




-
西红花Crocus sativus是鸢尾科Iridaceae番红花属Crocus著名药用植物,其干燥柱头(雌蕊)为名贵药材和香料,素有“植物黄金”之称[1−3]。西红花为不育同源三倍体(2n=3x=24),依赖球茎进行无性繁殖,其顶芽为典型的花叶混合芽[4],在发育过程中同时分化叶原基与花原基,通常仅顶芽可分化1~3朵花并抽生相应的叶片;而侧芽成花能力极低,多维持叶芽状态,这导致球茎繁殖系数低,单位产量有限[5−6]。因此,解析西红花花原基分化的分子调控机制,可为促进顶芽开花、诱导侧芽成花提供理论依据,对提高雌蕊产量具有重要实践意义。
高等植物从营养生长转向生殖生长的开花过程,是由复杂的基因网络精细调控,主要涉及光周期、春化、自主、赤霉素、温度及年龄等多条调控途径[7−9]。FLOWERING LOCUS T (FT)基因编码的成花素蛋白,是整合多条环境与内源信号通路的关键因子,在茎顶端分生组织触发花原基分化[10−12]。FT基因在植物中高度保守,如拟南芥Arabidopsis thaliana AtFT过表达导致极早开花,而其突变体则延迟开花[13],大豆Glycine max GmFT2a/GmFT5a在短日照下高表达并促进开花[14];矮牵牛Petunia × hybrida PhFT4亦具有类似功能[15]。因西红花是三倍体,其基因家族解析复杂。在西红花单倍型基因组组装完成之前,已有研究报道了其6个FT同源基因(CsatFT1~CsatFT6)[16−17]。KALIA等[16]研究发现:CsatFT1与CsatFT2在叶组织中高表达,可能参与营养生长;CsatFT3在顶芽中特异性表达,推测与成花诱导相关。然而,王桢等[17]研究发现:CsatFT3在顶芽中几乎不表达,且异源转化实验表明CsatFT1、CsatFT2和CsatFT3均能不同程度地促进拟南芥和烟草Nicotiana tabacum开花,提示西红花中可能存在多个功能重叠的FT同源基因,但其中与花原基分化直接相关、并决定成花与否的关键成员仍未明确。
近年来,XU等[18]完成了西红花单倍型基因组的组装工作,为系统解析其多倍体基因家族提供了基础。本研究基于该基因组数据,对西红花PEBP基因家族成员进行全面鉴定,并通过分析花原基分化关键阶段的转录组数据,结合多种逆境胁迫(低温、干旱、低氧)处理的成花/非成花样本,筛选出与成花密切相关的FT同源基因Csativus09980。通过系统进化、序列分析、亚细胞定位及拟南芥异源过表达等功能验证,明确Csativus09980在促进西红花花原基分化中的关键作用,为阐明西红花成花分子机制及分子育种提供新的基因资源与理论依据。
-
所用西红花球茎购自浙江建德。试验采用常规二段式栽培模式进行管理,即在每年11月室内开花采收后,将球茎移栽至大田,翌年5月初采收球茎并保存于室内阴凉通风处。
烟草种子与拟南芥哥伦比亚野生型(Col)种子,种植基质由蛭石、腐殖质、营养土按质量比1∶1∶1混匀配制,播种后覆盖保鲜膜保湿,并在22 ℃,16 h光照/ 8h黑暗环境中培养,待烟草和拟南芥长至10 d时进行移栽,新移栽的幼苗需要覆盖保鲜膜进行保湿,移栽7d后,去除保鲜膜,常规培养。
-
选取大小均一、质量范围为(20 ± 2) g的健康球茎,随机分为2个组,分别置于9 ℃(低温,非成花)和22 ℃ (常温,成花)的光照培养箱中培养,相对湿度为60%~70%。
-
选取(20 ± 2) g的球茎于休眠期去除褐色外皮,使球茎早期脱水从而达到早期极度干旱胁迫的目的。
-
选取(20 ± 2) g的球茎,放入人工气候箱中,通入按体积分数配制的低氧混合气体(3%O2 + 5%CO2 +92%N2),在相对湿度为60%~70%条件下培养。
-
根据已有文献报道[19],在花原基分化前后的6个关键发育时期[休眠期(S1)、休眠打破期(S2)、花原基分化早期(S3)、花原基分化中期(S4)、花原基分化晚期(S5)、花器官形成期(S6)],分别采集常温处理组和低温处理组的球茎顶芽组织。各处理在每个时期采集60个球茎顶芽,每20个顶芽混合为1个生物学重复,共设置3个生物学重复。样品迅速置于液氮中冷冻,−80 ℃保存,用于总RNA提取及转录组建库、测序。
-
为验证候选基因的表达与成花能力的相关性,对常温对照组、干旱处理组、低氧胁迫组于S6期剥取顶芽并在体视显微镜下观察有无花器官,分别采集成花与非成花顶芽样品。每个样品由10个球茎顶芽混合组成,设3个生物学重复。所有样品液氮速冻,−80 ℃保存,用于RNA提取及实时荧光定量PCR (RT-qPCR)分析。
-
从Pfam数据库中获得PEBP结构域(编号PF01161),利用HMMER软件对西红花基因组中的所有蛋白序列进行搜索,初步筛选得到29条含有PEBP结构域的候选氨基酸序列。采用在线分析工具SMART和美国国家生物技术信息中心保守结构域数据库(NCBI-CDD)对其进行保守结构域验证,结果表明上述序列均含有部分或完整的PEBP结构域。
采用DNAMAN软件对序列进行多重比对,并通过NCBI BLAST进行同源性分析。显示在29条含有PEBP结构域的序列中,共鉴定出17条FT-like 基因序列、5条MFT-like基因序列和7条TFL1-like基因序列。其中,17条FT-like序列中有3条(Csativus
38290 、Csativus53610 和Csativus53609 ) PEBP结构域不完整,故不参与后续分析。从GenBank数据库下载的代表性植物拟南芥及水稻Oryza sativa的FT、MFT和TFL同源蛋白序列,并与剩余26条含有完整PEBP结构域的序列一起,采用MEGA 7.0软件基于邻接法构建系统进化树,Bootstrap分析重复1 000次[20]。
-
收集新鲜的植物组织并在液氮中快速冷冻,每个样品3个生物学重复,提取样本RNA后进行质控检测,然后通过Illumina平台进行测序。原始测序数据去除接头序列和低质量读数后进行数据分析。
使用HISAT2软件将reads比对至参考基因组,随后利用StringTie软件进行转录本组装。采用R软件中的cor函数计算样品间的Pearson相关系数,并进行主成分分析(PCA)。利用DESeq2软件包进行差异表达基因分析,以假发现率(FDR)<0.05,且|log2FC| ≥1 (FC为表达量变化倍数)为筛选标准,鉴定显著差异表达基因[21]。采用R软件中的gplots包对差异表达基因进行层次聚类分析。进一步利用R软件中的clusterProfiler包进行功能富集分析,包括基因本体论(GO)分析和京都基因与基因组百科全书(KEGG)通路分析,显著性阈值为P<1 × 10−5。
-
使用TaKaRa MiniBEST Plant RNA Extraction Kit提取总RNA,NanoDrop定量。取1 μg RNA用PrimeScript™ RT reagent Kit with gDNA Eraser进行基因组DNA去除及cDNA合成。以Tublin为内参基因,使用TB Green® Premix Ex Taq™ II在QuantStudio™ 5实时荧光定量PCR系统上进行扩增。反应程序为95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,40个循环。采用2−ΔΔCt法计算基因相对表达量[22]。所用引物序列见表1。
表 1 引物序列
Table 1. Primer sequences used in this study
引物名称 正向引物(5′→3′) 反向引物(5′→3′) CsatFT1q ATGAGTAGGGATTCGCTGGTGCTCG AACCTCCCGAGACTGGTACGTCACC CsatFT2q TGGCTAAACAAACGAGCTACGACTA CAATCACTCTACCAACAACCAAAGG CsatFT3q ATATGGCCGCAGCTACTAGAGAAAC TCCACCATCACCAGAGTGTAGAAAT CsatFT2-1q CCGAGGGTCGAGGTCGGAG GCGCATATACCGTTTGCCGTC CsatFT2-2q CCACCGGTTTGTCTTCATGC CCGACTCCCTCTGACAGTTGA CsatFT2-2CDS扩增 ATGGTTAGGGAGAGGAATCCTTTGG GCTCCGGTGGGAGAAGGATTTAA CsatFT2-2过表达载体构建 ACACGGGGGACTCTTGACATGGTTA
GGGAGAGGAATCCTCCTTTACTAGTCAGATCTACTTAA
ATCCTTCTCCCACCGGAGCsatFT2-2亚细胞定位 ACACGGGGGACTCTTGACATGGTTA
GGGAGAGGAATCCTCCTTTACTAGTCAGATCTACAATC
CTTCTCCCACCGGAGC -
常温成花组各时期另取5个顶芽,乙醇-甲醛-冰乙酸(FAA)固定液固定48 h,再进行包埋处理,随后利用轮转式切片机完成切片,切片厚度为8 μm,经脱蜡、水化、番红-固绿染色、透明、封片后于光学显微镜观察并拍照[23]。
-
根据FT基因的引物序列,以基因组DNA (gDNA)和cDNA为模板进行PCR扩增。通过测序获得FT基因序列后,设计特异性正向和反向引物,用于构建超量表达载体。扩增出的cDNA特异片段通过DNA琼脂糖凝胶回收试剂盒进行回收,连接pMD20 T载体,热激转化大肠埃希菌Escherichia coli DH5α感受态细胞。挑选阳性克隆扩大培养后,提质粒,再通过相应的酶切位点,酶切,回收并连接pCAMBIA1302构建的过表达载体35S::Csativus09980。
-
以西红花顶芽cDNA为模板扩增Csativus09980基因的全长编码序列(CDS),通过同源重组技术将其克隆至pCAMBIA1302-35S::GFP双元载体。经测序验证无误后,将重组质粒35S::Csativus09980 -GFP与空载体35S::GFP分别通过电击转化法导入农杆菌Agrobacterium tumefaciens GV3101感受态细胞。于含有50 mg·L−1卡那霉素和25 mg·L−1利福平的Luria-Bertani (LB)液体培养基中将含有重组质粒和空载体的农杆菌培养至吸光度D(600)为0.6~0.8,离心收集菌体。将农杆菌培养物悬浮于缓冲液[10 mmol·L−1MgCl2、10 mmol·L−1 2-(N-吗啉)乙磺酸(MES)、100 μmol·L−1乙酰丁香酮(AS),pH 5.7],采用注射器将农杆菌悬浮液注入4周龄烟草叶片背面进行瞬时表达。培养48 h后,利用共聚焦显微镜观察绿色荧光蛋白(GFP)信号,以空载体pCAMBIA1302-35S::GFP为对照[24]。
-
将35S::Csativus09980-GFP重组质粒通过电击转化导入农杆菌GV3101,采用花序浸染法侵染野生型拟南芥花序。收获T0代种子,在含25 mg·L−1潮霉素的MS平板上筛选阳性转化苗,获得T1代植株。T1代自交收获T2代种子,种植T2代种子并筛选获得纯合株系用于表型分析。观察并记录转基因株系和野生型的莲座叶数量及开花时间。采用SPSS 27.0进行独立样本t检验,显著性水平为0.05。
-
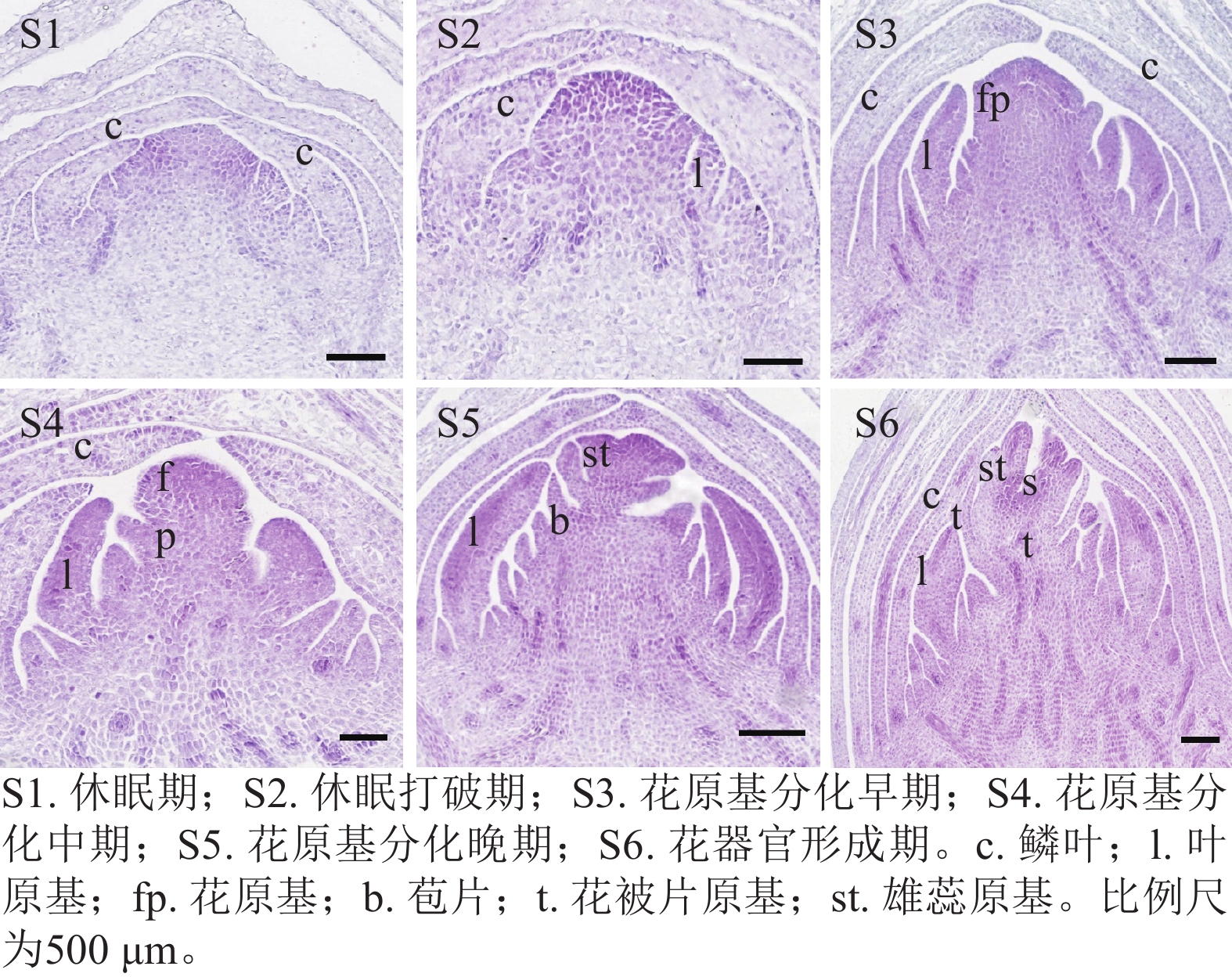
通过石蜡切片观察西红花球茎顶芽在6个连续发育时期的形态变化(图1)。结果发现:球茎休眠期(S1)顶端分生组织处于静止状态,休眠打破期(S2)生长锥开始活化并明显增大,花原基分化早期(S3)首先分化形成叶原基,花原基分化中期(S4)花原基在叶原基腋部开始形成,花原基分化晚期(S5)花被片和雄蕊原基依次分化,花器官形成期(S6)雌蕊等花器官清晰可辨。表明S3~S6期为西红花花原基分化的关键时期。
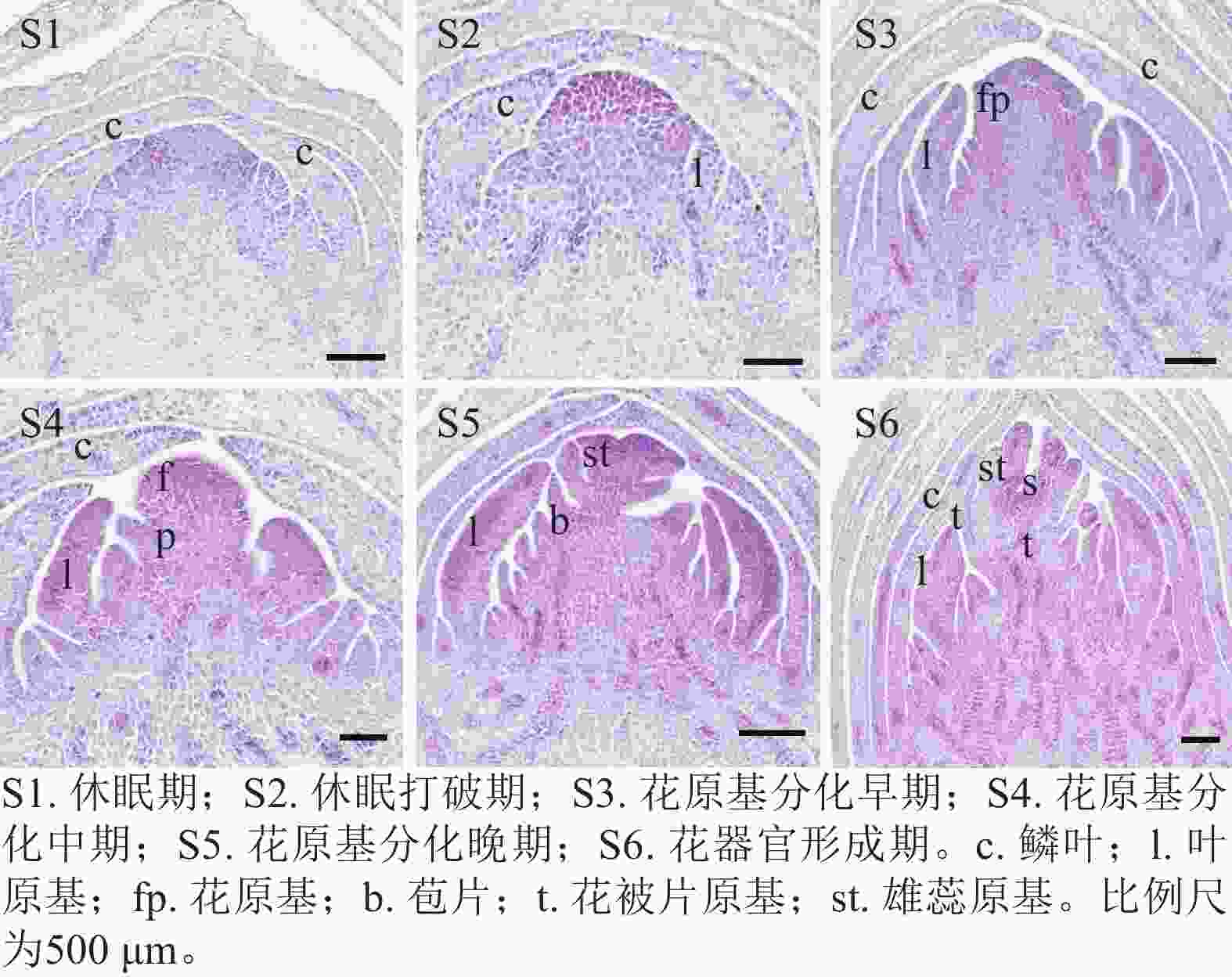
图 1 西红花花原基分化连续过程的形态特征
Figure 1. Morphological characteristics of the successive stages of floral primordium differentiation in C. sativus
-
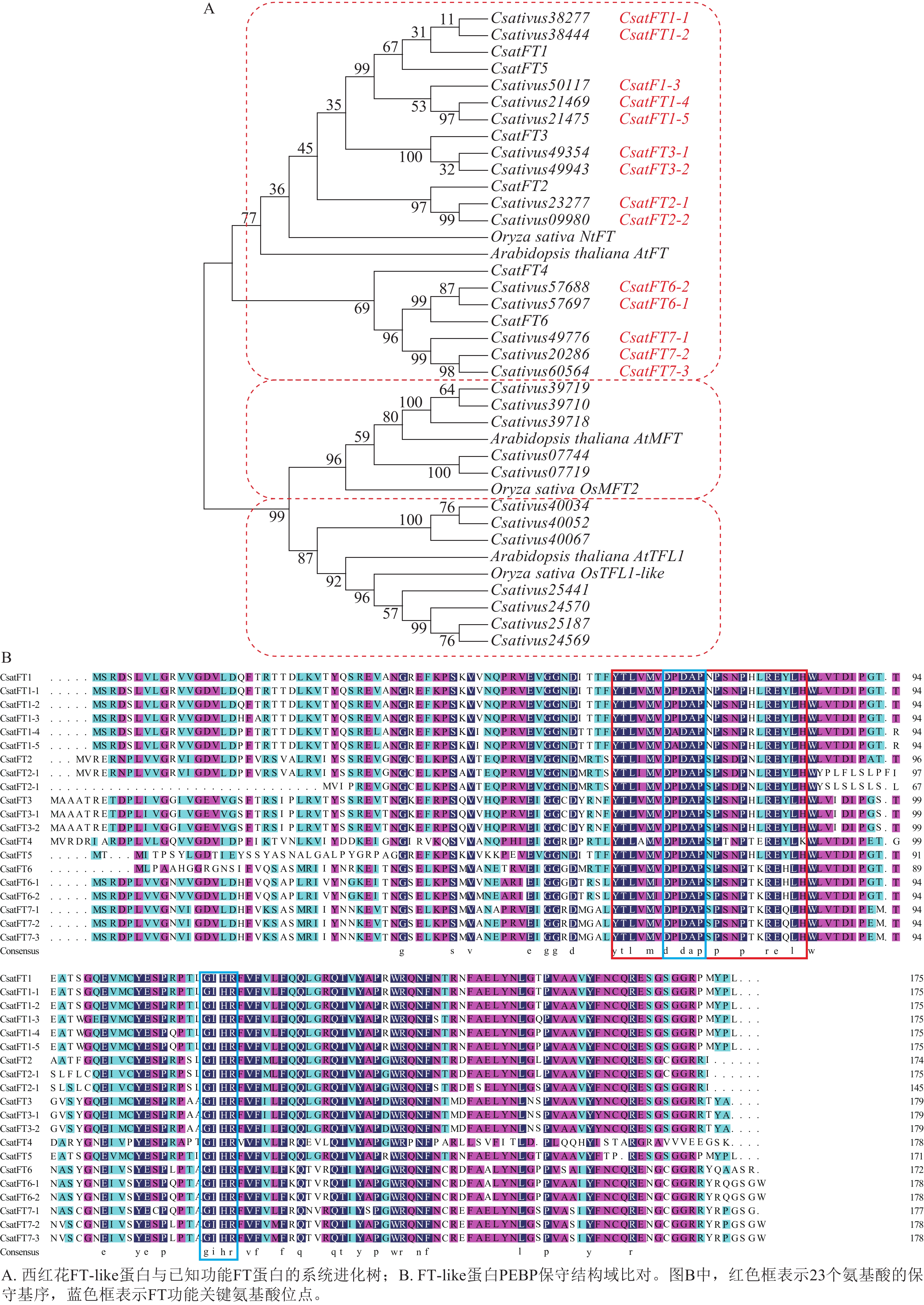
对西红花14个FT-like基因与已报道CsatFT1-6进行氨基酸序列比对,并构建系统进化树,根据序列同源性及进化关系对14个FT-like基因进行命名,可分为7个聚类,结果如图2A所示。氨基酸序列比对如图2B所示。除文献报道的CsatFT4外,其余序列均含有完整的PEBP家族保守结构域,其典型序列特征为[FYL]-x-[LVM]-[LIVF]-x-[TIVM]-[DC]-P-D-x-P-[SNG]-x(10)-H[25],并包含FT蛋白中与促开花功能相关的关键氨基酸残基(Tyr85/Gln140)[26]。此外CsatFT1-1、CsatFT1-2与CsatFT1以及CsatFT3-1、CsatFT3-2与CsatFT3的氨基酸序列完全一致,其余聚类间序列相似度均大于83% (附表1)。
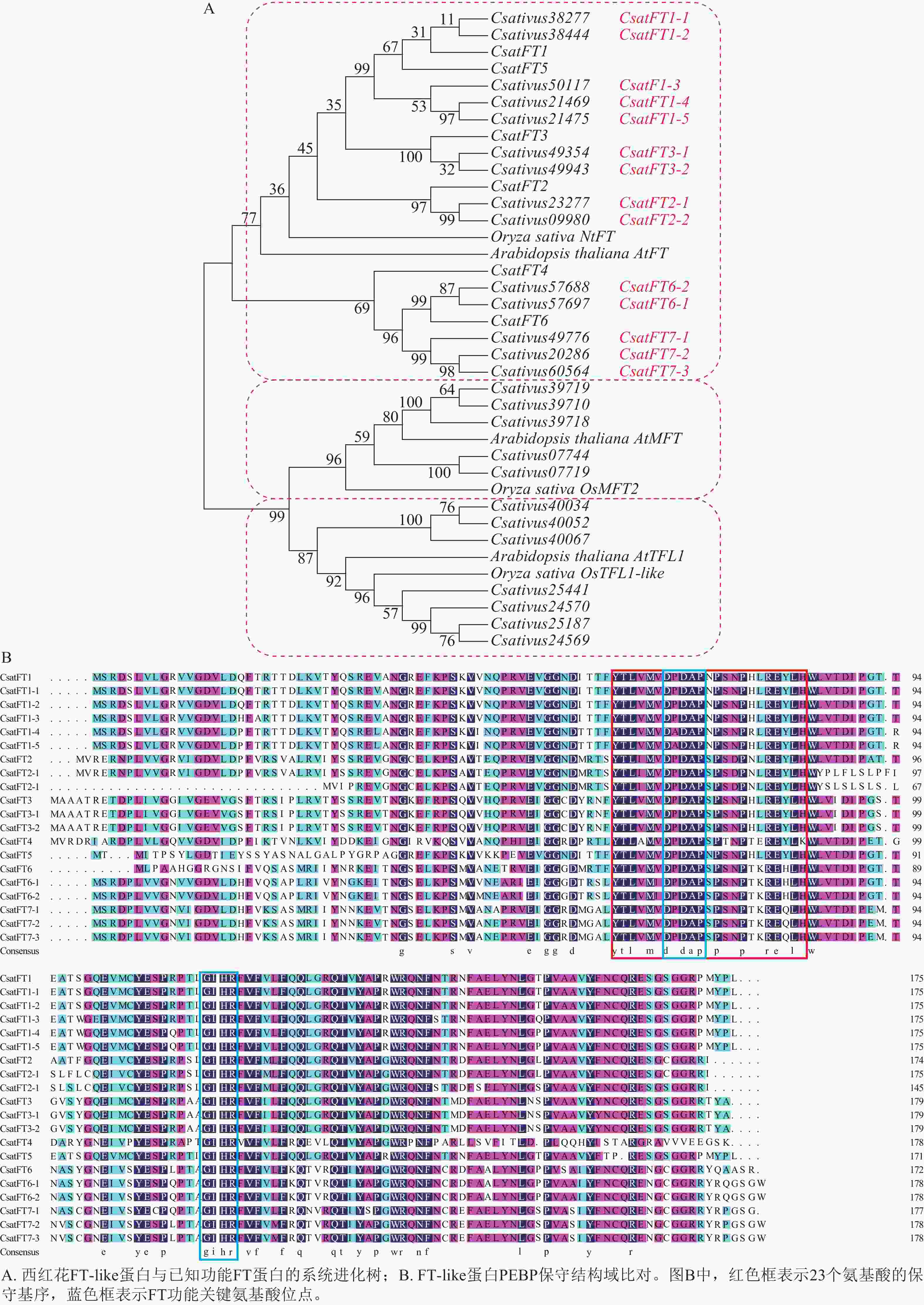
图 2 FT-like基因系统进化与序列比对分析
Figure 2. Phylogenetic and sequence alignment analyses of FT-like genes
-
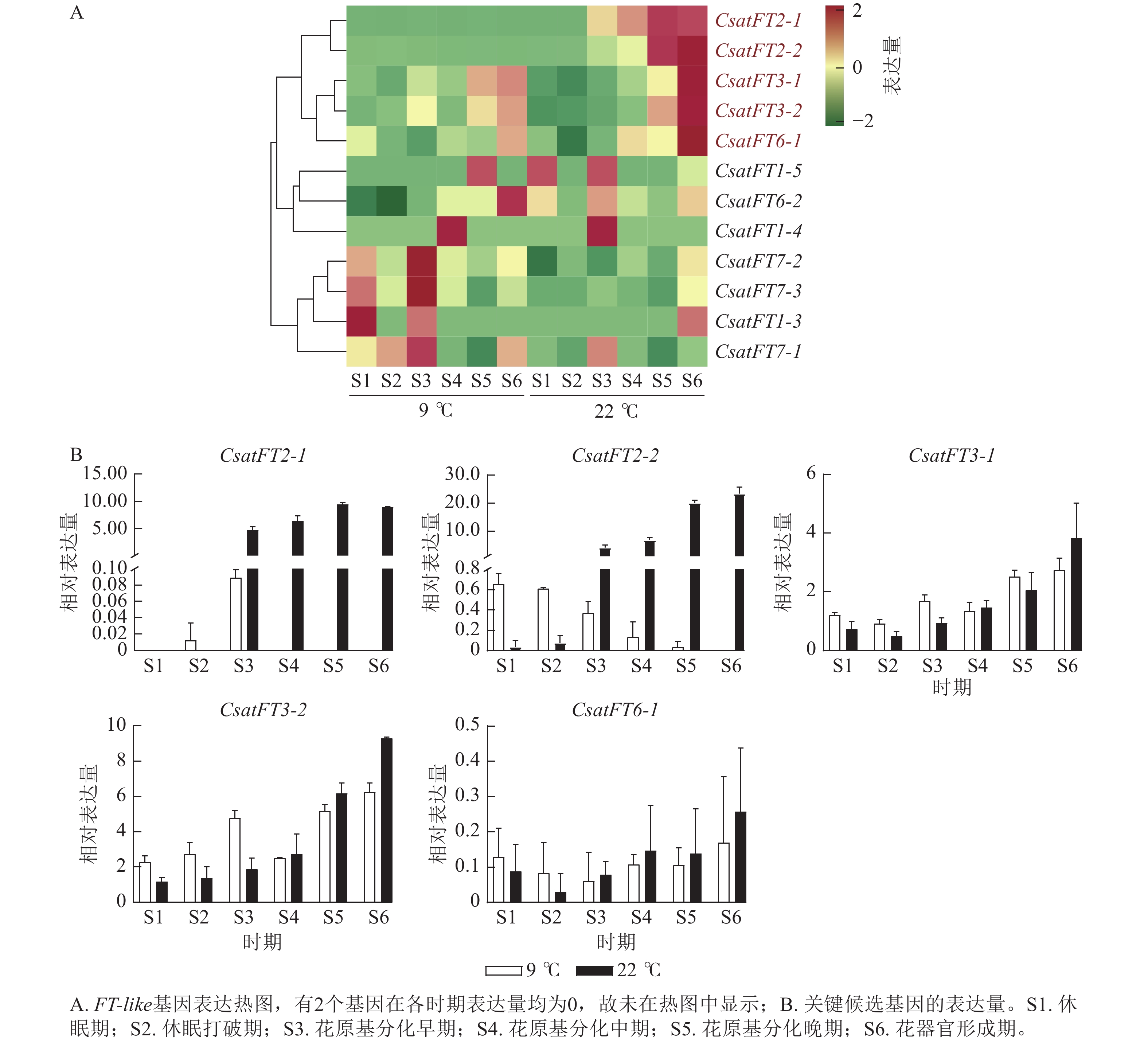
FT-like基因在常温(22 ℃,成花)与低温(9 ℃,非成花)条件下顶芽6个发育时期的表达模式如图3A所示。结果表明:CsatFT2-1和CsatFT2-2在常温成花顶芽中自花原基分化早期起表达量持续上调,且明显高于低温非成花顶芽(图3B)。CsatFT3-1、CsatFT3-2和CsatFT6-1在常温成花顶芽中虽呈上调趋势,但与低温非成花顶芽之间差异不明显,其中CsatFT6-1在2种条件下表达量均较低。CsatFT2-1和CsatFT2-2在非成花样本中表达量极低,甚至无法检测到,而在成花样本中随花芽分化进程逐渐升高,表明这2个基因可能在成花启动过程中发挥重要作用。
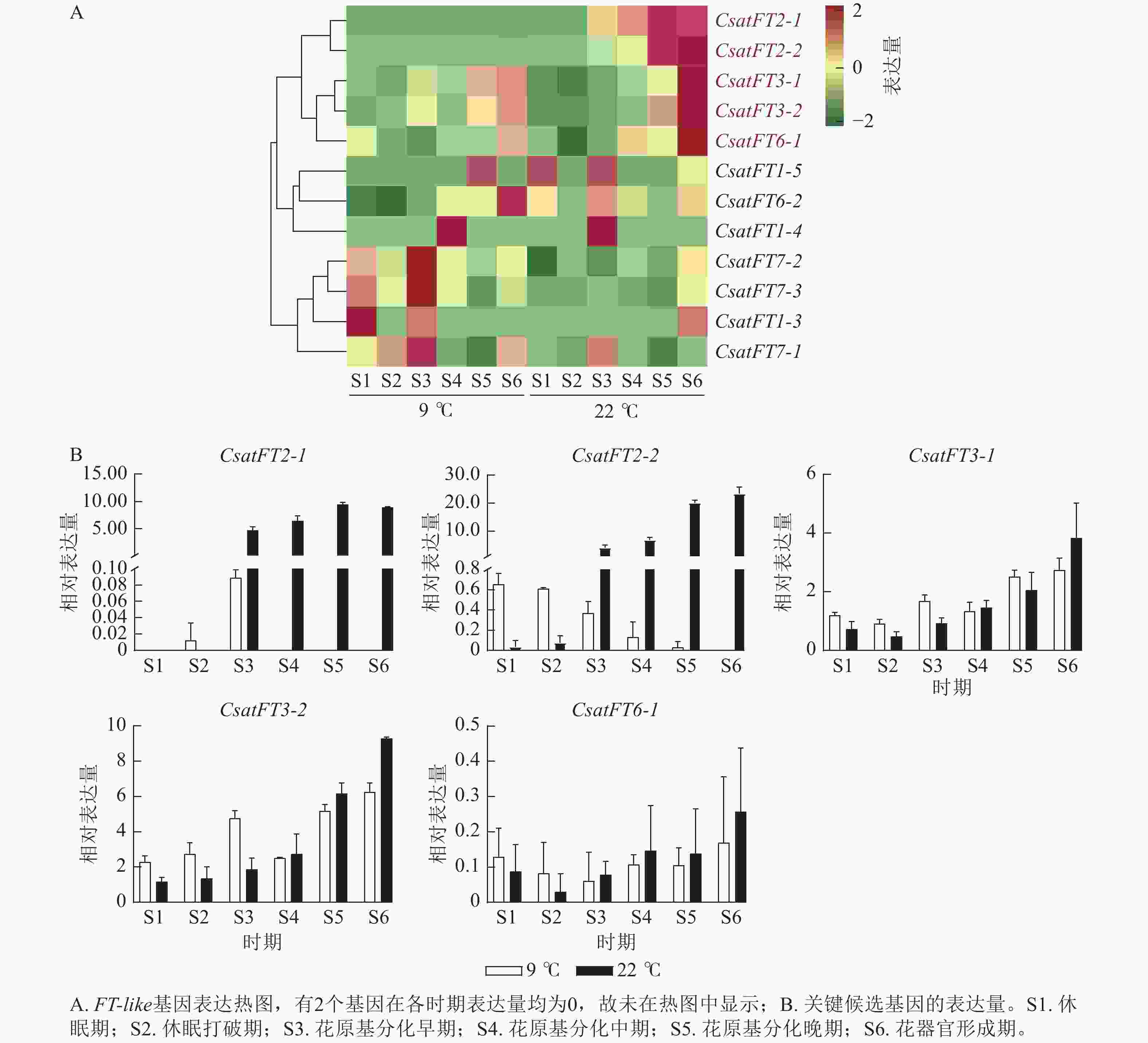
图 3 西红花FT-like基因表达谱分析
Figure 3. Expression-profile analysis of crocus FT-like genes
-
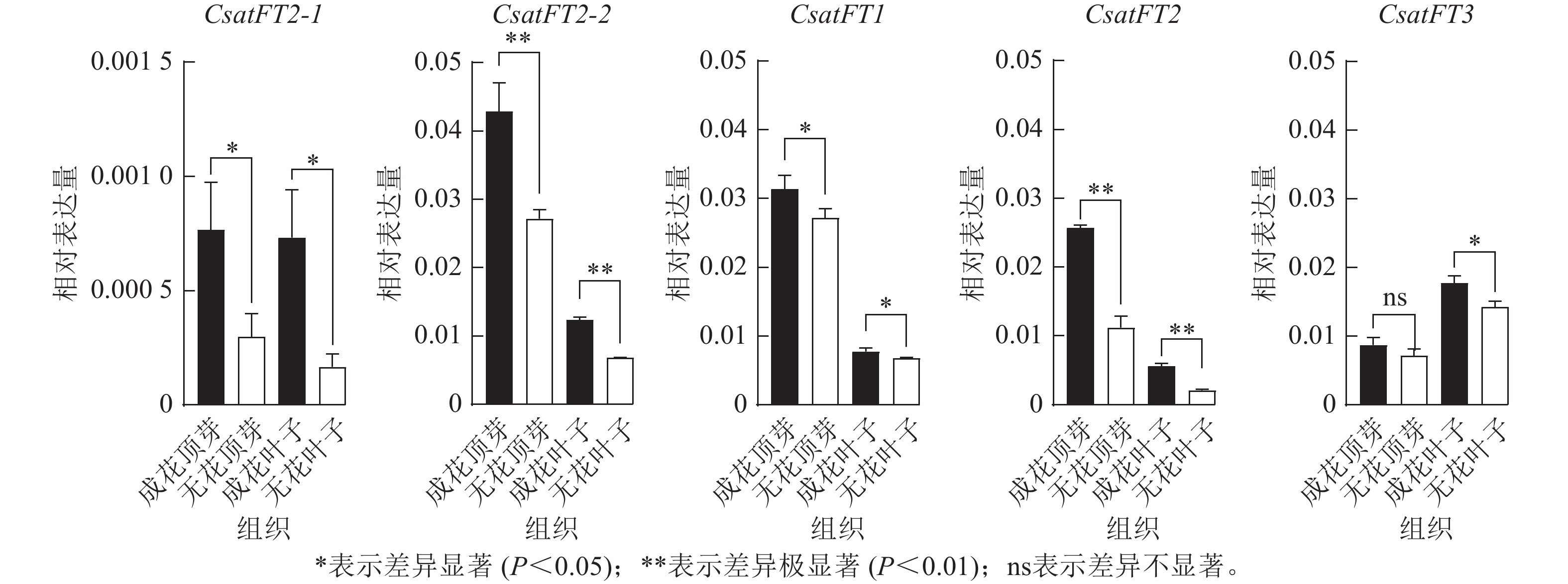
采用RT-qPCR技术分析花原基分化S6期西红花成花与无花球茎顶芽和叶片中CsatFT2-1、CsatFT2-2、CsatFT1、CsatFT2和CsatFT3的表达模式(图4)。结果表明:在成花顶芽中,CsatFT2-2与CsatFT2的表达量均显著高于无花顶芽(P<0.01);其中,CsatFT2-2的表达水平约为CsatFT2的2倍。在叶片中,尽管CsatFT2-2与CsatFT2在成花样本中的表达也显著高于无花样本(P<0.01),但其表达量明显低于顶芽组织。CsatFT3在成花叶片中表达显著上调(P<0.05),但在顶芽中的表达量仅为叶片的一半,且在成花与无花顶芽之间差异不显著。CsatFT1在顶芽和叶片中的表达差异最小。CsatFT2-1与CsatFT2-2在成花与非成花样本中的表达变化较为明显,但CsatFT2-1的表达量整体低于其他基因。综上,CsatFT2-2在成花顶芽中呈较高表达水平,提示其可能在西红花花原基分化及成花诱导过程中发挥重要作用。
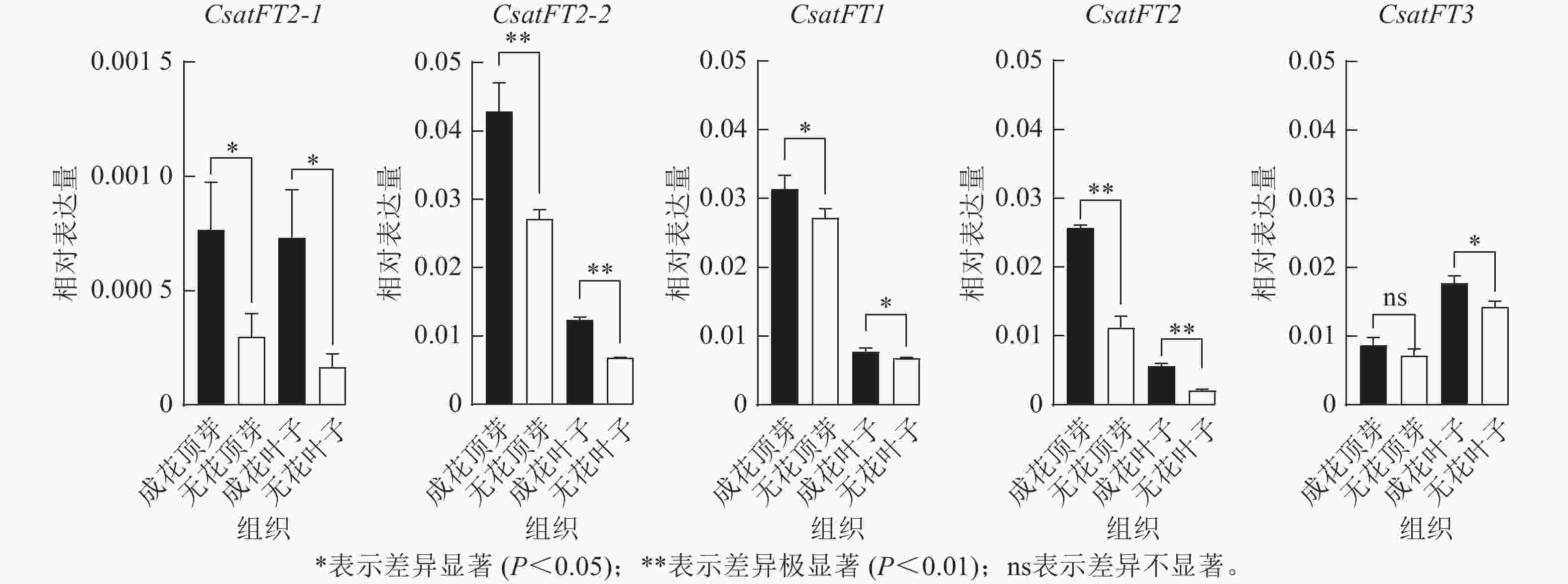
图 4 FT-like基因在顶芽和叶片组织中的表达模式分析
Figure 4. Expression patterns of FT-like genes in apical buds and leaves
-
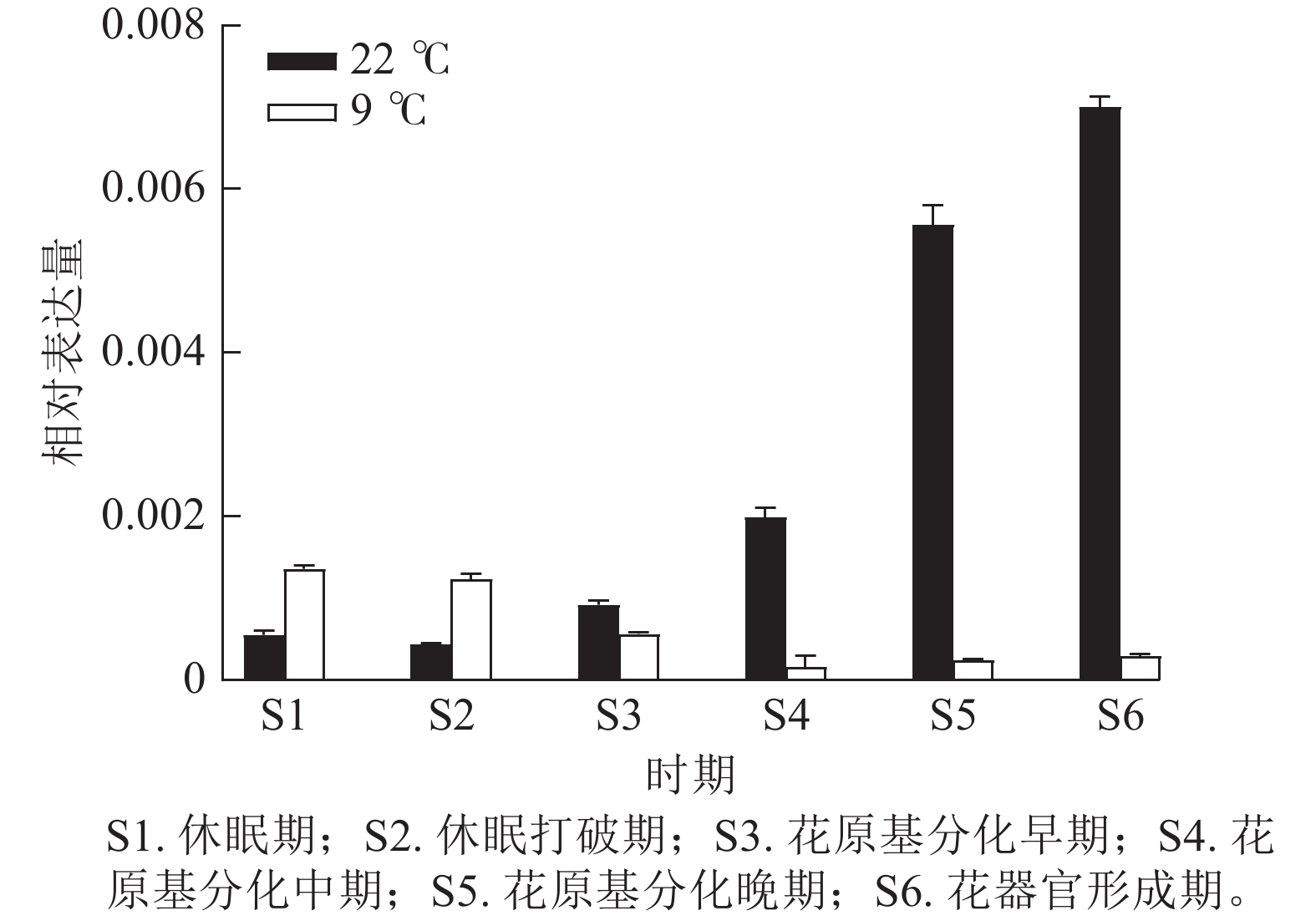
为验证转录组数据,通过RT-qPCR分析了CsatFT2-2在22 ℃ (成花)与9 ℃ (非成花)条件下顶芽不同发育阶段的表达模式(图5)。结果显示:在22 ℃条件下,CsatFT2-2的表达量在花原基分化及花器官形成阶段上调并持续增加;而在9 ℃非成花条件下,其表达受到明显抑制。表明低温抑制CsatFT2-2的表达,这与转录组测序结果一致。
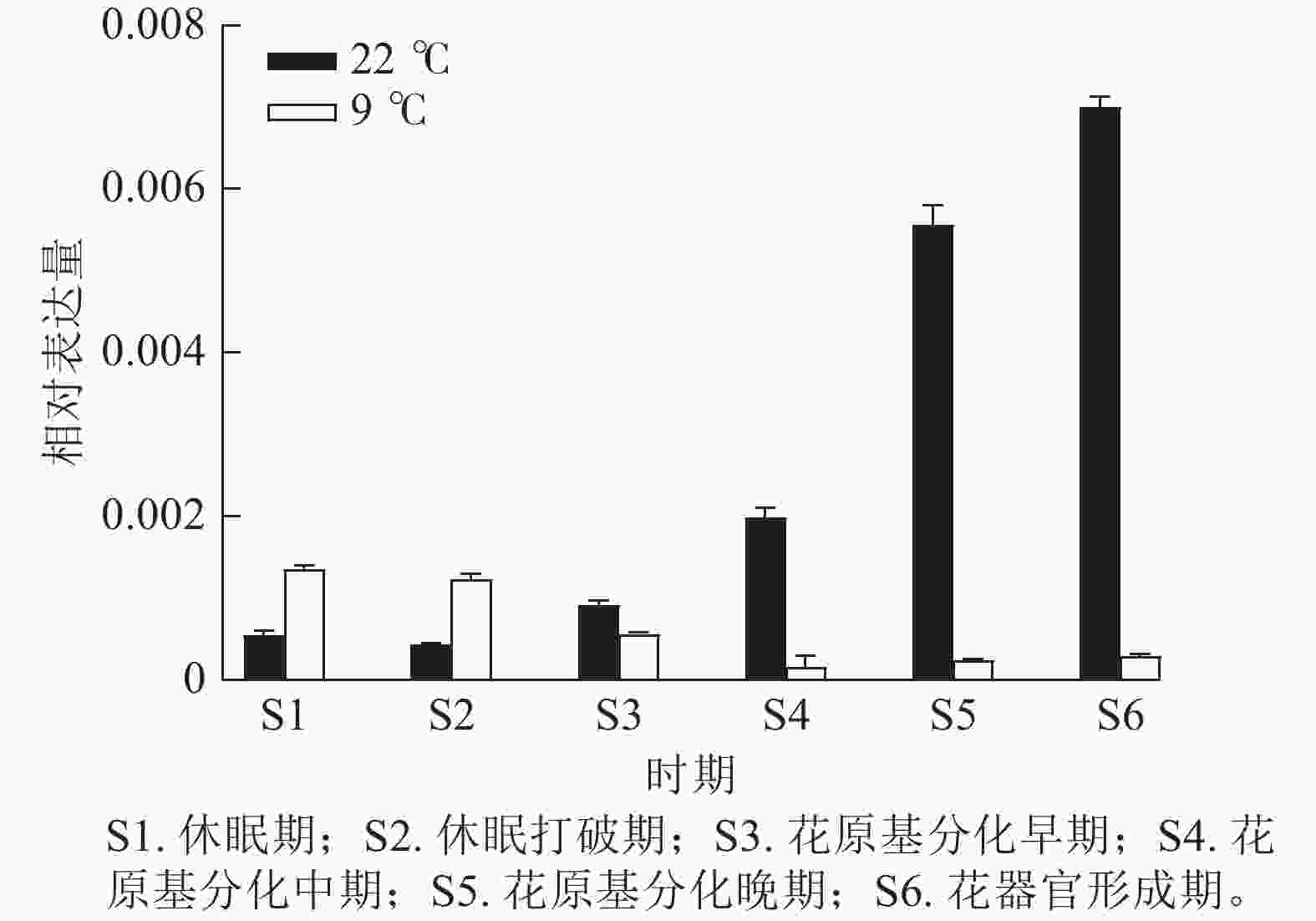
图 5 CsatFT2-2差异表达验证
Figure 5. Validation of differential expression of CsatFT2-2
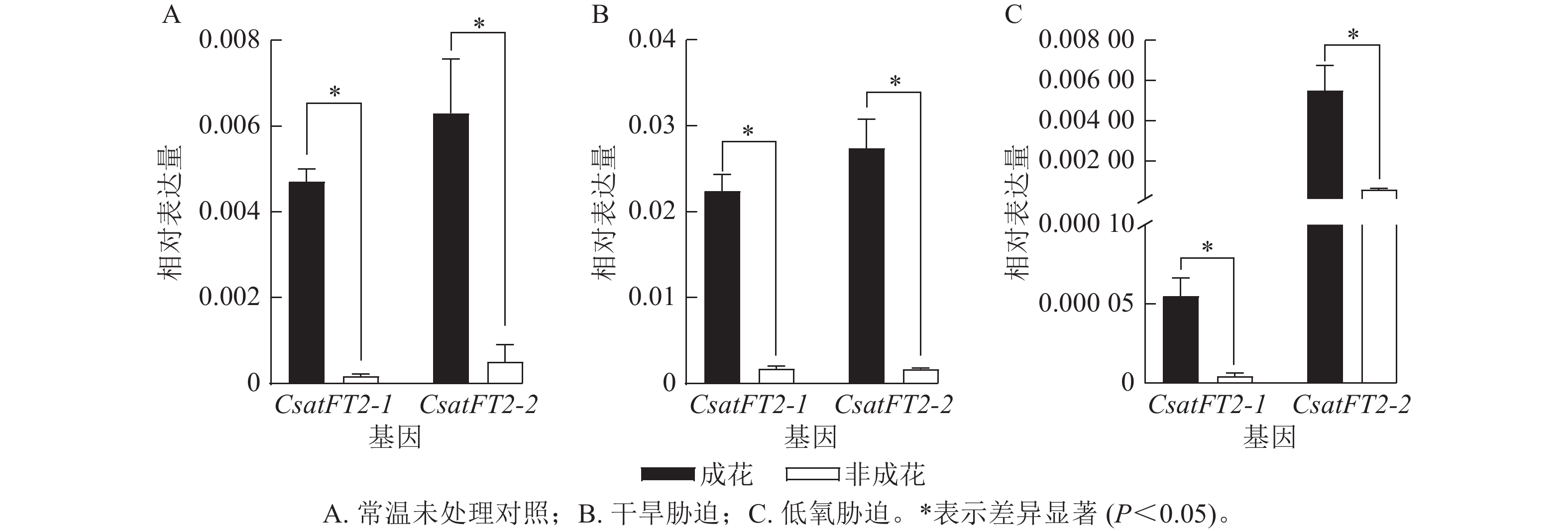
为了进一步验证CsatFT2-1与CsatFT2-2表达与成花能力的相关性,分析其在多种胁迫条件下(干旱、低氧)成花与非成花样本中的表达水平(图6)。结果表明:在成花顶芽中,CsatFT2-1和CsatFT2-2的表达量均显著高于对应的非成花(P<0.05)。在正常培养和干旱胁迫条件下的成花样本中,CsatFT2-2的表达量略高于CsatFT2-1;而在低氧胁迫成花样本中,CsatFT2-1的表达量显著低于CsatFT2-2 (P<0.05)。综上,推测CsatFT2-2在西红花开花过程中可能发挥更重要的调控作用。
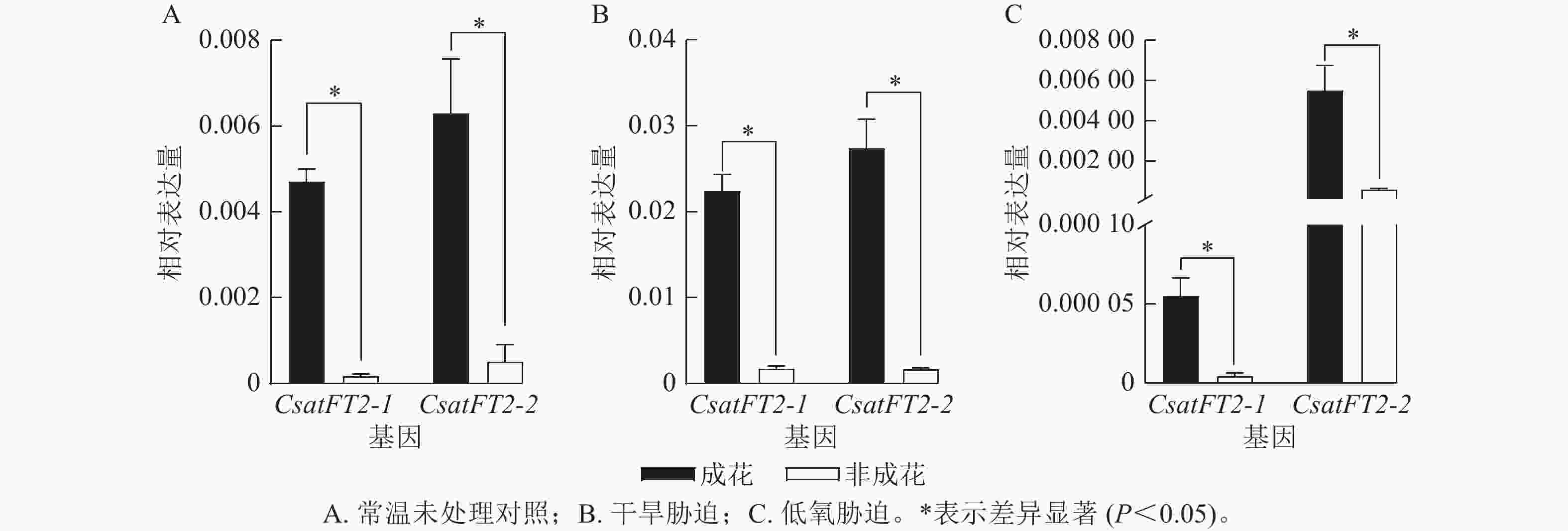
图 6 不同处理下CsatFT2-1和CsatFT2-2的表达分析
Figure 6. Expression analysis of CsatFT2-1 and CsatFT2-2 under different treatments
-
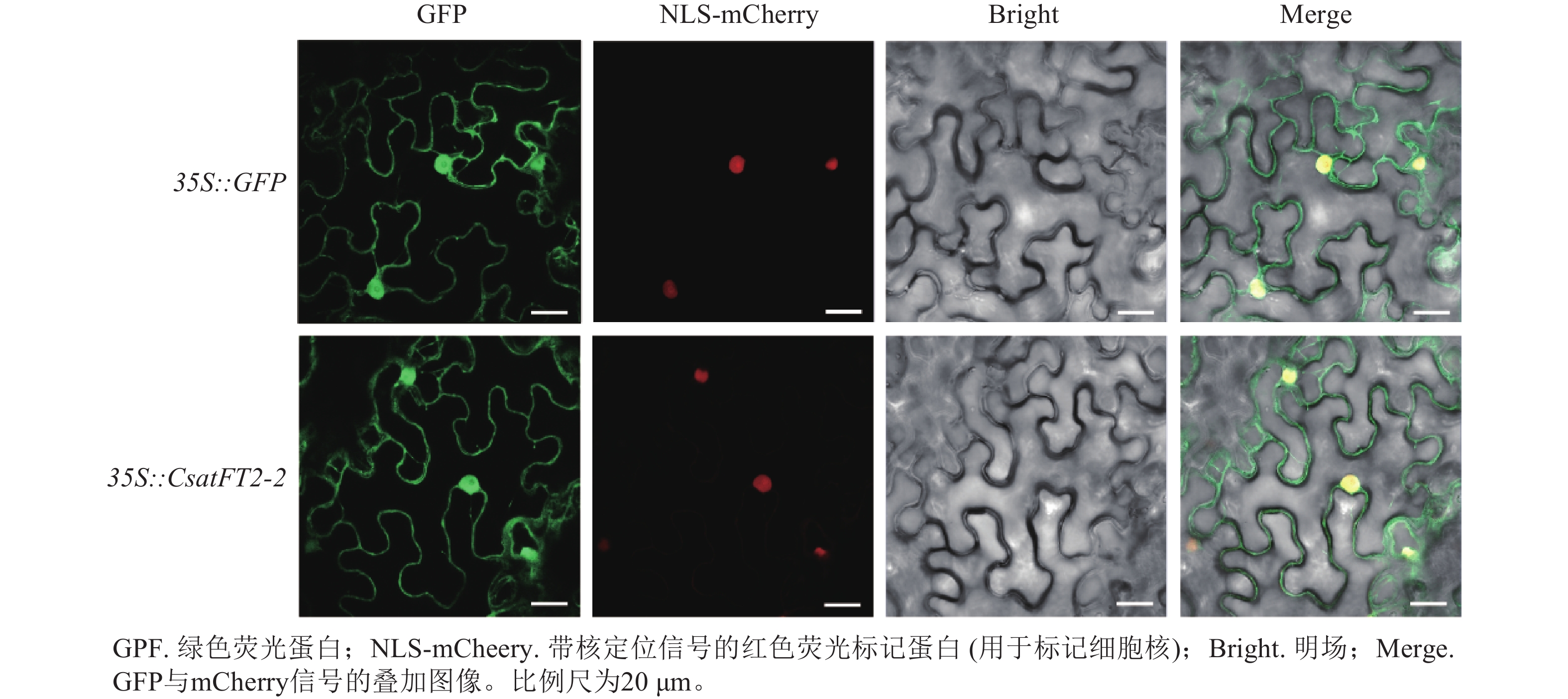
为探究CsatFT2-2蛋白在细胞内发挥功能的位点,将CsatFT2-2 -GFP融合蛋白在本氏烟草的叶片表皮细胞中进行瞬时表达。共聚焦显微镜结果显示:CsatFT2-2蛋白定位于细胞核和细胞周边区域,而空载体的GFP信号遍布整个细胞(图7),预示着CsatFT2-2蛋白可能通过在细胞核或膜系统的特异性分布,参与成花信号的感知与转导。
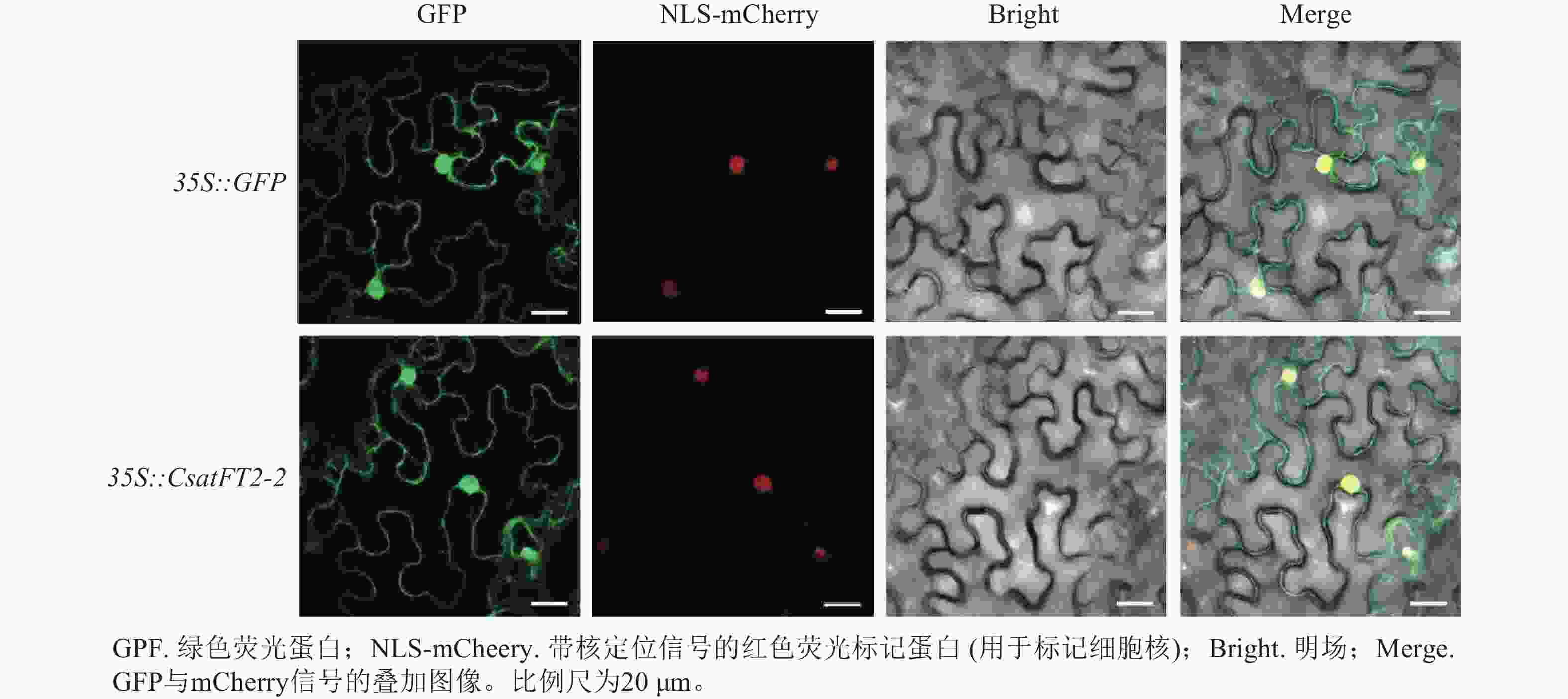
图 7 CsatFT2-2蛋白的亚细胞定位
Figure 7. Subcellular localization of CsatFT2-2 protein
-
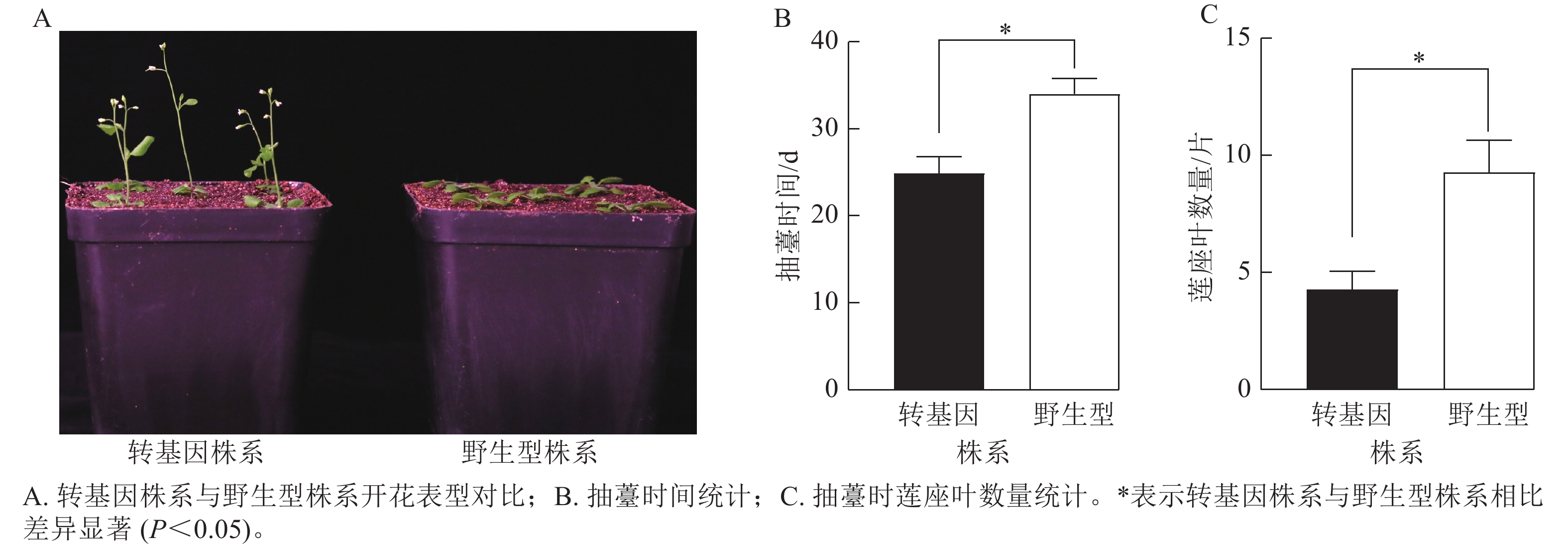
为了验证CsatFT2-2的生物学功能,将其在野生型拟南芥中过量表达。获得潮霉素抗性的T2代纯合株系后,观察其开花表型(图8A)。与野生型相比,过表达CsatFT2-2的转基因植株抽薹时间显著提前约9 d,且抽薹时的莲座叶数量显著减少约5片(图8B~C),表明CsatFT2-2具有促进开花的功能。
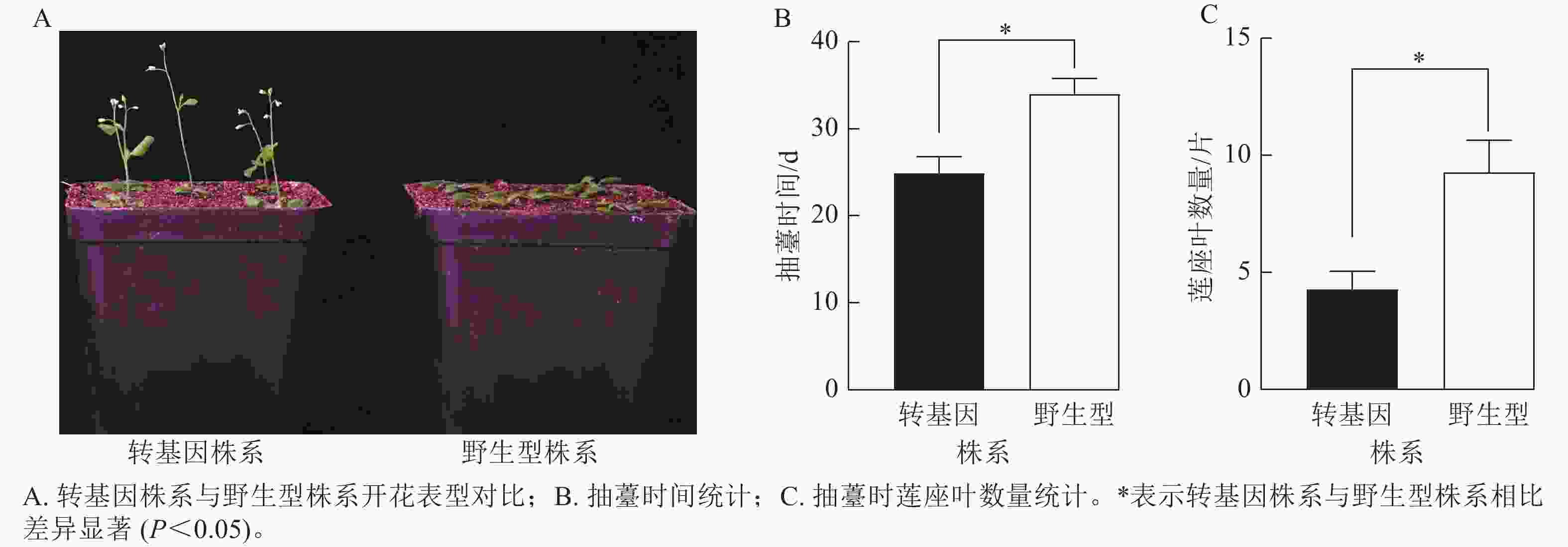
图 8 过表达CsatFT2-2对拟南芥开花时间的影响
Figure 8. Effect of CsatFT2-2 overexpression on flowering time in A. thaliana
-
目前关于西红花成花机制的研究集中于开花相关基因的分离与功能鉴定,如MADS-box家族、PEBP基因家族等[27−30]。这些基因在成花诱导及花器官发育中发挥重要作用,PEBP家族成员FT及其同源基因是保守的成花素信号,被认为在植物开花时间调控中具有关键功能。然而,对于FT及其同源基因在西红花开花数量方面的调控研究尚处于初步探索阶段[30]。
本研究基于西红花单倍型组装基因组对PEBP基因家族进行了系统鉴定。系统发育分析表明:西红花基因组中可能存在七类FT同源基因,而在花原基分化起始的关键时期,开花顶芽中仅有3类FT同源基因表达,这与FT类基因在植物开花诱导过程中具有功能分化和组织特异性表达的特点一致[11, 14−15]。通过比较花原基分化关键时期的转录组数据,并结合多种逆境胁迫条件下成花与非成花样本在顶芽和叶片中的表达差异分析,发现CsatFT2类的3个FT同源基因(CsatFT2、CsatFT2-1和CsatFT2-2)可能参与调控花原基分化过程,这与西红花及其他植物中FT同源基因参与开花调控的研究结果相一致[16−17]。序列分析表明:CsatFT2、CsatFT2-1和CsatFT2-2均具有典型FT蛋白特征,包括完整的PEBP结构域及保守的Tyr85和Gln140残基,这些残基已被证实对区分FT蛋白与TFL1蛋白至关重要[ 26]。表达分析结果显示:CsatFT2-2在花原基分化启动后表达量持续升高,而CsatFT2-1在成花顶芽花器官形成期的表达量明显低于CsatFT2-2;同时,在顶芽花器官形成期,CsatFT2-2的表达量约为CsatFT2的2倍,提示其在花原基分化过程中可能具有更为重要的调控作用。综上,CsatFT2-2可能在西红花花原基分化过程中发挥重要调控作用,这一结果与前人关于FT基因在开花调控中作为关键整合因子的研究结论一致[ 11]。
然而,由于西红花稳定、高效的遗传转化体系尚未建立,无法验证CsatFT2-2是否为调控西红花开花数量的重要基因。未来亟待突破西红花的遗传转化技术,编辑CsatFT2-2的基因序列,明确其在西红花本体中对花原基分化和开花数量的调控作用。另外2个关键的FT同源基因(CsatFT2、CsatFT2-1)可能具有促进开花的冗余功能,或在不同胁迫条件下发挥作用,仍需进一步研究。
-
本研究基于单倍型组装基因组数据对西红花FT同源基因进行了系统鉴定与命名。结合转录组分析和RT-qPCR结果,筛选出可能参与花原基分化的候选正调控基因CsatFT2-2,并通过异源表达对其功能进行了验证。结果表明:CsatFT2-2编码的蛋白具有典型的FT保守结构,在拟南芥中过表达CsatFT2-2可显著促进开花,表明其具有保守的促花功能;同时,CsatFT2-2表达水平与西红花成花过程呈显著相关性。本研究加深了对西红花成花分子机制的认识,为后续通过调控CsatFT2-2改良开花性状和提高产量提供了重要的候选基因资源。
本文已在中国知网网络首发,可在知网搜索、下载并阅读全文。
调控西红花花原基分化关键FT同源基因CsatFT2-2的鉴定与分析
DOI: 10.11833/j.issn.2095-0756.20250599
Identification and functional analysis of a key FT homolog, CsatFT2-2, regulating floral primordium differentiation in Crocus sativus
-
摘要:
目的 西红花Crocus sativus以干制花丝入药,经济价值高,但其复杂三倍体基因组导致开花整合基因FT同源基因尚未得到系统鉴定。解析西红花FT同源基因并筛选关键调控基因,对阐明其成花机制及分子育种具有重要意义。 方法 基于最新发布的单倍型组装基因组,挖掘并验证调控花原基分化的关键FT基因。通过生物信息学方法筛选含PEBP结构域的家族成员并构建系统进化树,明确其亚类;结合花原基分化关键阶段转录组数据,筛选得到5个与分化进程相关的FT-like基因。 结果 CsatFT2-1和CsatFT2-2在常温成花顶芽中自花原基分化早期起持续上调,且明显高于低温非成花顶芽;两者在非成花样本中表达极低,而在成花过程中逐步升高。相比之下,CsatFT3-1、CsatFT3-2和CsatFT6-1虽呈上调趋势,但差异不显著,其中CsatFT6-1整体表达水平较低。综合分析表明:CsatFT2类基因可能参与成花启动,其中CsatFT2-2在成花顶芽中呈特异性高表达,提示其在花原基分化及成花诱导中具有重要作用。该基因归属于开花促进型分支,具有完整保守的PEBP结构域。实时荧光定量PCR (RT-qPCR)结果显示:CsatFT2-2在常温诱导成花过程中表达量上调约20倍,而在低温、干旱及低氧等非成花条件下显著下调。亚细胞定位结果表明:CsatFT2-2蛋白定位于细胞核和细胞膜;在拟南芥Arabidopsis thaliana中异源过表达可使抽薹提前约9 d,莲座叶减少约54%,进一步验证其促花功能。 结论 明确FT同源基因参与西红花成花调控,并筛选到西红花花原基分化的关键正调控基因CsatFT2-2,为西红花成花机制解析及分子育种提供重要靶点。图8表2参30 Abstract:Objective Crocus sativus is an important medicinal plant whose dried stigmas possess high economic value. However, due to its complex triploid genome, FT homologs have not been systematically characterized. This study aims to identify key FT homologs involved in floral primordium differentiation, thereby providing candidate genes for elucidating flowering mechanisms and molecular breeding in saffron. Method Based on the recently published haplotype-resolved genome, FT genes associated with floral primordium differentiation were identified and functionally analyzed. Members of the PEBP gene family were identified through bioinformatics analysis, and a phylogenetic tree was constructed for subfamily classification. Transcriptome data from key stages of floral primordium differentiation were integrated to identify 5 FT-like genes potentially involved in this process. Result CsatFT2-1 and CsatFT2-2 were continuously upregulated from the early stages of floral primordium differentiation in flowering apical buds under normal temperature conditions and exhibited significantly higher expression levels than those in non-flowering buds under low-temperature conditions. Their expression levels were extremely low or undetectable in non-flowering samples but gradually increased during floral development. In contrast, CsatFT3-1, CsatFT3-2, and CsatFT6-1 showed increasing expression trends, but no obvious differences were observed between flowering and non-flowering samples, and CsatFT6-1 exhibited overall low expression levels. These results suggested that CsatFT2 genes may participate in floral initiation. Among them, CsatFT2-2 showed specifically high expression in flowering apical buds, indicating an important role in floral primordium differentiation and flowering induction. CsatFT2-2 belongs to the flowering-promotive clade and contains a conserved PEBP domain. RT-qPCR analysis showed that the expression level of CsatFT2-2 increased by approximately 20-fold during flowering induction under normal temperature conditions but was significantly downregulated under low temperature, drought, and hypoxic stresses. Subcellular localization analysis reveals that the CsatFT2-2 protein was localized in the nucleus and plasma membrane. Heterologous overexpression of CsatFT2-2 in Arabidopsis thaliana resulted in bolting approximately 9 days earlier and reduced rosette leaf number by approximately 54%, further confirming its flowering-promoting function. Conclusion This study systematically reveals the roles of FT homologs in flowering regulation in C. sativus and identified CsatFT2-2 as a key positive regulatory gene involved in floral primordium differentiation, providing an important molecular target for elucidating flowering mechanisms and molecular breeding in saffron. [Ch, 8 fig. 2 tab. 30 ref.] -
Key words:
- Crocus sativus /
- FT-like genes /
- floral primordium differentiation /
- gene function
-
图 1 西红花花原基分化连续过程的形态特征
Figure 1 Morphological characteristics of the successive stages of floral primordium differentiation in C. sativus
图 2 FT-like基因系统进化与序列比对分析
Figure 2 Phylogenetic and sequence alignment analyses of FT-like genes
图 4 FT-like基因在顶芽和叶片组织中的表达模式分析
Figure 4 Expression patterns of FT-like genes in apical buds and leaves
图 6 不同处理下CsatFT2-1和CsatFT2-2的表达分析
Figure 6 Expression analysis of CsatFT2-1 and CsatFT2-2 under different treatments
图 8 过表达CsatFT2-2对拟南芥开花时间的影响
Figure 8 Effect of CsatFT2-2 overexpression on flowering time in A. thaliana
表 1 引物序列
Table 1. Primer sequences used in this study
引物名称 正向引物(5′→3′) 反向引物(5′→3′) CsatFT1q ATGAGTAGGGATTCGCTGGTGCTCG AACCTCCCGAGACTGGTACGTCACC CsatFT2q TGGCTAAACAAACGAGCTACGACTA CAATCACTCTACCAACAACCAAAGG CsatFT3q ATATGGCCGCAGCTACTAGAGAAAC TCCACCATCACCAGAGTGTAGAAAT CsatFT2-1q CCGAGGGTCGAGGTCGGAG GCGCATATACCGTTTGCCGTC CsatFT2-2q CCACCGGTTTGTCTTCATGC CCGACTCCCTCTGACAGTTGA CsatFT2-2CDS扩增 ATGGTTAGGGAGAGGAATCCTTTGG GCTCCGGTGGGAGAAGGATTTAA CsatFT2-2过表达载体构建 ACACGGGGGACTCTTGACATGGTTA
GGGAGAGGAATCCTCCTTTACTAGTCAGATCTACTTAA
ATCCTTCTCCCACCGGAGCsatFT2-2亚细胞定位 ACACGGGGGACTCTTGACATGGTTA
GGGAGAGGAATCCTCCTTTACTAGTCAGATCTACAATC
CTTCTCCCACCGGAGC 下载: 导出CSV
下载: 导出CSV
-
[1] 高广春, 肖晨, 李军, 等. 西红花AP2/ERF基因家族鉴定及其响应激素诱导柱头脱辅基类胡萝卜素类成分生物合成的分析[J]. 中草药, 2025, 56(22): 8322−8333. GAO Guangchun, XIAO Chen, LI Jun, et al. Identification of AP2/ERF gene family in Crocus sativus and analysis of de-prothetic group apocarotenoid biosynthesis in stigmas induced by its responsive hormone[J]. Chinese Traditional and Herbal Drugs, 2025, 56(22): 8322−8333. DOI: 10.7501/j.issn.0253-2670.2025.22.022. [2] 王婷婷, 马琴国. 名贵的西域“贡品”−藏红花[J]. 中医健康养生, 2024, 10(7): 25−27. WANG Tingting, MA Qinguo. A precious tribute of the western regions-saffron[J]. TCM Healthy Life-Nurturing, 2024, 10(7): 25−27. [3] 杜雪, 李秀娟, 桂思琦, 等. 西红花真菌性病害与生防菌鉴定研究进展[J]. 浙江农林大学学报, 2021, 38(6): 1279−1288. DU Xue, LI Xiujuan, GUI Siqi, et al. Research progress of fungal diseases in Crocus sativus and identification of biocontrol bacteria[J]. Journal of Zhejiang A&F University, 2021, 38(6): 1279−1288. DOI: 10.11833/j.issn.2095-0756.20200809. [4] 兰卫, 朱金芳, 耿直. 新疆西红花的种植技术[J]. 新疆医科大学学报, 2010, 33(8): 874−875. LAN Wei, ZHU Jinfang, GENG Zhi. Planting techniques of Crocus sativus in Xinjiang[J]. Journal of Xinjiang Medical University, 2010, 33(8): 874−875. [5] 周琳, 邢鲁亭, 吕伟德, 等. 不同种源西红花倍性·生长性状及开花性状比较分析[J]. 安徽农业科学, 2025, 53(8): 158−161. ZHOU Lin, XING Luting, LÜ Weide, et al. Comparative analysis of ploidy, growth traits and flowering traits of Crocus sativus from different provenances[J]. Journal of Anhui Agricultural Sciences, 2025, 53(8): 158−161. DOI: 10.3969/j.issn.0517-6611.2025.08.034. [6] 吴光炎, 孙莉琼, 王康才, 等. 变温处理对西红花花芽分化及其生理生化的影响[J]. 西北植物学报, 2021, 41(8): 1338−1346. WU Guangyan, SUN Liqiong, WANG Kangcai, et al. Effects of variable temperature treatments on flower bud differentiation of Crocus sativus L.[J]. Acta Botanica Boreali-Occidentalia Sinica, 2021, 41(8): 1338−1346. DOI: 10.7606/j.issn.1000-4025.2021.08.1338. [7] 张雪梅, 苏晓兰, 迟敬楠, 等. 园艺植物成花调控途径及分子机制研究进展[J]. 植物生理学报, 2025, 61(8): 1086−1096. ZHANG Xuemei, SU Xiaolan, CHI Jingnan, et al. Research progress on flowering regulation pathways and molecular mechanisms in horticultural plants[J]. Chinese Bulletin of Plant Physiology, 2025, 61(8): 1086−1096. DOI: 10.13592/j.cnki.ppj.300152. [8] FORNARA F, de MONTAIGU A, COUPLAND G. SnapShot: control of flowering in Arabidopsis[J]. Cell, 2010, 141(3): 550−550. e2. DOI: 10.1016/j.cell.2010.04.024. [9] 肖和志. 香蕉MaSVP-L3基因克隆及互作蛋白鉴定[J]. 现代园艺, 2025, 48(9): 60−65. XIAO Hezhi. Cloning of banana MaSVP-L3 gene and identification of its interacting protein[J]. Contemporary Horticulture, 2025, 48(9): 60−65. DOI: 10.14051/j.cnki.xdyy.2025.09.001. [10] PENG Qiang, ZHU Chunmei, LIU Tao, et al. Phosphorylation of OsFD1 by OsCIPK3 promotes the formation of RFT1-containing florigen activation complex for long-day flowering in rice[J]. Molecular Plant, 2021, 14(7): 1135−1148. DOI: 10.1016/j.molp.2021.04.003. [11] 宋春晖, 李晓朋, 杨梓健, 等. 成花素基因FT调控植物开花和发育的研究进展[J]. 中国科学: 生命科学, 2025, 55(7): 1332−1346. SONG Chunhui, LI Xiaopeng, YANG Zijian, et al. Research progress on the regulation by the FLOWRING LOCUS T(FT) gene in plant flowering and development[J]. Scientia Sinica (Vitae), 2025, 55(7): 1332−1346. DOI: 10.1360/SSV-2024-0279. [12] PASRIGA R, YOON J, CHO L H, et al. Overexpression of RICE FLOWERING LOCUS T 1 (RFT1) induces extremely early flowering in rice[J]. Molecules and Cells, 2019, 42(5): 406−417. DOI: 10.14348/molcells.2019.0009. [13] 闫小玲, 郝琴, 申孜, 等. 毛竹PheFT1基因的表达、蛋白互作及生物学功能分析[J]. 林业科学, 2025, 61(4): 140−152. YAN Xiaoling, HAO Qin, SHEN Zi, et al. Expression, protein interaction and biological function analysis of PheFT1 gene in Moso bamboo[J]. Scientia Silvae Sinicae, 2025, 61(4): 140−152. DOI: 10.11707/j.1001-7488.LYKX20240069. [14] LIU Wei, JIANG Bingjun, MA Liming, et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation[J]. New Phytologist, 2018, 217(3): 1335−1345. DOI: 10.1111/nph.14884. [15] WU Lan, LI Fei, DENG Qiaohong, et al. Identification and characterization of the FLOWERING LOCUS T/TERMINAL FLOWER 1 gene family in Petunia[J]. DNA and Cell Biology, 2019, 38(9): 982−995. DOI: 10.1089/dna.2019.4720. [16] KALIA D, JOSE-SANTHI J, KUMAR R, et al. Analysis of PEBP genes in saffron identifies a FLOWERING LOCUS T homologue involved in flowering regulation[J]. Journal of Plant Growth Regulation, 2023, 42(4): 2486−2505. DOI: 10.1007/s00344-022-10721-2. [17] 王桢, 杨柳燕, 裴卫忠, 等. 西红花FT同源基因的表达及功能分析[J]. 植物研究, 2022, 42(2): 224−233. WANG Zhen, YANG Liuyan, PEI Weizhong, et al. Expression and functional analysis of FT homologous genes in saffron (Crocus sativus L.)[J]. Bulletin of Botanical Research, 2022, 42(2): 224−233. DOI: 10.7525/j.issn.1673-5102.2022.02.007. [18] XU Zhichao, CHEN Shanshan, WANG Yalin, et al. Crocus genome reveals the evolutionary origin of crocin biosynthesis[J]. Acta Pharmaceutica Sinica B, 2024, 14(4): 1878−1891. DOI: 10.1016/j.apsb.2023.12.013. [19] XI Xiaoyuan, LI Jing, SONG Jia, et al. CsERECTA alternative splicing regulates the flowering numbers depending on temperature in Crocus sativus L.[J]. Industrial Crops and Products, 2024, 218: 118971. DOI: 10.1016/j.indcrop.2024.118971. [20] 王树和, 周彩勤, 张奎望, 等. 金叶女贞棒孢叶斑病菌的生物学特性、致病性及系统发育[J]. 浙江农林大学学报, 2019, 36(6): 1174−1181. WANG Shuhe, ZHOU Caiqin, ZHANG Kuiwang, et al. Characterization of Corynespora cassiicola from golden vicary privet based on phylogeny, biology, and pathogenicity[J]. Journal of Zhejiang A&F University, 2019, 36(6): 1174−1181. DOI: 10.11833/j.issn.2095-0756.2019.06.015. [21] 郝力慧, 董彬, 朱绍华, 等. 牡丹响应高温胁迫的转录组分析及PsHSP基因表达[J]. 浙江农林大学学报, 2021, 38(4): 802−811. HAO Lihui, DONG Bin, ZHU Shaohua, et al. Transcriptome analysis and PsHSP gene expression of Paeonia suffruticosa in response to high temperature stress[J]. Journal of Zhejiang A&F University, 2021, 38(4): 802−811. DOI: 10.11833/j.issn.2095-0756.20200529. [22] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method[J]. Methods, 2001, 25(4): 402−408. DOI: 10.1006/meth.2001.1262. [23] 龚涵, 王林凯, 刘艳菊, 等. 热带地区3种油茶叶片解剖及光合生理特性比较[J]. 西南林业大学学报(自然科学), 2025, 45(11): 45−51. GONG Han, WANG Linkai, LIU Yanju, et al. Comparative study on leaf anatomy and photosynthetic characteristics of 3 tropical tea-oil Camellia[J]. Journal of Southwest Forestry University (Natural Sciences), 2025, 45(11): 45−51. DOI: 10.11929/j.swfu.202411032. [24] 申序, 牟东岭, 周金忠, 等. 龙眼DlSWC5基因克隆、亚细胞定位及表达特性分析[J]. 南方农业学报, 2024, 55(7): 2137−2147. SHEN Xu, MOU Dongling, ZHOU Jinzhong, et al. Cloning, subcellular localization and expression characterization of DlSWC5 gene in longan (Dimocarpus longan Lour.)[J]. Journal of Southern Agriculture, 2024, 55(7): 2137−2147. DOI: 10.3969/j.issn.2095-1191.2024.07.025. [25] 李超, 张彦楠, 刘焕龙, 等. 亚洲棉和雷蒙德氏棉PEBP家族基因的鉴定及该家族基因在陆地棉组织中表达分析[J]. 作物学报, 2015, 41(3): 394−404. LI Chao, ZHANG Yannan, LIU Huanlong, et al. Identification of PEBP gene family in Gossypium arboreum and Gossypium raimondii and expression analysis of the gene family in Gossypium hirsutum[J]. Acta Agronomica Sinica, 2015, 41(3): 394−404. DOI: 10.3724/SP.J.1006.2015.00394. [26] 赵田芸, 杨雄, 杨晓宇, 等. 毛白杨PtTFL1.1和PtTFL1.2基因的克隆和表达模式分析[J]. 中国细胞生物学学报, 2020, 42(6): 1008−1017. ZHAO Tianyun, YANG Xiong, YANG Xiaoyu, et al. Cloning and expression patterns of PtTFL1.1 and PtTFL1.2 in Populus tomentosa[J]. Chinese Journal of Cell Biology, 2020, 42(6): 1008−1017. DOI: 10.11844/cjcb.2020.06.0008. [27] TSAFTARIS A S, PASENTSIS K, POLIDOROS A N. Isolation of a differentially spliced C-type flower specific AG-like MADS-box gene from Crocus sativus and characterization of its expression[J]. Biologia Plantarum, 2005, 49(4): 499−504. DOI: 10.1007/s10535-005-0041-6. [28] HAGHIGHI R, SAYED TABATABAEI B E, ALI MOHAMMAD MIRMOHAMMADI MAIBODY S, et al. A flowering inhibitor of the temperature-dependent pathway in Crocus sativus L.[J]. Molecular Biology Reports, 2020, 47(3): 2171−2179. DOI: 10.1007/s11033-020-05316-7. [29] TSAFTARIS A, PASENTSIS K, KALIVAS A, et al. Isolation of a CENTRORADIALIS/TERMINAL FLOWER1 homolog in saffron (Crocus sativus L.): characterization and expression analysis[J]. Molecular Biology Reports, 2012, 39(8): 7899−7910. DOI: 10.1007/s11033-012-1634-8. [30] 周琳, 滕一涤, 吴立峰, 等. 西红花花芽分化及其花期调控研究进展[J]. 江苏农业科学, 2024, 52(14): 23−30. ZHOU Lin, TENG Yidi, WU Lifeng, et al. Research progress on flower bud differentiation and flowering period regulation of Crocus sativus[J]. Jiangsu Agricultural Sciences, 2024, 52(14): 23−30. DOI: 10.15889/j.issn.1002-1302.2024.14.004. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250599
 点击查看大图
点击查看大图
计量
- 文章访问数: 87
- HTML全文浏览量: 34
- PDF下载量: 22
- 被引次数: 0
