



-
中国林木的改良计划始于20世纪70年代,通过种源选择及引种试验提高林木的生长进程,改良林木材积与品质[1]。为了评估树木改良的潜在效应,需要掌握生物量分配是如何影响树木生长,为应对不断变化的环境条件,选择优质速生的优良基因型,以提高森林的生物量及碳封存能力[2-6]。生物量的分配模式决定该地区林木从地下汲取营养成分的方式,目前国内外均已开展相关研究。如JOHNSEN等[7]认为北部产地的黑云杉Picea mariana根部生物量比南方产地获得更多的分配比例;OLEKSYN等[8]研究12年生樟子松Pinus sylvestris生物量分配规律,表明北部、中部和南部种源根系生物量分别占总生物量的22%,19%和28%;MAJOR等[9]研究发现32年生黑云杉耐旱家系比不耐旱家系拥有更多的地上生长量,地下生物量更多的分配给树桩,而不是细根;朱航勇等[10]在研究兴安落叶松Larix gmelinii的生物量和碳储量的分配模式的基础上初步选择了生物量和碳储量大的优良种源。研究生物量及其分配模式可明确林木的生长变化规律,估测森林生态系统生物量和净生产力水平,进而指导生产造林[4-5, 11-12]。马尾松Pinus massoniana是松属Pinus植物在中国地理分布最广泛的树种,不仅是荒山绿化、水土保持的先锋树种,而且还是多功效的用材林和具有高附加值林产品的经济林树种,其分布区域位于21°41′~33°56′N,103°~123°E[13]。马尾松宽广的分布区域是开展不同产地生长及生物量等研究的优势条件[3, 14]。过往的研究表明不同产地的马尾松生产力存在显著差异,初步揭示马尾松生长具纬向变异模式[15-17],北部种源年均生长量较少,显著低于南部种源[15]。以往对马尾松林分的生产力和生物量的研究较多[14],而在相同立地条件下对不同种源开展生物量长期积累和分配规律研究较少。为此,选择引种至湖北的广东种源(南部种源)和湖北当地种源(北部种源)开展生产力形成过程研究,揭示不同种源生物量累积模式,阐明马尾松优势种源高生长量及较快生长速率的形成机制,为优质高产马尾松的遗传改良提供科学依据。
-
引种的广东省茂名市高州种源人工林与当地种源人工林均位于湖北省太子山林场管理局石龙林场(30°55′11″N,112°50′29″E),栽植立地类型、经营史相同,1972年栽植,位于同一林分相邻地块,株行距为2.0 m×1.5 m。试验林均利用1年生裸根苗造林,当年造林成活率在95%以上,造林后连续抚育3 a至幼林郁闭。分别在造林后第10年和第15年进行2次间伐,伐去试验林中的被压木,疏伐强度均为30%。海拔为80 m。该地属于亚热带季风湿润性气候,夏季多雨,冬春干旱,年平均气温为16.4 ℃,无霜期为240 d,年平均降水量为1 094.6 mm。立地为黄土壤,土壤厚度55 cm以上。
-
2016年9月,在引种的广东种源人工林和当地湖北种源人工林中各选择2块样地,样地设置25 m × 25 m。测量样地内每株树的树高、胸径及冠幅,按公式V=0.000 062 341 803×D1.31.855 149 7×H0.956 824 92估算活立木单株材积。比较2个种源单位面积的活立木蓄积量(表 1)可知:广东种源人工林单位面积蓄积量高于湖北种源人工林17.13%。每个种源分别选择12株,选择3个胸径DBH范围12 cm≤DBH<18 cm,18 cm≤DBH<26 cm和26 cm≤DBH<34 cm进行树干解析和生物量调查测定。解析木测定参考孟宪宇[18]方法获得各龄阶树高、胸径和材积生长量。采用全树收获法测定林木各组分生物量,解析木自地表伐倒后,测量树高,按2 m截段,分别测量每段的鲜质量。树冠按照长度平均分为3层,将每层枝叶分离并称量,分别取样500~1 000 g带回实验室。围绕树干基部为中心点,在直径3 m范围内挖取样株根系,将样木的粗根(>5 cm)和细根(≤5 cm)挖出。将细根全部挖掘出来在实际操作中不现实,且对根系总生物量的影响不大[11-12]。用凿子和抹布去除根系中的泥沙和土,测定粗根和细根总鲜质量,并取500~ 1 000 g根鲜样,保存在保鲜袋内,带回实验室,用流动水浸泡、漂洗。样品在105 ℃杀青后,在65 ℃下烘干至恒量,称质量并记录。通过烘干前后的质量,计算样品含水率。由样品的含水率和各部分鲜质量计算出各部分以及整株树木的干物质生物量。
表 1 取样林分概况
Table 1. Description of sampling sites
DBH/cm 广东种源人工林(GD) 湖北种源人工林(HB) 树龄/a 树高/m 胸径/cm 株数 树高/m 胸径/cm 株数 [26, 34) 20.74 28.50 24 18.88 28.34 23 44 [18, 26) 18.12 21.72 46 16.91 22.38 39 44 [12, 18) 12.21 14.77 29 11.20 14.57 20 44 -
利用马尾松解析木资料拟合胸径与各器官生物量的相对生长关系,模型为W=a(DBH)d。其中,W为各组分生物量,DBH为胸径,a和d为模型参数。计算决定系数r2和标准估计误差ESE[3]。引用相对差异ER(GD/HB)表示广东种源(GD)与湖北种源(HB)的差值比较[4],计算公式为ER(GD/HB)=(XGD-XHB)/XHB×100%,X代表不同种源人工林的生长量参数和生物量参数的均值。采用SPSS 19.0进行数据分析与曲线图绘制。
-
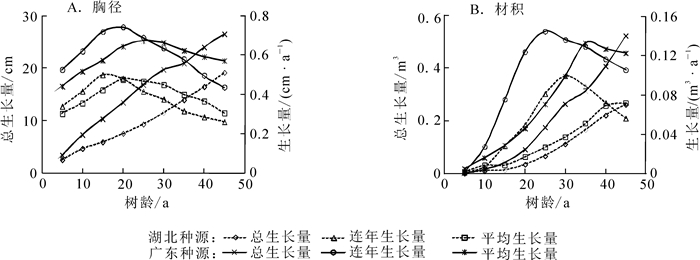
由图 1可看出:马尾松种源整体生长进程趋势较一致,胸径、材积的平均生长量与连年生长量在生长初期均逐年增加,属于快速生长阶段,之后逐年开始下降。但广东马尾松种源胸径和材积的总生长量、年平均生长量及连年生长量在各年龄段均高于湖北马尾松种源。在胸径方面,广东马尾松种源20年生连年生长量达最大值0.74 cm·a-1,26年生平均生长量达最大值0.67 cm·a-1;而湖北种源15年生连年生长量达最大值0.49 cm·a-1,20年生平均生长量达最大值0.44 cm·a-1。由连年生长量可看出:广东马尾松种源具有胸径生长量大,持续期时间长的特点,即使40年生时仍维持在0.40 cm·a-1以上。在材积方面,10年生之前材积生长缓慢,2个种源差异较小。随树龄的增加,广东种源在25年生左右连年生长量达最大值0.015 m3·a-1,35年生平均生长量达最大值0.011 m3·a-1;而湖北种源30年生连年生长量达最大值0.011 m3·a-1,42年生平均生长量达最大值0.007 m3·a-1。由材积的平均生长量与连年生长量的相交位置推断,广东马尾松种源的数量成熟龄到达时间比湖北种源早约10 a。

图 1 广东和湖北种源马尾松的不同年龄阶段生长量
Figure 1. Growth of Pinus massoniana in Guangdong and Hubei provenances at different ages
-
由表 2可知:广东种源人工林生长快速,在不同胸径范围内广东种源的树高、胸径及材积平均值均高于湖北种源人工林。12 cm≤DBH<18 cm,18 cm≤DBH<26 cm和26 cm≤DBH<34 cm 3个胸径范围内,广东种源人工林的整株生物量分别为124.1,238.7和437.5 kg·株-1,湖北种源人工林的整株生物量分别为98.1,167.3和307.4 kg·株-1;广东种源人工林的树干生物量分别为96.5,185.7和346.6 kg·株-1,湖北种源人工林的树干生物量分别为71.7,128.4和245.4 kg·株-1。在枝和叶的生物量上,除12 cm≤DBH<18 cm外,广东种源人工林在其他胸径范围上均高于湖北种源。
表 2 广东和湖北种源人工林生长量与生物量的均值及相对差值
Table 2. Average and relative differences of growth and biomass in Guangdong and Hubei man-made forest provenances
种源 DBH/cm 生长性状 生物量/kg 树高/m 胸径/cm 材积/m3 地上部分 树干 枝 针叶 地下部分 细根 粗根 整株 广东 [26, 34) 21.0 32.2 0.71 394.4 346.6 37.4 10.3 43.1 13.3 29.8 437.5 (1.4) (2.4) (0.16) (21.9) (27.5) (10.8) (1.7) (3.0) (2.2) (2.0) (23.1) [18, 26) 19.1 23.2 0.36 209.2 185.7 18.1 5.4 29.4 8.7 20.7 238.7 (1.7) (2.0) (0.08) (46.8) (44.4) (6.1) (1.6) (3.4) (1.2) (2.4) (48.6) [12, 18) 17.0 16.8 0.20 107.1 96.5 7.6 2.9 17.0 6.9 10.1 124.1 (0.7) (1.6) (0.06) (40.4) (41.4) (3.3) (0.2) (1.7) (1.3) (0.8) (42.1) 湖北 [26, 34) 19.6 29.9 0.57 278.4 245.4 27.0 6.0 28.9 8.7 20.2 307.4 (0.7) (2.5) (0.04) (32.1) (28.5) (2.9) (0.7) (6.5) (2.3) (4.1) (38.6) [18, 26) 16.2 21.5 0.29 149.4 128.4 16.2 4.6 17.9 5.3 12.7 167.3 (1.4) (1.6) (0.10) (49.5) (38.3) (8.9) (3.1) (3.5) (1.7) (2.0) (52.3) [12, 18) 14.7 15.8 0.16 87.1 71.7 11.8 3.5 11.0 3.6 7.4 98.1 (2.1) (2.0) (0.05) (24.3) (28.7) (6.8) (1.6) (3.7) (1.1) (2.6) (27.2) ER(GD/HB) [26, 34) 7.1 7.7 24.6 41.7* 41.2* 38.5 71.7* 49.1* 52.9 47.5* 42.3* [18, 26) 17.9 7.9 24.1 40.0 44.6* 11.7 17.4 64.2* 64.1 62.9* 42.6* [12, 18) 15.6 6.3 25.0 23.0 34.6* -35.6 -17.1 54.5* 91.6 36.5* 26.5 说明:括号内为标准差;*代表P<0.01检验水平上差异显著;ER(GD/HB)代表广东种源和湖北种源的相对差值 通过非参数检验可知:12 cm≤DBH<18 cm,18 cm≤DBH<26 cm和26 cm≤DBH<34 cm 3个胸径范围内,2个种源人工林在地下部分生物量和粗根生物量均存在显著差异(P<0.05),地下部分生物量ER(GD/HB)分别为54.5%,64.2%和49.1%,粗根的ER(GD/HB)分别为36.5%,62.9%和47.5%,而细根间生物量未达显著差异。除此之外,26 cm≤DBH<34 cm范围内,2个种源人工林在整株生物量、地上部分生物量、树干和针叶生物量差异显著;18 cm≤DBH<26 cm范围内,整株生物量和树干生物量差异显著。
-
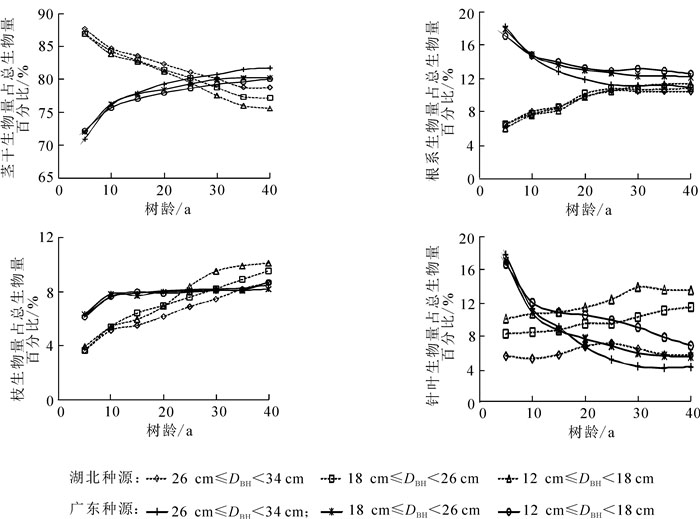
马尾松各部位生物量分配比例差别较大(表 3),总生物量的88.5%分配给了地上部分,其中,77.2%分配给树干,8.7%分配给枝,2.6%分配给针叶;11.5%分配给了地下部分。在根级别中,67.8%的根生物量分配给粗根,32.2%的根系生物量分配给细根。由表 3可知:2个种源人工林的树干生物量和地上部分生物量分配比例均随胸径范围的增加而增加,而地下部分生物量、树冠(树枝和针叶)生物量和细根生物量分配比例随胸径的增加而降低。随胸径范围的增加,湖北种源粗根系占总生物量比例逐渐减少,而广东种源呈先增加后减少的规律。广东种源较湖北种源分配较多比例的生物量给树干,随胸径范围的增加,树干分配比例相对差值分别为0.9%,1.4%和6.4%;而分配较少比例的生物量给树枝,相对差值分别为-49.1%,-21.7%,-2.7%,胸径范围为12 cm≤DBH<18 cm的树枝生物量分配比例差异最大。广东种源地上部分生物量分配比例少于湖北种源,随胸径的增加,相对差值分别为-2.8%,-1.8%和-0.5%;而地下部分(包括细根和粗根)被分配了较多比例的生物量,随胸径范围的增加,广东种源地下部分生物量分配比例比湖北种源分别高22.2%,15.2%,14.8%。由图 2可知:广东种源胸径和生物量较大,回归斜率显示广东种源(0.116 6)高于湖北种源(0.099),表明广东种源倾向分配更多的生物量给地下部分[19]。
表 3 各器官生物量占总生物量的百分比
Table 3. Biomass percentage of each organ in total biomass
生物量 百分比平均值/% 广东种源占总生物量的百分比/% ER(GD/HB)/% [26, 34) [18, 26) [12, 18) [26, 34) [18, 26) [12, 18) cm 各部位 地上部分 88.5 89.6 89.3 88.8 -0.5 -1.8 -2.8 树干 77.2 78.8 76.8 73.1 0.9 1.4 6.4 枝 8.7 8.8 9.7 12.0 -2.7 -21.7 -49.1 针叶 2.6 2.0 2.8 3.6 20.6 -17.7 -34.5 地下部分 11.5 10.4 10.7 11.2 14.8 15.2 22.2 细根 3.7 3.2 3.2 3.7 7.4 15.1 51.5 粗根 7.8 7.2 7.6 7.6 3.7 14.3 7.9 不同冠层分枝 上层枝 12.5 18.5 9.9 11.9 -37.9 -10.5 22.0 中层枝 40.1 48.1 29.0 39.0 -1.2 65.7 -25.7 下层枝 47.4 33.0 61.1 55.1 22.5 -30.4 0.3 不同冠层针叶 上层叶 25.9 48.3 21.7 25.7 -49.8 2.2 20.7 中层叶 45.3 38.3 43.5 48.6 29.2 6.5 56.2 下层叶 28.8 13.3 37.0 25.7 89.3 -14.8 7.3 说明:ER(GD/HB)代表广东种源和湖北种源的相对差值 
图 2 马尾松地下生物量与地上生物量的线性关系
Figure 2. Biomass linear relationship between underground and aboveground of Pinus massoniana
-
由表 4可知:马尾松各器官生物量基于胸径的生长方程拟合精度较高,可用于分析2个种源长期生长过程中生物量分配规律(图 3)。研究表明:在3个胸径范围内,湖北种源地下生物量分配比例在整个生长过程中均低于广东种源;在生长初期,湖北种源树干所占总生物量比例高于广东种源,而枝和叶的生物量分配比例低于广东种源;随着树龄增长,广东种源树干所占总生物量比例逐渐高于湖北种源,并趋于稳定;而枝、叶正好相反。说明广东种源在幼林期将更多的生物量分配给枝、叶和地下根系,来竞争有限的营养和光照资源,进而迅速生长积累更多的有机物质[20]。
表 4 马尾松广东和湖北种源生物量模型
Table 4. Parameter estimats of Richards model on Guangdong and Hubei provenance of Pinus massoniana
生物量 a d r2 ESE 湖北 广东 湖北 广东 湖北 广东 湖北 广东 树干 0.234 1 0.075 8 2.046 7 2.468 6 0.871 6 0.878 7 3.018 2.968 枝 0.006 4 0.007 6 2.573 8 2.471 3 0.790 2 0.833 2 1.507 1.611 叶 0.006 9 0.005 9 2.137 1 2.124 1 0.825 8 0.808 4 2.216 2.329 地上部分 0.279 0 0.163 3 2.049 9 2.255 0 0.834 7 0.873 0 5.828 5.926 细根 0.005 9 0.016 2 2.285 1 1.959 6 0.842 5 0.851 6 0.715 0.698 粗根 0.006 2 0.013 2 2.532 6 2.173 2 0.870 1 0.877 0 1.156 1.145 地下部分 0.014 0 0.029 1 2.381 8 2.084 6 0.867 1 0.935 5 1.823 1.685 说明:a和d为模型参数,r2为决定系数,ESE为标准估计误差 
图 3 各器官生物量在不同生长阶段占总生物量的百分比
Figure 3. Biomass percentage of each organ in total biomass at different growth stages
-
研究表明引种到湖北的广东种源生长性状及生物量均表现出快速生长的特点。由材积的平均生长量与连年生长量的相交位置推断,广东马尾松种源的数量成熟龄到达时间比湖北种源早约10 a,同刘青华等[15]得出的南部种源数量成熟早于中部和北部种源结论一致。湖北种源的数量成熟龄虽晚于广东种源,但每年的生长量维持在较低的水平。从幼龄期到主伐期间的不同生长发育阶段,马尾松南北种源间的胸径和材积的生长进程及生长量存在较大生长差异。由此可见,引种优良种源对于提高马尾松生产力的重要性。马尾松是喜光树种,生物量受到树高和胸径增长的影响,为竞争有限的营养和光照资源,通过调整分枝、针叶或根分布,加快高生长和干物质量积累,生长迅速的树木吸收更多的阳光进行光合作用,在地面上积累更多的有机物质,而当其达到主林层时,树冠更多的向水平方向生长,扩大冠面积[20]。广东和湖北种源的树干所占总生物量比例均随胸径的增加而增加,而树冠(树枝和针叶)生物量分配比例随胸径的增加而降低,这同以往的研究结论一致[21]。本研究中,随胸径的增加,广东种源人工林在地上部分及地下部分生物量均高于湖北种源。在大径材(26 cm≤DBH<34 cm)时广东种源与湖北种源人工林在地上部分、树干和针叶的生物量差值更大。表明快速生长种源的选择可增加总生物量,改变人工林的生产力大小。因此,栽植快速成长的种源将增加人工林生物量,进而增加固碳能力[7, 9, 15, 21-22]。
树木生物量分配随树体大小而变化,生长选择对树木生物量累积的影响很重要,通常用它们的生长特性(函数)来预测生物量分配利用的异速生长关系,可明确较高生长量及生长速率的形成机制[5, 23]。研究马尾松2个种源幼龄至成熟龄不同发育阶段的异速生长关系表明:生长前期的树木生物量分配变化幅度较大。生长迅速的广东种源在生长初期分配更多比例的生物量给树冠和地下根系,而树干生物量分配比例低于湖北种源;随着树龄增长,广东种源树干所占总生物量比例逐渐高于湖北种源,而树冠生物量的分配比例逐渐低于湖北种源,且趋于稳定。同BONGARTEN等[24]得出生长迅速的树木与生长缓慢的树木相比,倾向于将较少比例的生物量分配给针叶和分枝的结论类似。马尾松树干生物量平均值占总生物量平均值的77.2%,是整个树木的支撑。中龄林后,广东种源的树干生物量占总生物量比例高于湖北种源,随树龄的增加,2个源树干生物量分配比例差异逐渐减小。研究结果同XIANG等[3]研究不同产地的马尾松人工林生物量分配规律一致,即生长前期的树木生物量分配变化幅度较大,成熟龄树木主要是树干生物量保持了稳定的分配比例。ZHU等[4]和WANG等[5]分别利用不同的云杉Picea aspoerata和白桦Betula platyphlla种源同样揭示了生长快慢种源的生物量分配格局,认为生长快的种源分配较多的生物量给树干,使树干具有更高的生产力和固碳能力,进而选择出优良固碳种源。
本研究马尾松人工林地上部分生物量占总生物量的88.5%,地下部分生物量占11.5%,这与丁贵杰等[14]研究已达轮伐期的马尾松人工林的生物量分配相类似。研究表明:生长较快的树木,不仅将较高比例的生物量分配给树干,也会将更多的生物量分配给根[19]。引种的广东种源较湖北种源分配较多比例的生物量给树干,但地上部分总生物量分配比例少于湖北种源,相对差值分别为-2.8%(小树)、-1.8%(中等树)和-0.5%(大树),这部分差异是因为地下部分被分配了较多比例的生物量而引起的[19, 24]。不同的胸径范围内,2个种源的粗根生物量均存在显著差异,而细根的生物量差异不显著,说明在根层面上,广东种源相对于湖北种源,倾向分配更多的生物量给地下部分,种源间粗根生物量存在更大的差异。
Biomass allocation differences with Pinus massoniana in Guangdong and Hubei provenances
-
摘要: 采用解析木法与整株收获法分析已达轮伐期的马尾松广东种源和湖北当地种源的生长进程及地上、地下生物量分配格局,分析生长快速与缓慢种源人工林生长过程及生产力形成过程中生物量分配模式,探讨2个种源的长期适应性差异的机制。根据胸径(DBH)大小将解析木划分为3个等级:12 cm≤DBH < 18 cm,18 cm≤DBH < 26 cm和26 cm≤DBH < 34 cm。结果如下:①广东种源在25年生连年生长量达最大值0.015 m3·a-1,35年生平均生长量达最大值0.011 m3·a-1,湖北种源30年生时连年生长量达最大值0.011 m3·a-1,42年生时平均生长量达最大值0.007 1 m3·a-1。②广东种源分配较多比例的生物量给地下部分,2个种源的粗根生物量存在显著差异(P < 0.05),随胸径增加,相对差异(RE,GD/HB)分别为36.5%,62.9%和47.5%,而细根间生物量未达显著性差异。在胸径范围2 634 cm内,2个种源人工林在整株生物量、地上部分生物量、树干和针叶生物量差异显著(P < 0.05)。③湖北种源地下生物量分配比例在整个生长过程中均低于广东种源;在生长初期,湖北种源树干所占总生物量比例高于广东种源,而枝和叶的生物量分配比例低于广东种源;随着树龄增长,广东种源树干所占总生物量比例逐渐高于湖北种源,并趋于稳定;而枝、叶正好相反。因此,引种的广东种源生长性状及树干生物量均表现出快速增长的特点,数量成熟年龄较湖北种源提前,栽植快速成长的种源将增加人工林生物量。Abstract: To analyze biomass allocation in the process of productivity formation in man-made forests of fast- and slow-growing provenances, Masson pine (Pinus massoniana), an important man-made forest species in south China, was used. Different mechanisms of long-term adaptation in the two provenance types were also discussed. The allocation patterns of above- and below-ground biomass in Guangdong and Hubei provenances were analyzed by using the whole plant harvest method. According to differing diameter at breast heights (DBH), the analytic trees were grouped into three classes:12 cm ≤ DBH < 18 cm, 18 cm ≤ DBH < 26 cm, and 26 cm ≤ DBH < 34 cm. Results showed that (1) The maximum of annual growth of Guangdong provenance reached 0.015 m3·a-1 in 25 years and the average annual growth of Guangdong provenance reached 0.011 m3·a-1 in 30 years. In addition, the maximum of annual growth of Hubei provenance reached 0.011 and 0.007 m3·a-1 in 30 and 42 years, respectively. (2) A large proportion of biomass was distributed to the underground part in Guangdong provenance. There were significant differences in the crude root biomass of the two provenances (P < 0.05). With the increasing of chest diameter, the relative difference were 36.5%, 62.9% and 47.5%, respectively, while there was no significant difference in the biomass between the fine roots. In the DBH range from 26 to 34 cm, there were significant differences among the biomass of the whole plant, the aboveground part, the tree trunk and the conifer (P < 0.05). (3) The biomass allocation proportion of Hubei provenance for the whole growth process in the 3 DBH classes was lower than that of the Guangdong provenance. At the initial stage of growth, the total biomass of the trunk in the Hubei provenance was higher than that in the Guangdong seed source; whereas, the biomass allocation proportions of branches and leaves was lower. As tree age increased, the proportion of total biomass for the tree trunk in the Guangdong provenance was gradually higher than that in the Hubei provenance and tended to be stable. However, the results were opposite in the branches and leaves. Therefore, the growth characters and the biomass of tree trunks showed the characteristic of rapid growth in Guangdong provenance, while quantitative maturity age was earlier than that in Hubei provenance.
-
Key words:
- forest tree breeding /
- Pinus massoniana /
- biomass /
- growth process /
- allocation patterns
-
图 1 广东和湖北种源马尾松的不同年龄阶段生长量
Figure 1 Growth of Pinus massoniana in Guangdong and Hubei provenances at different ages
图 2 马尾松地下生物量与地上生物量的线性关系
Figure 2 Biomass linear relationship between underground and aboveground of Pinus massoniana
图 3 各器官生物量在不同生长阶段占总生物量的百分比
Figure 3 Biomass percentage of each organ in total biomass at different growth stages
表 1 取样林分概况
Table 1. Description of sampling sites
DBH/cm 广东种源人工林(GD) 湖北种源人工林(HB) 树龄/a 树高/m 胸径/cm 株数 树高/m 胸径/cm 株数 [26, 34) 20.74 28.50 24 18.88 28.34 23 44 [18, 26) 18.12 21.72 46 16.91 22.38 39 44 [12, 18) 12.21 14.77 29 11.20 14.57 20 44  下载: 导出CSV
下载: 导出CSV
表 2 广东和湖北种源人工林生长量与生物量的均值及相对差值
Table 2. Average and relative differences of growth and biomass in Guangdong and Hubei man-made forest provenances
种源 DBH/cm 生长性状 生物量/kg 树高/m 胸径/cm 材积/m3 地上部分 树干 枝 针叶 地下部分 细根 粗根 整株 广东 [26, 34) 21.0 32.2 0.71 394.4 346.6 37.4 10.3 43.1 13.3 29.8 437.5 (1.4) (2.4) (0.16) (21.9) (27.5) (10.8) (1.7) (3.0) (2.2) (2.0) (23.1) [18, 26) 19.1 23.2 0.36 209.2 185.7 18.1 5.4 29.4 8.7 20.7 238.7 (1.7) (2.0) (0.08) (46.8) (44.4) (6.1) (1.6) (3.4) (1.2) (2.4) (48.6) [12, 18) 17.0 16.8 0.20 107.1 96.5 7.6 2.9 17.0 6.9 10.1 124.1 (0.7) (1.6) (0.06) (40.4) (41.4) (3.3) (0.2) (1.7) (1.3) (0.8) (42.1) 湖北 [26, 34) 19.6 29.9 0.57 278.4 245.4 27.0 6.0 28.9 8.7 20.2 307.4 (0.7) (2.5) (0.04) (32.1) (28.5) (2.9) (0.7) (6.5) (2.3) (4.1) (38.6) [18, 26) 16.2 21.5 0.29 149.4 128.4 16.2 4.6 17.9 5.3 12.7 167.3 (1.4) (1.6) (0.10) (49.5) (38.3) (8.9) (3.1) (3.5) (1.7) (2.0) (52.3) [12, 18) 14.7 15.8 0.16 87.1 71.7 11.8 3.5 11.0 3.6 7.4 98.1 (2.1) (2.0) (0.05) (24.3) (28.7) (6.8) (1.6) (3.7) (1.1) (2.6) (27.2) ER(GD/HB) [26, 34) 7.1 7.7 24.6 41.7* 41.2* 38.5 71.7* 49.1* 52.9 47.5* 42.3* [18, 26) 17.9 7.9 24.1 40.0 44.6* 11.7 17.4 64.2* 64.1 62.9* 42.6* [12, 18) 15.6 6.3 25.0 23.0 34.6* -35.6 -17.1 54.5* 91.6 36.5* 26.5 说明:括号内为标准差;*代表P<0.01检验水平上差异显著;ER(GD/HB)代表广东种源和湖北种源的相对差值
下载: 导出CSV
表 3 各器官生物量占总生物量的百分比
Table 3. Biomass percentage of each organ in total biomass
生物量 百分比平均值/% 广东种源占总生物量的百分比/% ER(GD/HB)/% [26, 34) [18, 26) [12, 18) [26, 34) [18, 26) [12, 18) cm 各部位 地上部分 88.5 89.6 89.3 88.8 -0.5 -1.8 -2.8 树干 77.2 78.8 76.8 73.1 0.9 1.4 6.4 枝 8.7 8.8 9.7 12.0 -2.7 -21.7 -49.1 针叶 2.6 2.0 2.8 3.6 20.6 -17.7 -34.5 地下部分 11.5 10.4 10.7 11.2 14.8 15.2 22.2 细根 3.7 3.2 3.2 3.7 7.4 15.1 51.5 粗根 7.8 7.2 7.6 7.6 3.7 14.3 7.9 不同冠层分枝 上层枝 12.5 18.5 9.9 11.9 -37.9 -10.5 22.0 中层枝 40.1 48.1 29.0 39.0 -1.2 65.7 -25.7 下层枝 47.4 33.0 61.1 55.1 22.5 -30.4 0.3 不同冠层针叶 上层叶 25.9 48.3 21.7 25.7 -49.8 2.2 20.7 中层叶 45.3 38.3 43.5 48.6 29.2 6.5 56.2 下层叶 28.8 13.3 37.0 25.7 89.3 -14.8 7.3 说明:ER(GD/HB)代表广东种源和湖北种源的相对差值
下载: 导出CSV
表 4 马尾松广东和湖北种源生物量模型
Table 4. Parameter estimats of Richards model on Guangdong and Hubei provenance of Pinus massoniana
生物量 a d r2 ESE 湖北 广东 湖北 广东 湖北 广东 湖北 广东 树干 0.234 1 0.075 8 2.046 7 2.468 6 0.871 6 0.878 7 3.018 2.968 枝 0.006 4 0.007 6 2.573 8 2.471 3 0.790 2 0.833 2 1.507 1.611 叶 0.006 9 0.005 9 2.137 1 2.124 1 0.825 8 0.808 4 2.216 2.329 地上部分 0.279 0 0.163 3 2.049 9 2.255 0 0.834 7 0.873 0 5.828 5.926 细根 0.005 9 0.016 2 2.285 1 1.959 6 0.842 5 0.851 6 0.715 0.698 粗根 0.006 2 0.013 2 2.532 6 2.173 2 0.870 1 0.877 0 1.156 1.145 地下部分 0.014 0 0.029 1 2.381 8 2.084 6 0.867 1 0.935 5 1.823 1.685 说明:a和d为模型参数,r2为决定系数,ESE为标准估计误差
下载: 导出CSV
-
[1] 雷泽勇, 韩艳刚, 赵国军, 等.辽宁章古台樟子松生长过程分析[J].浙江农林大学学报, 2018, 35(2):324-330. LEI Zeyong, HAN Yangang, ZHAO Guojun, et al. Growth of Mongolian pine in Zhanggutai of Liaoning Province[J]. J Zhejiang A&F Univ, 2018, 35(2):324-330. [2] 王霓虹, 高萌, 李丹.长白落叶松人工林乔木层生物量分布特征及其固碳能力研究[J].植物研究, 2014, 34(4):554-560. WANG Nihong, GAO Meng, LI Dan. Tree layer biomass distribution and carbon storage capacity of Larix olgensis plantation[J]. Bull Bot Res, 2014, 34(4):554-560. [3] XIANG Wenhua, LIU Shaohui, DENG Xiangwen, et al. General allometric equations and biomass allocation of Pinus massoniana trees on a regional scale in southern China[J]. Ecol Res, 2011, 26(4):699-711. [4] ZHU H Y, WENG Y H, ZHANG H G, et al. Comparing fast- and slow-growing provenances of Picea koraiensis in biomass, carbon parameters and their relationships with growth[J]. For Ecol Manage, 2013, 307(6):178-185. [5] WANG X W, WENG Y H, LIU G F, et al. Variations in carbon concentration, sequestration and partitioning among Betula platyphylla provenances[J]. For Ecol Manage, 2015, 358:344-352. [6] GERSONDE R F, O'HARA K L. Comparative tree growth efficiency in Sierra Nevada mixed-conifer forests [J]. For Ecol Manage, 2005, 219(1):95-108. [7] JOHNSEN K H, SEILER J R, MAJOR J E. Growth shoot phenology and physiology of diverse seed sources of black spruce. Ⅱ. 23-year-old field trees[J]. Tree Physiol, 1996, 16:375-380. [8] OLEKSYN J, REICH P B, CHALUPKA W, et al. Differential above- and below-ground biomass accumulation of European Pinus sylvestris populations in a 12-year-old provenance experiment[J]. Scand J For Res, 1999, 14(1):7-17. [9] MAJOR J E, JOHNSEN K H, BARSI D C, et al. Fine and coarse root parameters from mature black spruce displaying genetic soil moisture interaction in growth[J]. Can J For Res, 2012, 42(11):1926-1938. [10] 朱航勇, 张含国, 张振, 等.整合生物量及碳汇的帽儿山地区兴安落叶松31年生种源选择[J].东北林业大学学报, 2014, 42(7):5-10. ZHU Hangyong, ZHANG Hanguo, ZHANG Zhen, et al. Integrating biomass and carbon sequestration of 31-year Larix gmelinii provenance selection in Maoershan[J]. J Northeast For Univ, 2014, 42(7):5-10. [11] 董点, 林天喜, 唐景毅, 等.紫椴生物量分配格局及异速生长方程[J].北京林业大学学报, 2014, 36(4):54-63. DONG Dian, LIN Tianxi, TANG Jingyi, et al. Biomass allocation patterns and allometric models of Tilia amurensis[J]. J Beijing For Univ, 2014, 36(4):54-63. [12] 汪金松, 张春雨, 范秀华, 等.臭冷杉生物量分配格局及异速生长模型[J].生态学报, 2011, 31(14):3918-3927. WANG Jinsong, ZHANG Chunyu, FAN Xiuhua, et al. Biomass allocation patterns and allometric models of Abies nephrolepis Maxim[J]. Acta Ecol Sin, 2011, 31(14):3918-3927. [13] 贺庆棠, 袁嘉祖, 陈志泊.气候变化对马尾松和云南松分布的可能影响[J].北京林业大学学报, 1996, 18(1):22-28. HE Qingtang, YUAN Jiazu, CHEN Zhibo. Possible effects of the climate changes on the distribution of Pinus massoniana and Pinus yunnanensis in South China[J]. J Beijing For Univ, 1996, 18(1):22-28. [14] 丁贵杰, 王鹏程.马尾松人工林生物量及生产力变化规律研究(Ⅱ)不同林龄生物量及生产力[J].林业科学研究, 2001, 15(1):54-60. DING Guijie, WANG Pengcheng. Study on change laws of biomass and productivity of Masson pine forest plantation(Ⅱ) biomass and productivity of stand at different ages[J]. For Res, 2001, 15(1):54-60. [15] 刘青华, 张蕊, 金国庆, 等.马尾松年轮宽度和木材基本密度的种源变异及早期选择[J].林业科学, 2010, 46(5):49-54. LIU Qinghua, ZHANG Rui, JIN Guoqing, et al. Variation of ring width and wood basic density and early selection of Pinus massoniana provenances[J]. Sci Silv Sin, 2010, 46(5):49-54. [16] 侯爱敏, 周国逸, 彭少麟.鼎湖山马尾松径向生长动态与气候因子的关系[J].应用生态学报, 2003, 14(4):637-639. HOU Aimin, ZHOU Guoyi, PENG Shaolin. Relationship between climatic factors and radial growth of Pinus massoniana in Dinghushan Mountain[J]. Chin J Appl Ecol, 2003, 14(4):637-639. [17] 程瑞梅, 封晓辉, 肖文发, 等.北亚热带马尾松净生产力对气候变化的响应[J].生态学报, 2011, 31(8):2086-2095. CHENG Ruimei, FENG Xiaohui, XIAO Wenfa, et al. Response of net productivity of Masson pine plantation to climate change in North subtropical region[J]. Acta Ecol Sin, 2011, 31(8):2086-2095. [18] 孟宪宇.测树学[M].北京:中国林业出版社, 2006. [19] GOODMAN A M, ENNOS A R. A comparative study of the response of the roots and shoots of sunflower and maize to mechanical stimulation[J]. J Exp Bot, 1996, 47(303):1499-1507. [20] HENRY H A L, AARSSENN L W. On the relationship between shade tolerance and shade avoidance strategies in woodland plants[J]. Oikos, 1997, 80:575-582. [21] PEICHL M, ARAIN M A. Allometry and partitioning of above and belowground tree biomass in an age-sequence of white pine forests[J]. For Ecol Manage, 2007, 253(1):68-80. [22] RETZLAFF W A, HANDEST J A, O'MALLEY D M, et al. Wholetree biomass and carbon allocation of juvenile trees of loblolly pine (Pinus taeda):influence of genetics and fertilization[J]. Can J For Res, 2001, 31(1):960-970. [23] STOVALL J P, FOX T R, SEILER J R. Allometry varies among 6-year-old Pinus taeda (L.) clones in the Virginia Piedmont[J]. For Sci, 2013, 59(1):50-62. [24] BONGARTEN B C, TESKEY R O. Dry-weight partitioning and its relationship to productivity in loblolly pine seedlings from 7 sources[J]. For Sci, 1987, 33:255-267. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.02.008
 点击查看大图
点击查看大图
计量
- 文章访问数: 2962
- HTML全文浏览量: 673
- PDF下载量: 119
- 被引次数: 0
