






-
全球竹类植物约有70余属1 642种,广泛分布于热带、亚热带及温带地区。中国是世界上竹类资源最为丰富的国家之一,拥有约43属近751种[1]。刚竹属Phyllostachys隶属于禾本科Poaceae竹亚科Bambusoideae,以其丰富的物种多样性、广泛的环境适应性及显著的经济和生态价值而备受关注,是中国特有竹属之一。自1843年建立刚竹属以来,对刚竹属的研究逐渐深入[2−4]。近年来,对刚竹属的研究取得了显著进展,揭示了刚竹属与其他属之间的亲缘关系[5];发现了刚竹属的种下分类群存在叶绿体基因序列差异[6];找出了刚竹属中具有增强植物抗逆性及促进植物生长功能的韧皮部蛋白,并进行鉴定及表达分析[7] ;通过对毛竹Ph. edulis ICE基因家族进行鉴定及分析,找出了响应毛竹抗寒关键家族成员[8]。生态学研究则进一步阐明了刚竹属竹种在生态系统中的重要作用,如竹林对降水的截流作用、对局部气候的调节作用以及扩张对土壤质量的影响[9−11]。在药用研究方面,则发现刚竹属竹种的竹笋、竹叶和竹茹等富含黄酮类、二萜酸类、木脂素类等多种活性成分,具有潜在药用价值[12]。
刚竹属是分布面积最大、经济价值最高、研究最为深入的竹属,但以下问题仍值得进一步研究:刚竹属各竹种之间表型性状有何差异?不同的表型性状之间是否具有密切联系?种间的差异是否大于种内的差异?能否通过某一表型性状预测其他表型性状?本研究以浙江安吉中国竹子博览园刚竹属111种(包括种下等级)竹种为研究对象,通过测定胸径、壁厚等表型性状和纤维性状,运用变异分析、相关性分析、聚类分析、拟合方程等方法,探究刚竹属竹种表型性状与纤维性状的变异规律和相关性,并建立性状特征拟合方程,为优良竹种选育提供理论依据。
-
试验地位于浙江省湖州市安吉县中国竹子博览园内(30°38′N,119°41′E)。该地区属于亚热带季风气候,年平均气温为15.0~17.0 ℃,极端最高气温为39.1 ℃,极端最低气温为−8.1 ℃,年平均日照时数为1 668.3 h,年平均无霜期为226.0 d。年平均降水量为1 478.0 mm。全年气候温和、光照充足、雨量充沛、四季分明,夏季高温多雨,水热条件好。无垂直气候特征,气候特点和地理环境非常适合竹类植物生长。土壤为冲积沙壤土,地形起伏小。
-
在试验地选取2年生刚竹属竹种,齐地伐倒,每个竹种取3株为1组重复,共3组重复,共计111种(包括种下等级)。测量每株的壁厚(mm)、胸径(mm)、壁腔比、秆高(m)、总节数(节)、枝下高节数(节)、枝条长度(cm)、节长(cm)共8个表型性状和纤维长度(mm)、纤维宽度(μm)、卷曲度(%)、扭结角度、细小纤维A类(%)、细小纤维B类 (%)、加权细小纤维 (%)、纤维化程度(%)共8个纤维性状。
表型性状:由游标卡尺、卷尺结合目测计数完成,在胸高处测量壁厚、胸径、壁腔比、节长,枝条长度选取从下至上第2分枝的最长枝条长度。依据袁金玲等[13]的方法,针对具有特殊秆型形态的竹种,在胸径位置前后选取10个竹节作为测量单元。每个竹节分别测量4个方向的节长与壁厚数据,取其平均值;若某一方向因秆型特殊(如畸形、缺裂等)无法测量,则选取剩余3个方向的数据计算平均值。
纤维性状:使用富兰克林纤维法测量,按五等分法将竹秆分上中下3个部分切成火柴大小细条,每个细条需包含竹青及竹黄[14]。卷曲度:长度加权平均纤维卷曲度;扭结角度:纤维扭结角度;细小纤维A类:长度小于0.2 mm的细小纤维所占总纤维投影面积的百分比;细小纤维B类:薄片状细小纤维(类型B)占长度的百分比。宽度小于10.0 μm且长度超过0.2 mm的颗粒被归入细小纤维B类。将这些颗粒的总长度除以所有长度超过0.2 mm的测量颗粒的总长度;加权细小纤维:长度加权平均算术分布中的细纤维百分比。
-
由表1可见:刚竹属竹种表型性状变异系数为21.75%~67.72%,其中胸径、壁厚、枝下高节数、秆高属于高度变异,变异系数分别为62.72%、41.12%、40.45%、40.39%;枝条长度、节长、总节数、壁腔比属于中等变异,变异系数分别为29.84%、23.92%、22.19%、21.75%。其中实心竹Ph. heteroclada f. solida竹腔接近实心,内径为0.1 mm,壁腔比达132.97,与其他竹种壁腔比差异较大,故不予统计。壁厚变异程度高的原因是由于厚皮毛竹Ph. edulis f. pachyloen、龟甲竹Ph. edulis ‘Kikko-chiku’、佛肚毛竹Ph. edulis f. ventricosa等变异品种壁厚明显大于其他竹种;胸径和秆高变异程度高的原因主要是由于毛竹及其变异品种胸径和秆高较大导致的。龟甲竹和元宝毛竹Ph. edulis ‘Yuanbao’秆型都具有特殊的膨大或凹陷,导致其节长明显低于其他竹种,分别为7.18和5.97 cm。
项目 壁厚/mm 胸径/mm 壁腔比 秆高/m 总节数/节 枝下高节数/节 枝条长度/cm 节长/cm 变异幅度 1.83~14.04 6.80~104.82 1.18~4.48 1.77~16.25 16.33~58.33 4.00~26.83 42.88~198.62 5.97~40.22 平均值±标准误 5.37±0.21 29.47±1.75 1.75±0.25 5.58±0.21 34.16±0.72 10.58±0.44 90.27±2.56 20.65±0.47 方差 4.87 341.54 0.16 5.08 57.48 21.94 725.50 24.39 变异系数/% 41.12 62.72 21.75 40.39 22.19 40.45 29.84 23.92 项目 纤维长度/mm 纤维宽度/μm 卷曲度/% 扭结角度 细小纤维A类/% 细小纤维B类/ % 加权细小纤维/% 纤维化程度/% 变异幅度 0.80~2.31 12.19~20.95 0.03~11.06 5.53~36.16 9.99~23.53 22.54~70.97 57.22~85.03 1.49~6.83 平均值±标准误 1.92±0.02 14.06±0.09 5.15±0.19 26.13±0.26 17.64±0.25 40.69±0.88 75.47±0.42 4.23±0.12 方差 0.04 0.97 3.87 7.65 6.86 85.50 19.21 1.70 变异系数/% 10.91 7.02 38.23 10.58 14.84 22.72 5.81 30.75 Table 1. Phenotypic and fiber characteristics in Phyllostachys
刚竹属竹种纤维性状变异系数为5.81%~38.23%,其中卷曲度、纤维化程度、细小纤维B类 为中等变异,变异系数分别为38.23%、30.75%和22.72%;细小纤维A类、扭结角度、纤维长度、纤维宽度、加权细小纤维为小变异,变异系数分别为14.84%、10.91%、10.58%、7.02%、5.81%。纤维性状无高度变异,整体变异程度低于表型性状,说明刚竹属竹种纤维性状遗传稳定性高于表型性状。
-
本次调查的刚竹属竹种有52个原种,其中18个原种具有种下等级,且10个原种的种下等级数量超过4个,共计111种刚竹属竹种。每个原种及其种下等级的总和为1个竹种类群,分别对这10个种下等级数量超过4个的类群表型性状进行变异程度分析(表2)。毛竹类群种下等级数量最多,共17个,其表型性状中胸径、壁腔比、秆高3个指标为高度变异,壁腔比、秆高、节长等指标的变异程度高于全属,该类群的特殊变种(变型)达8个,占47.05%。麻衣竹Ph. edulis f. exaurita、金丝毛竹Ph. edulis f. gracilis胸径明显小于其他毛竹变种(变型),厚皮毛竹壁腔比明显大于其他变种(变型),麻衣竹、元宝毛竹秆高明显低于其他变种(变型)。水竹Ph. heteroclada类群中种下等级数量为4个,其壁厚、壁腔比、枝下高节数属于高度变异且皆大于全属的变异程度,主要由于实心竹的特殊竹腔导致。水竹的2个变型黑水竹Ph. heteroclada f. denigrata和黄秆水竹Ph. heteroclada f. flaviculmis属于竹秆颜色变异,表型性状变异程度较小。桂竹Ph. reticulata、早竹Ph. violascens、黄槽竹Ph. aureosulcata类群中种下等级数量分别为7、5、5个,黄古竹Ph. angusta、篌竹Ph. nidularia、罗汉竹Ph. aurea、金竹Ph. sulphurea、乌哺鸡竹Ph. vivax类群中种下等级数量皆为4个。这些种下等级各表型性状变异系数皆小于全属变异系数,表型性状变异程度较小,其种内多为秆色、条纹或是沟槽等外观特征变异,遗传较为稳定。
类群 壁厚/mm 胸径/mm 壁腔比 秆高/m 总节数/节 枝下高节数/节 枝条长度/cm 节长/cm 毛竹类群 9.16±0.65(29.48) 60.44±6.45(44.03) 1.65±0.18(44.85) 7.81±0.90(47.50) 41.28±1.99(19.84) 15.51±1.48(39.26) 131.16±7.07(22.23) 21.62±1.98(37.84) 桂竹类群 4.63±0.26(14.69) 21.60±2.22(27.22) 1.86±0.08(11.29) 4.74±0.44(24.68) 31.95±1.90(15.71) 9.36±0.84(23.72) 80.8±6.46(21.16) 18.37±1.14(16.39) 早竹类群 5.42±0.43(18.00) 30.58±2.89(21.00) 1.60±0.02(3.00) 5.01±0.58(26.00) 39.43±2.19(12.00) 12.97±1.36(24.00) 75.57±4.00(12.00) 16.07±1.52(21.00) 黄槽竹类群 3.95±0.26(14.68) 21.60±2.00(20.69) 1.63±0.03(4.91) 5.71±0.51(19.96) 32.70±1.60(10.92) 9.23±0.88(21.24) 100.10±0.12(22.61) 23.05±1.20(11.63) 黄古竹类群 3.98±0.17(8.54) 17.14±0.79(9.22) 1.98±0.12(11.62) 4.01±0.23(11.47) 29.50±1.85(12.54) 9.00±1.39(30.78) 73.90±4.74(12.84) 18.10±0.61(6.69) 水竹类群 3.77±0.95(50.40) 11.82±1.15(19.46) 32.48±30.41(187.22) 3.08±0.50(32.79) 22.21±2.46(22.15) 5.67±1.21(42.68) 77.21±8.70(22.54) 17.24±2.33(26.97) 篌竹类群 3.63±0.29(15.98) 15.82±2.23(28.19) 2.00±0.12(12.00) 4.27±0.41(19.20) 28.46±2.93(20.63) 8.79±1.01(22.87) 73.46±3.71(10.11) 22.91±1.50(13.14) 罗汉竹类群 4.47±0.17(7.38) 16.73±0.61(7.29) 2.33±0.14(12.02) 3.05±0.15(9.84) 30.67±2.26(14.70) 10.33±1.51(29.24) 62.09±3.13(10.10) 16.26±1.02(12.55) 金竹类群 6.04±0.14(4.80) 31.58±1.09(6.90) 1.65±0.06(7.27) 5.58±0.32(11.47) 32.25±1.10(6.79) 7.42±0.56(14.96) 90.23±4.39(9.73) 22.49±1.22(10.80) 乌哺鸡竹类群 5.21±0.19(7.29) 31.16±0.95(6.13) 1.55±0.03(3.87) 6.04±0.50(16.56) 37.92±1.02(5.38) 12.06±2.54(36.48) 78.23±12.39(31.66) 19.31±1.56(16.16) 说明:数值为平均值±标准误。括号中数值为变异系数(%)。 Table 2. Phenotypic variation of the 10 clusts in Phyllostachys
-
对刚竹属竹种表型特征的相关分析结果表明:8个表型性状与8个纤维性状之间存在39对显著相关关系(P<0.05),其中5对显著负相关(P<0.05)。表型性状中,壁厚与胸径、秆高与总节数2对相关性较高,相关系数均超过0.80;胸径与其他7个表型性状之间皆为极显著相关(P<0.01),壁厚、秆高、总节数、枝下高节数、枝条长度、节长等6个性状与壁腔比相关性未达显著水平,壁腔比仅与胸径显著相关(P<0.05)。纤维性状中,细小纤维A类与加权细小纤维相关系数最大,达0.86;纤维长度与其他纤维性状显著相关关系最多;扭结角度和细小纤维B类呈显著负相关(P<0.05),纤维宽度不仅与卷曲度和纤维化程度呈显著正相关(P<0.05),还与表型性状中的胸径和枝下高节数呈显著相关(P<0.05)。
综上所述,刚竹属竹种不同表型性状之间、不同纤维性状之间相关性强,但表型性状与纤维性状关联不强;表型性状中,胸径与其他性状显著相关性强且皆为正相关,胸径较大时其他表型性状也较大,因此可以通过胸径估测其他7个表型性状。纤维性状中,纤维长度与纤维宽度的增加会导致粗糙纤维变少、纤维化程度增大。
-
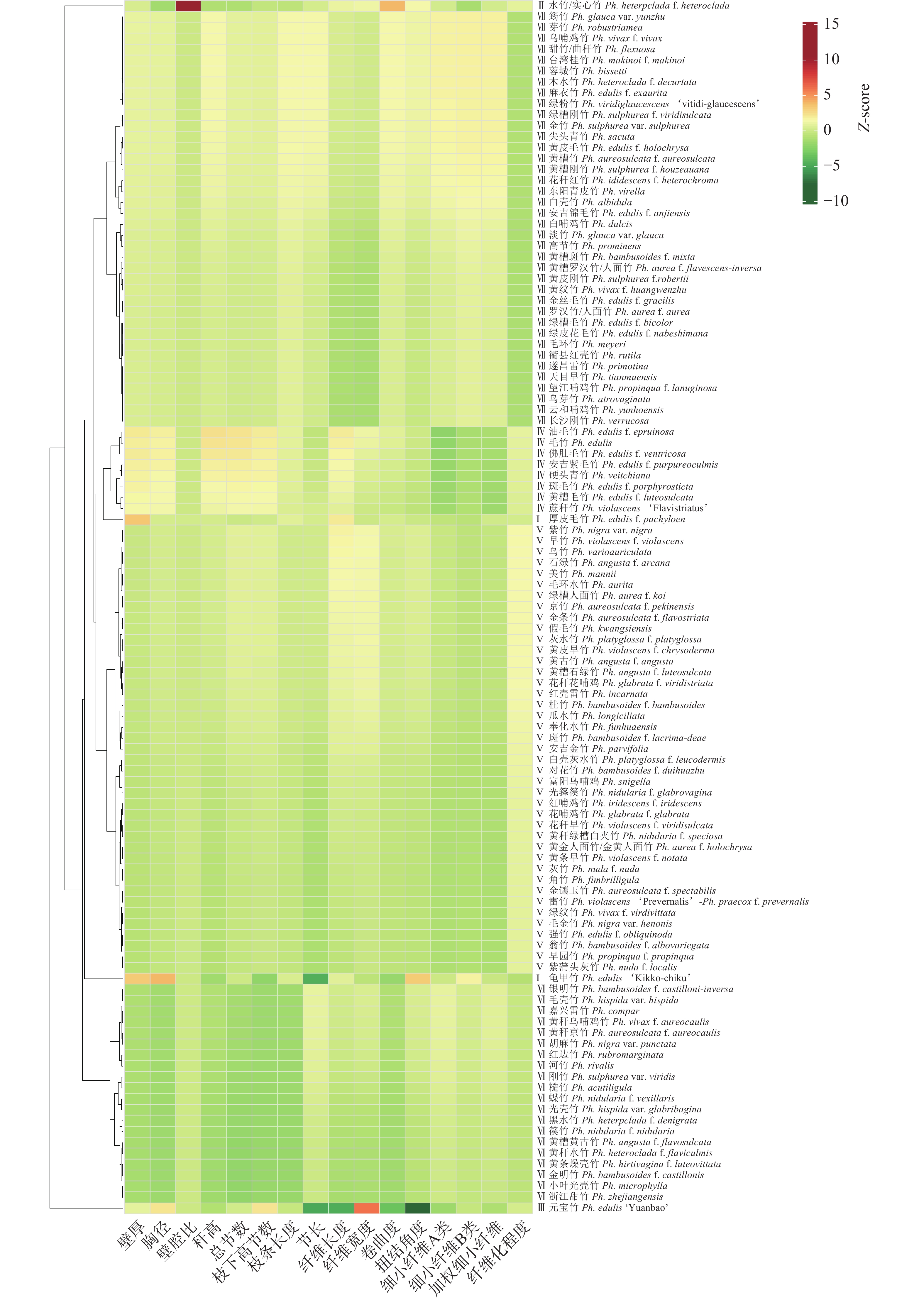
利用欧氏距离法对刚竹属竹种表型特征进行聚类分析(图1)。当欧氏距离为8时,刚竹属111种被分为7个大类,每大类都具有明显特征:第Ⅰ类仅包含2个竹种,分别为龟甲竹和厚皮毛竹,表现为壁厚明显大于其他竹种;第Ⅱ类仅包含实心竹1个竹种,性状表现为竹秆近实心,壁腔比明显大于其他竹种;第Ⅲ类仅包含元宝毛竹1个竹种,性状表现为竹节短缩坍塌,节长明显低于其他竹种;第Ⅳ类包含8个竹种,主要包括毛竹及其变种,性状表现为胸径、秆高和枝条长度明显大于其他竹种;第Ⅴ类包含41个竹种,这类竹种以雷竹Ph. violascens、桂竹等为代表,主要的性状表现是各表型指标皆略低于刚竹属平均值;第Ⅵ类包含20个竹种,是以河竹Ph. rivalis、糙竹Ph. acutiligula、篌竹等水竹组的竹种为代表,主要性状表现为壁厚、秆高、总节数、枝下高节数、节长5个表型性状明显低于其他竹种;第Ⅶ类包含38个竹种,其中刚竹组竹种有18个,以白哺鸡竹Ph. dulcis、乌哺鸡竹、淡竹Ph. glauca等的刚竹组竹种为代表,占比约47.37%,主要性状表现为各表型指标皆略高于刚竹属平均值,是最能代表刚竹属的类别。

Figure 1. Phenotypic characteristics of 111 species (including sub-species) of Phyllostachys by cluster analysis
-
为预测刚竹属竹种表型性状,建立刚竹属竹种表型性状拟合曲线方程。因胸径与其他表型性状相关性皆为极显著且测量相对容易,故以胸径(x)为自变量,其他表型性状为因变量建立拟合方程(表3)。每个性状皆建立线性和二次2种方程,除壁腔比的线性方程外,其他方程的决定系数(R2)皆大于0.800,均达到极显著水平(P<0.01),说明方程拟合值较高,能较好地预测表型性状指标。胸径与壁厚(y1)、秆高(y3)和总节数(y4)的相关性最强,二次拟合方程R2最高,拟合程度最好,分别为y1=0.227 7x−0.001 2x2(R2=0.987)、 y3=0.232 1x−0.001 0x2(R2=0.967)、 y4=1.621 2x−0.012 0x2(R2=0.973)。
性状 方程属性 拟合方程 R² 性状 方程属性 拟合方程 R² 壁厚(y1) 线性 y1=0.161 5x 0.936** 枝下高节数(y5) 线性 y5=0.331 5x 0.882** 二次 y1=0.227 7x−0.001 2x² 0.987** 二次 y5=0.449 0x−0.002 2x² 0.918** 壁腔比(y2) 线性 y2=0.043 8x 0.658** 枝条长度(y6) 线性 y6=2.726 9x 0.895** 二次 y2=0.087 1x−0.000 8x² 0.867** 二次 y6=3.911 6x−0.023 0x² 0.950** 秆高(y3) 线性 y3=0.179 4x 0.940** 节长(y7) 线性 y7=0.591 4x 0.811** 二次 y3=0.232 1x−0.001 0x² 0.967** 二次 y7=0.974 7x−0.007 4x² 0.921** 总节数(y4) 线性 y4=1.004 8x 0.868** 二次 y4=1.621 2x−0.012 0x² 0.973** 说明:x为胸径。**表示P<0.01。 Table 3. Phenotypic variation fitting equations in Phyllostachys genus
-
属作为种之上的分类等级,为植物遗传变异研究提供了更为宽泛的遗传信息和更为复杂的生物多样性视角。表型性状作为分类学中不可或缺的指标,在一定程度上能够直观反映植物的遗传多样性,为遗传学研究提供重要线索。近年来,多位学者深入开展了不同属植物种质资源的表型性状变异分析。何潇等[15]选取结缕草属Zoysia 43份种质资源作为研究材料,发现该属表型性状变异显著,变异范围达24.29%~64.14%,揭示了结缕草属在遗传多样性上的丰富性。这一变异范围与刚竹属相似。王欢利等[16]研究表明:25种椴树Tilia果实的表型性状变异主要源于种间差异,表型分化系数为76.76%~83.72%,种间变异是主导椴树属表型多样性的关键因素。曾慧杰[17]发现:忍冬属Lonicera种质间变异系数高达64.51%,表明种质资源间的遗传多样性显著。尽管刚竹属中部分竹种如毛竹和水竹存在特殊变种或变型,导致种下等级内的变异程度显著,但总体来说刚竹属种间的变异程度依然大于种内,说明种水平的选择育种优先度大于种内水平的选择育种。
本研究中刚竹属111种竹种均取自安吉中国竹子博览园,环境条件基本一致,对研究对象表型性状的变异影响较小,刚竹属表型性状的种间变异程度依然显著,而纤维性状的变异则表现出相对稳定的特点,说明刚竹属的遗传多样性较高,选择育种潜力大。秆形性状变异时,表型性状变异程度较大,这可能是秆形性状控制基因与所测表型性状控制基因连锁所致。毛竹和水竹等竹种的种内变异幅度较大可能是由于这些竹种在遗传上具有较高的可塑性或存在特殊的遗传机制,导致其在面对环境变化时能够产生更为显著的表型变异。这种现象提示其他竹种也可能具有较大的变异幅度。
-
竹类植物是植物界中一个庞大且复杂的类群,其分类问题历来是植物学界关注的热点与难点。《中国植物志》中文版记载刚竹属竹种50种,英文版记载刚竹属竹种51种。随着近年来分类工作的不断深入,该属下的变种、变型呈现出逐渐增多的趋势。安吉中国竹子博览园作为竹类植物的重要保育基地,现已收集到111种刚竹属竹种[18]。根据形态特征,刚竹属被进一步细分为水竹组和刚竹组。水竹组和刚竹组的分类主要基于地下茎的结构、竿的形态、箨鞘与箨片的特征、叶片的形态以及笋期等多个方面的差异[19]。蓉城竹Ph. bissetii、紫竹Ph. nigra在《中国植物志》里属于刚竹组,马乃训[20]在《中国刚竹属》中将蓉城竹和紫竹划到水竹组,高素萍等[21]基于随机扩增多态性DNA (RAPD)技术对刚竹属种间近缘关系进行聚类分析,发现紫竹与水竹组的遗传距离较远。朱芳明等[22]基于内转录间隔区(ITS序列)重建刚竹属及其近缘属的系统发育树,认为紫竹、蓉城竹归为刚竹组得到了较高的支持率。本研究中,蓉城竹被聚在第Ⅶ类,与大多数刚竹组竹种聚在一起;紫竹被聚在第Ⅴ类而并没有聚在以水竹组为主的Ⅵ类,说明紫竹的表型性状低于刚竹属平均值,与第Ⅵ类的水竹组具有一定差异,这一分类结果与高素萍等[21]的研究结果相似。本研究将17种毛竹及其种下等级聚类于不同的分组,其中8个与毛竹原种更为相似的大型变种被聚为一类,龟甲竹和厚皮毛竹被聚为一类,元宝毛竹单独聚为一类,其余种下等级皆被聚在刚竹组为主的第Ⅶ类,聚类结果还将实心竹单独聚为一类,而其种下等级黑水竹、黄秆水竹被聚为以水竹组为主的第Ⅴ类。水竹和毛竹的特殊种下等级跟原种具有较大差异,这可能是由于某些特殊生物或非生物因素导致的。说明毛竹和水竹的种下等级变异丰富,易出现较为特殊的种下等级,遗传信息多样,具有较大的选育空间。
-
表型性状之间的相关性是建立数学模型和拟合方程的重要依据。当2个或多个竹种的表型性状之间存在统计上的显著相关性时,就认为它们之间存在一定的关联或依赖关系。这种关系有助于理解竹子的遗传多样性规律。胸径作为竹子重要的表型性状之一,与其他表型性状之间相关性显著,且易于测量,通常被作为自变量建立拟合方程。马乃训等[23]分别用胸径和秆质量、胸径和秆高、秆高和秆质量等3对相关关系建立了刚竹属部分品种的拟合方程,并获得了较高的拟合度(平均R2为0.974)。这一结果表明:围绕种建立的拟合方程的预测精度要高于围绕属建立的拟合方程。
竹子的纤维品质与其表型特征密切相关。纤维细胞的形态、大小、排列方式等都会直接影响纤维的品质。目前,关于竹子纤维性状和表型性状之间相关性的研究还相对较少,无法证明竹子表型性状和纤维性状之间的相关关系。严艳兵等[24]对美洲黑杨Populus deltoides的材性与生长性状进行了相关分析,发现纤维长度和宽度与生长性状之间呈显著正相关。张晓艳[25]则以黑杨派无性系为研究对象,揭示了其生长与材质性状之间的联系,发现纤维长度与抗压强度和抗弯强度、木纤维壁率与冲击韧性呈正相关,而与木材基本密度呈负相关。这些研究结果表明:种内的纤维性状与生长性状和一些微观表型性状之间确实存在较强的相关性。在本研究中,并未发现在属内的尺度上竹子表型性状和纤维性状之间存在显著的相关性,这可能是由于多种因素共同作用的结果。竹子的纤维品质不仅受到基因的控制,还受到环境条件、生长过程等多种因素的影响,这些因素可能掩盖了表型性状与纤维性状之间的关系。还需要更深入地研究竹子的生长发育过程和纤维形成机制,以揭示属内表型性状与纤维性状的内在联系。
-
根据测定的表型性状和纤维性状,刚竹属111种被分为7个大类,每类都具有明显特征,支持刚竹组和水竹组的分类。刚竹属表型性状变异系数为21.75%~67.72%,其中胸径、壁厚、枝下高节数、秆高具有高度变异,变异系数分别为62.72%、41.12%、40.45%、40.39%,胸径与其他性状相关性极显著;纤维主要性状变异低,变异系数为5.81%~38.23%。
毛竹类群种下等级的变种数量最多,共17个,其表型性状中胸径、壁腔比、秆高3个指标为高度变异;水竹类群内竹种表型性状中壁厚、壁腔比、枝下高节数属于高度变异且皆大于全属的变异程度。毛竹、水竹类群的变异程度较高,具有较大的选育空间。
Variation patterns of phenotype and fiber characteristics of bamboo species in Phyllostachys
doi: 10.11833/j.issn.2095-0756.20240606
- Received Date: 2024-11-11
- Accepted Date: 2025-08-17
- Rev Recd Date: 2025-08-16
- Available Online: 2025-11-26
- Publish Date: 2025-12-20
-
Key words:
- Phyllostachys /
- phenotypic traits /
- fiber traits /
- fitting equation
Abstract:
| Citation: | WANG Jiye, YUAN Jinling, ZHAO Dandan, et al. Variation patterns of phenotype and fiber characteristics of bamboo species in Phyllostachys[J]. Journal of Zhejiang A&F University, 2025, 42(6): 1165−1173 doi: 10.11833/j.issn.2095-0756.20240606

|
 DownLoad:
DownLoad: