







-
物种多样性是维持植物群落生态系统的基础,是人类赖以生存的生物资源。研究植物群落物种多样性能更好地保护与利用多样性资源,已成为了植物生态学领域的热点[1-4]。群落物种多样性受自然界中普遍存在的干扰尤其是人为干扰的影响,日益频繁的人类活动已成为当前引起生态系统结构和功能变化的主要因素[5-6],因此,研究不同干扰强度对群落物种多样性的影响意义重大[7-8]。紫楠Phoebe sheareri又名紫金楠,也称金丝楠,系樟科Lauraceae楠属Phoebe中极为珍贵的中国特有树种。其木材坚硬致密,纹理直,有光泽,耐腐且富楠木芳香,是高级家具、雕刻的理想用材;同时可作为园林绿化树种。紫楠喜温暖、湿润的环境,常见于海拔1 000 m以下的山地沟谷阔叶林中,在江苏、浙江、安徽、江西、福建、湖南、贵州、广西等省均有自然散状分布。由于气候变迁,人类砍伐,旅游干扰,自然更新能力弱等原因,现存紫楠天然种质资源已接近枯竭[9]。对分布区的实地调查发现,现存较大的天然群落仅位于浙江省的杭州市灵隐、杭州市临安区、建德市、淳安县等部分区域[10]。目前,针对紫楠的研究主要集中在种子生理特征[11],幼苗光合特性[12],叶片细胞形态与生长特征[13-14]等方面,对紫楠天然群落物种多样性的研究仅涉及单个群落[15-16],而对其全分布区内群落物种多样性程度的研究未见报道。本研究在紫楠天然分布区内选择20个代表性的群落并研究物种组成,意在阐明干扰强度及生态因子对紫楠天然群落物种多样性的影响,为紫楠种质资源的保护、利用提供基础性资料。
HTML
-
紫楠群落的天然分布区地理位置跨度大(24.50°~32.05°N,106.38°~121.45°E),包括江苏、浙江、江西、湖南、贵州、广西等6个省份(自治区)的16个县(区),地理区划属于华东、华中、华南和西南地区,植被区划属于亚热带常绿阔叶林区域。研究区土壤以红壤、黄壤为主。地形差异显著,海拔为41~1 020 m,年平均日照时数为1 035.2~1 980.0 h,年均温为13.1~19.6 ℃,年均降水量为1 109.9~1 850.5 mm;年无霜期为216~322 d。植物区系组成的优势科为樟科、壳斗科Fagaceae和山茶科Theaceae等。
-
在对紫楠天然分布点实地踏查的基础上,选择生境条件、种群大小等代表性的20个天然群落进行样地调查(表 1)。采用全球定位系统(GPS)记录各样地的经纬度及海拔,用罗盘仪测定坡度,记录样地郁闭度,并查阅获得样地日照时数、气温、降水量和无霜期的年均值。植物群落干扰强度的确定至今仍缺乏科学的定量化标准与统一方法,本研究综合BHUYAN等[17]、吴甘霖等[18]、郝建锋等[19]的研究方法,将干扰强度划分为5个等级:种群远离居民区或核心景区道路,近年来未受抚育、砍伐、采摘或旅游等人为干扰,且无病虫害干扰称为1级干扰;离居民区较远(>5 km)或在核心景区道路两侧1 000 m以外,受抚育、砍伐、采摘或旅游等人为干扰较少的天然林称为2级干扰;离居民区较远(>2 km)或在核心景区道路两侧500 m以外,受人为干扰破坏,但受抚育、砍伐、采摘、旅游或病虫害等干扰较少的天然林称为3级干扰;离居民区较近(≤2 km)或在核心景区道路两侧500 m以内,受人为干扰破坏,同时受抚育、砍伐、采摘、旅游或病虫害等干扰较多的天然林称为4级干扰;离居民区较近(≤1 km)或在核心景区道路两侧200 m以内,反复遭受抚育、砍伐、采摘、旅游或病虫害等干扰破坏的天然林称为5级干扰。
干扰等级 群落 纬度(N) 经度(E) 海拔/m 坡度/(°) 郁闭度 年日照/h 年均温/℃ 年降水量/mm 年无霜期/d 1 ZJB 29°26′56″ 119°12′02″ 150 25 0.85 1 760.0 17.4 1 600.5 254 CA 29°28′02″ 119°07′56″ 310 30 0.90 1 950.5 17.0 1 430.0 263 SC 28°37′35″ 119°27′50″ 620 20 0.85 1 755.0 16.8 1 510.0 250 LS 25°37′52″ 109°55′14″ 734 35 0.85 1 465.5 18.1 1 705.5 314 2 ZJS 32°03′05″ 118°52′11″ 41 7 0.75 1 628.8 19.6 1 530.1 322 CH 30°07′48″ 119°07′29″ 315 30 0.80 1 850.0 15.5 1 450.5 225 LC 27°02′35″ 116°55′27″ 460 15 0.75 1 642.8 17.9 1 749.5 287 ZY 25°51′20″ 110°32′27″ 634 28 0.75 1 407.6 16.4 1 761.1 280 3 NH 29°26′34″ 121°24′01″ 140 38 0.70 1 900.5 16.1 1 300.0 216 TMS 30°19′49″ 119°26′19″ 510 13 0.75 1 820.0 14.5 1 400.0 222 QY 27°40′44″ 119°23′20″ 660 25 0.75 1 980.0 17.4 1 760.0 255 FJS 27°53′32″ 108°43′31″ 949 25 0.75 1 035.2 13.9 1 850.5 274 4 LJ 29°36′09″ 119°40′23″ 180 25 0.70 1 680.5 15.8 1 650.0 256 THY 30°23′18″ 119°33′29″ 460 40 0.70 1 900.0 15.9 1 425.5 223 ZJ 27°31′19″ 109°32′27″ 569 40 0.70 1 527.7 16.4 1 294.6 279 XS 28°29′24″ 106°23′12″ 1 020 30 0.70 1 053.0 13.1 1 109.9 248 5 XH 30°13′44″ 120°05′36″ 100 15 0.60 1 765.5 17.8 1 454.5 245 LHT 29°26′38″ 119°07′47″ 280 9 0.65 1 750.5 17.1 1 550.0 261 SY 28°23′43″ 119°24′04″ 350 20 0.60 1 840.0 17.7 1 700.0 247 XN 26°16′34″ 110°43′40″ 490 30 0.65 1 431.5 16.2 1 680.5 291 说明:ZJB:浙江朱家埠;CA:浙江淳安;SC:浙江遂昌;LS:广西龙胜;ZJS:江苏紫金山;NH:浙江宁海;LJ:浙江泷江;THY:浙江太湖源;LC:江西黎川;TMS:浙江天目山;ZJ:湖南芷江;CH:浙江昌化;ZY:广西资源;QY:浙江庆元;FJS:贵州梵净山;XS:贵州习水;XH:浙江西湖;LHT:浙江绿荷塘;SY:浙江松阳;XN:湖南新宁 Table 1. Habitats of sample plots of Phoebe sheareri
-
采用典型样方法。根据紫楠种群大小,每个群落设置1~2个20 m × 20 m的样地,并在其中设置2 m × 2 m的灌木样方和1 m × 1 m的草本样方。详细调查并记录乔木层树种名称、胸径、地径、高度、冠幅;灌木层(包括未满足乔木层测量标准的更新幼树、苗)植物名称、平均高度、冠幅、盖度及多度;草本层植物种名、平均高度、盖度及多度[20-22]。
-
以树种个体数为基础,计算各个种的重要值:重要值=(相对密度+相对频度+相对优势度)/3。
参考国内外植物群落研究中物种多样性指数的测度方法[18-19]:①物种丰富度Patrick指数以物种数(S)作为衡量物种丰富度的参数R=S。②物种多样性Shannon-Wiener指数H′=-∑PilnPi。③物种优势度Simpson指数D=1-∑Pi2。④反映群落内物种分布的均匀程度Pielou指数E1=H′/lnS。其中,Pi为i种的相对重要值。
2.1. 样地设置
2.2. 调查方法
2.3. 数据处理
-
在总面积为10 400 m2的26个紫楠种群样地内,共有维管束植物103科236属363种,其中蕨类植物12科16属19种,种子植物91科220属344种,包括裸子植物6科7属7种,被子植物85科213属337种。其中含5个属以上的科有樟科,壳斗科,蔷薇科Rosaceae,豆科Leguminosae,大戟科Euphorbiaceae,金缕梅科Hamamelidaceae,葡萄科Vitaceae,山茱萸科Cornaceae,禾本科Gramineae,菊科Compositae,荨麻科Urticaceae,茜草科Rubiaceae,百合科Liliaceae,虎耳草科Saxifragaceae,共14科,含164种植物;占总科数的13.59%,占总种数的45.18%,是紫楠群落的主要组成成分。仅含1个种的有45科,占总科数的43.69%,占总种数的12.40%。同时,天然紫楠群落还伴生有较多珍贵稀有的物种,如浙江楠Phoebe chekiangensis,浙江樟Cinnamomum chekiangensis,银杏Ginkgo biloba,柏木Cupressus funebris,南方红豆杉Taxus mairei等。
-
对20个紫楠天然群落内种子植物的91科进行分布区类型统计分析(表 2),共划分出11个分布区类型,占比较高的类型为世界分布(30科)、泛热带分布(25科)与北温带分布(15科),分别占32.97%,27.47%,16.48%;而中国特有分布的仅1科,为银杏科Ginkgoaceae。对20个紫楠天然群落内种子植物的220属进行统计分析(表 2),共划分出13个分布区类型,占比较高的类型为东亚分布(41属)、北温带分布(40属)、泛热带分布(35属)和热带亚洲分布(26属),分别占总属数的18.64%,18.18%,15.91%和11.82%,中国特有分布的共5属,为喜树属Camptotheca,杉木属Cunninghamia,青钱柳属Cyclocarya,牛鼻栓属Fortunearia和银杏属Ginkgo。
分布区类型 科 属 数量 比例/% 数量 比例/% 世界分布 30 32.97 13 5.91 泛热带分布 25 27.47 35 15.91 热带亚洲和热带美洲间断分布 7 7.69 6 2.73 旧世界热带分布 2 2.20 12 5.45 热带亚洲至热带大洋洲分布 2 2.20 6 2.73 热带亚洲至热带非洲分布 1 1.10 4 1.82 热带亚洲分布 1 1.10 26 11.82 北温带分布 15 16.48 40 18.18 东亚和北美间断分布 4 4.40 23 10.45 旧世界温带分布 0 0 8 3.64 地中海区、西亚至中亚分布 0 0 1 0.45 东亚分布 3 3.30 41 18.64 中国特有分布 1 1.10 5 2.27 合计 91 100.00 220 100.00 Table 2. Distribution area of families and genera of seed plants in the Phoebe sheareri communities
-
表 3列出了不同干扰强度下群落内平均重要值大于5%的优势种。在不同干扰强度下,群落各层次物种组成的种类和数量具有明显差异。1级干扰的乔木层中重要值大于5%的有5种,灌木层有5种;2级干扰的乔木层有5种,灌木层有4种;3级干扰和4级干扰的乔木层与灌木层都为4种;而5级干扰的乔木层与灌木层都仅为3种。乔木层常见树种为紫楠,薄叶润楠Machilus leptophylla,枫香Liquidambar formosana,杉木Cunninghamia lanceolata,柳杉Cryptomeria fortunei,红楠Machilus thunbergii,青冈栎Cyclobalanopsis glauca及黄果朴Celtis sinensis等;灌木层常见紫楠,薄叶润楠,茶Camellia sinensis,阔叶箬竹Indocalamus latifolius,紫麻Oreocnide frutescens,毛花连蕊茶Camellia fraterna,杜茎山Maesa japonica,并伴有香花崖豆藤Millettia dielsiana,络石Trachelospermum jasminoides等藤本植物。紫楠重要值在不同干扰强度下具有一定差异,乔木层中从高到低依次为5级干扰、1级干扰、2级干扰、3级干扰、4级干扰;灌木层中从高到低依次为2级干扰、1级干扰、3级干扰、4级干扰、5级干扰。
干扰强度 群落层次 优势种 重要值/% 1 乔木层 紫楠phoebe sheareri 35.63 薄叶润楠Machilus leptophylla 21.07 红楠Machilus thunbergii 11.90 黄果朴Celtis sinensis 11.57 青冈栎Cyclobalanopsis glauca 10.21 灌木层 紫楠Phoebe sheareri 17.24 薄叶润楠Machilus leptophylla 13.83 茶Camellia sinensis 12.72 香花崖豆藤Millettia dielsiana 11.67 毛花连蕊茶Camellia fraterna 11.36 杜茎山Maesa japonica 10.51 2 乔木层 紫楠Phoebe sheareri 30.37 枫香Liquidambar formosana 15.17 杉木Cunninghamia lanceolata 11.38 黄果朴Celtis sinensis 10.24 青冈栎Cyclobalanopsis glauca 10.07 灌木层 紫楠Phoebe sheareri 20.57 紫麻Oreocnide frutescens 17.15 茶Camellia sinensis 12.80 络石Trachelospermum jasminoides 10.29 3 乔木层 紫楠Phoebe sheareri 28.33 薄叶润楠Machilus leptophylla 21.36 枫香Liquidambar formosana 16.95 柳杉Cryptomeria fortunei 16.65 灌木层 高粱泡Rubus lambertianus 14.28 络石Trachelospermum jasminoides 10.94 紫楠Phoebe sheareri 10.88 阔叶箬竹Indocalamus latifolius 10.26 4 乔木层 紫楠Phoebe sheareri 21.92 杉木Cunninghamia lanceolata 16.48 薄叶润楠Machilus leptophylla 16.34 枫香Liquidambar formosana 10.02 灌木层 薄叶润楠Machilus leptophylla 23.76 紫楠Phoebe sheareri 10.30 茶Camellia sinensis 10.53 5 乔木层 紫楠Phoebe sheareri 42.01 枫香Liquidambar formosana 17.17 薄叶润楠Machilus leptophylla 10.93 灌木层 薄叶润楠Machilus leptophylla 18.61 紫楠Phoebe sheareri 10.55 茶Camellia sinensis 10.05 Table 3. Dominant species and their importance values of communities under different disturbance intensity
-
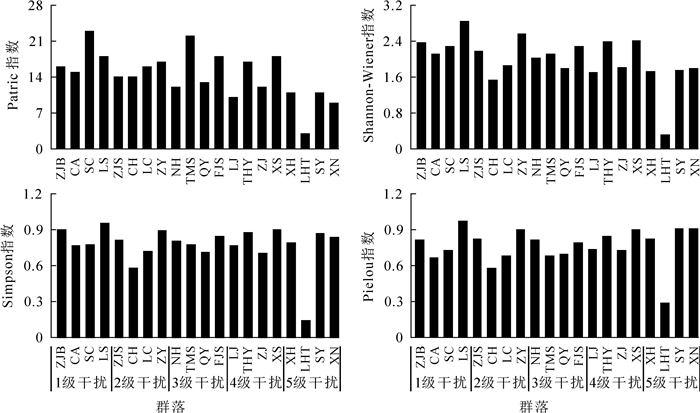
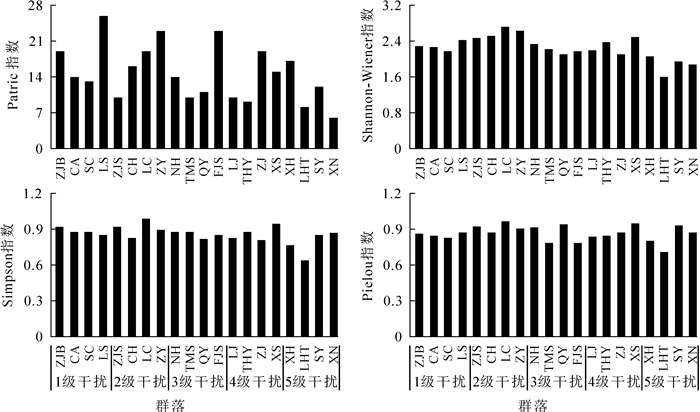
受干扰、环境的影响,不同群落物种组成与数量差异明显。从图 1和图 2可见:Shannon-Wiener指数、Simpson指数、Pielou指数的变化趋势基本一致。乔木层中这3个指数最大值都出现在1级干扰的LS,Patrick指数最大为SC;最小值都出现在5级干扰的LHT。灌木层Patrick指数最大值和最小值分别出现在1级干扰的LS和5级干扰的XN,而Shannon-Wiener指数、Simpson指数和Pielou指数的最大值和最小值均出现在2级干扰的LC和5级干扰的LHT。

Figure 1. Species diversity in the tree layer of Phoebe sheareri communities

Figure 2. Species diversity in the shrub layer of Phoebe sheareri communities
-
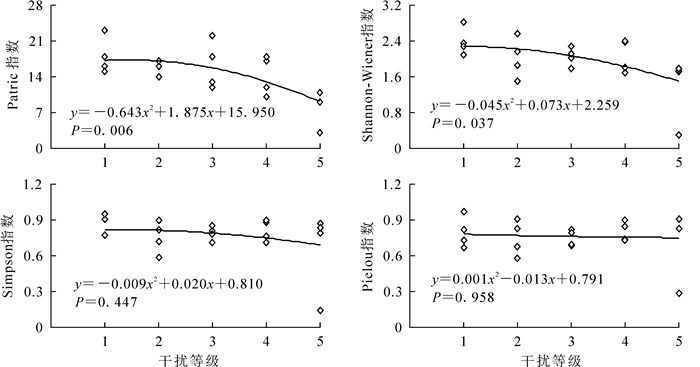
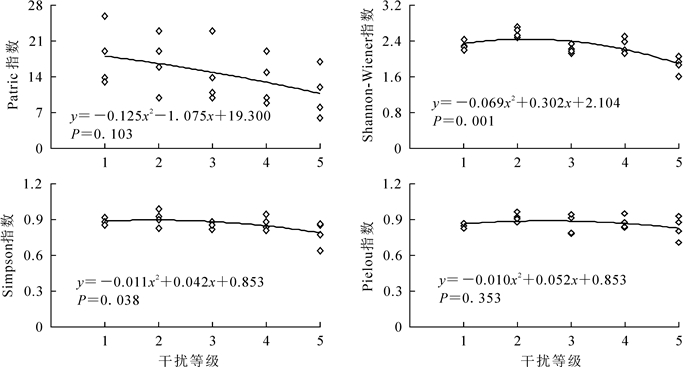
运用Quadratic曲线对物种多样性指数与干扰强度的回归分析(图 3和图 4)可见:4个多样性指数值整体上均随干扰强度的增加而减小。其中,乔木层Patrick指数与干扰强度呈极显著相关,Shannon-Wiener指数与干扰强度呈显著相关,说明干扰越强,群落乔木层物种丰富度与多样性程度越低。灌木层Shannon-Wiener指数与干扰强度呈极显著相关,Simpson指数与干扰强度呈显著相关。在2级干扰强度下,灌木层Shannon-Wiener指数和Simpson指数达最大值,而之后随干扰强度增加而减小。

Figure 3. Relationship between disturbance intensity and species diversity index in the tree layer

Figure 4. Relationship between disturbance intensity and species diversity index in the shrub layer
各群落物种多样性指数与其地理生态因子间进行Pearson相关分析(表 4)表明:乔木层和灌木层的4个多样性指数均与经度、纬度呈负相关,均与海拔高度、坡度呈正相关,均与年日照时长呈负相关。其中,乔木层Patrick指数与海拔、郁闭度分别呈显著与极显著正相关,Shannon-Wiener指数与郁闭度呈显著正相关;灌木层Patrick指数与经度、纬度都呈显著负相关,Shannon-Wiener指数与郁闭度呈显著正相关。但乔木层和灌木层的物种多样性指数与坡度、年均日照、年均温、年降水量、年无霜期等生态因子间的相关性均未达到显著水平。
群落层次 多样性指数 r 纬度 经度 海拔 坡度 郁闭度 年日照 年均温 年降水量 年无霜期 乔木层 R -0.067 -0.181 0.485* 0.115 0.596** -0.180 -0.284 0.074 -0.075 H' -0.244 -0.366 0.388 0.407 0.479* -0.321 -0.105 0.012 0.183 D -0.224 -0.303 0.248 0.386 0.193 -0.286 -0.075 0.061 0.132 E1 -0.322 -0.400 0.239 0.394 -0.036 -0.373 0.000 0.072 0.227 灌木层 R -0.492* -0.494* 0.370 0.269 0.381 -0.416 -0.008 0.229 0.303 H' -0.059 -0.162 0.154 0.216 0.472* -0.117 -0.036 -0.065 0.115 D -0.120 -0.188 0.193 0.152 0.376 -0.171 -0.062 -0.020 0.152 E1 -0.269 -0.159 0.110 0.248 0.051 -0.013 0.193 0.005 0.199 说明:*表示显著相关(P < 0.05),**表示极显著相关(P < 0.01) Table 4. Correlation between geo-ecological factors and diversity index in the Phoebe sheareri community
3.1. 紫楠群落物种组成
3.2. 种子植物科、属分布区类型
3.3. 不同干扰强度下紫楠群落优势树种的比较
3.4. 群落物种多样性特征
3.5. 物种多样性与干扰强度及生态因子的关系
-
本研究表明:调查样地占地面积为10 400 m2,共有维管束植物103科236属363种,其中蕨类植物12科16属19种,种子植物91科220属344种,紫楠伴生植物的科、属、种类型较为丰富。紫楠群落物种组成丰富度显著高于多数针叶树种[23],而与分布较广的麻栎Quercus acutissima[24],米槠Castanopsis carlesii[25],甜槠Castanopsis eyrei[26]等阔叶树种的群落相当。在全国15个种子植物科(属)的分布区类型中,紫楠群落伴生种中科的分布区类型出现11种,属的分布区类型出现13种,以北温带和泛热带成分占明显优势。可见紫楠适合的生境范围较广,适与多种植物相互伴生组成群落,其伴生植物区系具有热带向北温带过渡的特征,符合紫楠天然群落常见于亚热带常绿阔叶林的特点。
干扰强度导致不同植物群落差异显著[27]。通过分析群落内平均重要值大于5%的优势种发现,干扰强度越大,群落整体物种组成与数量都越少。5级干扰紫楠天然群落,乔木层多以紫楠为主,树种组成相对单一,灌木层中紫楠往往因受干扰较大而难以生长,尤其是位于浙江建德绿荷塘楠木林的紫楠群落,乔木层仅有紫楠、薄叶润楠、刨花楠Machilus pauhoi 3种,且紫楠占绝对优势地位,天然混交林逐渐向紫楠纯林演变,一旦出现气候灾害或病虫害,群落最终会走向灭亡[28]。
不同群落的物种多样性主要表现在丰富度Patrick指数、多样性Shannon-Wiener指数、优势度Simpson指数和均匀度Pielou指数的差异。乔木层4个多样性指数最小值都出现在5级干扰的LHT,表明重度干扰会降低群落的多样性水平,这与郝建锋等[29]的研究结果相符。灌木层Patrick指数最大的为1级干扰的LS,Shannon-Wiener指数、Simpson指数、Pielou指数的最大值都出现在2级干扰的LC,可见适度的干扰可扩大林窗面积,对灌木层物种多样性具有促进作用,印证了中度干扰假说[30]。
紫楠群落物种多样性指数值整体上均随干扰强度的增加而减小。乔木层Patrick指数与干扰强度呈极显著相关,Shannon-Wiener指数与干扰强度呈显著相关,说明干扰越强,群落乔木层物种丰富度与多样性程度越低,与李建民等[31]的研究结果相符。2级干扰强度时,群落灌木层Shannon-Wiener指数、Simpson指数和Pielou指数均大于1级干扰,但在3级干扰强度下,灌木层物种多样性指数随干扰强度的增加而减小,进一步佐证了中度干扰假说[32]。对物种多样性指数与地理生态因子进行Pearson相关分析结果表明:乔木层、灌木层物种多样性指数与郁闭度显著相关,而与年均日照、年均温、年降水量、年无霜期等气候条件均无显著相关性;群落郁闭度在一定程度上代表了干扰强度,说明与气候条件相比,干扰强度对群落物种多样性的影响更大[33]。
紫楠为珍贵商品木材“金丝楠木”的原植物种,在人为干扰和环境变化下,现存天然种群面积日益减少且资源片段化极其严重,其天然资源保护与合理利用刻不容缓。因此,建议各地政府应切实加强对紫楠现存资源的就地保护,在分布较集中的浙江杭州、临安、建德、淳安,南部的广西龙胜、资源以及西部的贵州梵净山等地建立重点保护区,尽可能最大限度保存野生珍贵资源;针对干扰较强的群落(如LHT和XH等)进行适度人工干预,提高群落物种多样性水平,促进紫楠种群自然更新与发展;开展人工育苗与造林,不断扩大紫楠资源分布与基因库;开展分子遗传学等研究,进一步了解紫楠种群遗传结构及其濒危成因,确定重点保护单元,制定科学保护策略。
 DownLoad:
DownLoad: