






-
刺五加Acanthopanax senticosus为五加科Araliaceae五加属Eleutherococcus多年生落叶小灌木, 是第三纪孑遗植物,在《中国植物红皮书——稀有濒危植物》中被列为濒危植物,其体内含有苷类、黄酮类、有机酸类、香豆素类、多糖类、挥发油、微量元素和氨基酸等多种化学成分[1−2],全株均可入药,具有治疗心血管疾病和抗衰老等保健功能[3],在《野生药材资料保护管理条例》中被列为国家三级保护物种。野生刺五加是森林资源的重要组成部分,对于维持生态平衡有重要的作用。刺五加分布区主要在中国黑龙江省(小兴安岭、张广才岭等)、吉林省(汪清、长白山等)、辽宁省(新宾、本溪等),其中小兴安岭及长白山北部蕴藏量尤为丰富[4]。目前,关于野生刺五加的研究主要在药理药效[5−6]、生殖生态[7−8]、资源保护[9]及栽培利用[10]等方面,关于野生刺五加资源的分布规律和种群结构鲜有报道。
植物种群结构是种群最基本的特征[11]。本研究以中国东北地区3个区域内野生刺五加种群为研究对象,采用静态生命表、存活曲线、死亡曲线和生存分析等方法研究野生刺五加种群年龄结构与动态特征,旨在揭示野生刺五加种群的生存现状,预测种群未来的演替趋势,以期为东北地区野生刺五加种群的可持续发展提供理论依据。
-
2021年4—9月,在查阅相关文献和野外踏查的基础上,选择小兴安岭、张广才岭和长白山区域内野生刺五加分布相对密集的地段进行野外调查。各地区概况如下。
小兴安岭位于黑龙江省北部(46°22′~50°10′N,126°04′~129°13′E),山脉呈西北—东南走向,海拔为500~1000 m,山势和缓,属低山丘陵地形。年平均气温为−2.0~2.0 ℃, 无霜期为 100.0~130.0 d,最冷为1月,平均气温为−20.0~−25.0 ℃,最热为7月,平均气温为19.0~21.0 ℃,年平均降水量为500.0~700.0 mm,多集中在6—8月,年均相对湿度为70%,属于北温带大陆性季风气候。土壤以暗棕色森林土为主,有机质丰富。小兴安岭地区森林植被丰富,是中国重点的林木基地,森林覆盖率达到了72.6%,红松Pinus koraiensis蓄积量达 4300 多万m3,中国一半以上的红松都生长在此。植被类型以红松-落叶阔叶混交林为主,与刺五加种群伴生的主要乔灌木树种有红松、水曲柳Fraxinus mandshurica、臭冷杉Abies nephrolepis、色木槭Acer mono、白桦Betula platyphylla、暴马丁香Syringa reticulata var. amurensis、东北山梅花Philadelphus schrenkii、忍冬Lonicera japonica、舞鹤草Maianthemum bifolium等。
张广才岭位于黑龙江省东南部(45°20′~46°25′N,127°36′~128°39′E),山势高峻,地形复杂,属于流水侵蚀山地,平均海拔为800 m。年平均气温为2.3~3.7 ℃,年平均冻结期为160.0~229.0 d,季节冻土深度为1.8~2.5 m。年平均降水量为440.0~640.0 mm,多集中在6—9月,占全年降水量的50%~70%,年平均蒸发量为1100~2500 mm,属于中温带大陆性季风气候。土壤以黑钙土为主,pH为7.0~7.5。植被类型主要为红松、云杉Picea asperata、冷杉Abies fabri及各种阔叶树的针阔混交林和灌丛等,与刺五加种群伴生的主要乔灌木树种有落叶松Larix gmelinii、蒙古栎Quercus mongolica、胡桃楸Juglans mandshurica、黄檗Phellodendron amurense、五味子Schisandra chinensi、疣枝卫矛Euonymus pauciforus、胡枝子Lespedeza bicolor等[12]。
长白山位于吉林省东南部(41°41′~42°25′N,127°42′~128°16′E),山脉呈东北—西南走向,海拔高度多为1000 m以上。属于温带大陆性山地气候,具有明显的垂直气候变化带谱特征,年平均气温为3.0~7.0 ℃,最低气温为−44.0 ℃,年日照时数不足2300.0 h,无霜期为100 d,年平均积雪深度为50 cm,年降水量为700.0~1400.0 mm[13],多集中在6—9月,冰雪覆盖期长达9个月。土壤主要为腐殖质暗棕壤,土壤肥沃,pH为5.5~6.5。长白山地区地形复杂,地貌组合差异大,长白山植被从低海拔到高海拔依次形成阔叶林、针阔混交林、针叶林、岳桦矮曲林、高山苔原5个分布带,与刺五加种群伴生的主要乔灌木树种有胡桃楸、色木槭、蒙古栎、紫椴Tilia amurensis、东北山梅花、五味子、金银忍冬Lonicera maackii、白花碎米荠Cardamine leucantha、白屈菜Chelidonium majus等。
-
在3个区域按地形设置样地。小兴安岭区域设置了14个样地,张广才岭区域设置了30个样地,长白山区域设置了77个样地。每个样地内布设1个10 m×10 m的乔木样方,在每个乔木样方内设置1个5 m×5 m的灌木样方,再于灌木样方的4个角分别设置1个1 m×1 m的草本样方[14]。对样地内刺五加进行逐木调查,测量其基径、株高、郁闭度等,并对样地内的其他乔木、灌木和草本植物进行全面调查。同时记录各样地地理位置、经纬度、海拔、坡位、坡度、坡向等环境因子,并对各样地种群龄级与环境因子进行相关性分析,其中坡度(上限排除法):0°~0.5°为平原,0.5°~2.0°为微斜坡,2.0°~5.0°为缓斜坡,5.0°~15.0°为斜坡,15.0°~35.0°为陡坡,35.0°~55.0°为峭坡,55.0°~90.0°为垂直壁;坡向:北坡、东北坡、东坡、东南坡、南坡、西南坡、西坡、西北坡分别赋值为1、2、3、4、5、6、7、8;坡位:上坡位赋值为1、下坡位赋值为2。各样地信息见表1。
样地 株数/株 北纬(N) 东经(E) 平均海拔/m 平均郁闭度/% 平均坡度/(°) 平均株高/m 平均基径/cm 小兴安岭 533 46°58′48″ 128°39′36″ 376.08 76.81 12.20 1.12±2.12 a 1.48±0.09 a 张广才岭 171 45°33′00″ 128°19′12″ 253.56 57.00 14.71 0.78±0.30 a 0.63±0.02 b 长白山 436 42°28′12″ 127°50′24″ 825.81 72.00 17.81 0.94±0.76 a 0.74±0.04 b 说明:平均株高和平均基径为平均值±标准差。同列不同小写字母代表不同种群间差异显著(P<0.05)。 Table 1. Information on the distribution of wild A. senticosus populations in different regions
-
本研究采用种群的高度级代替龄级进行分析。将野生刺五加茎干高度以20 cm为1个龄级进行分株统计,第1龄级为0~20 cm (上限排除法),第2龄级为20~40 cm,第3龄级为40~60 cm,以此类推。小兴安岭区域共划分11个龄级,张广才岭区域共划分9个龄级,长白山区域共划分10个龄级,并以龄级为横坐标,株数为纵坐标,绘制不同地区野生刺五加种群年龄结构图。
-
为了定量描述不同区域环境下野生刺五加的种群动态,参考陈晓德[15]种群与群落结构动态量化分析法[16−17],计算种群从n到n+1龄级的种群个体数量(Vn)、整个种群结构的数量变化动态指数($ {V}_{\mathrm{p}n} $)、考虑未来外部干扰时的种群年龄结构指数(${V'_{{\rm{p}}n}})$、种群对完全随机干扰所承担的最大风险概率(Pmax) ,定量描述野生刺五加种群动态特征。Vn、Vpn、$ {V'_{{\rm{p}}n}} $取正、负、0值时分别反映种群个体数量的增长、衰退和稳定的结构动态关系。
本研究中静态生命表的编制参考韩忠明[18]的方法,龄级划分与1.3节中的方法相同。在编制静态生命表的过程中,当出现种群的死亡率为负值的情况时采用分段匀滑技术[19]对种群个体的存活数据进行处理[20],再据此编制野生刺五加种群的静态生命表。静态生命表中,n代表龄级,an为n龄级内存活的个体数,an*为匀滑后n龄级内存活的个体数,ln为标准化存活个体数,$ {\mathrm{l}\mathrm{n}l}_{n} $为ln的自然对数,dn为n~n+1龄级间的标准化死亡数量,$ {q}_{n} $为n~n+1龄级间的死亡率,$ {L}_{n} $为n~n+1龄级间存活的个体数,$ {T}_{n} $为从n龄级到超过n龄级的总个体数,$ {e}_{n} $为进入n龄级个体的生命期望寿命,$ {k}_{n} $为各龄级间的消失率。
-
以龄级为横坐标,标准化存活数的自然对数(lnln)为纵坐标,绘制野生刺五加种群的存活曲线,参照HETT等[21]的方法,分别采用指数函数y=aebn和幂函数y=anb模型检验野生刺五加种群的存活曲线类型,得到检验方程。其中:y、a分别代表n龄级内存活数和种群形成初期的个体数,b为死亡率。再以龄级为横坐标,以死亡率为纵坐标,绘制刺五加种群的死亡率曲线。
-
为了更好地分析3个区域野生刺五加种群的动态变化规律,引入4个生存函数,即生存率函数(Sn)、累计死亡率函数(Fn)、死亡率密度函数(ftn)、危险率函数(λtn)[22],并参考毕晓丽等[23]的计算方法,计算这些生存函数估计值,其中:t 为时间,n 为时间段龄级。
-
由于生境条件及人类干扰活动的空间异质性,东北地区不同区域环境下野生刺五加种群分布特征的差异明显(表1)。本研究调查野生刺五加共计1140株,小兴安岭区域有533株,平均株高为1.12 m,平均基径为1.48 cm;张广才岭区域有171株,平均株高为0.78 m,平均基径为0.63 cm;长白山区域有436株,平均株高为0.94 m,平均基径为0.74 cm。3个区域中刺五加最大基径为14.80 cm,最大株高为2.20 m,均出现在小兴安岭区域,且小兴安岭地区的刺五加基径显著粗于其他两地。
相关性分析(表2)显示:小兴安岭地区对刺五加种群龄级影响较大的环境因子是经度、海拔、坡度和坡向,小兴安岭刺五加种群龄级与坡度、坡向和海拔的正相关性较大,与经度的负相关性较大。张广才岭地区对刺五加种群龄级影响较大的环境因子是经纬度,种群龄级与经纬度的负相关性较大。长白山地区对刺五加种群龄级影响较大的环境因子是经纬度和坡度,种群龄级与经纬度的正相关性较大,与坡度的负相关性较大。
样地 经度 纬度 海拔 坡度 坡向 坡位 郁闭度 小兴安岭 −0.224** 0.098* 0.134** 0.154** 0.159** −0.053 0.016 张广才岭 −0.191* −0.190* 0.103 0.108 0.052 −0.129 −0.093 长白山 0.359** 0.298** −0.006 −0.325** −0.115* 0.033 −0.117* 说明: **表示在P<0.01水平(双侧)上极显著相关; *表示在P<0.05水平(双侧)上显著相关。 Table 2. Correlation coefficients of age class and environmental factors of wild A. senticosus populations in different regions
-
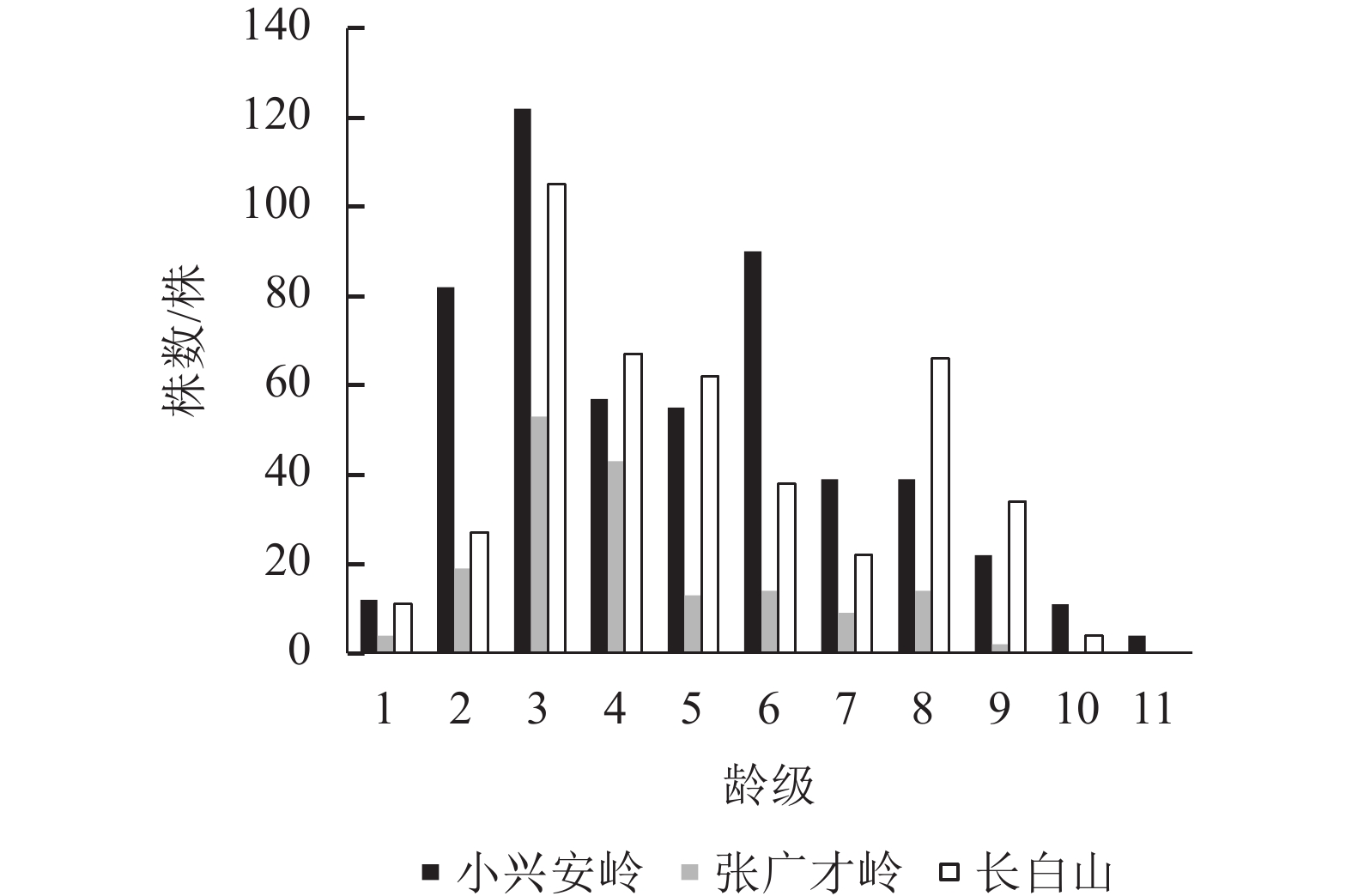
图1显示:3个区域的野生刺五加种群数量均随着龄级的增加呈先升后减的趋势,主要集中在第3~6龄级,说明3个区域的野生刺五加种群均以成龄期植株占优势,种群的稳定性都较好。但各区域1龄级的幼苗数量极少,分别占总数的2.25%、2.34%和2.52%。动态数量化分析结果显示(表3):V1和V2均小于0,也说明了3个区域幼龄级植株数量少,呈负增长的趋势,这极大地限制了成龄期植株的发展潜力和种群延续。3个区域野生刺五加种群的总体数量变化动态指数$ {V'_{{\rm{p}}n}} $均小于$ {V}_{{\rm{p}}n} $,但都大于0,其中$ {V'_{{\rm{p}}n}} $更加趋于0,表明忽略外部干扰时均为增长型种群;3个区域受随机干扰所承担的风险概率极大值 (Pmax) 分别为0.01、0.03和0.01,趋近于0,说明对外界随机干扰均具有较高的敏感性。
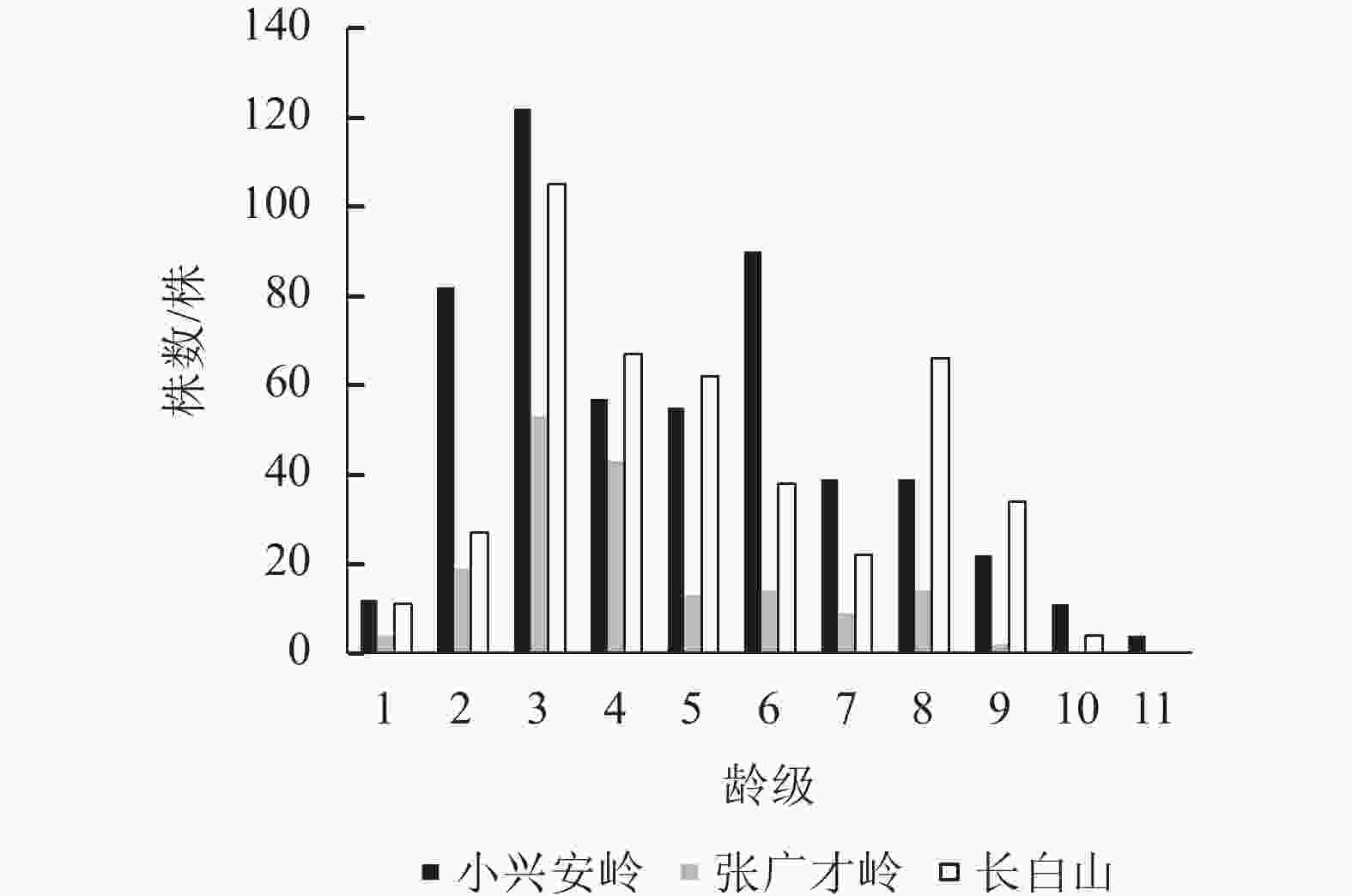
Figure 1. Age-class structure of wild A. senticosus populations in different regions
样地 年龄结构动态变化指数/% $ {{P}_{\max}}$ V1 V2 V3 V4 V5 V6 V7 V8 V9 V10 V11 Vpn $ {V'_{ {\rm{p} }n} }$ 小兴安岭 −85.37 −32.79 53.28 3.51 −38.89 56.67 0.00 43.59 50.00 63.64 − 32.33 0.24 0.01 张广才岭 −78.95 −64.15 18.87 69.77 −7.14 35.71 −35.71 85.71 − − − 40.68 0.99 0.03 长白山 −59.26 −74.29 36.19 7.46 38.71 42.11 −66.67 48.48 88.24 − − 40.87 0.34 0.01 说明:Vn是种群n~n+1龄级的种群个体数量,${V_{{\rm{p}}n} }$是整个种群结构的数量变化动态指数,${V'_{ {\rm{p} }n} }$是考虑未来外部干扰时的种群年龄结构指数,Pmax是种群对完全随机干扰所承担的最大风险概率。−表示无此项。 Table 3. Dynamic change index of wild A. senticosus populations in different regions
-
随种群龄级的增加,3个区域野生刺五加种群的个体标准化存活数(ln)逐渐减少(表4)。小兴安岭和长白山区域种群的期望寿命值(en)也随龄级的增加逐渐降低,而张广才岭区域种群的期望寿命值随龄级增加呈先增后减的变化,在第5龄级达到最大值,表明该阶段的种群生存质量最高,生理代谢旺盛,且具有较高的生存环境质量。
样地 龄级 高度级/cm an an* ln lnln dn qn Ln Tn en kn 小兴安岭 1 0~20 12 308 1000 6.91 188 0.19 906 3454 3.45 0.21 2 20~40 82 250 812 6.70 169 0.21 728 2548 3.14 0.23 3 40~60 122 198 643 6.47 146 0.23 570 1820 2.83 0.26 4 60~80 57 153 497 6.21 130 0.26 432 1250 2.52 0.30 5 80~100 55 113 367 5.91 107 0.29 314 818 2.23 0.34 6 100~120 90 80 260 5.56 88 0.34 216 505 1.94 0.41 7 120~140 39 53 172 5.15 68 0.40 138 289 1.68 0.50 8 140~160 39 32 104 4.64 46 0.44 81 151 1.45 0.58 9 160~180 22 18 58 4.06 29 0.50 44 70 1.21 0.69 10 180~200 11 9 29 3.37 6 0.21 26 26 0.99 0.23 11 200~220 4 7 23 3.14 − − − − − − 张广才岭 1 0~20 4 143 1000 6.91 358 0.36 821 2678 2.68 0.44 2 20~40 19 92 642 6.47 248 0.39 518 1678 2.61 0.49 3 40~60 53 56 394 5.98 160 0.40 314 1036 2.63 0.52 4 60~80 43 33 235 5.46 92 0.39 189 641 2.73 0.50 5 80~100 13 20 142 4.96 46 0.32 120 407 2.86 0.39 6 100~120 14 14 97 4.57 20 0.21 87 264 2.72 0.23 7 120~140 9 11 77 4.34 16 0.20 69 167 2.17 0.23 8 140~160 14 9 61 4.12 32 0.52 45 91 1.48 0.74 9 160~180 2 4 29 3.38 − − − − − − 长白山 1 0~20 11 162 1000 6.91 198 0.20 901 3242 3.24 0.22 2 20~40 27 130 802 6.69 176 0.22 714 2341 2.92 0.25 3 40~60 105 101 627 6.44 154 0.25 550 1627 2.60 0.28 4 60~80 67 77 473 6.16 132 0.28 407 1077 2.28 0.33 5 80~100 62 55 341 5.83 109 0.32 287 670 1.96 0.39 6 100~120 38 38 232 5.45 87 0.38 188 383 1.65 0.47 7 120~140 22 23 145 4.97 65 0.45 112 195 1.35 0.60 8 140~160 66 13 79 4.37 43 0.55 58 83 1.05 0.79 9 160~180 34 6 36 3.58 21 0.59 25 25 0.69 0.89 10 180~200 4 2 15 2.70 − − − − − − 说明:n. 龄级;an. 第n龄级内存活的个体数;an*. 匀滑后n龄级内存活的个体数;ln. 标准化存活个体数(一般以1 000为基数);$ {\mathrm{l}\mathrm{n}l}_{n} $. ln的自然对数;dn. n~n+1龄级的标准化死亡数量;$ {q}_{n} $. n~n+1龄级间的死亡率;$ {L}_{n} $. n~n+1龄级之间存活的个体数;$ {T}_{n} $. 从n龄级到超过n龄级的总个体数;$ {e}_{n} $. 进入n龄级个体的生命期望寿命;$ {k}_{n} $. 各龄级间的消失率。−表示无此项。 Table 4. Static life tables of wild A. senticosus populations in different regions
-
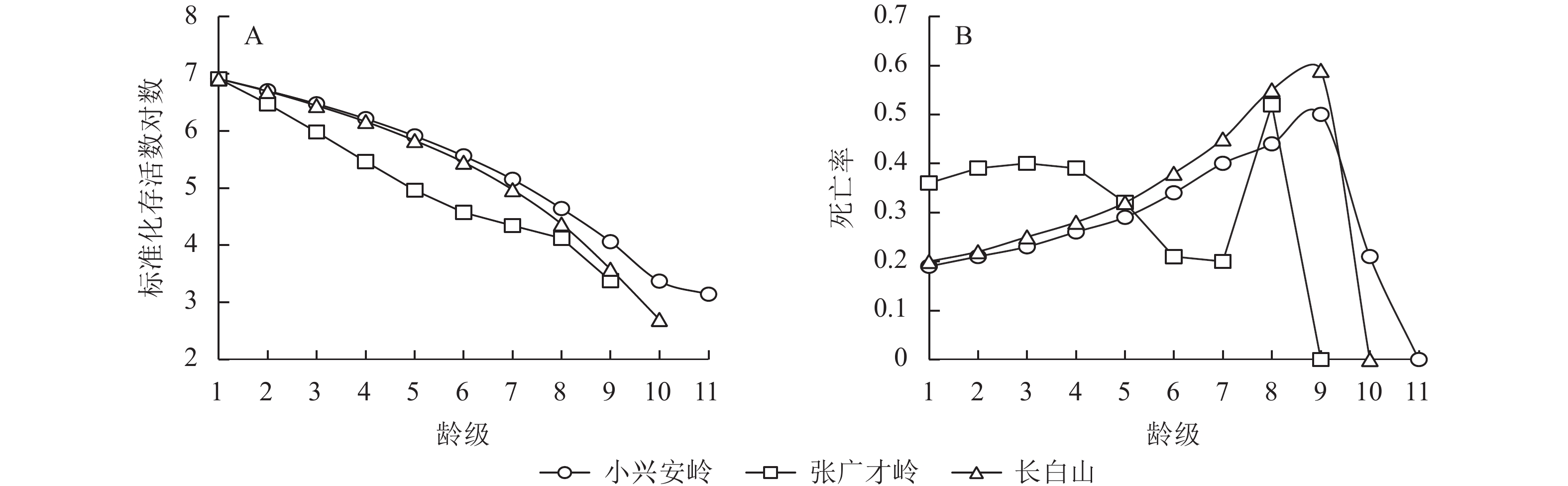
由图2可知:小兴安岭和长白山种群在1~8龄级间的存活曲线变化一致,下降幅度均较张广才岭区域更大,第8龄级之后张广才岭和长白山区域种群存活曲线下降趋势加快,而小兴安岭种群存活曲线趋于平稳(图2A)。幂函数和指数函数2种模型的拟合结果显示:3个区域野生刺五加种群的拟合结果均达到了显著水平(P<0.05),且3个区域下指数模型的决定系数(R2)均大于幂函数模型(表5),P值均小于幂函数模型,因此,认为3个区域野生刺五加种群的存活曲线为Deevey-Ⅱ型。小兴安岭和长白山种群只存在1个死亡高峰,在第9龄级,而张广才岭种群存在2个死亡高峰,分别在3龄级和8龄级,最高峰出现在8龄级(图2B)。

Figure 2. Survival (A) and killing curves (B) of wild A. senticosus populations in different regions
种群 检验方程 F R2 显著性 小兴安岭 $ {y={8.405n}^{-0.312}}$ 22.295 0.712 0.001 $ {y={8.272\mathrm{e} }^{-0.08n}}$ 119.018 0.930 0.000 张广才岭 $ {y={7.700n}^{-0.302}}$ 49.698 0.877 0.000 $ {y={7.626\mathrm{e} }^{-0.084n}}$ 390.944 0.982 0.000 长白山 $ {y={8.467n}^{-0.334}}$ 14.676 0.647 0.005 $ {y={8.569\mathrm{e} }^{-0.094n}}$ 54.898 0.873 0.000 Table 5. Examination of the survival curves of wild A. senticosus populations in different regions
-
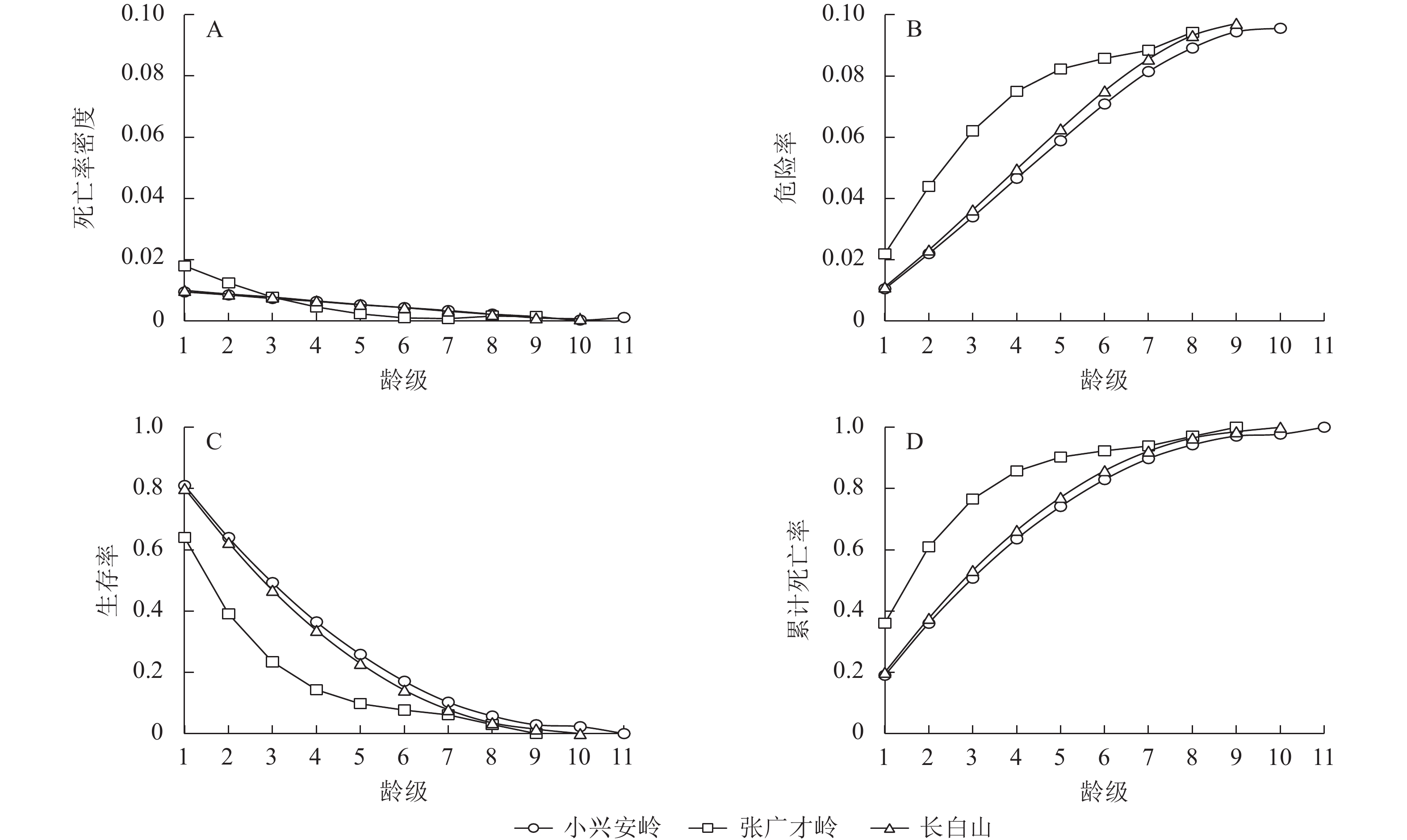
如图3所示:以龄级为横坐标,4个生存函数估计值为纵坐标,绘制不同区域野生刺五加种群生存函数曲线。3个区域种群的死亡密度均在第1龄级出现最高峰,危险率曲线总体上保持逐渐上升的趋势(图3A~B)。3个区域种群均在第3龄级左右生存率和累积死亡率达到平衡,这反映出第3龄级以后野生刺五加种群将进入衰退期(图3C~D)。

Figure 3. Survival function curves of wild A. senticosus population in different regions
-
刺五加喜温暖湿润气候,适宜生长在土壤较为湿润、腐殖质层深厚的混交林、山地阔叶林下及林缘[18, 24]。本研究通过对小兴安岭、张广才岭和长白山3个区域内野生刺五加种群的调查发现:小兴安岭和长白山区域刺五加植株数量多,茎干粗壮,龄级分布范围广,张广才岭区域植株数量少,茎干细矮,龄级分布范围窄,说明小兴安岭和长白山区域气候环境更适宜野生刺五加生长。王书越等[25]基于MaxEnt模型对野生刺五加在东北地区的空间分布进行评估,也证实了野生刺五加的生态适宜区主要在长白山山脉及小兴安岭地区,与本研究结果相符。相关性分析结果显示:影响3个区域野生刺五加种群龄级的环境因子主要有海拔、经纬度、坡向和坡度,王书越等[25]的研究也表明刺五加群落的存活同坡度、坡向、光照条件以及所处林型都有一定联系。小兴安岭和张广才岭区域种群龄级随经度的升高而减小,而长白山区域种群龄级随经度的升高而增加。植物形态特征反映植物对异质环境的响应。在不同生境下,植物采取不同形态特征,调整生存策略,以增加种群的生存适合度和竞争能力[26]。说明刺五加种群对异质性生境采取了不同的适应政策,也表明野生刺五加种群分布是其与环境长期相互作用、相互适应的结果。
-
植物种群结构反映植物对自然环境的适应性[27−28],比较不同区域环境下的野生刺五加种群结构特征有助于了解该物种与其生存环境之间的适合度[29]。本研究发现:3个区域野生刺五加种群的年龄结构均呈非典型金字塔型,种群呈现增长型,但1~3龄级呈衰退趋势,如果没有幼龄植株补充,野生刺五加种群整体的长期稳定性将难以维持,这与其他一些濒危植物情况相似[30]。野生刺五加种子萌发的实生苗很少,现在生存的刺五加种群多为无性植株产生,主要通过根茎上的潜伏芽转化植株[31],这样基因多样性减少,导致出现绝种的风险,在一定程度上也限制了野生刺五加种群的自然更新。3个区域种群出现死亡高峰的频率不同,小兴安岭和长白山区域种群都只有1次死亡高峰,均在第9龄级,这是因为当植物进入老龄级后,生理机能衰退,种群更容易受到外界干扰。张广才岭区域种群出现2次死亡高峰,第1次在第3龄级,是从幼龄期向成龄期过渡阶段。韩忠明[18]研究表明:刺五加幼苗在第3龄级开始脱离母株,并且由营养生长转向生殖生长,此时植株对营养空间的需求增大,种群自疏和他疏作用增强,导致了较高的死亡率。第2次死亡高峰在第8龄级,其原因与其他2个地区相同,即植株的生长发育到达了生理衰退期。
-
植物种群的发展是复杂且漫长的过程,不仅有物种内和物种间的竞争,同时还伴有物种和环境的适应过程[32]。长期野外定位观测发现:造成野生刺五加种群急速减少的主要外在因素是人们过度采伐[33]。野生刺五加的活性成分在落叶后至萌发前后最高[34]。对于张广才岭区域野生刺五加种群来说,首先要保护好现有的成年植株,并提高成年植株的种子生产能力,然后建立一定规模的刺五加苗圃,规模化生产优质种苗,并将达到出圃标准的幼苗经野外驯化后进行回归种植[35]。其次,要保证野生刺五加无性繁殖个体的成活率,加大就地保护力度。可以利用规定采收期和采收方式,防止人类对野生刺五加资源的破坏,应在秋季落叶后至春季萌发前采收, 其他时期不允许采收。根茎是野生刺五加种群扩增的主要繁殖方式,应禁止采挖。
-
3个区域野生刺五加种群第1龄级和第2龄级的植株数量均少于第3龄级,说明幼龄级植株数量少。虽然种群总体呈现增长型,但1~3龄级呈衰退趋势。如果没有幼龄个体的补充,野生刺五加种群整体的长期稳定性将难以维持。4个生存函数分析显示:3个区域野生刺五加种群的生存率曲线和累计死亡率曲线总体表现为逐渐下降或逐渐上升的趋势,且各生存函数曲线斜率均表现为前期、中期高于后期,表明3个区域野生刺五加种群数量均具有幼龄期和成龄期种群结构波动较大,而老龄期稳定的特点。
Population structure and dynamics of wild Acanthopanax senticosus in different regions of Northeast China
doi: 10.11833/j.issn.2095-0756.20230377
- Received Date: 2023-06-25
- Accepted Date: 2023-10-17
- Rev Recd Date: 2023-10-13
- Available Online: 2024-03-21
- Publish Date: 2024-04-01
-
Key words:
- Northeast China /
- wild Acanthopanax senticosus /
- age structure /
- quantitative dynamics /
- survival analysis
Abstract:
| Citation: | WEI Hongling, XIE Shengnan, TIAN Xuchen, et al. Population structure and dynamics of wild Acanthopanax senticosus in different regions of Northeast China[J]. Journal of Zhejiang A&F University, 2024, 41(2): 333-342. DOI: 10.11833/j.issn.2095-0756.20230377

|
 DownLoad:
DownLoad: