







-
森林生物量是量化森林生态系统结构和功能的重要指标之一,对揭示森林生态系统的固碳潜力、生产力及物质循环等关键过程具有重要的指示作用[1-3]。林下植被作为森林的重要组成部分,是研究森林生态系统的功能及稳定性的基础之一[4]。以往研究表明:林下植被多样性越高越有利于促进森林凋落物分解、养分循环以及改善土壤等[5-8]。中国是全球拥有最大面积人工林的国家,人工林面积增长速率快,约占全球人工林面积三分之一,对减缓全球气候变化和支撑国家生态文明建设具有重要作用[9]。然而,目前人工林的树种结构单一、地力衰退、稳定性和抗逆性较差、生产力和生物多样性逐渐降低等生态问题日益凸显[10]。因此,研究森林生物量及其林下植被多样性,有助于认识森林生态系统的结构功能和稳定性机制,可为人工林的可持续经营与管理提供科学依据。
目前国内外学者主要探讨了不同林龄、间伐强度、海拔等对森林生物量与林下植被多样性的影响。如:赵金龙等[11]研究发现:河北省平泉市辽河源自然保护区不同林龄油松Pinus tabuliformis各器官生物量分配存在显著差异,幼龄林和近熟林各器官生物量从大到小依次为干、根、枝、叶、干皮、球果,而中龄林和成熟林则是干、根、枝、干皮、叶、球果;刘思泽等[12]通过对川东低山区29年生马尾松P. massoniana人工林研究发现:随着间伐强度增加,林下植物多样性指数先增后降。近年来,不同树种对生物量及林下植被多样性的影响也逐渐受到关注,如葛露露等[13]研究发现:福州市滨海沙地不同树种人工林生物量存在差异,从大到小依次为木麻黄Casuarina equisetifolia、尾巨桉Eucalyptus urophylla、纹荚相思Acacia aulacocarpa。另外,树种可以通过改变林下微环境(如光照、水分、温度等),从而影响林下植被多样性[14]。如MESTRE等[15]研究发现:落叶林比常绿林有着更高的林下植被多样性。然而,此类研究均在不同区域或不同林型等开展,其结果可能会受到土壤、地形、海拔、降水、土地利用方式等立地条件的影响,难以量化树种对生物量及林下植被多样性的影响效应。
同质园(common garden)研究具有立地条件基本一致的特点,能最大程度地降低由气候、土壤、地形等环境因素导致的差异[16]。目前国内同质园研究主要在植物叶片功能和遗传变异[17-19]、土壤养分[20-21]等方面,而关于亚热带树种对林分生物量及林下植被多样性的影响研究还鲜有报道。据四川省第9次森林资源清查结果显示:四川省森林覆盖率为38.03%,森林面积为1 840万 hm2,人工林主要以马尾松[22]、云杉Picea asperata[23]等针叶纯林以及速生外来树种桉树Eucalyptus林为主,其面积和蓄积量分别为502万 hm2、25 546万 m3。人工纯林极易引发各种生态问题,如松材线虫Bursaphelechus xylophilus病、火灾等[10]。营建乡土阔叶树种林分既能兼顾珍贵用材生产,又能增加林下植被多样性和生态服务功能,已逐渐成为替代亚热带地区大型针叶林的主要森林经营模式[24]。然而,如何科学选择适生乡土阔叶树种是培育人工林和提升森林生产力及生态服务功能亟须解决的关键问题。大叶樟Cinnamomum platyphyllum、油樟C. longepaniculatum、天竺桂C. japonicum、樟树C. camphora、桤木Alnus cremastogyne、香椿Toona sinensis、红椿T. ciliata是四川常见的珍贵乡土阔叶树种,具有较高的经济与生态价值,但至今缺乏必要关注,因此,本研究以亚热带同质园这7个树种林分作为研究对象,探讨树种对生物量分配特征及对林下植被多样性的影响,为营建亚热带人工林筛选适生的珍贵乡土阔叶树种及其管理提供科学依据。
-
同质园位于四川省崇州市桤泉镇四川农业大学现代农业研发基地(30°55′N,103°49′E),海拔为516 m,属四川盆地亚热带湿润季风气候,该地区年平均气温为16.0 ℃,年平均降水量约1 015.2 mm,年平均日照时数为1 161.5 h,平均无霜期约285 d。
-
2015年春季,采用随机区组设计建立了同质园试验区。样地前期为农耕地,经营历史相同,土壤均一,地势平坦。种植前,对同质园进行了翻耕,使样地土壤更加均一,本底土壤理化性质基本一致。统一选用2年生树苗,按2 m×2 m的株行距挖穴造林,初植林分密度为1 600株·hm−2。每个样地面积为25 m×25 m,每个树种设置3个重复样地,在各样地边缘设置宽度为1 m的排水沟和缓冲带。造林后进行统一管理,使其自然生长,并且围建栅栏以防止人为活动的干扰(图1)。
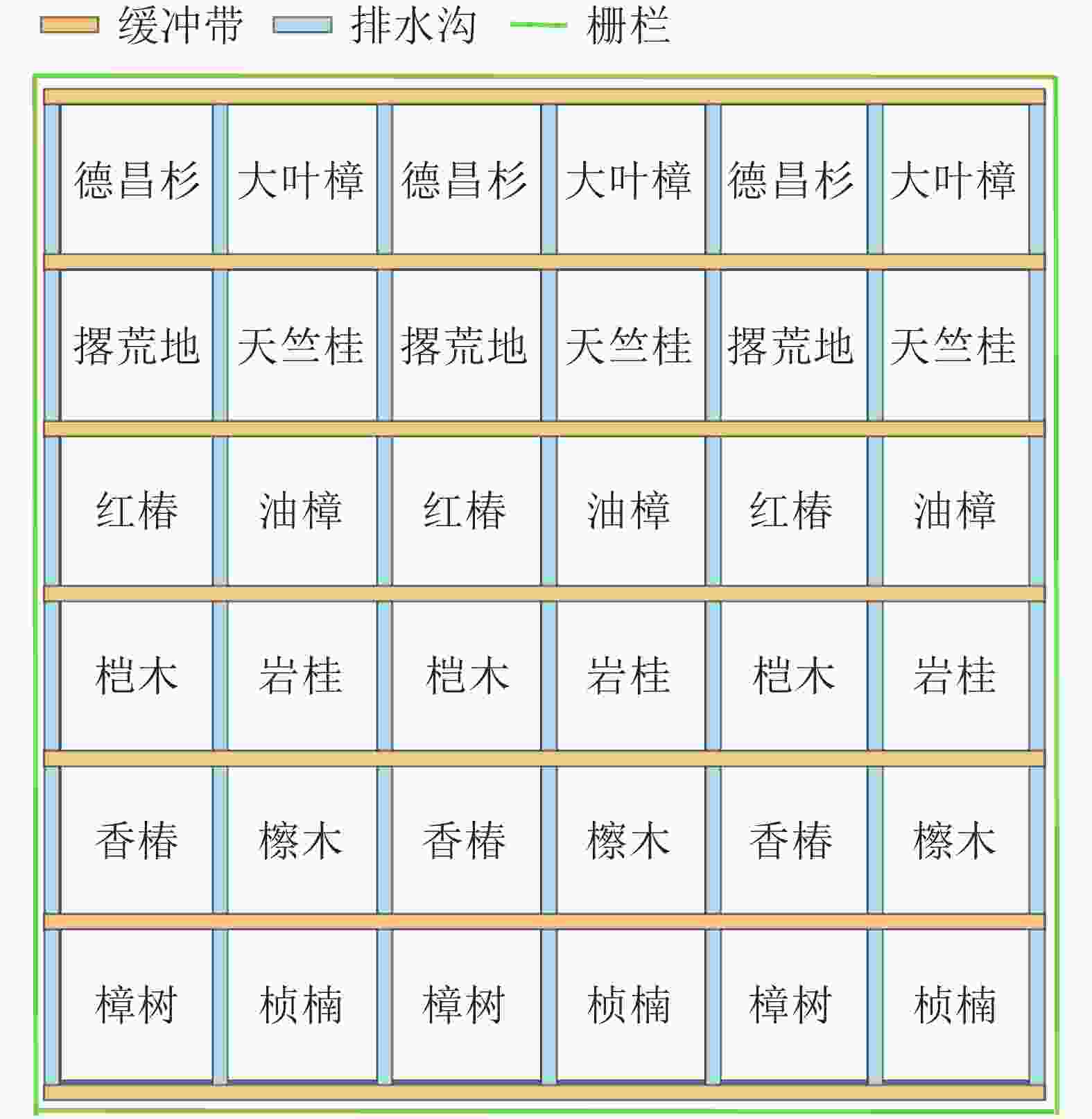
Figure 1. Common garden layout diagram
-
2020年7月底,选择长势良好,现存林分密度基本一致,当年种植存活率为90%以上的大叶樟、油樟、天竺桂、樟树、桤木、香椿、红椿等7个单一树种林分作为研究对象,每个林分3个重复,共21个样地。存活率低且生长较差的树种(德昌杉Cunninghamia unica、桢楠Phoebe zhennan、岩桂Cinnamomum pauciflorum、檫木Sasafras tsumu等)未纳入本研究。对每个样地的乔木进行每木检尺,用LI-RD 1000激光测树仪(美国)测量树高,用胸径尺测量每株乔木在树高1.3 m处的胸径,计算平均树高和平均胸径。
在每个样地分别随机布置2个5 m × 5 m的灌木样方和1 m × 1 m的草本样方,调查灌木和草本样方内的所有植物种类、数量和高度等。同时,用手捡法采集草本样方中现存凋落物,装入信封保存,带回实验室,放入65 ℃恒温烘箱中烘干至恒量,用电子天平秤称量,计算单位面积现存凋落物量。同时,为比较功能群植物对生物量及林下植被多样性的影响,按照叶寿命分为常绿树种(大叶樟、油樟、樟树和天竺桂)和落叶树种(桤木、红椿和香椿)。
-
采用实际伐木计算并验证相近林龄同种同属或其他多种树种通用的生物量模型[25-27]。如表1所示:通过平均树高(H)和平均胸径(DBH),采用二元回归方程计算单株林木各器官生物量(W),方程为W=a(DBH2H)b。其中,a和b为方程参数。
树种 器官 a b 相关系数 大叶樟、油樟、
天竺桂、樟树叶 0.059 871 0.574 327 0.989 1 枝 0.006 652 1.051 841 0.986 8 干 0.055 603 0.850 193 0.962 7 根 0.184 736 0.616 421 0.968 6 桤木 叶 0.010 593 0.813 953 0.860 0 枝 0.014 972 0.875 639 0.920 0 干 0.027 388 0.898 869 0.900 0 根 0.036 227 0.728 875 0.900 0 红椿、香椿 叶 0.013 347 0.689 300 0.968 1 枝 0.025 860 0.807 600 0.975 0 干 0.038 352 0.907 600 0.984 7 根 0.014 732 0.874 600 0.975 3 Table 1. Regression equation of biomass of different organs of different tree species
由于各群落林下灌木只有构树Broussonetia papyrifera或无灌木,灌木层多样性差异较小,因此,本研究着重分析草本层的多样性,通过公式计算不同群落草本层的Simpson多样性指数(λ)和Shannon-Wiener多样性指数(H′)[28]。
其中:Pi为第i种的个体数占调查物种个体总数的比例,i=1,2,3,
$\cdots $ ,n。 -
采用Excel 2016对数据进行整理,单因素方差(one-way ANOVA)分析不同树种群落对乔木层各器官生物量、占比、根冠比及林下草本层生物多样性的影响;利用独立样本t检验分析常绿树种和落叶树种对乔木层各器官生物量、生物量占比、根冠比及林下草本层生物多样性的影响。采用Duncan多重比较法检验显著性(P<0.05)。采用SPSS 22.0软件进行数据分析,用Origin 9.5进行绘图,文中数据均为平均值±标准误。
-
由表2所示:不同树种的平均树高、平均胸径、现存凋落物量和郁闭度均具有显著差异(P<0.05)。树高从大到小依次为桤木、大叶樟、红椿、樟树、香椿、油樟、天竺桂;胸径为红椿、桤木和大叶樟最大,其次是油樟、香椿和樟树,最小的是天竺桂;郁闭度从大到小依次为大叶樟、油樟、天竺桂、樟树、桤木、红椿、香椿。现存林分密度最大的是红椿,最小是大叶樟。另外,7个树种林分的林下灌木植物主要是构树Broussonetia papyrifera,林下草本植物主要是空心莲子草Alternanthera philoxeroides、茜草Rubia cordifolia、竹叶草Oplismenus compositus、蛇莓Duchesnea indica、华西凤尾蕨Pteris occidentalisinica等。
树种 平均树高/m 平均胸径/cm 现存凋落物量/(kg·hm−2) 郁闭度 现存密度/(株·hm−2) 林下灌木 林下草本 大叶樟 10.8±0.3 a 10.6±0.3 a 700.9±106.4 b 0.85±0.03 a 1 184 构树 空心莲子草、茜草等 油樟 7.0±0.3 d 7.7±0.1 b 388.9±92.5 c 0.90±0.25 a 1 274 空心莲子草、蛇莓等 天竺桂 6.1±0.2 e 5.1±0.3 c 160.2±45.9 cd 0.73±0.26 a 1 248 构树 空心莲子草、华西凤尾蕨等 樟树 9.0±0.5 bc 7.1±0.3 b 986.8±197.0 a 0.89±0.04 a 1 248 空心莲子草、竹叶草等 桤木 11.0±0.3 a 10.7±0.0 a 136.9±40.4 cd 0.66±0.53 b 1 251 构树 空心莲子草、茜草等 红椿 9.7±0.3 b 11.2±0.4 a 17.1±3.9 d 0.67±0.58 b 1 290 构树 空心莲子草、茜草等 香椿 8.7±0.1 c 7.7±0.3 b 15.6±2.1 d 0.67±0.05 b 1 274 构树 空心莲子草、竹叶草等 说明:同列不同小写字母表示树种间差异显著(P<0.05) Table 2. General situation of plantation in common garden
-
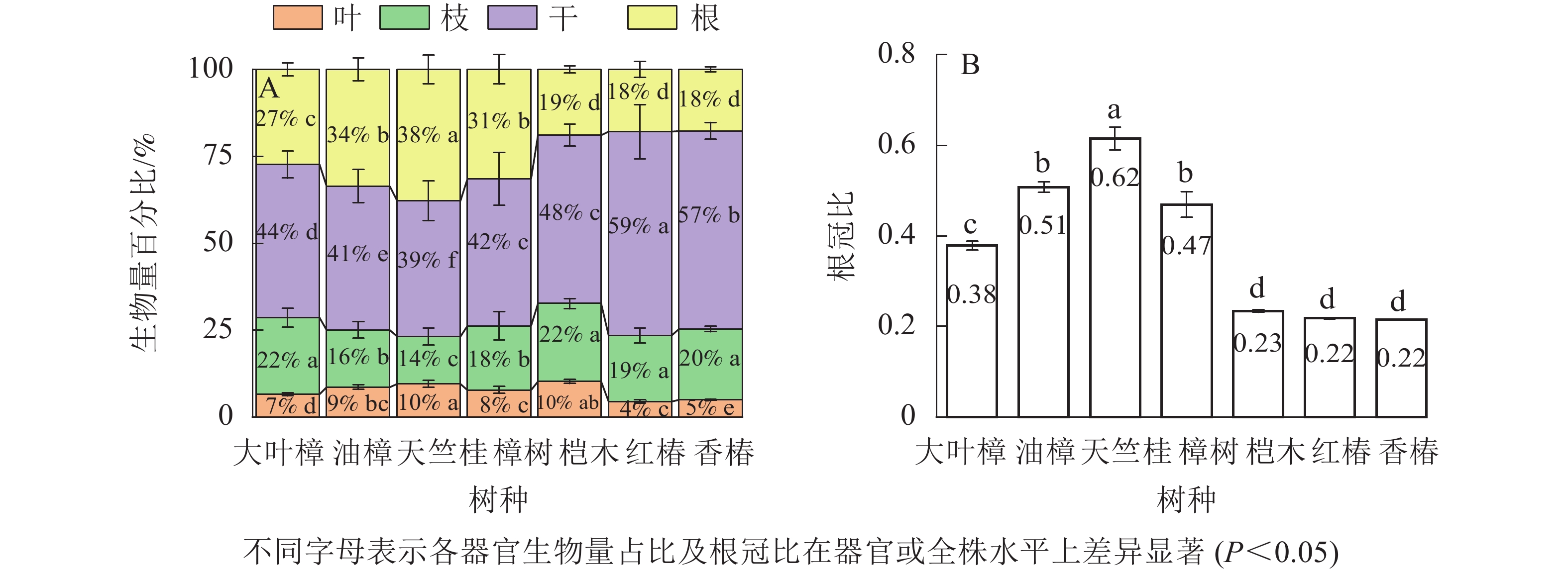
图2所示:不同树种的全株生物量及各器官生物量存在显著差异(P<0.05)。大叶樟的全株生物量最高,其次是桤木、红椿和樟树,而油樟、香椿和天竺桂较低。大叶樟和桤木的叶生物量较高,油樟和樟树次之,而红椿、天竺桂和香椿最低。大叶樟的枝生物量显著高于其他树种(P<0.05),油樟和天竺桂最低。干生物量从大到小依次为大叶樟、红椿、桤木、樟树、香椿、天竺桂、油樟。大叶樟的根生物量显著高于其他树种(P<0.05),最低的是香椿。

Figure 2. Biomass of different tree species
如图3所示:不同树种叶、枝、干、根生物量占全株生物量占比分别为4%~10%、14%~22%、39%~59%、18%~38%。叶生物量占比从大到小依次为天竺桂、桤木、油樟、樟树、大叶樟、香椿、红椿;大叶樟、桤木、香椿和红椿的枝生物量占比显著高于其他树种(P<0.05),天竺桂最小;树干生物量占比从大到小依次为红椿、香椿、桤木、大叶樟、樟树、油樟、天竺桂;根生物量占比表现为天竺桂最高,其次是油樟和樟树,桤木、香椿和红椿最低。不同树种根冠比为0.22~0.62,从大到小依次为天竺桂、油樟、樟树、大叶樟、桤木、红椿、香椿。
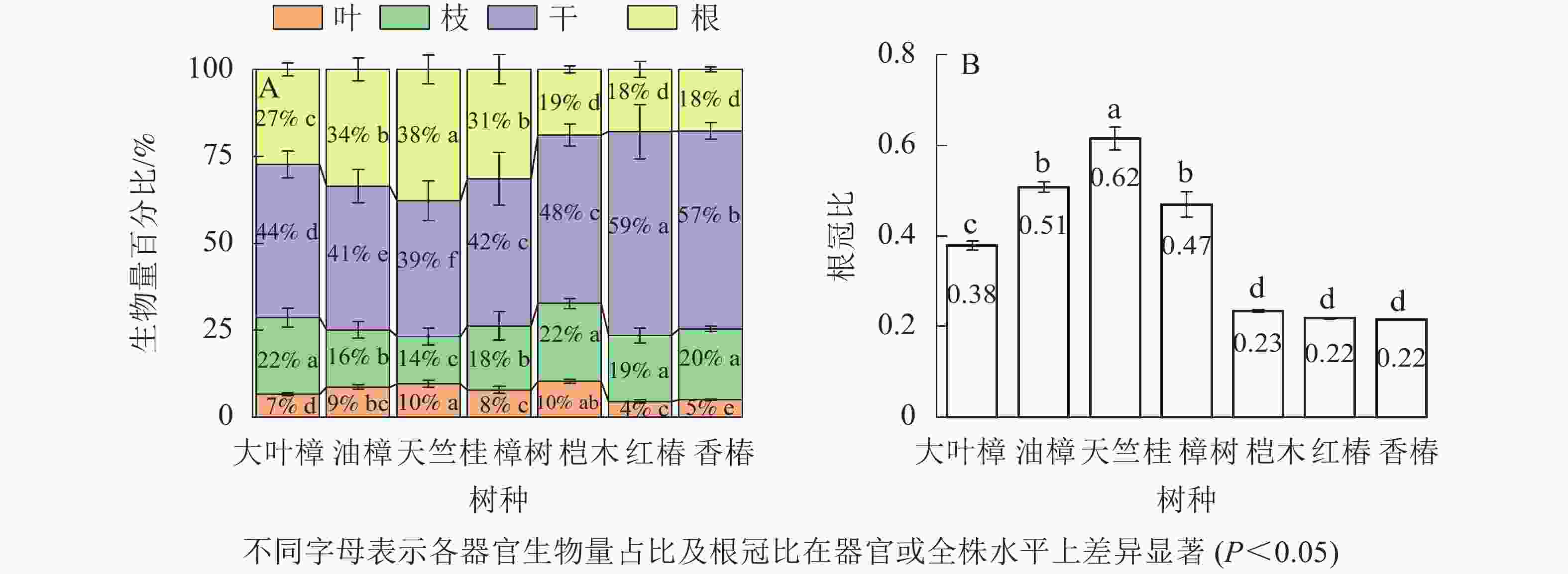
Figure 3. Biomass ratios of organs and root-shoot ratio in different tree species
由图4所示:常绿和落叶树种的全株、叶、枝及干生物量无显著差异(P>0.05),但常绿树种的根生物量显著高于落叶树种。除叶生物量占比外,常绿和落叶树种各器官生物量占总生物量比例存在显著差异(P<0.05),例如常绿树种的根生物量占比和根冠比显著高于落叶树种(P<0.05),而枝和树干生物量占比显著低于落叶树种(P<0.05)。
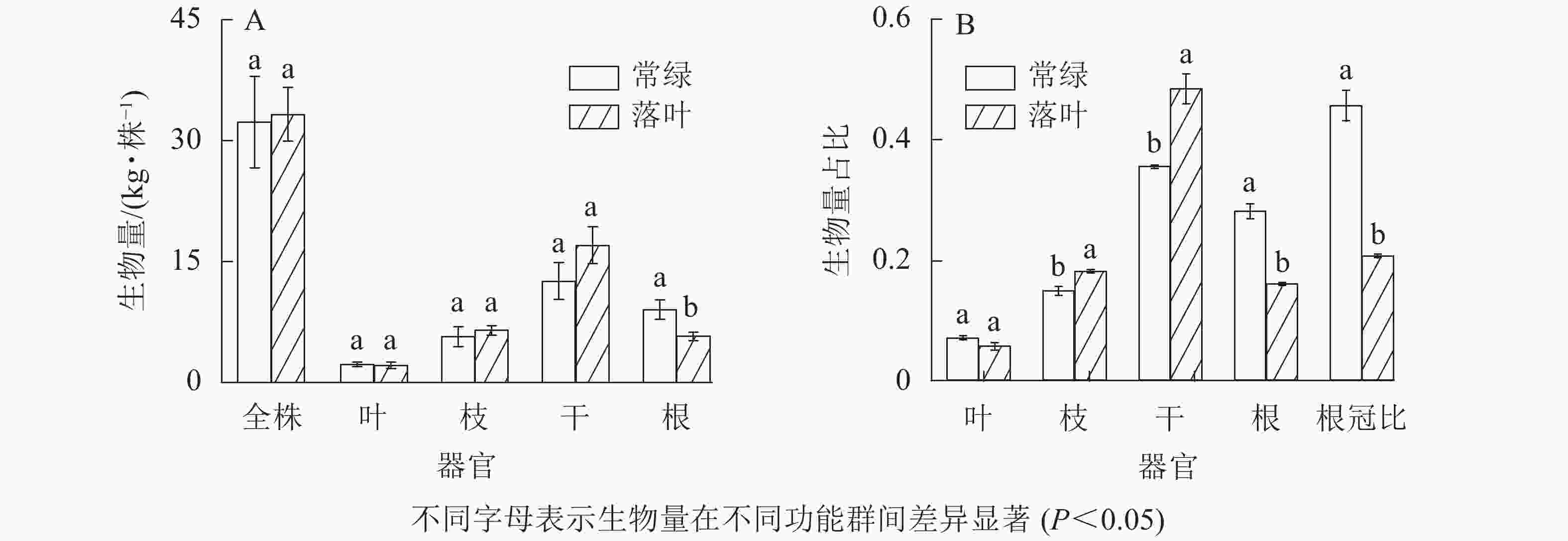
Figure 4. Biomass, biomass proportion and root-shoot ratio of each organs in different functional groups
-
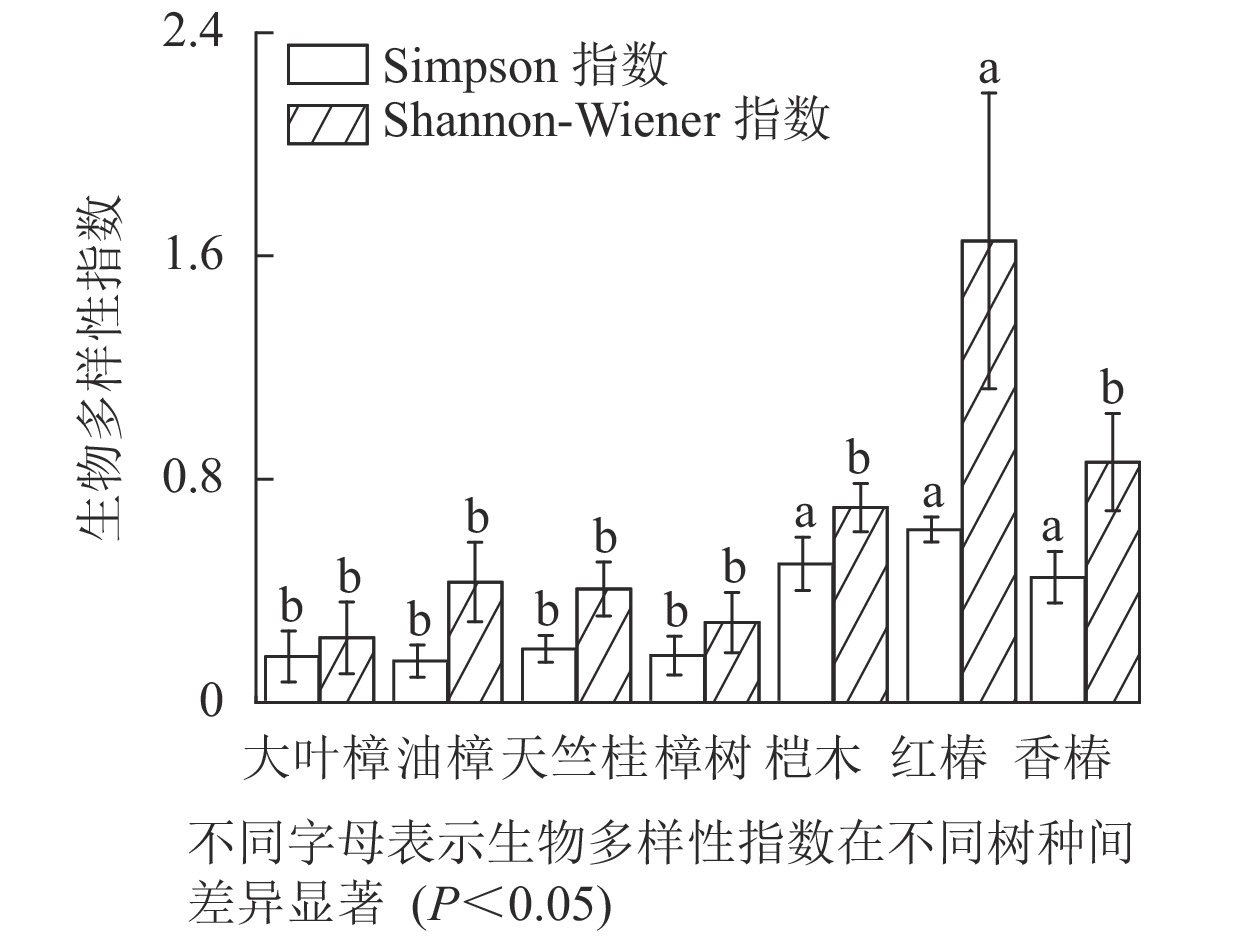
如图5所示:不同树种林分的林下草本多样性存在显著差异,红椿、桤木和香椿林分的林下草本的Simpson多样性指数显著高于天竺桂、大叶樟、樟树和油樟林分(P<0.05)。红椿林分的林下草本的Shannon-Wiener多样性指数显著高于其他树种林分(P<0.05),并且香椿、桤木、油樟、天竺桂、樟树和大叶樟林分之间无显著差异。
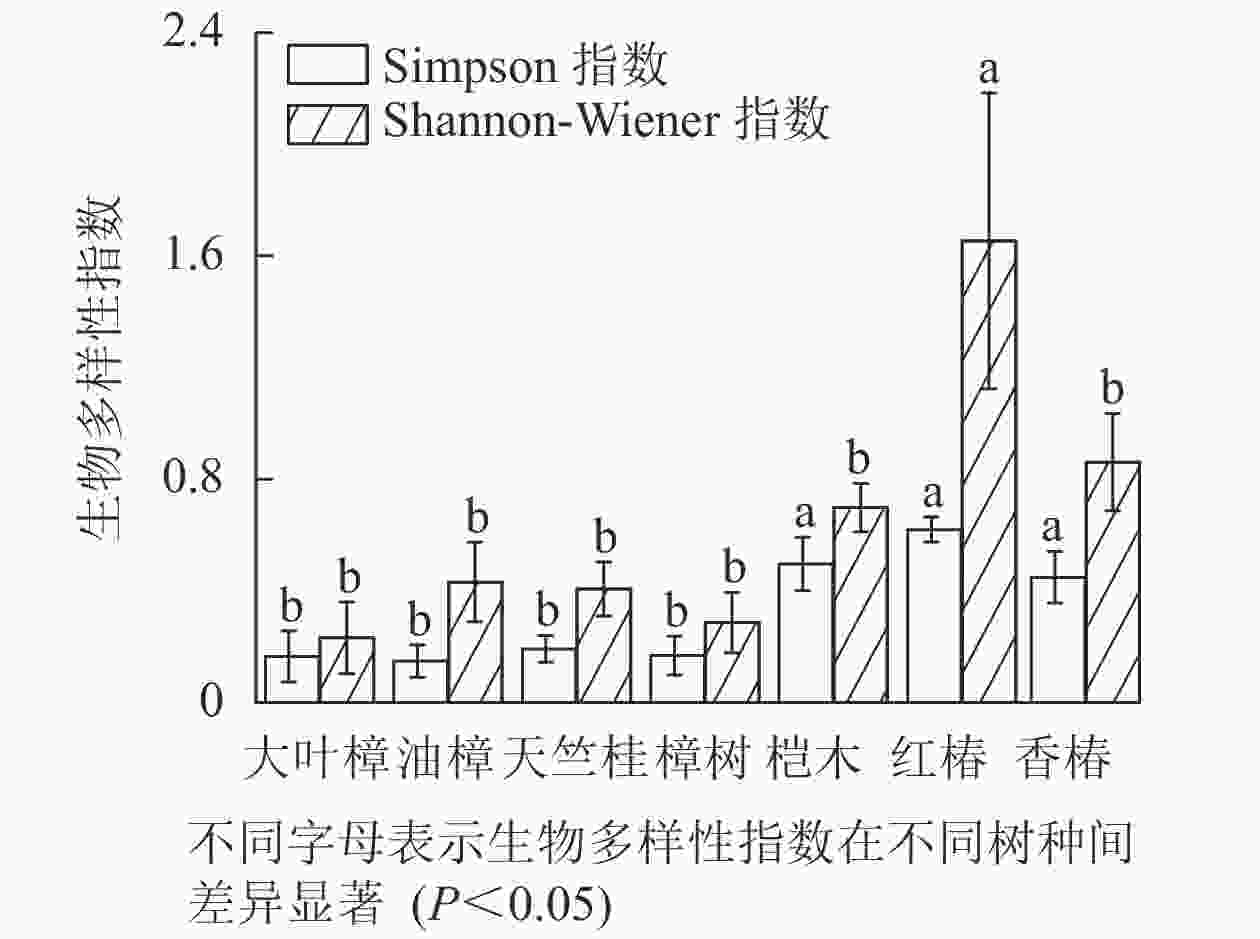
Figure 5. Diversity of understory herbaceous plants in different tree species stands
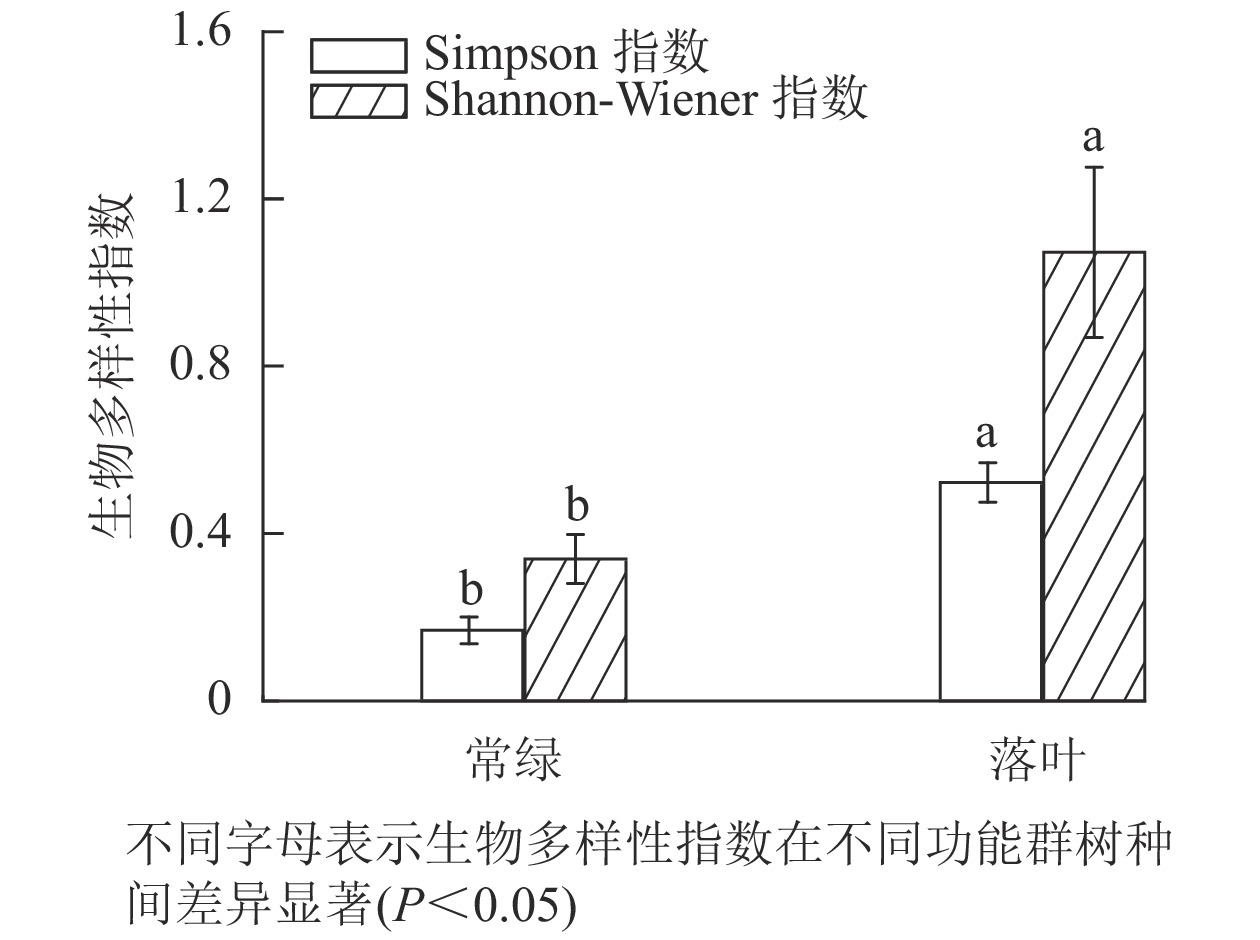
不同功能群树种林分的林下植被多样性存在显著差异,落叶树种林分的林下草本层Simpson多样性指数和Shannon-Wiener多样性指数均显著高于常绿树种林分(图6)。
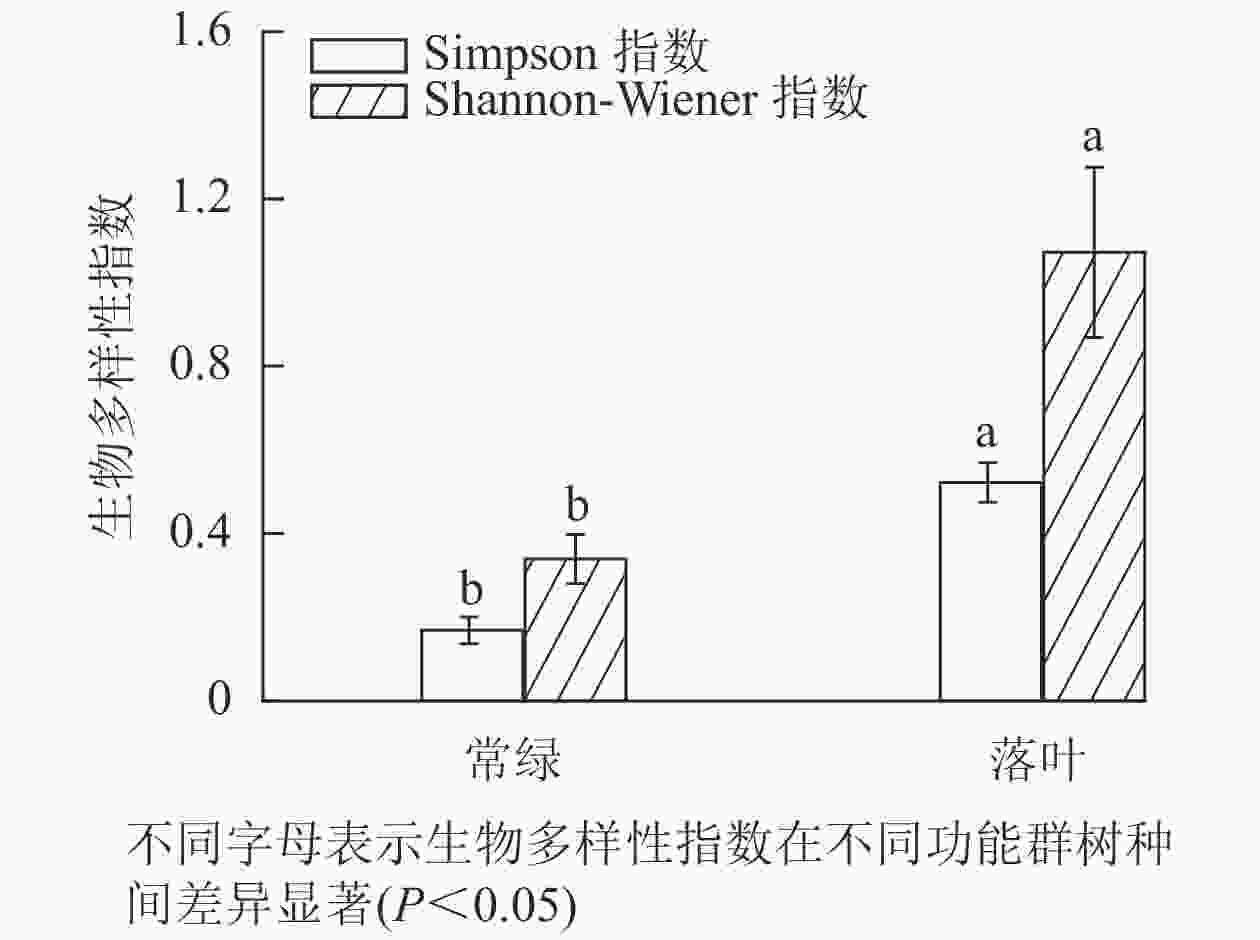
Figure 6. Diversity of understory herbaceous plants in different functional groups
-
生物量是研究物质循环、能量流动、全球气候变化以及碳平衡的基础,乔木生物量的空间分配状况也反映了植物对光、水和养分的利用效率[11]。本研究表明:樟属Cinnamomum植物(大叶樟、樟树、油樟和天竺桂)的平均林分总生物量为80.76 t·hm−2,低于姚迎九等[25]研究的樟树林总生物量(111.08 t·hm−2),而桤木林总生物量为103.37 t·hm−2,高于李贵祥等[26]研究的高原桤木人工林总生物量(23.22 t·hm−2),并且香椿属Toona植物(香椿和红椿)的平均林分总生物量为146.2 t·hm−2,高于王延茹等[29]研究的香椿生物量结果。这可能是由于试验地区降水、海拔、土壤、造林密度以及林龄等的差异引起的,本研究位于成都平原,降水充沛、光照充足,土壤在造林前为农耕地,养分含量丰富,利于林木的生长。本研究发现:不同树种之间的生物量存在显著差异,从大到小依次为大叶樟、桤木、红椿、樟树、油樟、香椿、天竺桂,与葛露露等[13]研究结果一致。这可能是遗传特性差异不同树种对环境的选择和适应不同,进而导致其生长发育能力也不同。且不同树种因其生态习性、生长特性和林分自疏强度不同导致乔木层生物量差异,并且随着生长时间的延长,这种差异越明显[13]。树种各器官生物量分配存在显著差异,从大到小依次为干、根和枝、叶,这与部分学者[30-32]的研究结果相似。这可能是因为干是树木的主体部分,具有运输养分和支撑的重要作用,植物光合作用固定的碳主要储存在干中,形成木质部,并且随林龄增长,干生物量占比会不断增加,而枝、叶生物量比例下降。
本研究发现落叶与常绿树种的全株生物量无显著差异,与FAYOLLE等[33]的研究结果不同,可能是由于调查的树种、数量和试验方式(同质园试验)不同导致的。然而,本研究发现:常绿树种的根生物量占比和根冠比显著高于落叶树种,枝和干生物量占比显著低于落叶树种。这可能是因为本研究的常绿树种凋落物质量(如叶片厚度、质地、化学性质等)相较于落叶树种较差,分解缓慢[34],导致土壤肥力低于落叶树种,因此,需要加大对根生物量的投入,增强对土壤养分的吸收能力。也可能与植物功能群落养分利用策略不同有关,常绿树种属于保守型养分利用策略,其根系发达,能在贫瘠土壤环境下生长发育,而落叶树种属于获取型养分利用策略,其根吸收的养分更多地投入地上部分的生长[35]。
-
高艳鹏等[14]研究发现:刺槐Robinia pseudoacacia、油松纯林的多样性指数高于榆树Ulmus pumila和侧柏Platycladus orientalis纯林,说明不同树种林分的林下植被多样性差异显著,这一结果与本研究结果一致。本研究结果表明:红椿、桤木和香椿林分的林下草本Simpson多样性指数显著高于天竺桂、樟树、大叶樟和油樟林分,而红椿林分林下草本的Shannon-Wiener多样性指数显著高于其他6个树种林分。这可能是因为不同树种可以通过冠幅和郁闭度调控林地的微环境(如光照、水分等),进而有利于其他植物的入侵和生长[23, 36]。如本研究发现:郁闭度较小的树种林下植被群落逐渐出现构树、茜草和空心莲子草等繁殖能力强,喜光耐寒的植物。以往研究表明:林下植被多样性较高有利于促进森林凋落物分解[6-7]。因此,本研究结果说明红椿相较于其他树种可能更有利于促进养分循环,进而改善土壤肥力。
本研究结果表明:落叶树种林分的林下草本Simpson多样性指数和Shannon-Wiener多样性指数均显著高于常绿树种林分,与MESTRE等[15]的研究结果相同。可能存在以下原因:①郁闭度不同导致林下微环境存在差异。本研究中常绿树种林分郁闭度(0.73~0.90)显著高于落叶树种林分(0.66~0.67),常绿树种林分由于郁闭度较高,林下光照不足,水分被截留等原因,林下植被可能难以生长。②土壤肥力影响林下植被生长。常绿树种的凋落物质量相较于落叶树种较差,凋落物分解和养分归还较慢,从而导致土壤肥力较差。不利于林下植被的生长发育。③常绿树种释放化感物质抑制林下植被的生长及养分的循环。如陈洪等[37]研究发现:樟树的凋落叶分解对林下植被生长发育有抑制作用。
-
本研究通过调查亚热带同质园中不同树种林分的生物量和林下植被多样性,发现不同树种全株生物量差异显著,大叶樟的生物量最高,其次是桤木和红椿,天竺桂最低;不同树种各器官生物量差异显著,整体从大到小依次为干、根、枝、叶;除根生物量外,不同功能群各器官生物量均无显著差异;然而,常绿和落叶树种各器官生物量占总生物量比例差异显著,常绿树种的根生物量占比和根冠比显著高于落叶树种,枝和干生物量占比则相反;落叶树种林分的林下植被多样性显著高于常绿树种林分,红椿、桤木和香椿林分的林下草本Simpson指数显著高于天竺桂、樟树、大叶樟和油樟林分,红椿林分的林下草本Shannon-Wiener指数显著高于其他6个树种林分。因此,该地区培育落叶树种更有利于人工林的物质循环和生物多样性保育,并且选择桤木和红椿作为培育树种,更有利于亚热带人工林的可持续经营与管理。
Difference of biomass and understory vegetation diversity among different subtropical plantations in common gardens
doi: 10.11833/j.issn.2095-0756.20210562
- Received Date: 2021-08-13
- Accepted Date: 2021-12-16
- Rev Recd Date: 2021-12-10
- Available Online: 2022-07-20
- Publish Date: 2022-08-20
-
Key words:
- subtropical plantation /
- common garden /
- broad-leaved tree species /
- biomass /
- understory vegetation diversity
Abstract:
| Citation: | LIU Xuan, XIAO Sa, ZHU Peng, et al. Difference of biomass and understory vegetation diversity among different subtropical plantations in common gardens[J]. Journal of Zhejiang A&F University, 2022, 39(4): 717-726. DOI: 10.11833/j.issn.2095-0756.20210562

|



 DownLoad:
DownLoad: