







-
月季Rosa chinensis在世界各地广泛种植,花色艳丽,香气浓郁,适应性强,且品种繁多,目前已经成为城市绿化的生力军,同时也是重要的切花材料;其花、根和叶均可入药,且含有多种营养物质,具有较高的观赏价值、食用价值和药用价值[1−3]。月季栽培产业是中国当前经济大环境中的朝阳产业之一,取得了较大的经济和社会效益,但随着其栽培和应用面积的不断扩大,月季病害已经成为影响月季产业发展的重要因素。
丛赤壳属Nectria是丛赤壳科Nectriaceae的模式属,由Rossman在1989年建立,无性型为瘤座孢属Tubercularia。该属的真菌是广泛分布于北半球温带地区的典型木本植物病害病原菌,还能侵染一些草本植物,为害时在寄主植物枝条上形成近球形到球形、红色到暗红色的柔软肉质子实体[4−6]。国内已报道的该属病害不多,文献报道的有血红丛赤壳N. haematococca引起的大豆Glycine max茎腐病和黄檗Phellodendron amurense枝干腐烂病;假毛丛赤壳N. pseudotrichia引起的草莓Fragaria ananassa茎腐病、山茶Camellia japonica溃疡病和香椿Toona sinensis溃疡病;朱红丛赤壳N. cinnabarina引起的核桃Juglans regia枯枝病以及松丛赤壳菌N. cucurbitula引起的油松Pinus tabuliformis枯枝病[7−13]。此外,2015—2020年,国内学者报道了丛赤壳属的3个新物种,即N. zangii、榆生丛赤壳N. ulmicola和N. berberidis[5−6, 14]。结合现有资料,丛赤壳属近40种真菌,国内分布有17种[5−6, 14−15]。
目前,常见的月季真菌性病害有Calonectria blephiliae引起的月季丽赤壳褐斑病、灰葡萄孢Botrytis cinerea引起的月季灰霉病、蔷薇拟日规壳Gnomoniopsis rosae引起的月季黑斑病以及毡毛单囊壳菌Podosphaera pannosa引起月季白粉病等,病原菌侵染部位集中在叶片上,部分病原菌同时会侵染花和嫩芽嫩梢[3, 16−18]。对月季枝干病害研究较少,目前已报道的病原菌有蔷藤盾壳霉Coniothyrium fuckelii等[19]。在月季的栽培过程中,枯枝病对月季的伤害是致命的,病原菌多从枝梢或侧枝进行侵染致病后再从顶端逐渐向下蔓延到主干,严重时导致全株死亡。
2023年9月,在辽宁省宽甸县的绿化带上发现一种月季枯枝病,该病造成月季枝条干枯死亡,表皮皱缩,同时枝条上附着有橙色与黑色的扁球形子实体,根据其形态,初步认为由丛赤壳属真菌引起。鉴于该病害发病严重,且目前对月季枯枝病相关研究较少,本研究对该病害的病原物的种类进行了鉴定,同时对病原物的生物学特性进行了探究,并开展了药剂筛选试验,以期为该病害的防治与月季的合理管护提供理论参考。
-
从辽宁省宽甸县绿化带上采集病枝,装入信封后带回实验室。采用组织分离法分离病原菌:将带有分生孢子器的枝条在质量分数为2%次氯酸钠和体积分数为75%的酒精中分别浸泡1 min,然后用无菌水冲洗5 min、再用吸水纸吸干表面残余水分,最后将分生孢子器切开后用无菌针挑至马铃薯葡萄糖琼脂培养基(PDA)上,切开面朝下,置于培养箱中28 ℃避光培养,待形成菌落后,在体视显微镜下挑取尖端菌丝纯化多次,直至获得纯菌落[20]。
月季‘香紫绒’‘Mirandy’扦插苗购于市场,在实验室内栽培,待萌发出新枝并开花后,对其枝条消毒处理,采用烧伤(使用酒精灯外焰灼烧2~3 s,听到表皮破裂的响声为止)、刺伤(使用昆虫针刺透枝干表皮,刺孔7~8个)和无伤(健康)3种接种方式,以培养7 d的病原菌PDA平板的菌落边缘菌饼为接种体进行致病性测验,接种完成后将沾有无菌水的脱脂棉用封口膜缠绕在接种部位作保湿处理,48 h后除去。以空白PDA为对照,每处理10组重复[20]。
-
在OLYMPUS-SZX2-FOF体视显微镜下随机选择20个分生孢子器观察特征并测量大小;将分生孢子器制成纵切片后在OLYMPUS-BX51显微镜下对分生孢子器内部形态、分生孢子梗和分生孢子进行观察和拍照,并随机选择50个孢子进行测量。对获得的纯菌落形态及产孢情况进行持续观察,记录菌落形态和颜色等特征以及产器和产色素情况。
-
以培养7 d的病原菌纯菌丝为材料,使用博迈德植物基因组提取试剂盒抽提病原菌DNA基因组,并使用引物对ITS1(5′-CCGTAGGTGAACCTGCGG-3′)/ITS4(5′-TCCTCCGCTTATTGATATGC-3′)、LR0R(5′-GTACCCGCTGAACTTAAGC-3′)/LR7(5′-TACTACCACCAAGATCT-3′)、RPB2-b6F(5′-TGGGGYATGGTNTGYCCYGC-3′)/RPB2-b7R(5′-GAYTGRTTRTGRTCRGGGAAVGG-3′)和EF1-728(5′-CATCGAGAAGTTCGAGAAGG-3′)/EF1-986(5′-TACTTGAAGGAACCCTTACC-3′)分别扩增病原菌的内转录间隔区基因(ITS)、核糖体大亚基(LSU)、RNA聚合酶Ⅱ第2亚基基因(RPB2)和转录延伸因子(Tef-1α)等4个基因片段[20−22]。扩增反应体系和条件根据试剂说明书设置,所得产物经质量分数为1%的琼脂糖凝胶电泳检测后送至上海生工公司进行测序。获得序列信息后进行Blast相似性检索,根据每个基因的Blast结果下载相关序列后备用(表1)。系统发育分析在Phylosuite v1.2.3中进行,在MAFFT v7中独立排序后使用Gblocks自动剪切并串联,并进行适当的手动调整;利用ModelFinder v2.2.0根据贝叶斯信息准则(BIC)选择贝叶斯推理系统发育和最大似然系统发育的最佳拟合模型;使用MrBayes 3.2.6进行贝叶斯推理系统发育分析,其中初始25%的采样数据被丢弃作为预模拟运行,每1 000代采样4个马尔可夫链(MCMC),直到分裂偏差频率小于0.01;使用IQ-TREE进行最大似然系统发育分析,根据获得的最佳模型进行5 000次超快bootstrap;获得的系统发育树文件使用iTOL v7进行可视化,并通过Adobe Illustrator 2024进行处理[23−25]。此外,鉴于部分参考菌株缺失RPB2和(或)Tef-1α序列,因此根据以上方法构建一棵基于ITS-LSU 片段的二基因树,进一步佐证结果的可靠性。最后结合系统发育与形态学结果将序列上传至美国国家生物技术信息中心(NCBI)数据库。
病原菌 菌株 登录号 ITS LSU RPB2 Tef-1α N. antarctica CBS115033 HM484556 HM484560 – HM484516 N. asiatica MAFF 241439 HM484701 HM484563 JQ014140 – N. berberidicola 10779 KY498619 KY498621 – – N. berberidicola CBS 128669 JF832662 JF832712 – JF832538 N. berberidis XJAU 3355-1-2 OM701869 OM701965 – OM891361 N. cinnabarina CBS 125165 HM484548 HM484562 – HM484527 N. dematiosa CBS 126570 HM484557 HM484561 JQ014144 HM484534 N. dematiosa CBS 125125 HM484676 HM484717 – HM484645 N. eustromatica CBS 121896 HM534896 HM534896 HM534886 HM534875 N. magnispora CBS 129361 JF832664 JF832685 – JF832540 N. magnispora CBS 129362 MH865276 MH876720 – N. mariae CBS 125294 JF832629 JF832684 – JF832542 N. nigrescens CBS 125148 HM484707 HM484720 JQ014123 HM484672 N. polythalama CBS 128672 JF832638 JF832695 – JF832523 N. pseudocinnabarina CBS 129366 JF832642 JF832697 – JF832533 N. pseudotrichia CFCC 52122 MG231953 MG231974 MG231995 MG232016 N. tibetensis 10849 KY797280 KY797283 – KY797291 N. triseptata HAMS252485 KM026503 KM026504 – KM026506 N. ulmicola CFCC 52117 MG231959 MG231980 MG232001 MG232022 N. ulmicola CFCC 52118 MG231960 MG231981 MG232002 MG232023 Allantonectria miltina A.R.4391 HM484547 HM484572 – HM484524 说明:朱红丛赤壳N. cinnabarina;假毛丛赤壳N. pseudotrichia;榆生丛赤壳N. ulmicola。 Table 1. Sequences in the phylogenetic tree
-
以查氏培养基为基础培养基,以相同摩尔碳质量的蔗糖、葡萄糖、乳糖、果糖、麦芽糖和可溶性淀粉替换碳源,相同摩尔氮质量的酵母浸粉、蛋白胨、磷酸氢二铵、硫酸铵、硝酸钠和氯化铵替换氮源,制作不同碳氮源培养基,且分别以无碳无氮源为对照;把PDA培养基的pH分别调整至5.11、6.31、7.17、8.33和9.36制作不同pH培养基。在以上各种培养基中接种培养7 d的5 mm病原菌菌饼后,在25 ℃全黑暗培养,每组3次重复,每隔2 d采用十字交叉法测量菌落直径。并在培养箱中设置12、20、25、30和35 ℃的培养温度,使用PDA平板,其余同上,通过以上处理筛选病原菌的最适碳氮源、pH和温度[20, 26]。
-
以质量分数50%多菌灵悬浮剂(安徽广信农化)、75%百菌清可湿性粉剂(山东百农思达)、40%五氯硝基苯可湿性粉剂(山东鑫星)、80%代森锰锌可湿性粉剂(四川国光)和70%甲基硫菌灵可湿性粉剂(日本曹达)等5种杀菌剂作为供试药剂,使用二甲基亚砜(DMSO)溶解后加入熔融的PDA中,制作带药平板,以添加DMSO为对照。首先进行初筛,将其有效成分质量浓度分别调整为0.01、0.10、1.00、10.00和100.00 mg·L−1,接种病原菌菌饼后在25 ℃全黑暗下培养,每组3次重复。待对照满板后用十字交叉法测定菌落直径,计算菌丝生长抑制率,明确5种杀菌剂对病原菌菌丝的生长抑制作用,再根据初筛结果设置质量浓度同初筛方法进行复筛。数据用Excel 2019进行处理,计算毒力回归方程与抑制中浓度(EC50)[26]。菌丝生长抑制率=(对照菌落直径−处理菌落直径)/(对照菌落直径−5 mm)×100%。
-
通过 SPSS 24.0 单因素方差分析进行数据统计及差异显著性分析,显著性水平为 0.05;使用Origin 2022绘图。
-
通过持续观察病原菌分生孢子器在PDA平板上的生长情况,不断纯化培养后最终获得1株纯菌落菌株,且该菌株培养至后期产生的分生孢子器与受害枝条上的分生孢子器相似,将其转入斜面培养基保存,编号为R1007。
依据科赫氏法则开展致病性测验(图1),无伤(健康)接种与刺伤接种处理未发病,而烧伤处理组接种27 d后,接种位置表面出现颗粒状隆起且迅速扩展,后续逐渐转变为橙色,且相比于对照组,接菌组的烧伤部位表皮皱缩情况明显加剧。从发病部位再次分离出的病原菌形态学特征与病原菌R1007一致,以形成的分生孢子器为材料提取DNA进行分子验证,其ITS序列与R1007的ITS序列相似性达100%,结合形态学与分子证据可知R1007为月季枯枝病的病原菌。

Figure 1. Pathogenicity assay results
-
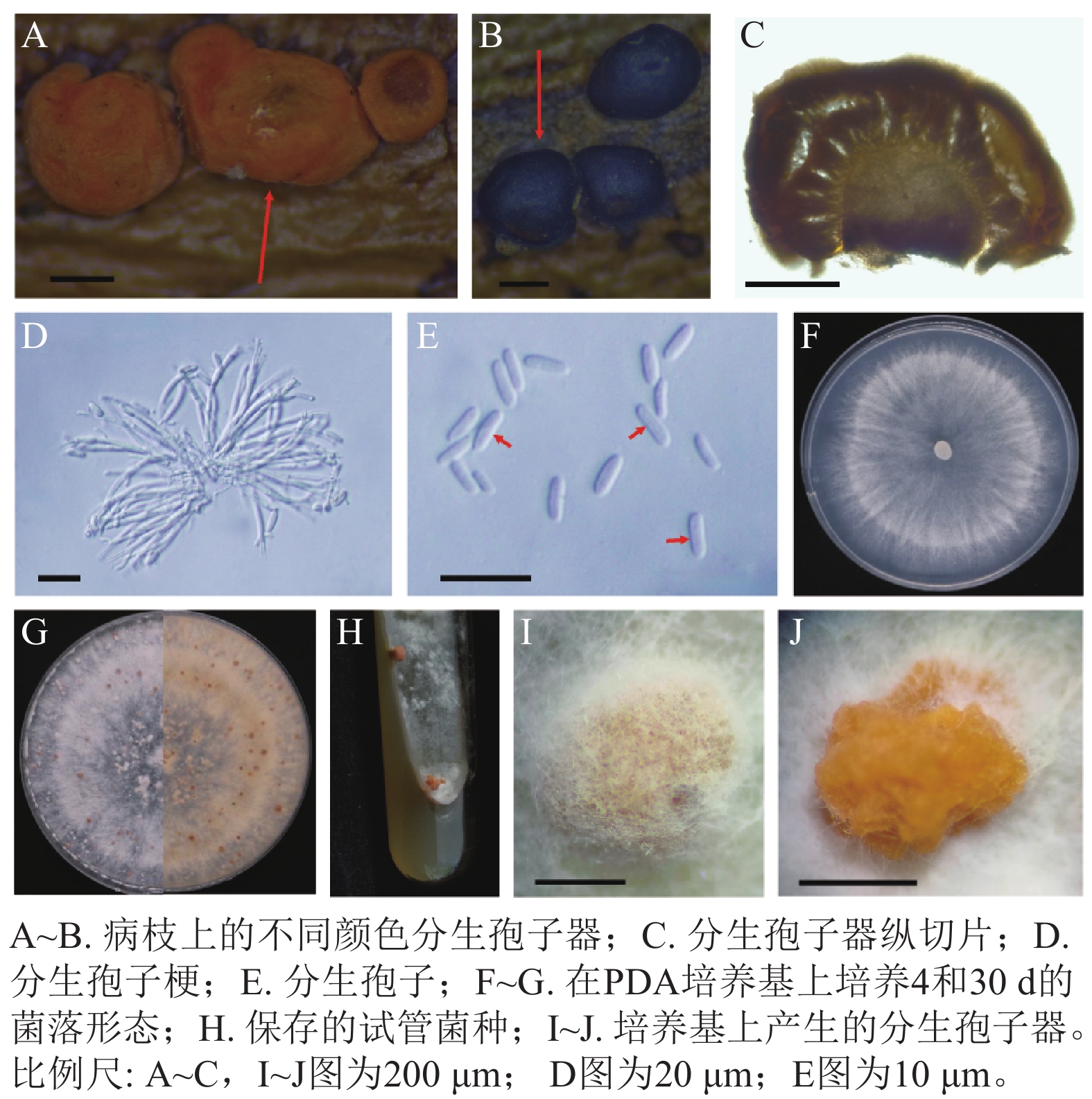
病原菌分生孢子器半埋生于寄主树枝表皮下,近圆形至不规则形,橙红色,老后转变为黑色,直径为240~630 μm (n=30),表面光滑,单生,内部脑状(图2A~C)。分生孢子梗无色透明,顶端具有小梗,不分叉(图2D)。分生孢子大小为(4.75~7.25) μm× (1.25~2.25) μm,平均为5.5 μm×2.0 μm (n=50),单孢,椭圆形,部分微弯曲,无隔(图2E)。未发现有性型。菌落在PDA培养基上呈辐射状生长,绒毛状,最初为纯白色,后期稍显污白色,质地均匀,边缘波浪状或平整,25 ℃全黑暗培养条件下最快6 d可以长满PDA培养基,之后经光照刺激,14 d 即可产生橙色的分生孢子器,菌落背面逐渐产生橙红色色素(图2F~J)。
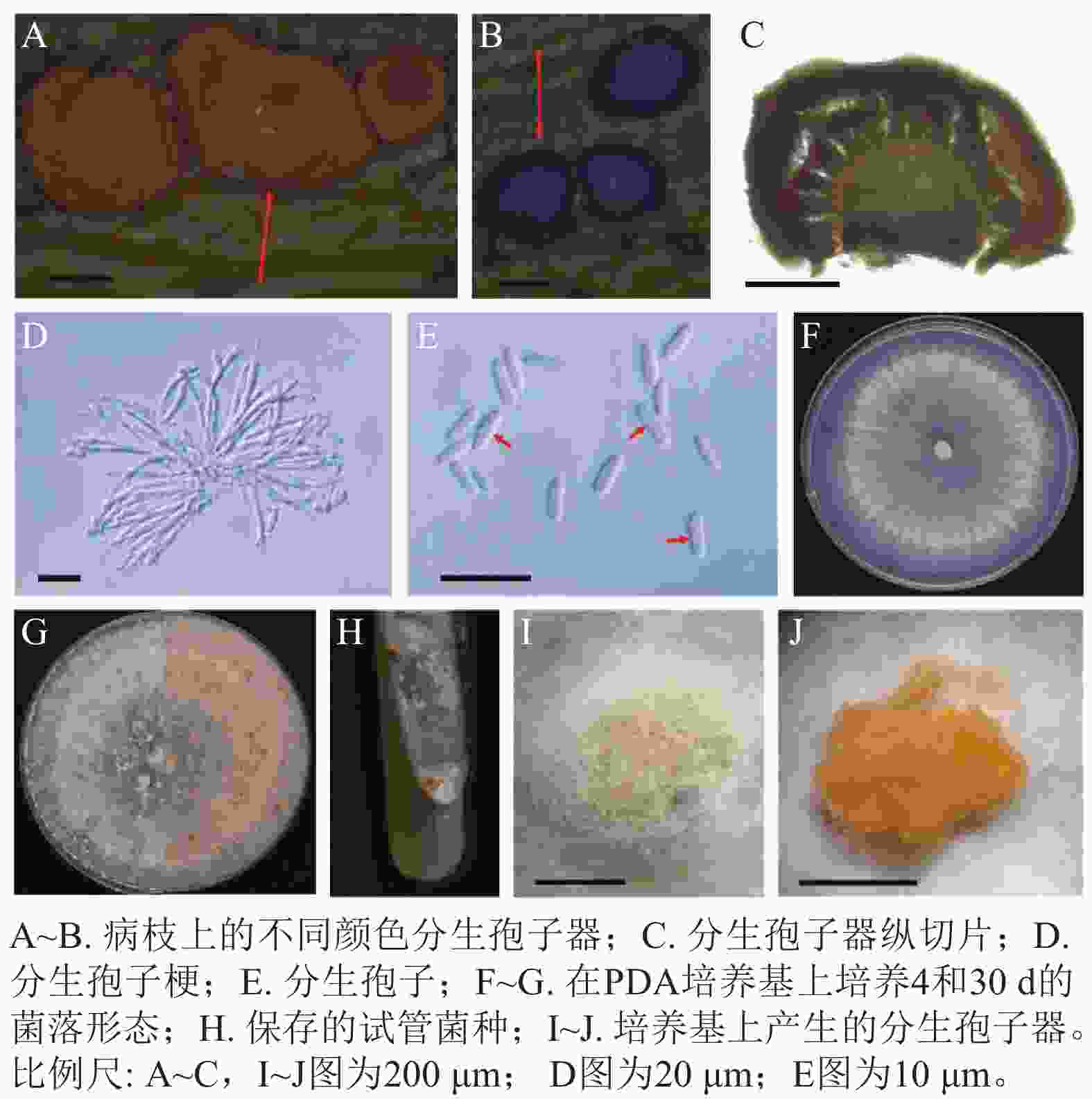
Figure 2. Morphological characteristics of the pathogen
-
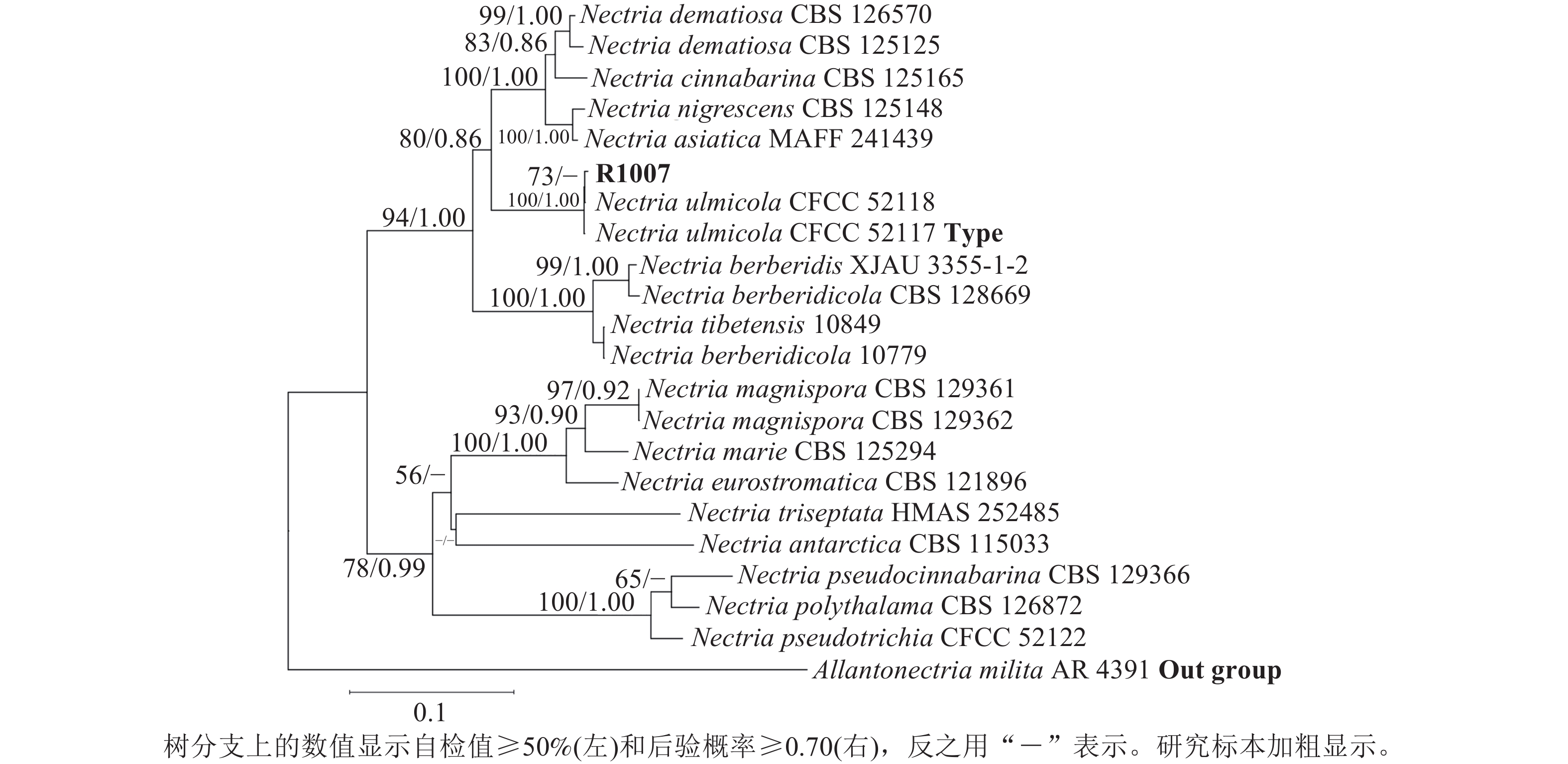
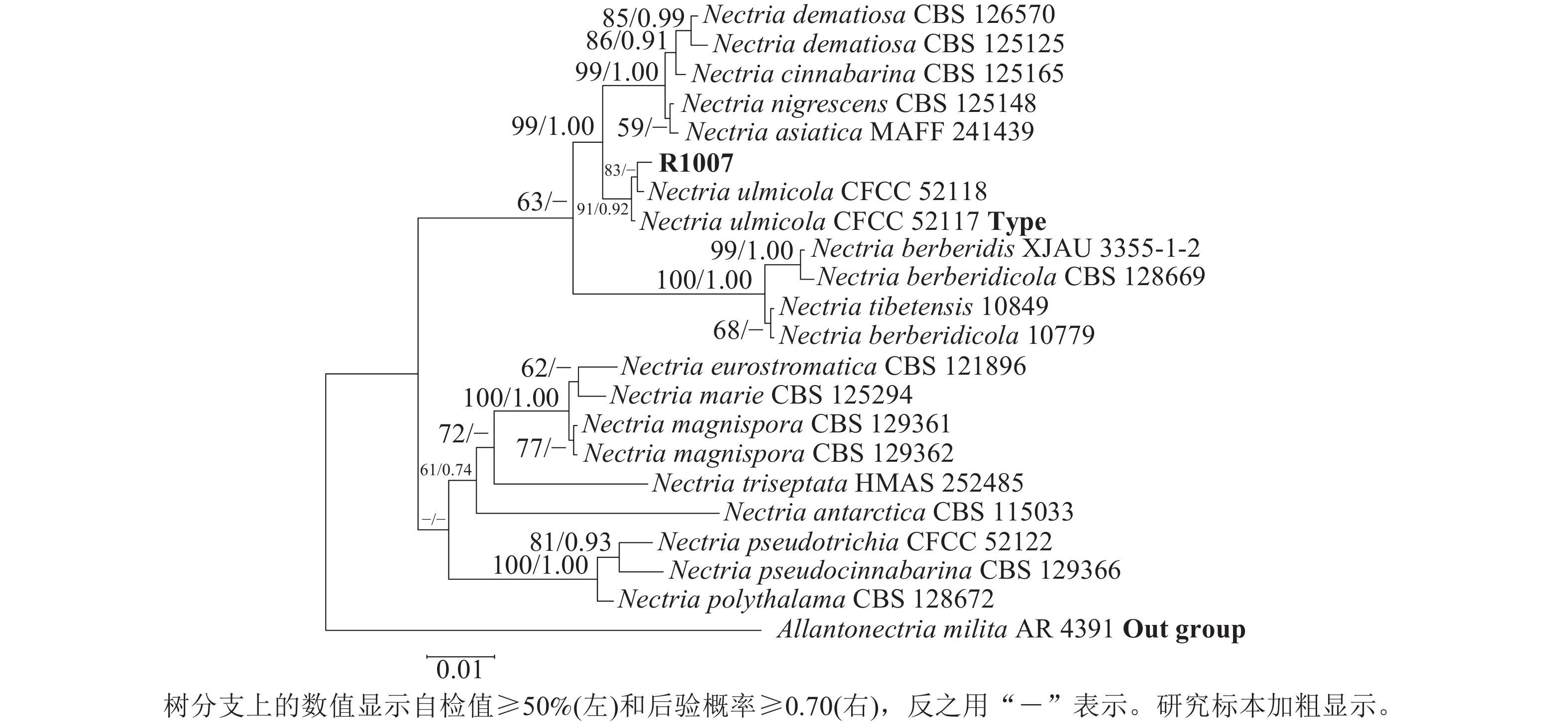
测序分别获得582 bp (ITS/PQ573830)、793 bp (LSU/PQ573831)、758 bp (RPB2/PQ589804)和437 bp (Tef-1α/PQ615445)的基因序列,Blast检索结果显示:与上述4条序列相似性最高的物种均为N. ulmicola。其中,ITS序列与其他序列相似性最高为100.00% (MG231959/N. ulmicola),得分最高为1 040 分(OK090962/N. ulmicola);LSU序列与其他序列相似最高为99.75% (MG231980/N. ulmicola),得分最高为1 445分(MG231980/N. ulmicola);RPB2序列与其他序列相似最高为99.86% (MG232001/N. ulmicola),得分最高为1 362分 (MG232001/N. ulmicola);Tef-1α序列与其他序列相似最高为99.77% (MG232022/N. ulmicola),得分最高为795分(MG232022/N. ulmicola)。2个数据集各自构建的贝叶斯树与最大似然树拓扑结构一致,因此仅分析贝叶斯树。菌株R1007与榆生丛赤壳的模式菌株CFCC52117以及CFCC52118聚为一支,且得到了充分的支持(图3)(BS/PP=100/1.00);二基因树的拓扑结构与四基因树在拓扑结构和支持率上具有一定差异(图4),但R1007仍与CFCC52117和CFCC52118形成一个稳定分支(BS/PP=91/0.92)。综上,结合形态学特征、Blast检索结果以及系统发育推断,确定本研究中引起月季枯枝病的病原菌为榆生丛赤壳。
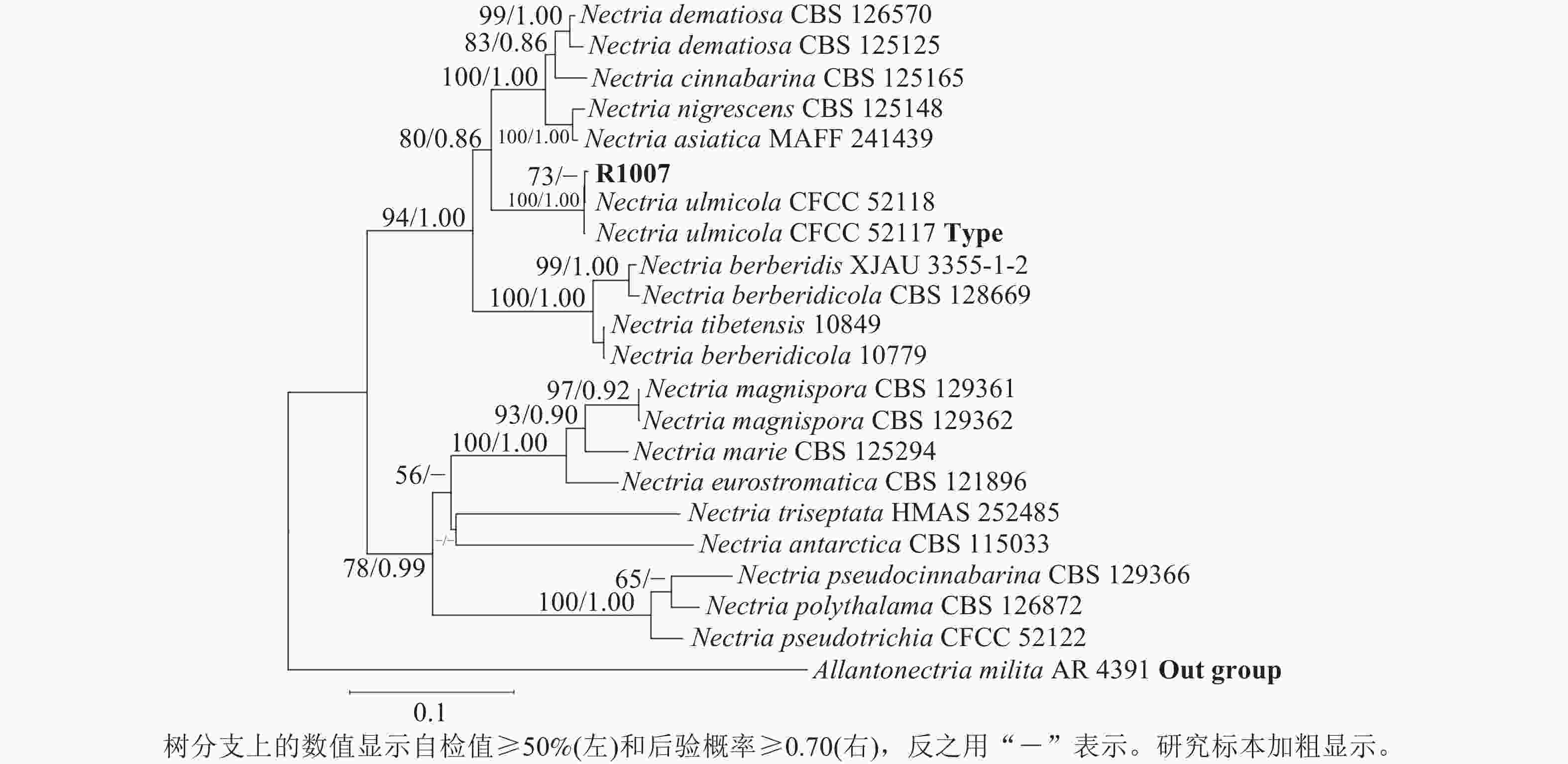
Figure 3. A four-gene phylogenetic tree based on ITS-LSU-RPB2-Tef-1α
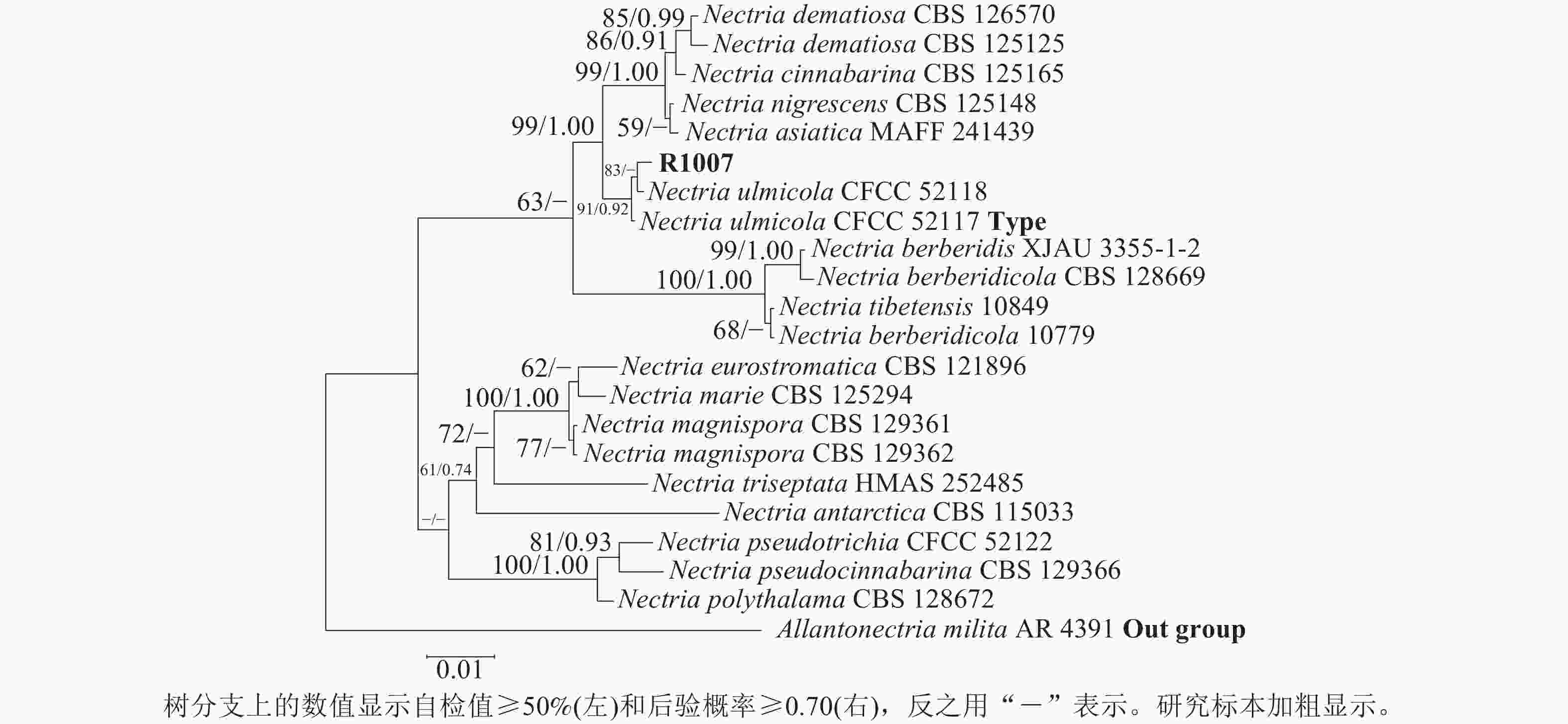
Figure 4. A two-gene phylogenetic tree based on ITS-LSU
-
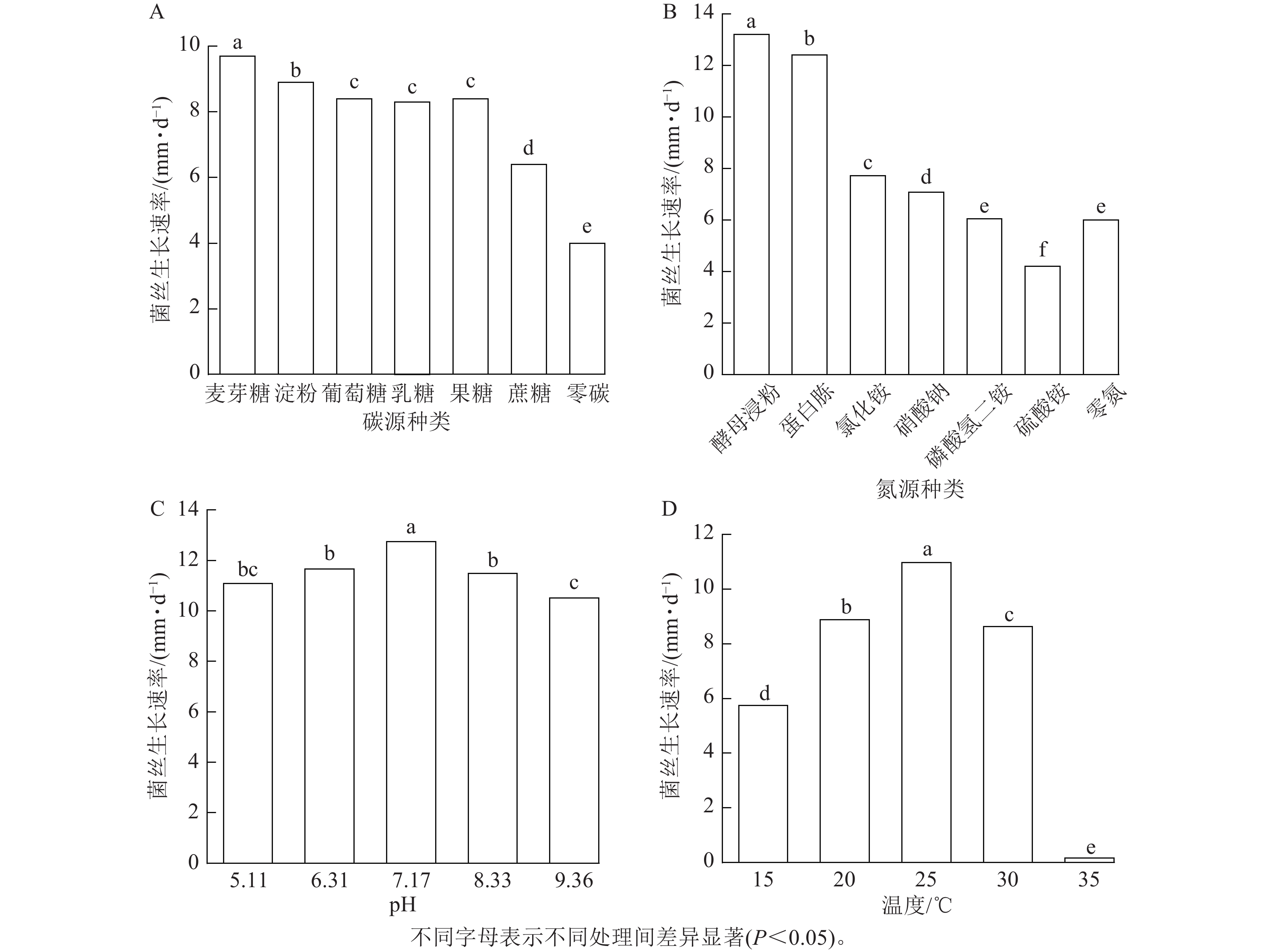
由图5A可见:病原菌对碳源利用广泛,单糖、二糖、多糖都能利用。碳源为麦芽糖时病原菌长势最好,其菌丝生长速率为9.7 mm·d−1,生长第8天即可以布满整个平板。对可溶性淀粉的利用能力次之,对葡萄糖、乳糖和果糖的利用效果相近,菌丝生长速率分别为8.4、8.3和8.4 mm·d−1,对蔗糖的利用能力最差,生长速率为6.4 mm·d−1。综上,麦芽糖更适合病原菌的生长。
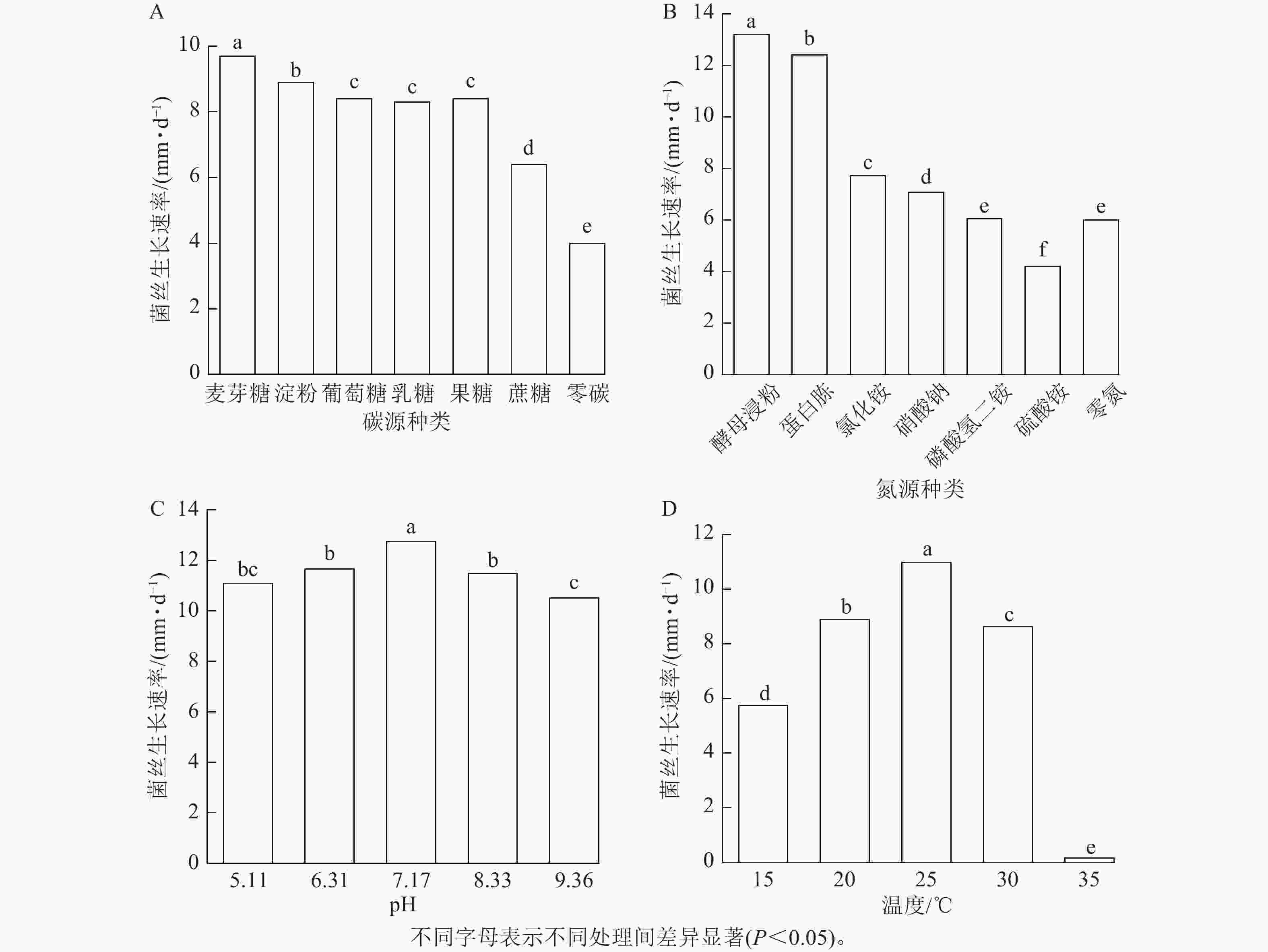
Figure 5. Mycelial growth rate under different conditions of carbon sources (A), nitrogen sources (B), pH (C), and temperature (D)
-
由图5B可见:不同氮源对病原菌菌丝的生长影响区别明显。在以酵母浸粉为氮源时菌丝生长速率可以达13.2 mm·d−1,6 d即可长满整个平板。蛋白胨的促进效果稍弱,菌丝的生长速率为12.4 mm·d−1。对其他几种无机氮源的利用效果较差,菌丝生长速率为4.2~7.7 mm·d−1,氮源为硫酸铵时甚至次于无氮处理。综上,酵母浸粉对病原菌的促进效果最佳,且病原菌对有机氮源表现出明显的偏好性。
-
由图5C可见:在pH为5.24~9.36的范围下,病原菌菌丝均能正常生长,且长势良好。pH=7.17时,菌丝的生长速率最快,为12.7 mm·d−1,不同处理下菌丝生长速率均超过10 mm·d−1,pH波动对病原菌的生长影响不大,可以看出病原菌对酸碱度适应能力较强,但偏好微碱性环境。
-
由图5D可见:在25 ℃时,菌丝生长速率最快,生长速率达10.97 mm·d−1,显著高于其他温度处理。虽然病原菌在15~35 ℃范围内均能生长,但在35 ℃下生长速率极低,生长速率为0.16 mm·d−1,几乎没有生长。综上,病原菌最适生长温度为25 ℃,35 ℃接近其致死温度。
-
供试的5种不同药剂对病原菌菌丝的生长均有抑制效果(表2)。其中抑菌效果最好的是50%多菌灵,其EC50为0.338 8 mg·L−1,其次是75%百菌清和40%五氯硝基苯,EC50分别为8.820 0、9.200 3 mg·L−1,70%甲基硫菌灵抑菌效果最差,EC50高达373.594 0 mg·L−1。
杀菌剂 毒力回归方程 相关系数(R2) 抑制中浓度(EC50)/(mg·L−1) 75%百菌清 y = 1.238 2x + 3.828 8 0.973 1 8.820 0 50%多菌灵 y = 4.442 2x + 7.088 2 0.988 5 0.338 8 70%甲基硫菌灵 y = 0.740 4x + 3.095 4 0.993 1 373.594 0 80%代森锰锌 y = 1.730 6x + 2.634 7 0.932 6 23.270 2 40%五氯硝基苯 y = 2.305 5x + 2.778 0 0.991 0 9.200 3 说明:杀菌剂前百分比为有效成分的质量分数。y为抑制率,x为杀菌剂质量浓度的对数值。 Table 2. Inhibitory effect of 5 fungicides for R1007
-
月季具有重要的应用价值,本研究结合形态学特征、多基因系统发育以及致病性测定,将发生于辽宁省宽甸县的月季枯枝病病原菌鉴定为榆生丛赤壳;单因素试验表明:榆生丛赤壳的最佳碳氮源分别为麦芽糖和酵母浸粉,病原菌喜好pH为7.17的弱碱性环境,最适生长温度为25 ℃;多菌灵对榆生丛赤壳菌丝生长抑制效果最强,且EC50较低,可用于该病害的防治。榆生丛赤壳是中国学者分离自榆树Ulmus pumila枝干上的一个新物种[5]。由于寄主和个体间的差异,形态学特征与原文描述稍有不同,病原菌孢子平均大小略小,但其他分类学特征与分子生物学结果支持R1007为榆生丛赤壳这一结论,特别是R1007与榆生丛赤壳模式菌株CFCC52117的ITS序列相似性为100%[5]。在致病性测验中只有烧伤处理组发病结果与病原菌的致病性和寄主植物的抗病性有直接关系。丛赤壳属真菌为弱致病菌,难以直接穿透表皮或通过气孔进行侵染,因此无伤接种条件下接种失败。在刺伤处理下植株并未发病,原因除病原菌致病性较弱外,还可能是由于枝条遭到创伤之后产生的应激响应,触发系统获得性抗性进一步增强了植株抗病能力,且除去保湿棉后将接种材料直接暴露于室内,给植株补充水分时直接浇至盆中,接种部位一直处于干燥状态使得病原菌无法正常定殖甚至快速死亡,从而导致接种失败[27]。而在烧伤处理下植株发病且病原菌很快产生分生孢子器,这可能是烧伤引发植物细胞的广泛死亡,导致大量的营养物质释放供病原菌生长与繁殖,且因为细胞严重受损,植物的天然免疫反应也不再有效。另外,推测榆生丛赤壳腐生性极强,侵入受伤组织后定殖成功并逐渐适应,产生大量的毒素、分解酶等物质,进一步扩展至健康组织,最终导致枝条干枯,这是植物病原真菌经长期进化而发展出的侵染宿主策略之一,从而克服和规避宿主的防御反应,以此促进自身的繁殖和侵染[28]。这也说明了在园林作业中合理的管护至关重要,通过科学的管理措施最大程度地减少物理性伤害的发生,从而提高植株的健康水平和抗病能力。在油松枯枝病的研究中接种材料在保湿室内培养,香椿溃疡病接种时在人工气候箱内培养,病原菌可以保持较高的侵染能力,同时后者直接移除香椿枝干表皮,造成植株抗性极大下降[11, 13]。
病原菌的生物学特性与病害的发生与流行密切相关,明确病原菌的生物学特性在探索病原菌致病机制和筛选生防菌等方面具有关键意义[20]。多数丛赤壳属病害研究仅鉴定了病原物,对该属病原菌生物学特性的研究尚属空白,防治研究也少[8−12]。已知N. asiatica、N. cinnabarina和N. nigrescens在PDA上的最佳生长温度为25 ℃,多菌灵是对油松枯枝病病原菌N. cucurbitula抑制效果最好的杀菌剂之一,上述结果与本研究类似[4, 13]。
-
引起辽宁省宽甸县的月季枯枝病病原菌为榆生丛赤壳;该病原菌的最佳碳氮源分别为麦芽糖和酵母浸粉,喜好弱碱性环境,最适生长温度为25 ℃,推荐将多菌灵应用于该病害的防治。本研究是国内首次报道由榆生丛赤壳引起的月季枯枝病,同时也是第一次对丛赤壳属真菌引起的病害进行系统研究,对掌握该属病害发生和传播的季节、区域以及病原菌与环境之间的关系等具有一定的参考意义。
Identification, characterization and fungicide screening of the causal pathogen of branch blight in Rosa chinensis
doi: 10.11833/j.issn.2095-0756.20250213
- Received Date: 2025-03-19
- Accepted Date: 2025-06-11
- Rev Recd Date: 2025-06-10
- Available Online: 2026-04-02
- Publish Date: 2026-04-02
-
Key words:
- Rosa chinensis branch blight /
- new disease /
- biological characterization /
- fungicide screening
Abstract:
| Citation: | WU Chonggao, ZHU Tong, CUI Yunlong, et al. Identification, characterization and fungicide screening of the causal pathogen of branch blight in Rosa chinensis[J]. Journal of Zhejiang A&F University, 2026, 43(2): 361−369 doi: 10.11833/j.issn.2095-0756.20250213

|
 DownLoad:
DownLoad: