







-
草本植物是森林生态系统的重要组成部分,在维持森林生态系统稳定性方面发挥着重要的作用。虽然草本层生物量仅占整个森林生物量的1%左右,却包括林内90%甚至更多的物种,很大程度上影响森林生态系统的能量流动和物质循环[1-2]。自20世纪80年代以来,中国学者针对长白山阔叶红松Pinus koraiensis林生态系统结构、生物多样性评估和变化动态等领域开展了大量研究。基于草本植物的重要地位,林下草本层也成为了生态学家的关注热点。郝占庆等[3]、夏福才等[4]、赵秀海等[5]、于凤英等[6]最早对草本植物群落特征做了详细的研究,之后李步杭等[7]、常新华等[8]又进一步研究了阔叶红林草本植物的生长期动态及空间分布格局,但这些研究主要是针对原始阔叶红松林草本植物,其目的在于了解原始林的林下层植物资源,而处于不同演替阶段的天然阔叶红松林草本植物的物种多样性研究少见报道。近几十年来,由于人为不合理利用森林资源,在中国东北部地区大面积的原始林被各演替阶段的次生林取代。作为对上层林木空间结构变化最为敏感的草本植物是森林生态系统物种多样性评价不可缺少的一部分,其多样性的研究为森林抚育管理提供有利的理论依据。本研究区林下草本植物大都属于浅根性植物,对土壤环境的变化有着较强的敏感性。由于该区域的森林植物群落是长期自然恢复的结果,林下土壤发育较好,但目前少有专门针对这种森林群落下草本植物与土壤因子的相关性研究,大部分研究主要集中在人工林,如崔宁洁等[9]对不同林龄马尾松Pinus massoniana人工林林下植被多样性与土壤理化性质的研究;杨东等[10]对海堤防护林草本物种多样性与土壤肥力的研究等。土壤作为植物生长的重要物质基础,是森林生态系统中物质和能量交换的重要参与者和载体,为植物生长提供所必须的矿质营养和水分,土壤条件的差异会直接影响着植物的生长发育,从而会影响物种多样性[11]。因此,研究森林群落演替过程中物种多样性与土壤的相关性具有重要的意义。本研究区植被类型为天然阔叶红松林,通过对林内草本植物的调查分析,以及物种多样性与土壤因子的相关性分析,探讨草本植物物种多样性的生长期动态及其在不同土壤条件下的动态特征,旨在了解天然阔叶红松林草本植物资源和群落的生态特征,以期为森林抚育管理、多样性保护和持续利用提供参考,为加快森林生态系统的进一步发育演替以及合理开发资源提供基础数据和理论依据。
HTML
-
研究地点位于吉林省蛟河林业实验区管理局辖区。2009年,在该研究区域建立21.12 hm2(320 m × 640 m)固定监测样地(43°57.897′~43°58.263′N,127°42.789′~127°43.310′E),海拔高度为425.3~525.8 m,属于长白山系张广才岭山脉。研究样地内地势较平坦,部分区域崎岖,属于典型的温带大陆性山地气候,年平均气温为3.8 ℃,冬夏温差较大,最热月份7月平均气温达21.7 ℃,最冷月份1月平均气温为-18.6 ℃。平均年降水量695.9 mm,主要集中在7-8月。土壤为山地暗棕色森林土,土层厚度较薄,为20~50 cm。该区原始植被为阔叶红松林,经采伐破坏后形成的天然林,林龄为近熟林。木本植被种类有63种(包括3种藤本植物),主林层乔木树种主要有红松,蒙古栎Quercus mongolica,水曲柳Fraxinus mandshurica,紫椴Tilia amurensis,胡桃揪Juglans mandshurica,春榆Ulmus japonica,色木槭Acer mono和千金榆Carpinus cordata等树种。
-
研究样地内,通过全站仪测量将整个样地划分为20 m × 20 m正方形连续样方,样方的顶点均埋有石桩标记。采用机械布点法均匀布设了272个1 m × 1 m的草本小样方。于2014年5-9月的月初调查,记录草本植物种名、盖度(长轴、短轴的长度)、株数、高度等参数。调查的土壤因子包括土壤速效氮、速效磷、速效钾、全氮、全磷、全钾、有机质和pH值。具体调查方法如下:在各草本小样方的东侧设置1个土壤采样点,在采样点的表层(0~15 cm)各取1个500 g的土壤样品,装入写有编号的自封袋中带回实验室测定。
室内土壤样品分析方法依据《森林土壤分析方法》的操作步骤进行:采用重铬酸钾外加热法测定有机质;用凯氏定氮法测定土壤全氮,碱解-扩散法测定速效氮;用酸溶钼锑抗比色法测定土壤全磷,用浸提-钼锑抗比色法测定有效磷;用浓硫酸-高氯酸加热法测定土壤全钾,分光光度计法测定速效钾;用pH计测定土壤pH值。
-
物种多样性采用Gleason指数(G),Simpson指数(D)和Shannon-Wiener指数(H)表征,采用Pielou指数(E)和生态优势度(λ)来描述样地内草本植物物种分布均匀程度和优势种分布状况。Gleason指数(G):G=S/lnA; Simpson指数(D):D=1-∑Pi2;Shannon-Wiener指数(H):H=-∑PilnPi;Pielou指数(E):E=D/lnS;生态优势度指数(λ):λ= ∑[ni(ni-1)]/[N(N-1)];重要值(Pi):Pi=(相对密度+相对盖度+相对频度)/3。其中: Pi是种i的相对重要值;S是样地内物种数;A为样方面积; N为样地内所有草本植物的总株数;ni是样地内物种i的株数[12-13]。
采用Jaccard指数计算月份间的物种相似性系数:J=c/(a+b-c)。其中:a为月份a的物种数;b为月份b的物种数;c为a和b共有的物种数。
利用Excel软件初步处理野外调查的数据、土壤理化性质,计算物种多样性指数,利用CANOCO 4.5软件的冗余分析方法(redundancy analysis,RDA)来研究物种多样性和土壤因子的对应关系。
2.1. 样方布设及数据采集
2.2. 数据分析方法
-
从表 1可以看出:在研究样地内共调查到草本植物91种,隶属于34科80属,其中蕨类植物有7种,隶属于4科6属。以毛茛科Ranunculaceae,百合科Liliaceae,伞形科Umbelliferae和菊科Compositae等世界性大科的物种数最多,分别含有12,10,8和7种。根据统计分析结果表示,样地内含10个种以上的科只有2个(毛茛科和百合科),所含的属数为10属,且大部分种为早春植物,如:山韭Allium senescens,朝鲜顶冰花Gagea lutea,黑水银莲花Anemone amurensis,多被银莲花Anemone raddeana,菟葵Eranthis stellata,侧金盏Adonis amurensis和东北扁果草Isopyrum manshuricum等。其他科属有90%以上只含有1~3个物种,其中只含1个种的科有16个,占总科数的47.1%;有74个属均只含有1个物种,占总属数的92.5%。由此可见,样地内草本植物少种科、少种属现象很明显,而单种属在样地内占有绝对优势。
科 属 种 毛茛科 Ranunculaceae 10 12 百合科 Liliaceae 10 10 伞形科 Umbelliferae 7 7 菊科 Compositae 5 6 蔷薇科 Rosaceae 4 4 罂粟科 Papaveraceae 3 5 唇形科 Labiatae 3 3 莎草蕨科 Cyperaceae 2 5 蹄盖科 Athyriaceae 2 3 石竹科 Caryophyllaceae 2 3 荨麻科 Urticaceae 2 3 小檗科 Berberidaceae 2 2 合计 52 63 鳞毛蕨科 Dryopteridaceae 2 2 禾本科 Gramineae 2 2 豆科 Leguminosae 2 2 兰科 Orchidaceae 2 2 茜草科 Rubiaceae 2 2 虎耳草科 Saxifragaceae 2 2 铁线蕨科 Adiantaceae 1 1 凤仙花科 Balsaminaceae 1 1 紫草科 Boraginaceae 1 1 桔梗科 Campanulaceae 1 1 金粟兰科 Chloranthaceae 1 1 十字花科 Cruciferae 1 1 合计 18 18 木贼科 Equisetaceae 1 1 牻牛儿苗科 Geraniaceae 1 1 金丝桃科 Hypericaceae 1 1 柳叶菜科 Onagraceae 1 1 紫萁蕨科 Osmundaceae 1 1 酢浆草科 Oxalidaceae 1 1 芍药科 Paeoniaceae 1 1 透骨草科 Phrymaceae 1 1 花芌科 Polemoniaceae 1 1 堇菜科 Violaceae 1 1 合计 10 10 Table 1. Species composition of herbs in plots
-
由于不同生长季节的气候条件和上层乔木群落结构的不同,草本植物在生长季内的生长动态会存在一定的差异。据调查,5-9月的草本植物物种数分别为71种、77种、74种、72种和60种。由表 2可以看出,物种的组成存在着明显的差异,5月与其他月份的差异最为显著,与其他月份的草本植物物种相似度最低,尤其是与9月的相似度只有0.489;夏季(6月、7月、8月)草本植物的相似度最大,6月和7月的相似度高达0.936。5月初,积雪刚刚融化,生活周期短暂的早春草本植物大量萌发,主要有黑水银莲花,东北延胡索Corydalis yanhusuo和东北扁果草Isopyrum manshuricum,大量的早夏植物也相继萌发,如荷青花Hylomecon japonicum,美汉草Meehania urticifolia和北重楼Paris verticillata,藜芦Veratrum nigrum等。6月,虽有部分早春植物死亡,但早夏植物已全部萌发,晚夏植物轮叶百合Lilium distichum和珠芽艾麻Laportea bulbifera等也相继萌发,因此6月草本植物物种最为丰富。从7月到9月,早夏和晚夏植物逐渐枯萎,物种数逐渐减少,物种的组成也发生了较大的变化,9月的物种组成与其他月份也相差较大。
月份 5 6 7 8 6 0.663 7 0.648 0.936 8 0.607 0.863 0.92 9 0.489 0.7 3 0.740 0.76 Table 2. Comparability index of herbs from May to September
与物种数不同,草本植物的个体总数从5月到9月急剧减少,各月份的个体数量差异显著。Gleason指数、Simpson指数和Shannon-Wiener指数的变化呈单峰型分布(表 3),在6月达到最大值,其值分别13.726 8,0.923 9和3.093 3,其次是7月和5月。9月取得最小值,分别为10.790 0,0.847 8和2.554 2。Pielou指数也具有相似的变化趋势,先增加后减小,但7月达到最大,9月依然取得最小值;生态优势度呈现逐月递增的趋势,9月最高,5月最小,仅有0.099 5,明显低于其他月份,表明5月的草本植物优势种的地位不突出。均匀度指数反映的是物种个体数目分配的均匀程度;生态优势度指数反映了各物种种群数量的变化情况,生态优势指数越大,说明群落内物种数量分布越不均匀,优势种的地位越突出[14]。
月 份 Gleason 指数 Simpson 指数 Shannon-矶iener 指数 Pielou 指数 生态优势度指数 物种数 属数 科数 个体总数 5 2.657 2 0.9 5 2.924 8 0.686 0.099 5 7 6 25 4 333 6 3.726 8 0.923 9 3.092 9 0.700 5 0. 24 6 77 70 34 23 2 3 7 3. 92 0 0.92 5 3.043 3 0.7 8 7 0. 36 7 74 67 33 9 0 6 8 2.860 8 0.897 2 2.890 3 0.675 8 0. 73 2 72 65 32 3 423 9 0.790 0 0.847 8 2.554 2 0.623 8 0.234 7 60 53 28 658 Table 3. Changes dynamic of species diversity of herbs from May to September
-
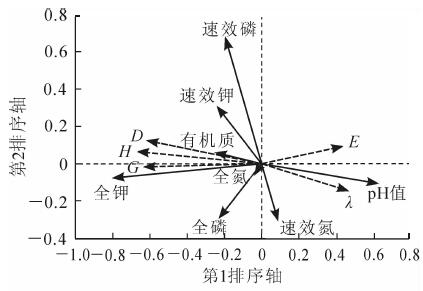
对草本植物多样性的5个指数与8个土壤因子进行冗余分析排序。分析结果显示:土壤因子能够解释39.2%的草本植物多样性变化。冗余分析中前2个排序轴的物种与环境相关性较高,共解释物种和环境关系总方差的99.3%,排序结果可信。在草本多样性与土壤冗余分析中(表 4),第1排序轴主要反映了土壤全钾和pH值的变化趋势,这2个因子与第1排序轴的相关系数分别为-0.509 9和0.391 3;第2排序轴主要反映的了土壤速效磷的变化趋势,其与排序轴的相关系数为0.141 4,说明影响草本植物多样性变化的主要土壤因子有土壤全钾、速效磷和pH值。由于并非每个土壤因子都与物种多样性的相关关系都达到显著水平,应用前向选择和蒙特卡洛检验分析(Monte Carl permutation test)每个土壤因子对物种多样性的显著性关系。结果表明:土壤全钾和pH值分别对物种多样性变化有着显著性的解释作用(P=0.002,F=26.089;P=0.002,F=31.382),两者对物种多样性变化的解释量占所有土壤因子解释量的66.0%。
土壤参数 第1 排序轴 第2 排序轴 有机质 一0.159 2 0.012 1 全氮 一0.025 7 一0.011 5 速效氮 0.053 0 一0.062 0 速效磷 一0.125 6 0.141 4 全磷 一0.145 6 一0.060 0 速效钾 一0.152 9 0.062 7 全钾 一0.509 9 一0.015 9 pH 值 0.391 3 一0.021 3 Table 4. Correlations between soil factors and the ordina tion axis
在排序图中,每个环境因子箭头长度反映的是土壤因子与多样性指数变化的关系强弱,箭头连线和排序轴夹角表示某一土壤因子与排序轴的相关性,夹角越小相关性越高[15]。在冗余分析排序图中(图 1),物种丰富度指数(G),多样性指数(D和H)与土壤全钾、速效钾、有机质、全磷和速效磷呈正相关,与pH值、速效氮呈负相关;而均匀度指数(E)和生态优势度指数(λ)与土壤pH值呈正相关,与土壤全钾呈负相关。沿冗余分析第1排序轴,从左到右,物种丰富度指数(G),多样性指数(D和H)随显著性影响因子土壤全钾质量分数的增加而增加,随土壤pH值的增加而减小;均匀度指数和生态优势度指数随土壤全钾质量分数的降低而增加,随土壤pH值的降低而减小。沿第2排序轴从下往上,随土壤速效磷质量分数的降低,物种丰富度指数(G),多样性指数(D和H),均匀度指数(E)减小。

Figure 1. RDA ordination of soil factors and species diversit
3.1. 草本植物物种组成分析
3.2. 草本植物生长期动态分析
3.3. 草本植物物种多样性和土壤因子的关系
-
草本植物是森林生态系统生物多样性的重要组成部分,其物种组成和结构也是森林自然恢复状况及森林生态系统健康结构的重要标志。对草本植物群落的物种多样性研究能更好地认识与了解森林群落组成、结构、功能、演替动态和稳定性。在该天然阔叶红松林样地内草本植物丰富,共调查到草本91种,隶属于34科80属。其中以世界广布科的毛茛科、百合科、伞形科的物种数最多;少种科属占总数的90%以上,单种属在样地内占有绝对优势,单科单属、单属单种现象在该研究区内非常明显。
阔叶红松林作为温带森林的一个特殊森林层片,早春生活周期短暂的草本植物在乔灌木树种处于休眠状态时就开始生长,林分郁闭前完成开花、结果等各个生活阶段,林分郁闭后结束生长进入休眠期[16],夏季植物在林分郁闭前后萌发进行其生活周期。因此,阔叶林内早春、夏季、秋季的草本植物会存在很大的差异。该天然阔叶红松林的林下层草本植物以1年生植物为主,5月早春植物成片生长占有优势,林分郁闭时萌发的早夏植物和晚夏植物在夏末秋初才逐渐枯萎,物种组成在这一完整的生长期内存在较大的差异,尤以5月和9月的差异最为明显,物种的相似度极低,而夏季6月和7月的差异最小。这与李步杭等[7]的调查结果相似,同时Jacaard相似系数随调查间隔时间的延长而变小在时间尺度上物种的相似度会降低。而草本植物植株个体总数从5月到9月呈急剧减少的趋势,可能是由于一些植物长期的生态适应,只能在上层林分未郁闭前进行生活史,郁闭后植株的地上部分会大量死亡导致物种个体总数减少,也可能是植株间相互竞争的影响使植物自然稀疏[17]。Gleason指数与物种数变化趋势一致,以物种数和物种株数为基础构建的Shannon-Wiener指数和Simpson指数在生长季内的变化趋势也基本相同,2个指数反映了该群落草本层多样性在夏季初最高,早春次之,9月最低的特点。这与郝占庆等[3]对长白山北坡草本植物的研究结果一致;但常新华等[8]、夏富才等[18]对天然次生林的研究结果却发现草本层物种多样性的最高值出现在7月。分析其原因有可能是他们研究的次生林中主要阔叶树种多为阳性树种(白桦Betula platyphylla和山杨Populus davidiana等),在春季回温后这些阳性树种花先叶开放,郁闭较晚,草本植物物种数和多样性在该月达到最大。郭利平等[19]在研究中发现长白山红松阔叶林演替的发展过程中,白桦、山杨、黄檗Cortex phellodendri等阳性或半阴性树种减少,而紫椴、水曲柳、色木槭等阴性树种增加。上层乔木的演替变化过程中,其组成和结构通过影响光照、降水以及凋落物等,会对草本层产生直接或间接的影响[2]。
土壤是植物群落的主要环境因子之一,土壤的性质与植物群落组成和物种多样性有着密切的关系,并且多年来一直是生态学家的研究热点[20]。对植物物种多样性及其与土壤环境因子关系的研究,是群落生态学研究和生物多样性保护与管理的基础[11]。本研究显示:该天然阔叶红松林内草本多样性指数与土壤全钾、速效磷和pH值相关性较大(P<0.05),其中与土壤全钾和pH值的关系达到显著相关水平(P<0.01),而其他因子与物种多样性的关系不明显,表明土壤全钾和土壤酸碱度对该天然阔叶红松林草本物种多样性具有显著的影响作用。GARTLAN等[21]研究显示土壤钾的水平与热带植物群落物种多样性之间存在着显著(P<0.05)的相关关系,这与本研究结果相似。土壤酸碱度是土壤内元素有效性和速效性的制约因子[22];本研究区的土壤pH值为pH 6.9~4.7,安树青等[22]研究也表明:对紫金山次生森林群落物种多样性影响较大的土壤因子是土壤厚度、含水量和酸碱度,土壤酸性增加(pH 6.3~5.0),多样性上升,这一研究结果与本研究一致。曾歆花等[23]发现:在植被自然恢复过程中,乔灌草植株生物量的增加使得年凋落物量也随之增加,降水的集中等会影响林下土壤表层养分性质的改变和分布。因此,随着森林群落的演替发展,土壤因子与物种多样性的关系也不尽相同。夏富才等[24]对林龄为老龄林的阔叶红松林草本植物多样性影响因子的研究中表示,速效钾是影响草本植物生长的主要因子。同时他对原始阔叶林与次生林草本植物的环境影响对比研究中发现,次生林中草本植物多样性与土壤养分的相关关系不明显,原始阔叶红松林中土壤有机质含量和土壤酸碱度对草本多样性的影响较为明显[25]。于洋等[26]研究发现天然红松林草本Shannon-Wiener指数与有机质含量呈极显著负相关(P<0.01),全氮、速效钾与草本物种数呈显著负相关(P<0.05)。此外,不同的地理气候条件和土壤因子对物种多样性的影响也不同。丛静等[27]通过对神农架保护区典型植被物种多样性和环境因子的关系分析得出,土壤全氮含量和pH值是影响植物多样性分布格局的主要因素;刘丽丽等[28]研究表示夏蜡梅Sinocalycanthus chinensis群落物种多样性与土壤因子有机质含量相关性较大。
草本植物多样性受多种因素的影响,例如地形、海拔、光照、林隙、土壤肥力等。夏富才等[25]对长白山阔叶红松林林下草本植物的研究表明,林隙分布和土壤酸碱度是影响草本植物多样性的主要因素;曹云生等[29]对不同森林群落地形因子进行的典范分析认为:坡向是影响草本多样性空间差异的主要地形制约因子,海拔与各多样性指数呈正相关;也有研究表示,海拔在一定范围内变动时草本多样性指数波动不大,坡度与草本多样性关系较明显[30]。草本多样性受多种环境因子的制约,各种环境因子又相互作用共同影响其多样性。由于研究区地形较平坦,海拔的变化差异也很小,最大高程差仅为100 m,故本研究仅分析了草本植物物种多样性与土壤因子的关系,在冗余分析土壤因子中,虽然土壤因子只解释了39.2%的物种多样性变化格局,尚有60.8% 的变化部分未能得到相应的解释,说明还有一些未知的环境因子或其他因素在植物生长过程中起了重要的作用。张春雨等[31]在长白山次生林植物空间分布及环境解释研究中曾提到在纯粹的环境因子解释能力较低。环境因子未能解释的部分主要反映了群落自身互作及人类活动(采伐或清林等经营行为)对植物种群格局的影响,以及一些未涉及的环境因子、干扰事件或随机因子的影响[21]。本研究区域虽属于典型的低海拔、人为活动频繁区域,但近年来在半封育的管理下,人类的采伐破坏活动减少,物种多样性与珍稀植物也愈趋丰富,故未能解释的部分很大程度上可能依赖于群落自身的演替以及一些随机干扰因子的影响(如全球气候变化)。要全面了解天然阔叶红松林的草本植物物种多样性,需将草本植物的生理生态特征与森林群落自身特征和环境因子等结合起来做进一步的研究。
 DownLoad:
DownLoad: