比最不明显,RCP 2.6和RCP 4.5情景下森林碳汇从2064年开始已经表现为碳源。碳汇累积量在2064年之前一直处于增加状态,之后除了RCP 8.5情景继续增加外,其他2种情景已经停止增加甚至有下降的现象。3种情景下的NEP累积量相对于2015年分别增加了8.51、9.32、10.97 Pg。总体来说,1979—2079年间丽水市森林NEP呈先增加再减少的趋势,并且在RCP 8.5情景下减少最慢。

图 3 历史(1979—2015年)及不同气候情景下未来(2016—2079年)丽水市森林NEP的年际变化
Figure 3. Historical (1979-2015) and future (2016-2079) interannual changes of forest NEP in Lishui City under different climate scenarios
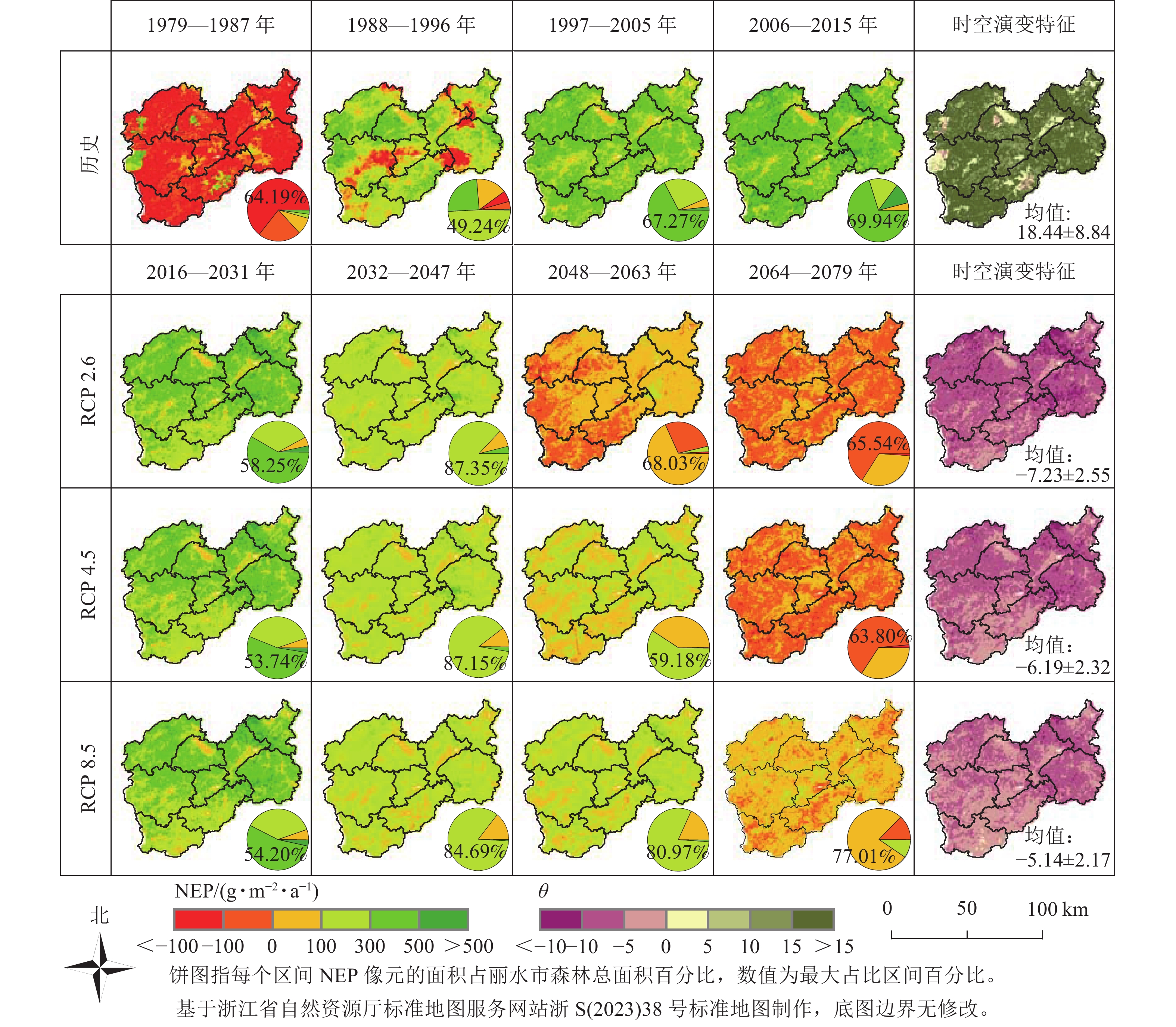
图4为1979—2079年丽水市NEP以及时空演变特征的空间分布结果。从历史来看,1988—1996年碳汇能力增强,NEP<0的区域减少到9.74%,2015年时NEP>0的区域已达100%,丽水所有区域在历史上森林碳汇上升趋势都比较高。3种气候情景下,2016—2031年丽水市整体表现为碳汇,超过50%的区域为300<NEP<500 g·m−2·a−1,主要分布在北部区域;2032—2047年森林全市碳汇能力下降,超过80%的区域为100<NEP<300 g·m−2·a−1;2048—2063年森林碳汇能力持续下降,RCP 2.6下降最明显,主要在西部区域;2064—2079年,依旧持续下降。RCP 8.5气候情景77.01%的区域集中在0<NEP<100 g·m−2·a−1,RCP 2.6和RCP 4.5情景下NEP<0的区域增加到66.49%和65.46%,积累量有下降的现象,且下降趋势高值区主要分布在东部和北部。

图 4 1979—2015基准年以及不同气候情景下2016—2079年丽水市NEP的空间分布和时空演变特征
Figure 4. Spatial distribution of NEP and linear trend in Lishui City during 1979-2015 base year and 2016-
2079 under different climate scenarios -
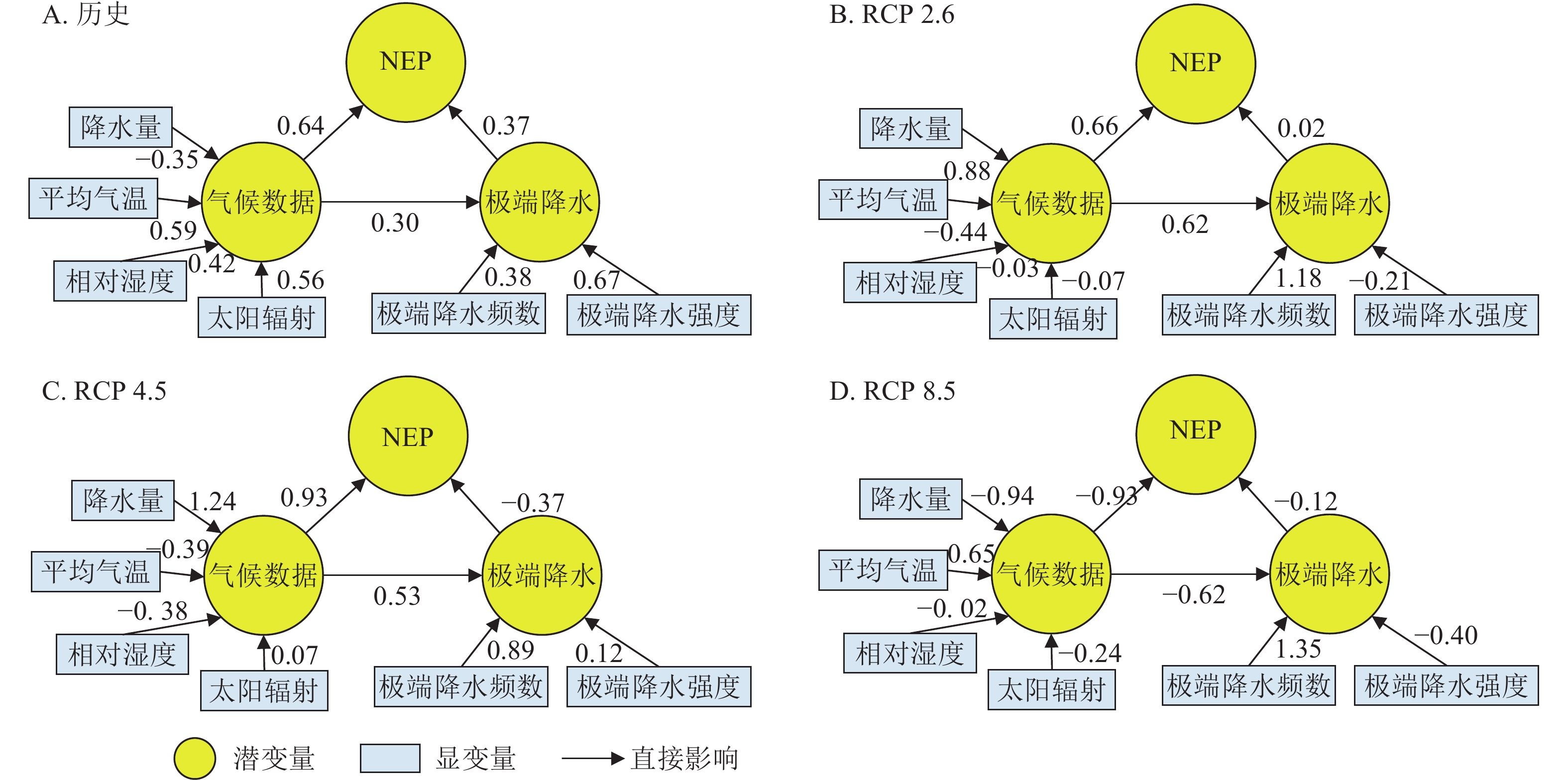
分析图5可知:1979—2079年丽水市气候因子对NEP的通径系数绝对值最大的均为降水,可见降水是森林生态系统NEP变化的重要驱动因子。由图5A得出:气候变化对NEP变化的直接影响为0.64,对极端降水的直接影响为0.30,气候变化通过影响极端降水进而影响NEP的间接影响为0.11 (0.30×0.37=0.11),极端降水对NEP的直接影响为0.37,因此1979—2015年丽水市极端降水对NEP产生积极影响。由图5B可知:在RCP 2.6情景下,2016—2079年丽水市气候变化对NEP变化的直接影响为0.66,对极端降水的直接影响为0.62,气候变化通过影响极端降水进而影响NEP的间接影响为0.01 (0.62×0.02=0.01),极端降水对NEP的直接影响不显著。由图5C可知:在RCP 4.5情景下,2016—2079年丽水市气候变化对NEP变化的直接影响为0.93,对极端降水的直接影响为0.53,气候变化通过极端降水对NEP产生的间接影响为0.20 (0.53×0.37=0.20),极端降水对NEP的直接影响为0.37,并且对NEP产生消极影响。由图5D可知:在RCP 8.5情景下,2016—2079年丽水市气候变化对NEP变化的直接影响为0.93,对极端降水的直接影响为0.62,气候变化通过影响极端降水进而影响NEP的间接影响为0.07 (0.62×0.12=0.07),极端降水对NEP的直接影响为0.12,对NEP变化产生消极影响。

图 5 历史(1979—2015年)及不同气候情景下未来(2016—2079年)下极端降水对NEP的影响
Figure 5. Effects of extreme precipitation on NEP under historical (1979-2015) and different climate scenarios (2016-2079)
-
本研究通过百分位阈值法研究了浙江省丽水市近百年的极端降水状况,结果表明:1979—2015年丽水市极端降水事件呈上升趋势,在丽水市东南部地区的上升趋势较大。RCP 2.6情景下,2016—2079年极端降水大部分呈下降趋势,RCP 4.5及RCP 8.5情景下,极端降水主要呈上升趋势,RCP 4.5情景相对于RCP 8.5情景的极端降水量、频数及强度增加更为明显。这与前人研究结果一致[19]。人类活动的加剧及地形影响,使浙江省温室气体增加,气温上升从而导致极端降水事件更容易发生[2−3]。极端降水在浙江西部及南部等地上升趋势较大的原因主要是由于南部及西部地区多丘陵山地,湿润气流遇到山脉等阻挡时被迫抬升,气温降低,容易形成地形雨。丽水位于浙江省西南部,全市多山地,比浙江的中部盆地和东北部冲积平原更容易发生极端降水事件,因此丽水市未来应加强洪涝灾害的预防。
长江流域及以南地区极端降水增多主要是由极端降水频数增加导致的[20]。本研究采用偏最小二乘法分析得知:1979—2079年极端降水频数对极端降水事件的通径系数大于极端降水强度对极端降水事件的通径系数,说明丽水市极端降水量的增加亦主要受极端降水频数的影响。另外,本研究是以年尺度分析极端降水事件变化。极端降水与季节有较强的相关性,并且极端降水的开始时间及持续时间都将直接或间接影响[7]丽水市气候的变化,因此,未来应将这些指标纳入丽水市极端降水变化特征的研究。
-
本研究发现:1979—2015年期间,丽水市森林NEP从1989年由碳源逐渐转为碳汇,这主要是由于1989年前人为干扰严重,导致森林质量下降,1989年以后,随着退耕还林政策的实施,全省森林质量得到改善,森林面积、年龄逐渐增加,使得碳汇能力增强[21]。2016—2079年3种气候情景下,丽水市森林碳汇均呈下降趋势,主要是受到森林年龄继续增加及气候变化的影响[10],其中随着未来极端降水事件的增加,森林NEP也呈下降趋势。
本研究通过InTEC模型模拟丽水市森林NEP的时空分布,虽然总体上精度较好,但仍然存在一些误差。首先,森林年龄对InTEC模型的模拟结果影响较大[10]。丽水市森林多属异龄林,森林资源清查的年龄数据为森林平均年龄,与森林实际情况有差别,通过空间插值后的数据也会存在误差。其次,本研究采用的3种气候情景的气象数据空间分辨率较低,通过线性插值生成的1 km分辨率气象数据对森林碳通量和极端降水空间解析不够细致,导致结果存在误差,但对长时间(百年尺度)碳通量演变和极端降水变化趋势仍具有一定参考价值。另外,本研究假设丽水市森林区域未来保持不变,然而实际上随着社会经济的发展,森林区域会发生改变,给模型模拟结果带来不确定性。因此,未来应进一步提高丽水森林空间分布、土壤、地形、气象等数据分辨率,更精确地模拟丽水市森林碳通量时空格局。
-
偏最小二乘通径模型分析结果表明:降水是影响NEP变化的重要因子。1979—2015年极端降水对NEP产生了积极影响。这与WANG等[5]的研究结果一致,可能得益于植被对降水有一定程度的适应和抵抗能力,当极端降水超过阈值时,会对生态系统造成消极影响[4]。一方面可能导致植被根系严重缺氧、呼吸减慢,最终引起植被死亡;另一方面极端降水频次和强度会加剧土壤侵蚀,导致地表水土流失,造成颗粒和有机碳从陆地生态系统流向河流生态系统,从而改变生态系统碳源汇的大小和空间分布。可见,气候变化强度的增加对生态系统消极影响的也会变得严重[22]。虽然偏最小二乘通径分析因子间存在直接和间接影响,但受模型算法限制无法得到超过生态系统抵抗力和恢复力的阈值,未来可以通过极端降水和气候综合敏感性分析、多情景模拟比较,进一步揭示极端降水对NEP影响的阈值,阐明极端降水影响森林NEP的关键。
-
浙江省丽水市历史、RCP 4.5及RCP 8.5情景下极端降水量、频数及强度均呈增加趋势,RCP 2.6呈下降趋势。极端降水事件上升趋势较高的地区主要位于丽水市南部和东南部地区。
1979—2015年丽水市森林NEP以18.44 g·m−2·a−1的速度增加,在1989年由碳源转变为碳汇。2016—2079年,3种气候情景下丽水市森林NEP均呈下降趋势,但碳汇总量呈上升趋势,说明在未来气候情景下,丽水市森林生态系统依然具有较高的碳汇潜力。
降水是影响NEP变化的重要驱动因子。虽然历史模拟显示极端降水对丽水NEP的影响为0.37,但未来气候变化情景下,除RCP 2.6情景不显著外,其余2种情景均对NEP产生显著消极影响,说明随着气候变化强度加剧,极端降水将对丽水市森林生态系统碳汇能力产生负面作用,应当积极采取森林经营措施,提高森林对极端气候事件抵抗力,使森林更好地服务实现“双碳”目标。
Spatiotemporal evolution of forest carbon sink in Lishui City and its response to extreme precipitation
-
摘要:
目的 模拟并分析浙江省丽水市森林净生态系统生产力(NEP)时空演变趋势,揭示其对极端降水的响应机制,为丽水市森林固碳潜力时空评价提供科学依据。 方法 综合样地调查、遥感观测、InTEC模型,模拟3种气候情景(RCP 2.6、RCP 4.5和RCP 8.5)下1979—2079年丽水市森林NEP时空格局,分析极端降水时空特征,构建结构方程模型,探讨极端降水对NEP 的影响。 结果 ①1979—2015年丽水市极端降水频率和强度呈增加趋势。未来气候情景下除RCP 2.6之外,RCP 4.5和RCP 8.5极端降水量、频率和强度仍持续增加,其中莲都区、缙云县以及景宁畲族自治县等地区极端降水事件上升趋势较强。②1979—2015年森林NEP 呈增加趋势,增长速度为18.44 g·m−2·a−1。2016—2079年,3种气候情景下森林NEP 均呈下降趋势,但碳汇总量呈上升趋势,最大NEP 累积达10.97 Pg。③1979—2015年极端降水对NEP 产生积极影响,2016—2079年除RCP 2.6影响不显著,其他2种情景下极端降水对NEP均产生消极影响,但RCP 8.5情景下极端降水较RCP 4.5对NEP 的影响更小。 结论 丽水市森林碳汇能力较强,在未来气候情景下仍具有较高的碳汇潜力,然而极端降水对NEP的影响也不容忽视。图5参22 -
关键词:
- 极端降水 /
- 森林碳汇 /
- 净生态系统生产力(NEP) /
- InTEC模型 /
- 浙江省丽水市
Abstract:Objective The objective is to simulate and analyze the spatiotemporal evolution trend of forest net ecosystem productivity (NEP) in Lishui City, and reveal its response mechanism to extreme precipitation, so as to provide data support for spatiotemporal assessment of forest carbon sink potential in Lishui. Method The spatiotemporal pattern of forest NEP in Lishui from 1979 to 2079 was simulated under three climate scenarios (RCP 2.6, RCP 4.5, and RCP 8.5) by integrating plot survey, remote sensing observation and InTEC model. The spatiotemporal characteristics of extreme precipitation were analyzed and a structural equation model was constructed to explore the impact of extreme precipitation on NEP. Result (1) The frequency and intensity of extreme precipitation in Lishui during 1979-2015 showed an increasing trend. In the future climate scenario (except RCP 2.6), the extreme precipitation, frequency and intensity of RCP 4.5 and RCP 8.5 would continue to increase, with a higher trend of extreme precipitation events in areas such as Liandu District, Jinyun County and Jingning She Autonomous County. (2) Forest NEP showed an increasing trend from 1979 to 2015, with a growth rate of 20.18 g·m−2·a−1. From 2016 to 2079, forest NEP showed a downward trend under the three climate scenarios, but the total carbon sink showed an increasing trend, with the maximum NEP accumulation reaching 10.97 Pg. (3) From 1979 to 2015, extreme precipitation had positive effects on NEP, while it had negative impacts on NEP under the other two scenarios (RCP 4.5 and RCP 8.5) during 2016-2079, except for RCP 2.6 scenario under which extreme precipitation had no significant effect on NEP. However, extreme precipitation under RCP 8.5 had less impact on NEP than that under RCP 4.5. Conclusion Forests in Lishui demonstrate strong carbon sink capacity, and have high carbon sink potential under future climate scenarios. However, the impact of extreme precipitation on NEP should not be ignored. [Ch, 5 fig. 22 ref.] -
图 1 历史(1979—2015年)及不同气候情景下未来(2016—2079年)丽水市极端降水的空间分布
Figure 1 Spatial distribution of extreme precipitation in Lishui City under historical (1979-2015) and future (2016-2079) climate scenarios
图 2 历史(1979—2019年)及不同气候情景下未来(2019—2079年)丽水市年极端降水累计次数的年际变化
Figure 2 Interannual changes of the cumulative number of extreme precipitation in Lishui City under historical (1979-2019) and different climate scenarios (2019-2079)
图 3 历史(1979—2015年)及不同气候情景下未来(2016—2079年)丽水市森林NEP的年际变化
Figure 3 Historical (1979-2015) and future (2016-2079) interannual changes of forest NEP in Lishui City under different climate scenarios
图 4 1979—2015基准年以及不同气候情景下2016—2079年丽水市NEP的空间分布和时空演变特征
Figure 4 Spatial distribution of NEP and linear trend in Lishui City during 1979-2015 base year and 2016-
2079 under different climate scenarios -
[1] 徐雨晴, 肖风劲, 於琍. 中国森林生态系统净初级生产力时空分布及其对气候变化的响应研究综述[J]. 生态学报, 2020, 40(14): 4710 − 4723. XU Yuqing, XIAO Fengjin, YU Li. Review of spatio-temporal distribution of net primary productity in forest ecosystem and its responses to climate change in China [J]. Acta Ecologica Sinica, 2020, 40(14): 4710 − 4723. [2] 丁楠, 沙晓军, 高颖会, 等. 1961—2010年浙江省极端降水特征分析[J]. 中国农村水利水电, 2017(2): 104 − 107. DING Nan, SHA Xiaojun, GAO Yinghui, et al. The characteristics of extreme precipitation in Zhejiang Province from 1961 to 2010 [J]. China Rural Water and Hydropower, 2017(2): 104 − 107. [3] GAO Lu, HUANG Jie, CHEN Xingwei, et al. Contributions of natural climate changes and human activities to the trend of extreme precipitation [J]. Atmospheric Research, 2018, 205: 60 − 69. [4] WEI Siyu, HAN Guangxuan, CHU Xiaojing, et al. Prolonged impacts of extreme precipitation events weakened annual ecosystem CO2 sink strength in a coastal wetland [J/OL]. Agricultural and Forest Meteorology, 2021, 310: 108655[2023-12-23]. doi: 10.1016/j.agrformet.2021.108655. [5] WANG Miaomiao, ZHAO Jian, WANG Shaoqiang, et al. Detection and attribution of positive net ecosystem productivity extremes in China’s terrestrial ecosystems during 2000−2016 [J/OL]. Ecological Indicators, 2021, 132: 108323[2023-12-23]. doi: 10.1016/j.ecolind.2021.108323. [6] LIU Bingjun, CHEN Junfan, CHEN Xiaohong, et al. Uncertainty in determining extreme precipitation thresholds [J]. Journal of Hydrology, 2013, 503: 233 − 245. [7] 卢珊, 胡泽勇, 王百朋, 等. 近56年中国极端降水事件的时空变化格局[J]. 高原气象, 2020, 39(4): 683 − 693. LU Shan, HU Zeyong, WANG Baipeng, et al. Spatio-temporal patterns of extreme precipitation events over China in recent 56 years [J]. Plateau Meteorology, 2020, 39(4): 683 − 693. [8] PIAO Shilong, HE Yue, WANG Xuhui, et al. Estimation of China’s terrestrial ecosystem carbon sink: methods, progress and prospects [J]. Science China (Earth Sciences), 2022, 65(4): 641 − 651. [9] CHEN Jing, CHEN Wenjun, LIU Jane, et al. Annual carbon balance of Canada’s forests during 1895–1996 [J]. Global Biogeochemical Cycles, 2000, 14(3): 839 − 849. [10] MAO Fangjie, DU Huaqiang, ZHOU Guomo, et al. Simulated net ecosystem productivity of subtropical forests and its response to climate change in Zhejiang Province, China [J/OL]. Science of the Total Environment, 2022, 838: 155993[2023-12-31]. doi: 10.1016/j.scitotenv.2022.155993. [11] WANG Shaoqiang, CHEN Jingming, JU Weimin, et al. Carbon sinks and sources in China’s forests during 1901−2001 [J]. Journal of Environmental Management, 2007, 85(3): 524 − 537. [12] LI Xuejian, DU Huaqiang, MAO Fangjie, et al. Mapping spatiotemporal decisions for sustainable productivity of bamboo forest land [J]. Land Degradation &Development, 2020, 31(8): 939 − 958. [13] 俞静芳, 余树全, 张超, 等. 应用CASA模型估算浙江省植被净初级生产力[J]. 浙江农林大学学报, 2012, 29(4): 473 − 481. YU Jingfang, YU Shuquan, ZHANG Chao, et al. Net primary productivity estimates for Zhejiang Province based on the CASA model [J]. Journal of Zhejiang A&F University, 2012, 29(4): 473 − 481. [14] 肖晶晶, 李正泉, 郭芬芬, 等. 浙江省1901—2017年降水序列构建及变化特征分析[J]. 气候变化研究进展, 2018, 14(6): 553 − 561. XIAO Jingjing, LI Zhengquan, GUO Fenfen, et al. Construction and analysis of annual precipitation series from 1901 to 2017 in Zhejiang Province [J]. Climate Change Research, 2018, 14(6): 553 − 561. [15] 孔锋, 史培军, 方建, 等. 全球变化背景下极端降水时空格局变化及其影响因素研究进展和展望[J]. 灾害学, 2017, 32(2): 165 − 174. KONG Feng, SHI Peijun, FANG Jian, et al. Advances and prospects of spatiotemporal pattern variation of ex-treme precipitation and its affecting factors under the background of global climate change [J]. Journal of Catastrophology, 2017, 32(2): 165 − 174. [16] LI Yangguang, HAN Ning, LI Xuejian, et al. Spatiotemporal estimation of bamboo forest aboveground carbon storage based on landsat data in Zhejiang, China [J]. Remote Sensing, 2018, 10(6): 898 − 898. [17] SAXTON K E, RAWLS W J, ROMBERGER J S, et al. Estimating generalized soil-water characteristics from texture [J]. Soil Science Society of America Journal, 1986, 50(4): 1031 − 1036. [18] MAO Fangjie, DU Huaqiang, ZHOU Guomo, et al. Coupled LAI assimilation and BEPS model for analyzing the spatiotemporal pattern and heterogeneity of carbon fluxes of the bamboo forest in Zhejiang Province, China [J]. Agricultural and Forest Meteorology, 2017, 242: 96 − 108. [19] 莫康, 刘光裕, 贺治国, 等. 气候变化下浙江省极端降水时空变化研究[J]. 中国农村水利水电, 2021(3): 74 − 79, 87. MO Kang, LIU Guangyu, HE Zhiguo, et al. Research on the spatial-temporal changes of extreme precipitation in Zhejiang Province under climate changes [J]. China Rural Water and Hydropower, 2021(3): 74 − 79, 87. [20] 江志红, 丁裕国, 陈威霖. 21世纪中国极端降水事件预估[J]. 气候变化研究进展, 2007, 3(4): 202 − 207. JIANG Zhihong, DING Yuguo, CHEN Weilin. Projection of precipitation extremes for the 21st century over China [J]. Advances in Climate Change Research, 2007, 3(4): 202 − 207. [21] 蔡丽莎, 陈先刚, 郭颖, 等. 贵州省退耕还林工程碳汇潜力预测[J]. 浙江林学院学报, 2009, 26(5): 722 − 728. CAI Lisha, CHEN Xiangang, GUO Ying, et al. Carbon sequestration potential with the Grain for Green Program in Guizhou Province [J]. Journal of Zhejiang Forestry College, 2009, 26(5): 722 − 728. [22] PIŃSKWAR I. Complex changes of extreme precipitation in the warming climate of Poland [J]. International Journal of Climatology, 2022, 42(2): 817 − 833. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240141
 点击查看大图
点击查看大图
计量
- 文章访问数: 2005
- HTML全文浏览量: 553
- PDF下载量: 69
- 被引次数: 0
