

 下载:
下载:

-
新型城镇化建设的推进为苏南水网地区城乡建设带来了经济的快速发展,但同时也给区域生态环境保护带来压力,蓝绿交织的水网地域特色景观正面临逐渐消失的危机。蓝绿空间是水网地区自然生态空间的重要组成部分,也是水网地区地域特色景观的载体[1]。水网地区的耕地以水田为主,具有涵养水源、调蓄雨洪的生态功能[2],与林地、草地等共同构成绿色空间。蓝色空间则指河流、湖泊、湿地和坑塘等共同构成的水域空间[3]。目前,国内外学者对于蓝绿空间的研究多集中于景观格局演变[4−5]、空间营建与调控[6−8]、冷岛效应[9]以及健康服务功能评价[10−11]等方面,研究区域多集中于城市群区域或城市市域范围,对具有水网地域特征的城镇地区蓝绿空间格局的研究相对较少。
苏南水网地区属于太湖流域,具有得天独厚的地理条件,形成了水绿共生的整体格局。快速城镇化在促进苏南水网地区社会经济高速发展的同时,量大面广的居住区、工业园区等人工开发建设活动对蓝绿空间的干扰程度日趋增强。据Landsat 8影像数据统计,2000—2022年,苏州市吴江区建设用地面积由14653.57 hm2增加至31008.33 hm2,而蓝绿空间面积由107543.79 hm2减少至91790.00 hm2。建设用地不断扩张使得蓝绿空间面临着面积下降、景观格局破碎、生态效益减弱等问题,水网地区的可持续发展受到冲击。模拟预测蓝绿空间未来发展格局,分析蓝绿空间格局未来发展趋势,总结蓝绿空间格局未来演变特征,是保护和恢复水网地区生态环境,促进苏南水网地区城乡可持续发展的关键。
本研究以具有典型苏南水网特征的苏州市吴江地区为代表,在总结2000—2020年蓝绿空间格局演变规律的基础上,借助CA-Markov模型,模拟2030年吴江南片区蓝绿空间土地利用多情景发展状态,并对多情景发展的蓝绿空间演变进行定量分析,结合形态学空间格局分析(MSPA)和景观连通性评价科学解读蓝绿空间景观生态结构与生态格局特征,为水网地区蓝绿空间的保护提供理论帮助与数据支持,以期促进水网地区城乡可持续发展。
-
吴江南片区隶属于江苏省苏州市吴江区,地处江苏省东南部,位于31°02′~31°13′N,120°51′~120°42′E,包括了横扇街道、平望镇、七都镇、震泽镇、盛泽镇和桃源镇,面积约606.1 km2。该片区自然环境条件优越,地势平坦,属于典型的水网平原,耕地、林地、河流和湖泊资源丰富,形成了水网地区蓝绿交错的独特格局。但在快速城镇化建设过程中,吴江区蓝绿空间面临着面积缩减,功能脆弱,空间破碎化等问题。
-
①2000、2010、2020年吴江区遥感影像从地理空间数据云平台Landsat 8卫星获取,分辨率均为30 m。以此为基础数据,结合研究区实地调查结果,使用ENVI 5.3对遥感影像进行监督分类和人工目视解译,根据GB/T 21010—2017《土地利用现状分类》的分类依据和标准,将土地利用类型划分为耕地、林地、草地、水域和建设用地等5类,经检验3期图像精度均达到85%以上。②行政区划边界数据来源于最新发布的吴江区行政区划图,通过地图数字化处理得到行政区划边界矢量数据。③研究区数字高程(DEM)数据为30 m分辨率的ASTER GDEMV 3数据,来源于中国科学院地理空间数据云(
http://www.gscloud.cn/ ),坡度数据由DEM高程数据提取得到。④城镇居民点数据来源于国家统计局官方网站发布的统计用区划和城乡划分代码等相关信息,将其整理成格式化文本数据,通过地理编码等处理得到村镇居民点空间分布矢量点数据。 -
CA-Markov模型由Markov模型和离散动力学元胞自动机(cellular automata,CA)模型[12−13]构成,能够基于土地利用转移概率矩阵与土地利用变化适宜性图集,模拟预测未来格局的演变趋势。本研究基于IDRISI 7.0中的CA-Markov模型,对吴江南片区的土地利用格局变化进行预测,将模拟得到的吴江南片区2020年土地利用类型图与实际情况进行比对,检验模型精度。在CA-Markov模型模拟可信度较高的情况下,以2020年吴江南片区土地利用图为基础年数据,分别选取3种情景下的转移概率和适宜性图集运行CA-Markov模型,得到2030年吴江南片区多情景土地利用模拟预测图像。
-
多标准评价模型(multi-criteria evaluation,MCE)由约束条件和适宜因子2个部分组成,是土地利用多情景模拟转换规则的核心。本研究借鉴前人的研究结果[14−16],并结合研究区实际情况和模拟效果,从自然因素和社会因素2个方面选择高程、坡度、距村镇居民点距离、距河流和湖泊距离及人均国内生产总值(GDP)共6类因子,作为制作适宜性图集的主要影响因子,约束条件则根据不同的发展情景进行分别设置。
-
情景分析旨在描述和分析事物发展的各种可能性,对比不同发展路径下的状态,为科学规划决策提供依据[17]。本研究基于CA-Markov模型和MCE模型,综合考虑研究区不同情景的发展需求及转换规律,参照《苏州市国土空间总体规划(2021—2035)》《苏州市吴江区国土空间规划近期实施方案》《长江三角洲区域一体化发展规划纲要》等相关文件,设定吴江南片区3种土地利用变化模拟情景:①自然发展情景。自然发展情景是基于2010—2020年吴江南片区的土地利用变化趋势,按照目前的城镇化发展模式和土地利用转移速率,不对耕地、林地、草地和水域与建设用地间的相互转换进行限制,模拟自然发展情景下2030年吴江南片区蓝绿空间土地利用演变状况。②生态保护优先情景。生态保护优先情景是以推动生态文明建设为首要,结合苏南水网地区水网密布,蓝绿交织的地域特征,考虑将永久基本农田保护区和生态保护红线设置为限制转化区,对建设用地随意侵占耕地、林地、草地及水域实施管控,降低蓝绿空间向建设用地的转化速率。③城镇发展优先情景。该情景是以适应苏南水网地区的经济与城镇发展需求为前提,土地利用情景设置中的新增建设用地有可能会占用原有建设用地范围周边的部分耕地、林地、草地及水域。
-
形态学空间格局分析(morphological spatial pattern analysis, MSPA)是一种图像处理方法,可以识别目标像元集与结构要素之间的空间拓扑关系,能够有效地确定景观的类型和结构[18]。本研究借助MSPA分析蓝绿空间格局的形态变化及演变趋势,基于3种模拟情景下得到的2030年吴江南片区土地利用类型图,结合当地实际情况,利用ArcGIS平台的重分类工具将生物多样性潜在力较好的耕地、林地、草地和水域分类为前景像元,建设用地作为背景像元,运用Guidos Toolbox软件的MSPA工具对数据进行腐蚀、膨胀等运算,得到3种模拟情景下2030年吴江南片区蓝绿空间景观组成类型及其占比。
-
景观连接度是评价景观结构单元之间连续性的重要指标[19],也是度量水网乡村蓝绿空间连通性的关键指标,数值越高,越有利于蓝绿空间格局整体生态效益的发挥。可能连通性指数(PC)综合考虑了物种迁移能力和扩散概率,是现阶段较为成熟的评价指标[20]。
选取对本研究有重要意义的核心区景观要素,运用Conefor 2.6,参考相关文献[15, 21−22]并结合本研究的研究尺度,设置斑块连通距离阈值为500 m,连通的概率根据实际情况设为0.5,对研究区的核心区进行景观连接度评价,筛选出3种模拟情景下景观连接度最大的9个斑块。按照连接重要性指数($ \mathrm{d}\mathrm{I} $)评价各核心区斑块在整体景观中的重要值,作为评判核心区景观连接度的指标,记作可能连通性重要值(dPC),反映斑块重要性[23]。
-
利用2000、2010年的土地利用类型数据,获得2000—2010年土地利用类型的转移概率矩阵(表1)。以2010年的土地利用类型图作为基础年数据,以10 a为迭代周期,比例误差设为0.15,预测2020年吴江区南片区土地利用类型图(图1),与2020年实际土地利用分布图进行对比验证,得到Kappa值为0.886 2,表示模拟的可信度较高,可用来预测研究区后续土地利用变化情况。
表 1 2000—2010年吴江南片区土地利用类型转移概率矩阵
Table 1. Probability matrix of land use type transfer in southern Wujiang District from 2000 to 2010
2000年土地利用类型 2010年土地类型转移概率/% 耕地 林地 草地 水域 建设用地 耕地 63.74 0.03 0.02 4.58 31.63 林地 6.02 68.85 0.19 14.30 10.64 草地 0.01 0.73 72.42 26.83 0.01 水域 13.93 0.05 0.01 76.64 9.37 建设用地 19.95 0.06 0.01 4.21 75.77 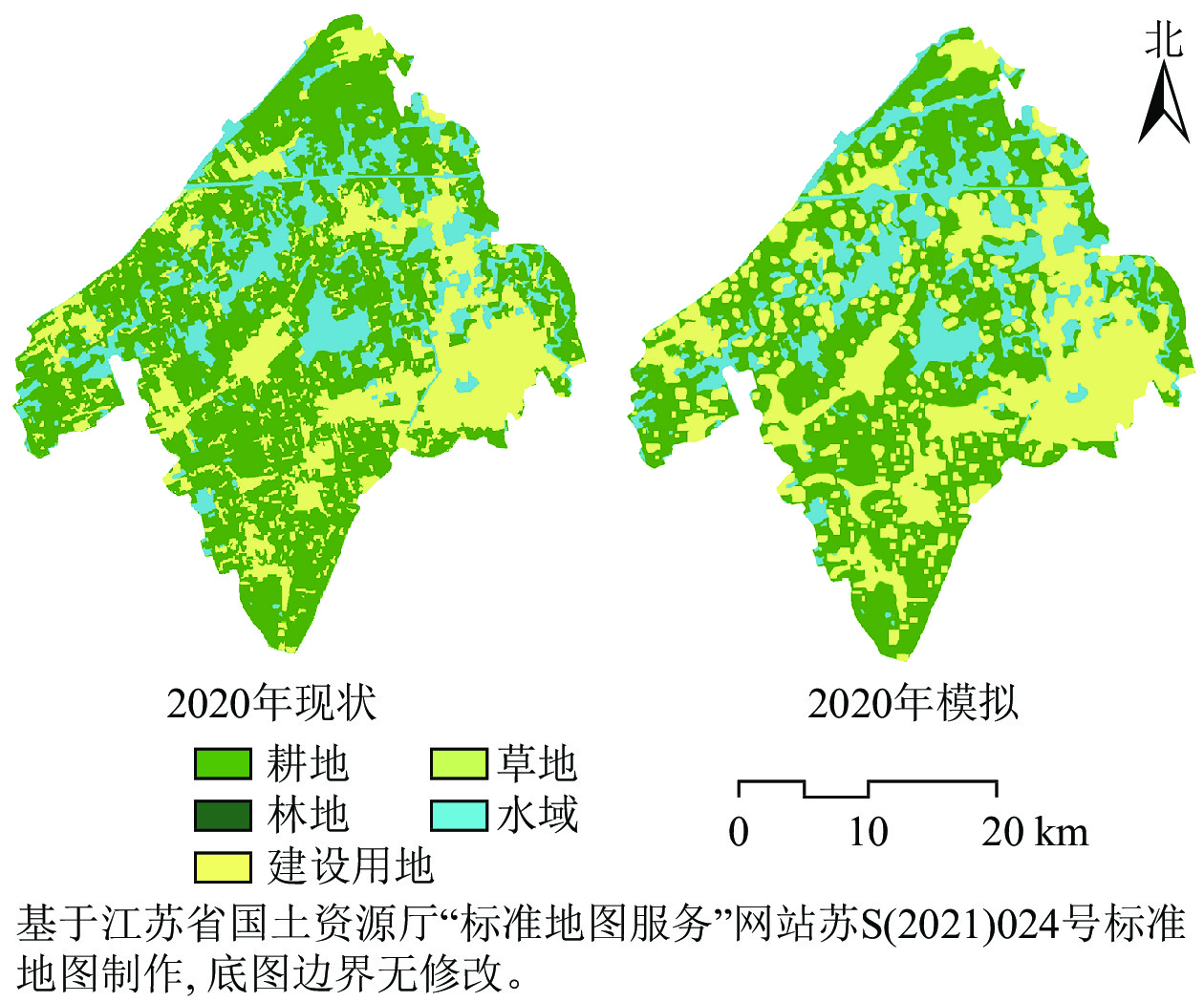
图 1 2020年吴江南片区土地利用现状与模拟
Figure 1. Landuse status and simulation of southern Wujiang District in 2020
-
以2020年吴江南片区土地利用空间分布图为基础数据,通过MCE模型,分析各类土地利用适宜性及吴江南片区2010—2020年的土地利用类型转移概率矩阵(表2),并限制不同情景下各类土地利用的约束条件,利用CA-Markov模型预测3种情景下2030年吴江南片区蓝绿空间土地利用的变化及各类土地利用的面积、比例(图2,表3)。
表 2 2010—2020年吴江南片区土地利用类型转移概率矩阵
Table 2. Probability matrix of land use type transfer in southern Wujiang District from 2010 to 2020
2010年土地利用类型 2020年土地利用类型转移概率/% 耕地 林地 草地 水域 建设用地 耕地 65.13 0.01 0.01 3.44 31.41 林地 0.90 75.78 0.13 13.45 9.74 草地 15.42 0.01 84.55 0.01 0.01 水域 13.37 0.01 0.01 75.57 11.04 建设用地 14.27 0.01 0.01 5.13 80.58 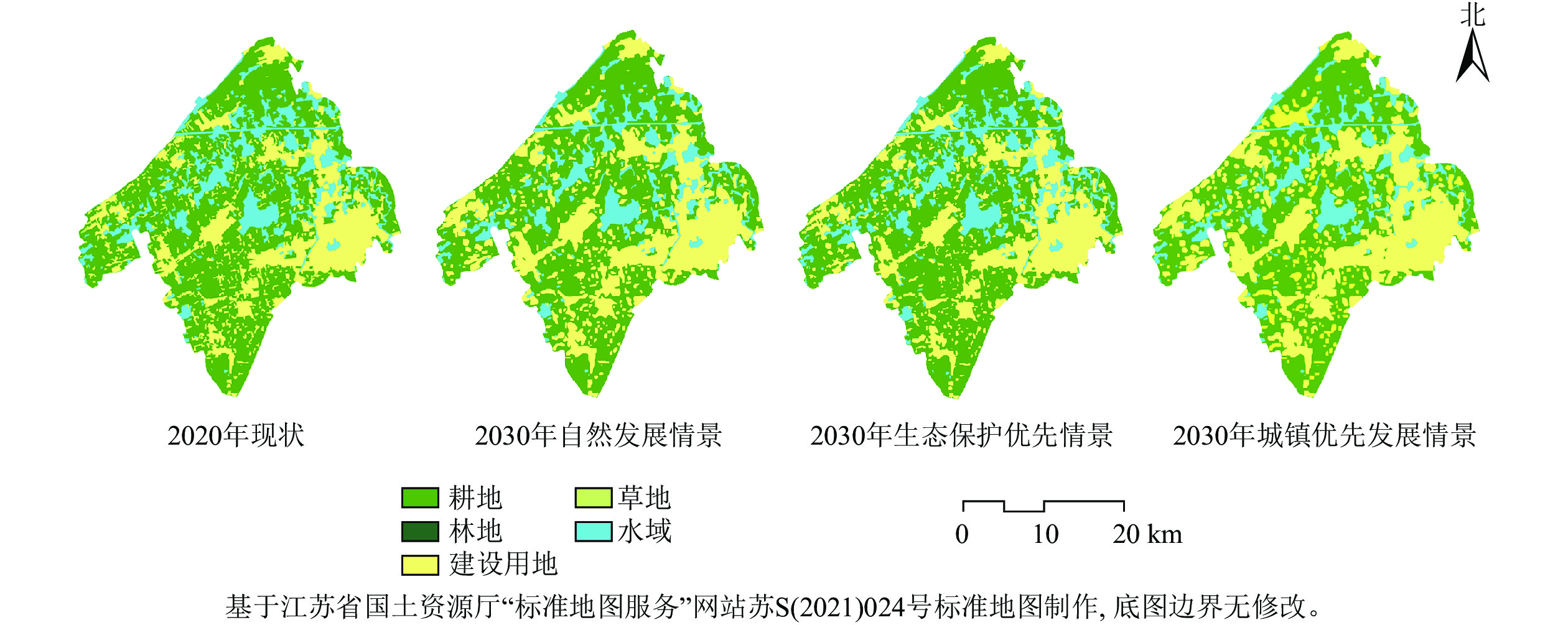
图 2 吴江南片区蓝绿空间土地利用变化
Figure 2. Landuse change of blue and green space in southern Wujiang District
表 3 2020—2030年吴江南片区蓝绿空间土地利用数据
Table 3. Land use data of blue and green space in southern Wujiang District from 2020 to 2030
类型 2020年现状 2030年自然发展情景 2030年生态保护优先情景 2030年城镇发展优先情景 面积/hm2 占比/% 面积/hm2 占比/% 面积/hm2 占比/% 面积/hm2 占比/% 耕地 33 837.03 55.84 31 826.79 52.52 33 058.35 54.36 28 946.79 47.84 林地 142.29 0.23 129.78 0.21 148.59 0.24 30.41 0.05 草地 19.44 0.03 17.64 0.03 22.14 0.04 17.64 0.03 水域 9 838.98 16.24 8 237.16 13.59 9 886.59 16.26 7 697.16 12.72 建设用地 16 763.22 27.66 20 390.76 33.65 17 693.10 29.10 23 810.58 39.36 由图2和表3可知:2020—2030年,3种发展情景下的耕地、林地、草地和水域的土地利用面积及占比变化差异较大。①自然发展情景下,耕地、林地和水域面积随时间推移而缩减,分别减少了3.32%、0.02%和2.56%,建设用地面积持续增加,增幅约为5.99%,基本延续过去10 a吴江南片区土地利用的发展规律。②生态保护优先情景下,基于生态文明建设的需求,限制了耕地、林地和水域等向建设用地的无序转化,蓝绿空间土地利用相对稳定,耕地、林地、草地和水域的面积及占比有所增加。相对于自然发展情景,生态保护优先情景下耕地和水域增幅最为显著,分别为1.84%和2.67%,建设用地增幅减少,从3 627.54 hm2减少到929.88 hm2。③城镇优先发展情景下,蓝绿空间土地利用发生了较为剧烈的变化,相较于生态保护优先情景,城镇发展优先情景下耕地、林地面积减幅较大,其占比分别下降了6.59%、0.19%,水域面积减少了2 189.43 hm2,而建设用地面积显著增加,增幅达10.25%。
-
由表4和图3可知:2020—2030年,①自然发展情景下的7类蓝绿空间景观要素基本延续前10 a的自然发展规律,核心区、边缘区等景观要素占比呈现出整体下降的趋势,孤岛下降率明显,孤岛面积从54.54 hm2减少至30.30 hm2,变化率为−44.44%。孤岛面积下降说明可供生物在吴江南片区蓝绿空间之间迁移的“踏脚石”数量正在减少。②生态保护优先情景下,蓝绿空间景观要素多呈现良好的发展态势,其中核心区的增幅最大,增长了490.87 hm2。核心区可作为蓝绿网络中的源地,在生物多样性保护方面发挥着重要作用,核心区面积的增加反映了吴江南片区蓝绿空间的生态功能有所增强。生态保护优先情景下的桥接区和环道区占比分别为0.38%和0.48%,较自然发展情景分别减少了0.21%和0.05%。桥接区是连通各核心区间的廊道,环道区是核心区内部的廊道,两者是物种迁移、能量流动的重要通道,桥接区、环道区数量的减少反映了吴江南片区蓝绿空间景观破碎化程度的改善。③城镇优先发展情景下,受建设用地沿分布区域向内外扩张的影响,蓝绿空间核心区面积大幅减小,核心区内部的孔隙面积持续增加。孔隙是蓝绿空间核心区内部的孔洞,孔隙面积增幅越大代表蓝绿空间核心区内部零散的建设用地越多,蓝绿空间的破碎化程度越高。城镇优先发展情景下孔隙占比较生态保护优先情景增长了2.77%,表明城镇优先发展情景下吴江南片区蓝绿空间正面临着外部面状、内部点状的生态破坏,对维持蓝绿空间景观格局稳定造成不利影响。
表 4 2020—2030年吴江南片区蓝绿空间MSPA景观类型变化
Table 4. Changes of MSPA landscape types in the blue and green space of southern Wujiang District from 2020 to 2030
类别 2020年实际
面积/hm2自然发展情景 生态保护优先情景 城镇优先发展情景 2030年预测
面积/hm2面积变化/
hm2变化率/
%2030年预测
面积/hm2面积变化/
hm2变化率/
%2030年预测
面积/hm2面积变化/
hm2变化率/
%核心区 31 312.51 29 258.13 −2 054.37 −6.56 31 803.37 490.87 1.57 23 804.05 −7 508.46 −23.98 孤岛 54.54 30.30 −24.24 −44.44 42.42 −12.12 −22.22 96.96 42.42 77.78 孔隙 4 005.72 3 508.79 −496.93 −12.41 2 817.94 −1 187.18 −29.65 4 496.59 490.87 12.25 边缘区 6 708.52 6 211.60 −496.93 −7.41 5 987.37 −721.15 −10.75 6 787.31 78.78 1.17 环道区 351.49 309.06 −42.42 −12.07 290.88 −60.60 −17.24 103.02 −248.46 −70.69 桥接区 454.51 327.25 −127.26 −28.00 230.28 −224.22 −49.33 696.91 242.40 53.33 支线 963.55 569.65 −393.91 −40.88 569.65 −393.91 −40.88 612.07 −351.49 −36.48 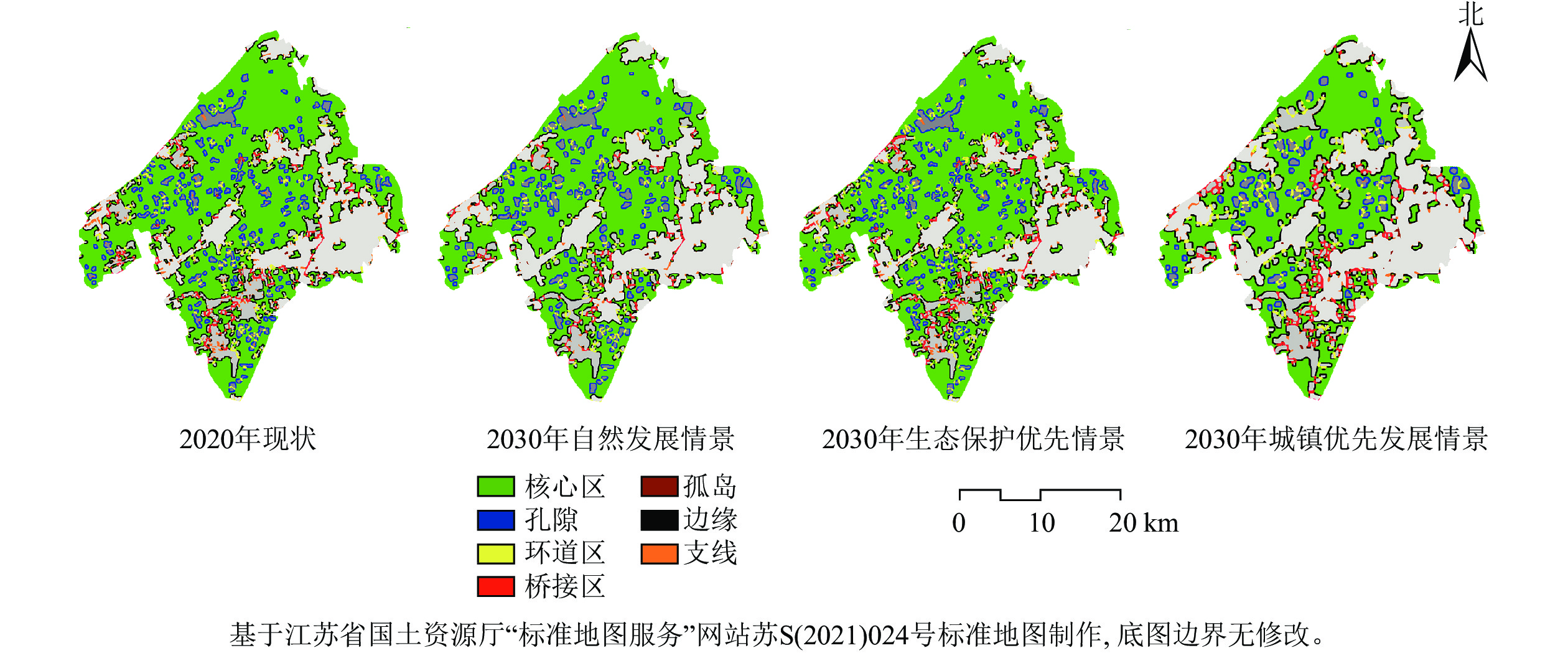
图 3 吴江南片区蓝绿空间形态学空间格局分析图
Figure 3. Analysis of morphological spatial pattern of blue and green space in southern Wujiang District
-
由图4可知:2020—2030年,①自然发展情景下,蓝绿空间整体景观连通性呈现略微下降的趋势。与2020年吴江南片区蓝绿空间景观连通性现状相比,毕家湾、沈庄漾区域景观连通性均有下降,可能连通性重要值分别下降了0.004 3、2.909 9。②生态保护优先情景下,蓝绿空间研究区核心区重要斑块数量较多、相对面积较大,且斑块较为集中,蓝绿空间连通性有所增强。相较于自然发展情景,沈庄漾和严士浜区域景观连通性增长较为显著,可能连通性重要值分别增加了2.963 5和11.699 6。③城镇发展优先情景下蓝绿空间受建设用地快速扩张的影响,连通性呈现出快速下降趋势,其中长荡—大龙荡、严士浜区域斑块所受影响较大。相对于生态保护优先情景,长荡—大龙荡区域斑块的可能连通性重要值从98.477 0下降至88.643 4,降幅约为9.98%。严士浜区域景观连通性受镇域内建设用地快速扩张的负面影响,同样呈现大幅下降趋势,可能连通性重要值由12.814 5下降至0.353 7,降幅约为97.24%。
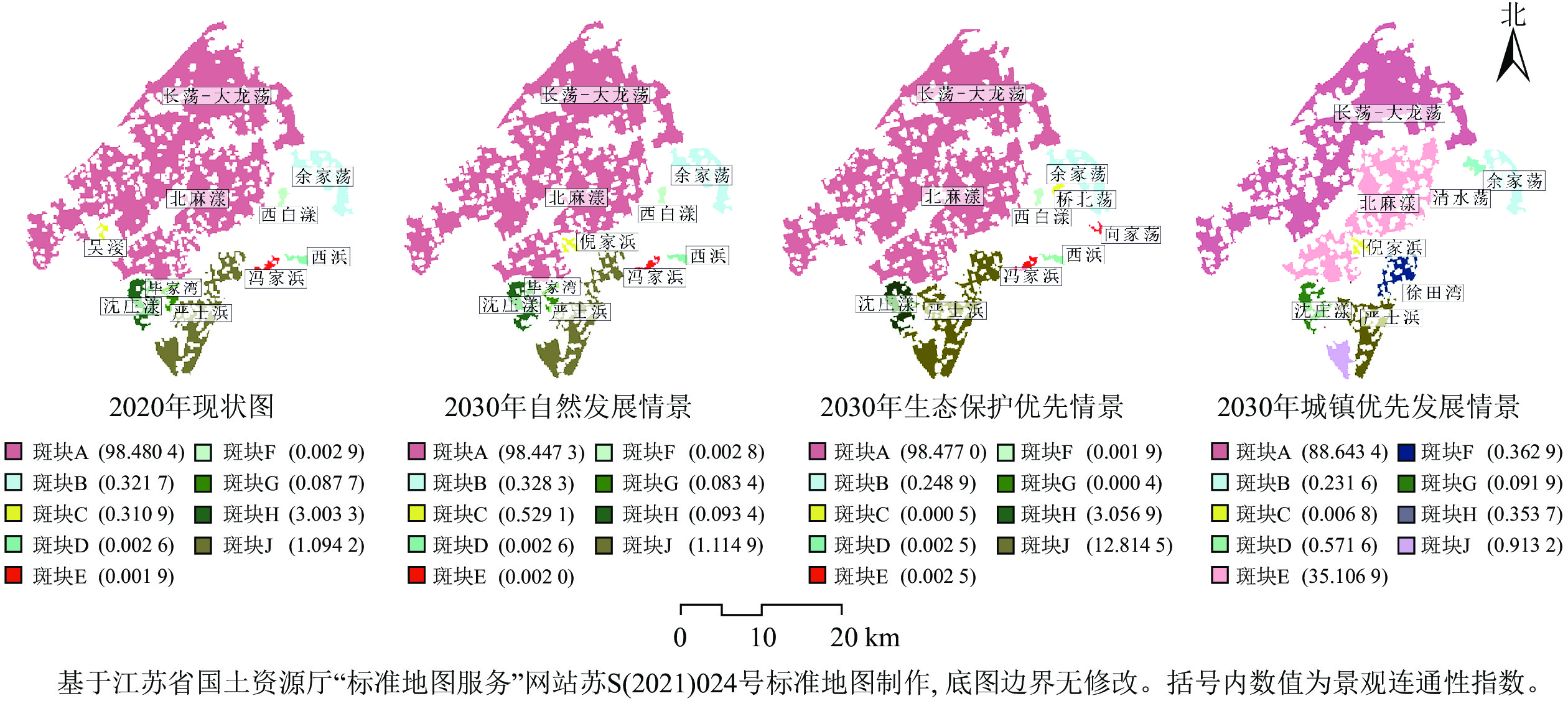
图 4 吴江南片区蓝绿空间景观连通性变化
Figure 4. Change of landscape connectivity of blue and green space in the southern Wujiang District
-
本研究基于苏州市吴江南片区2000、2010和2020年3期土地利用数据,通过参数调配设置CA-Markov模型,模拟自然发展情景、生态保护优先情景和城镇发展优先情景等3种情景下吴江南片区2030年蓝绿空间的发展趋势,并运用MSPA和景观连通性评价对比各情景模拟下的蓝绿空间格局。主要结论如下:①从多情景模拟下的蓝绿空间土地利用分析来看,自然发展情景下,耕地、林地、草地和水域面积均有不同程度减少,基本延续过去10 a吴江南片区土地利用的发展规律。在生态保护优先情景下,蓝绿空间用地面积呈现整体上升趋势,建设用地面积增速明显放缓,反映出生态优先保护情景对控制非蓝绿空间用地无序扩张具有显著成效。城镇优先发展情景下,耕地、林地、草地和水域等蓝绿空间用地面积减少速率较2020年明显加快。②从多情景模拟下的蓝绿空间形态学空间格局分析结果来看,自然发展情景下,各蓝绿空间形态类型基本延续前10 a的发展趋势,呈现负向增长,表明如不调整未来发展模式,吴江南片区仍将面临生态环境保护的压力。生态保护优先发展情景下,蓝绿空间核心区等形态类型呈现出较为明显的增长趋势,表明蓝绿空间的整体生态效益有所提升。城镇优先发展情景下,蓝绿空间核心区、孔隙等形态类型面积及占比呈现负向发展趋势,反映出吴江南片区蓝绿空间核心区内部的集聚程度受到较大影响,斑块景观连通性下降,亟须加强保护力度。③从多情景模拟下的蓝绿空间景观连通性评价来看,蓝绿空间核心区的连通性程度在自然发展情景呈现出略微下降的趋势,毕家湾、沈庄漾区域的景观连通性降低程度相对明显。生态保护优先发展情景下,蓝绿空间景观连通性整体呈现上升趋势,表明以生态保护建设为首要的发展模式对吴江南片区蓝绿空间景观破碎化程度改善具有明显作用。在城镇优先发展情景下,受人为活动和建设用地快速扩张等因素影响,长荡—大龙荡等区域的景观连通性下降幅度较为剧烈,反映出未来吴江南片区蓝绿空间连通性降低程度还将持续加剧。
-
CA-Markov模型的模拟主要基于土地利用数据分析,虽然解译多时段的土地利用数据能更细致地体现时空变化,但受区域土地利用数据遥感影像图精度限制,导致模拟结果存在一定偏差。在后续研究中应进一步提升土地利用数据的分辨率,提高研究结果的精准性。同时,在多情景设置时,本研究结合前人的研究,情景设置主要考虑了自然地理和政策法规,对社会、经济、人文等方面指标的权衡作用尚未全面考虑,未来将进一步完善多情景模拟的指标体系设置,对每个情景做更全面的定量分析比较。苏州市吴江区蓝绿空间具有典型水网生态结构特征,伴随快速城镇化进程,水网地区蓝绿空间生态呈现复杂性。面对蓝绿空间面积减少,破碎化加剧等问题,未来应该加强对蓝绿空间保护,坚守生态保护红线,管控建设用地快速扩张,保护蓝绿空间用地规模,强化蓝绿空间生态环境治理。同时,建议整合现有蓝绿空间生态资源,通过科学识别重要蓝绿空间资源,构建吴江南片区蓝绿空间生态网络等一系列重要举措,增强蓝绿空间连通性,优化城镇复合生态系统,缓解地区生态矛盾,促进人地关系和谐。
Multi-scenario simulation and prediction of blue-green spatial pattern in the southern Jiangsu water network area
-
摘要:
目的 模拟预测苏南水网地区蓝绿空间格局多情景发展趋势,探究促进地区可持续发展的情景方案。 方法 以江苏省苏州市吴江南片区为例,设置自然发展、生态保护优先和城镇发展优先等3种情景,借助CA-Markov模型,对2030年蓝绿空间格局变化趋势进行多情景模拟分析,并运用形态学空间格局分析(MSPA)和景观连通性评价解读蓝绿空间景观生态结构与生态格局特征。 结果 不同发展情景下蓝绿空间规模与格局变化差异显著。自然发展和城镇发展优先情景下,蓝绿空间面积呈现加速减少趋势,分别减少了3 626.37、7 145.74 hm2。蓝绿空间核心区、孔隙等形态类型负向变化剧烈,蓝绿空间破碎度增加,连通性降低。生态保护优先情景与自然发展和城镇发展优先情景相比,蓝绿空间面积分别增加了2 904.30、6 423.67 hm2,蓝绿空间核心区等形态类型增幅明显,孔隙率显著降低,蓝绿空间破碎度降低,连通性增强。 结论 自然发展和城镇发展优先情景对水网地区蓝绿空间呈现负向影响趋势,生态保护优先情景在维持蓝绿空间面积与格局稳定,改善蓝绿空间连通性方面效果显著。图4表4参23 -
关键词:
- 蓝绿空间 /
- CA-Markov模型 /
- 多情景模拟 /
- 形态学空间格局分析 /
- 水网地区
Abstract:Objective The objective is to simulate and predict the multi-scenario development trend of the blue-green spatial pattern in the southern Jiangsu water network area, and explore scenario plans to promote sustainable development in the region. Method Taking the southern Wujiang District of Suzhou City, Jiangsu Province as sampling area, three scenarios were set up: natural development, ecological protection priority, and urban development priority. Based on CA-Markov model, a multi-scenario simulation analysis was conducted to simulate and analyze the changing trend of the blue-green spatial pattern in 2030. Morphological spatial pattern analysis (MSPA) and landscape connectivity evaluation were used to interpret the ecological structure and characteristics of the blue-green spatial landscape. Result There were significant differences in the scale and pattern of the blue-green space under different development scenarios. Under the priority scenario of natural development and urban development, the area of the blue-green space showed an accelerated reduction trend, with a reduction of 3 626.37 and 7 145.74 hm2 respectively. The shape types of the core area and pore of the blue-green space changed sharply in the negative direction, resulting in an increase in fragmentation and a decrease in connectivity in the blue-green space. Compared with the priority scenarios of natural development and urban development, the priority scenario of ecological protection increased the area of the blue-green space by 2 904.30 and 6 423.67 hm2, and the core area of the blue-green space and other morphological types increased significantly, with a significant reduction in porosity, a reduction in the fragmentation of the blue-green space, and an increase in connectivity. Conclusion The priority scenarios of natural development and urban development have a negative impact on the blue-green space in the water network area, and the priority scenario of ecological protection has a significant effect on maintaining the stability of the area and pattern of the blue-green space and improving its connectivity. [Ch, 4 fig. 4 tab. 23 ref.] -
Key words:
- blue-green space /
- CA-Markov model /
- multi-scenario simulation /
- MSPA /
- water network area
-
核桃Juglans regia 为胡桃科Juglandaceae胡桃属Juglans植物[1],其种仁含油量高,有“木本油料之王”的称号[2]。同时,核桃木材坚实,是良好的硬木材料。作为重要的经济林树种,核桃大多种植于土壤贫瘠的山坡沟坎,不与粮争地[3]。然而,核桃树体高大,与其他果树相比对矿质营养元素需求量较高,大量元素与核桃产量和品质形成紧密相关[4],因此,提高核桃树体对矿质元素的吸收能力对于提高核桃产量和品质至关重要[5]。研究表明:在核桃树各器官中种仁的氮素质量分数最高,核桃树吸收累积的矿质营养元素中氮素被商品核桃(种仁、硬壳)携走的比例也最高,叶片次之[6]。可见,氮素可能是提高核桃产量和品质的关键营养元素。然而,过量施用氮肥会导致严重的环境问题,因此,提高氮素利用效率是提高核桃产量与品质的重中之重[7]。
土壤中氮素主要分为无机氮和有机氮两大类,植物根系能够吸收利用的主要是无机氮,主要以硝态氮和铵态氮形式存在[8−9]。植物根系对无机氮转运调节途径可分为2种,即高亲和力(HATs)和低亲和力(LATs)的氮转运系统[10],HATs 在外部铵态氮( ${\rm{NH}}_4^ +$)、硝态氮(${\rm{NO}}^ -_3 $)浓度低于 0.5 mmol·L−1时介导吸收大部分的无机氮,而 LATs 则是在 ${\rm{NH}}_4^ + $、${\rm{NO}}_3^ - $浓度高于 0.5或 1.0 mmol·L−1时介导吸收无机氮[10]。植物对铵态氮和硝态氮的吸收主要由铵转运蛋白和硝酸转运蛋白介导。土壤中同时含有植物可吸收利用的硝态氮和铵态氮时,由于植物吸收硝态氮需要先将其还原成铵态氮后才能进行同化利用,消耗的能量更多,所以植物对铵态氮表现出明显的偏好性,而且当植物受盐胁迫及活性氧的伤害时,铵态氮具有缓解作用,因此,介导植物对铵态氮吸收的铵转运蛋白在植物氮同化中起着重要作用[11]。铵转运蛋白基因主要有两大类族,分为AMT1和AMT2。在已知的铵转运蛋白中大部分 AMT1 家族的转运蛋白属于高亲和转运体[12],如拟南芥Arabidopsis thaliana中有6个编码铵转运蛋白被鉴定,包括5个AMT1家族基因,1个AMT2家族基因。AMT1家族基因中,AtAMT1.1和AtAMT1.3对拟南芥根系铵态氮吸收的贡献率最高,为30%,AtAMT1.2、AtAMT1.5对拟南芥根系高亲和铵态氮吸收的贡献率略低于AtAMT1.1和AtAMT1.3[13],AtAMT1.4在花粉中特异性表达[14],在水稻Oryza sativa铵转运蛋白基因家族中AMT1家族有基因3个,其中OsAMT1.1和OsAMT1.2为高亲和力转运体[15];而 AMT2 家族以低亲和为主,在拟南芥AMT2家族基因中AtAMT2.1 在低亲和范围内适度促进拟南芥根系对铵态氮吸收,主要在铵从根部到地上部运输中发挥作用[16]。AMT2 型蛋白通常在植物的不同组织中包括根、芽和叶中都有表达,如AtAMT2.1基因在拟南芥各器官中均有表达,主要表达在拟南芥的维管束及上皮层[17],在拟南芥中AtAMT2.1与AtAMT1s之间还存在协同作用。此外,毛果杨Populus trichocarpa PtAMT2.1主要在叶片中,PtAMT2.2在叶柄中高表达。除此之外,玉米Zea mays[18]、番茄Lycopersicon esculentum [19]、欧洲油菜Brassica napus [20]等高等作物均鉴别出了AMT基因,但到目前为止,研究大多集中于高亲和的铵转运蛋白AMT1基因家族,对低亲和的铵转运蛋白AMT2基因家族的研究较少。因此,阐明AMT2的生物学功能和调控机制,对于提高核桃自身的氮效率和提高肥料利用效率都具有重要意义。
本研究以核桃JrAMT2过表达株系为供试材料,采用实时荧光定量聚合酶链式反应(qRT-PCR)及生理检测的方法鉴定JrAMT2基因在核桃植株体内的表达模式,进一步对核桃JrAMT2基因进行生物学功能分析,为核桃优良品种选育提供理论依据。
1. 材料和方法
1.1 植物材料
实验材料来自浙江农林大学省部共建亚热带森林培育国家重点实验室保存的核桃野生型(WT)以及课题组2019年获得的核桃JrAMT2过表达阳性植株 [21],其中JrAMT2基因构建于PCMBIA1300植物表达载体,载体抗性为卡那霉素(kanmycin,Kan),利用根癌农杆菌Agrobacterium tumefaciens GV3103菌株介导将构建好的35S::JrAMT2::GFP过表达载体转化到核桃野生型体细胞胚中,植物筛选标记为潮霉素(hygromycin,Hyg)。本研究所用核桃组培苗为野生型体胚和JrAMT2过表达阳性体胚经脱水萌发获得,温室苗则由上述组培苗经生根、炼苗、驯化获得。
1.2 实验方法
1.2.1 生物信息分析运用
SOPMA(
https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html )网站在线分析JrAMT2蛋白质二级结构,运用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/ )在线软件对JrAMT2 蛋白进行跨膜结构域的预测,通过 GSDS(http://gsds.gao-lab.org/ )软件在线分析JrAMT2基因结构。1.2.2 植株的培养
同一JrAMT2过表达阳性体胚萌出的植株为1个株系,选取3个核桃JrAMT2阳性株系,命名为JrAMT2-1、JrAMT2-2、JrAMT2-3,每个株系继代培养至50株以上,培养条件:温度为 25 ℃,湿度为 75%~80%,光照强度为2~15 klx,光照周期为16 h光照8 h黑暗,培养基为Driver&Kunivuki&McGranahan (DKW)培养基。
采用2步生根法获得核桃驯化植株,选取阳性体胚萌发后经4次继代培养的核桃组培苗作为实验材料。第1步进行根诱导,5~8 cm长的光生芽,转移到补充有10 mg·L−1吲哚丁酸钾(K-IBA)的DKW固体培养基,在黑暗中培养7 d,诱导根原基的发生;第2步,不定根诱导结束后将其转移到粗蛭石∶DKW培养基比例(体积比)为3∶2的固体培养基中,温度为25 ℃,湿度为75%~80%,光照强度为2~15 klx,光照周期为16 h光照8 h黑暗,培养时间为21~28 d,形成不定根,获得核桃不同株系生根植株[22]。
核桃苗不定根形成后取出用清水冲洗,多菌灵浸泡,移栽到泥炭∶蛭石∶珍珠岩比例(体积比)为2∶1∶1的混合土中,将移栽驯化成活后获得的核桃再生植株在温度为 25 ℃,湿度为 75%~80%,光照强度为2~15 klx,光照周期为16 h光照8 h黑暗的条件下培养[23],获得核桃不同株系温室植株。
1.2.3 核桃JrAMT2基因阳性鉴定
选用阳性体胚萌发后经4次继代培养的核桃组培苗进行绿色荧光蛋白(GFP) 检测、PCR及RT-qPCR验证,引物见表1。从再生植株顶芽开始向下截取 1.5 cm,培养14 d 后观察植株表型,每个株系均5个生物学重复。选取植株顶芽、叶片、茎段混样提取DNA及RNA。 PCR 反应程序为:94 ℃预变性 2 min;98 ℃变性10 s,55 ℃退火温度 30 s,68 ℃延伸 2 min,共 32 个循环;68 ℃延伸 7 min,PCR 反应产物进行质量分数为1.2%琼脂糖凝胶电泳。使用 The iQ5 Real-Time PCR Detection System 仪器进行RT-qPCR,测定转基因植株中JrAMT2的相对表达量。反应程序为:95 ℃ 10 min;95 ℃ 10 s,60 ℃ 31 s,40 个循环;95 ℃ 15 s;60 ℃ 1 min;95 ℃ 30 s;60 ℃ 15 s。通过 2−ΔΔCt方法计算定量结果[24]。
表 1 引物Table 1 Primers引物 序列(5′→3′) 用途 Actin-F GCCGAACGGGAAATTGTC 内参 Actin-R AGAGATGGCTGGAAGAGG 内参 QJrAMT2-F AGCAAATGGGGTTCCAGGTT 定量 QJrAMT2-R TGTCTCCCGCAGATAGAAGGTA 定量 GFP-F ATGGTGAGCAAGGGCGAGGA 鉴定 GFP-R TTACTTGTACAGCTCGTCCA 鉴定 JrAMT2-F CATGAATACCACACCGGCCTA 鉴定 取核桃野生型及核桃JrAMT2过表达株系组培苗的根、茎、小叶,根纵切、茎横切临时切片分别放置于体视荧光显微镜(Carl Zeiss Stereo D13covery V12,Axio Cam MRc system)在明场和蓝光(488 nm)激发条件下利用 ZEN lite 成像软件连续拍照[25-26]。
1.2.4 生长参数、铵态氮和硝态氮测定
分别选取生长状态一致的核桃野生型与3个核桃JrAMT2过表达株系组培苗,从顶尖向下剪取1.5 cm茎段,含2~4片复叶,培养14 d,每个株系均5个生物学重复,测量株高及节间数。选取长势相同的核桃野生型及JrAMT2过表达植株采用2步生根法驯化,移栽后用直尺测量统计不同株系生长0、20、40 d的株高、节间长、叶片长度及叶片宽度。
分别选取生长状态一致的核桃野生型与3个核桃JrAMT2过表达株系组培苗,培养14 d,采用2步生根法获得核桃生根植株,植株分为地上部分及地下部分2个部分,清洗植株,分别测定地上部分及地下部分鲜质量,于105 ℃下杀青,80 ℃烘干至恒量,称取干质量。使用苏州科铭生物技术有限公司的植物铵态氮(ZATD-1-G)和植物硝态氮(ZXTD-1-G)试剂盒测定地上部分及地下部分铵态氮和硝态氮质量分数,每个株系均5个生物学重复。
1.2.5 植株叶绿体观察、叶绿素质量分数及叶绿素荧光测定
分别选取生长状态一致的核桃野生型与3个核桃JrAMT2过表达株系组培苗,取顶尖向下第2节间处复叶。将该叶片置于0.35 mol·L−1氯化钠中研磨破碎至絮状,取悬液于高倍光学显微镜下观察,找到视野中单个完整的叶肉细胞,观察细胞中的叶绿体, 使用image J软件计算叶绿体表面积与单层细胞表面积比率。每个株系均5个生物学重复。
采用丙酮浸取法测定叶绿素质量分数。分别取0.1 g核桃野生型与3个核桃JrAMT2过表达株系组培苗长势一致的叶片,剪碎,置于15 mL离心管中,加入10 mL体积分数 80%丙酮溶液,于室温黑暗处浸提,直至管内材料褪色变白,以80%丙酮溶液为对照,测定663和646 nm处吸光值,每个株系均5个生物学重复。wt=[(wa+8wb)×Vt×(mFW×1 000)−1],叶绿素 a 质量分数(wa) =20.3×D(646),叶绿素 b 质量分数(Cb)=8.04×D(663)。其中:wt为叶绿素质量分数(mg·g−1),Vt为提取液总体积(mL),mFW为叶片鲜质量(g),D(646)和D(663)分别为646和663 nm处的吸光度。
叶绿素荧光测定使用M-PEA(multi-function plant efficiency analyser)多功能植物效率分析仪(英国Hansatech公司)测定。选取生长状态一致的核桃野生型与3个核桃JrAMT2过表达株系温室苗由上向下第3片叶片暗处理30 min,在饱和脉冲光(5 000 μmol·m−2·s−1) 下进行快速叶绿素荧光诱导动力学曲线(OJIP曲线) 的测定和绘制。参照SCHANSKE等[27]的方法分析叶绿素荧光诱导动力学参数(JIP-test)。
1.2.6 数据分析
利用SPSS 26软件进行单因素方差分析(one-way ANOVA)和多重比较(邓肯法),显著性水平为0.05。使用GraphPad Prism 7.0软件绘图。
2. 结果与分析
2.1 核桃JrAMT2基因生物信息学分析
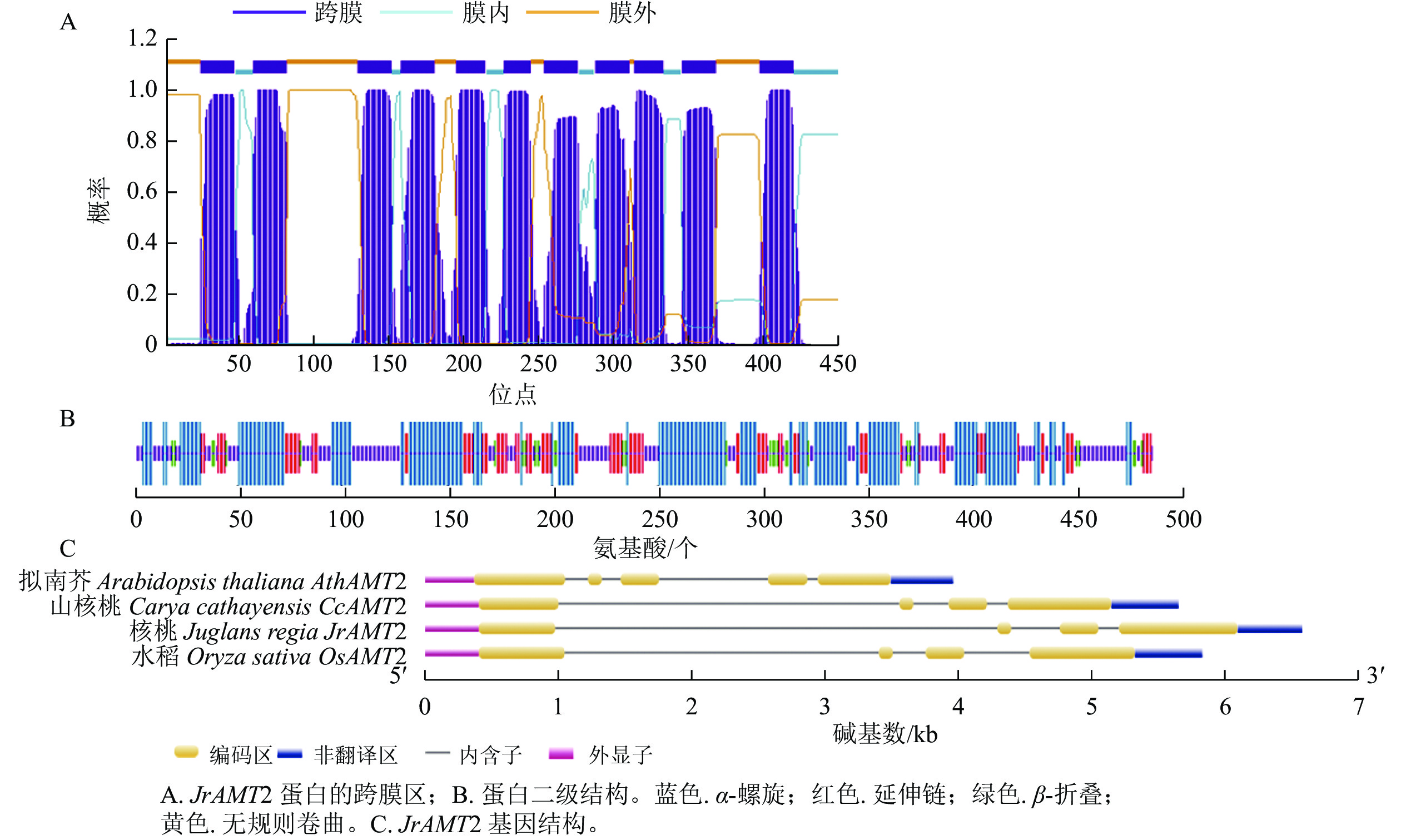
核桃 JrAMT2基因的全长为 1 464 bp,起始密码子为 ATG,终止密码子为 TGA。经过美国国家生物技术信息中心(NCBI)在线序列比对显示:该基因的序列编码的氨基酸序列属于 Ammonium Transporter Family,编码487个氨基酸(图1),蛋白分子量为52.458 kD,预测分子式为C2433H3715N601O646S23,含有7 418个原子,理论等电点为7.11,不稳定指数为35.61,脂溶指数为101.56,亲水性平均值为0.472。该段蛋白质中包含20种常见氨基酸:亮氨酸质量分数最高,达到11.7%,其次分别为甘氨酸11.1%,丙氨酸10.9%,缬氨酸8.6%,半胱氨酸质量分数最低,仅为1.0%。JrAMT2蛋白中分别含有29个酸性氨基酸残基(Asp+Glu)和29个碱性氨基酸残基(Arg+Lys) (表2)。
表 2 JrAMT2基因氨基酸组成Table 2 Composition of JrAMT2 amino acids氨基酸 数量/个 占比/% 氨基酸 数量/个 占比/% 氨基酸 数量/个 占比/% 丙氨酸 53 10.90 组氨酸 9 1.80 苏氨酸 27 5.50 精氨酸 11 2.30 异亮氨酸 25 5.10 色氨酸 18 3.70 天冬酰胺 16 3.30 亮氨酸 57 11.70 酪氨酸 17 3.50 天冬氨酸 15 3.10 赖氨酸 18 3.70 缬氨酸 42 8.60 半胱氨酸 5 1.00 甲硫氨酸 18 3.70 吡咯赖氨酸 0 0.00 谷氨酰胺 11 2.30 苯丙氨酸 24 4.90 晒半胱氨酸 0 0.00 谷氨酸 14 2.90 脯氨酸 24 4.90 甘氨酸 54 11.10 丝氨酸 29 6.00 对核桃JrAMT2蛋白跨膜区的预测结果表明:该蛋白N端在膜外,C端存在膜内,共含有11个跨膜螺旋,跨膜螺旋区段分别位于24~46、59~81、129~151、158~180、195~214、227~244、254~276、288~310、314~333、346~368和398~420,推测JrAMT2属于跨膜蛋白,并在N端存在信号肽(图2A)。
对JrAMT2 蛋白二级结构的预测结果显示:JrAMT2的氨基酸组成中有4种构象,其中 α- 螺旋有 201 个氨基酸,占比 43.12%;延伸链有 93个氨基酸,占比19.10%;β- 转角有 30个氨基酸,占比 6.16%;无规则卷曲有154个氨基酸,占比 31.62%。JrAMT2 铵转运蛋白主要由 α- 螺旋和无规则卷曲组成(图2B)。
对JrAMT2基因结构的分析显示:JrAMT2基因由4个外显子,3个内含子组成。与拟南芥AtAMT2基因相比,拟南芥基因结构多了1个外显子和1个内含子,与山核桃Carya cathayensis CcAMT2基因[28]、栓皮栎Quercus suber QsAMT2基因[29]相比,外显子和内含子数量相同,同源性较高(图2C)。
2.2 核桃JrAMT2过表达植株阳性验证
在成功构建35S::JrAMT2::GFP的过表达载体并通过农杆菌介导转化核桃体胚后,对同一无性系JrAMT2体胚经脱水萌发获得的再生植株进行阳性鉴定。利用PCR技术,以核桃JrAMT2过表达植株3个株系(JrAMT2-1、JrAMT2-2和JrAMT2-3) DNA为模板,进行外源GFP基因(729 bp)的PCR验证,检测到大小约为 750 bp的电泳条带,与GFP基因大小符合(图3A);进行目的基因JrAMT2加外源GFP基因全长(2193 bp)的PCR验证,检测到大小为2000 bp的电泳条带,与目的基因大小符合(图3B);以核桃JrAMT2过表达植株3个株系的cDNA为模板,利用实时定量PCR技术对JrAMT2基因表达量进行检测,结果显示:核桃JrAMT2过表达植株3个株系JrAMT2基因相对表达量分别为野生型的10.58、12.80和14.94倍,显著上调(图3C),表明核桃JrAMT2过表达植株中JrAMT2基因稳定表达。
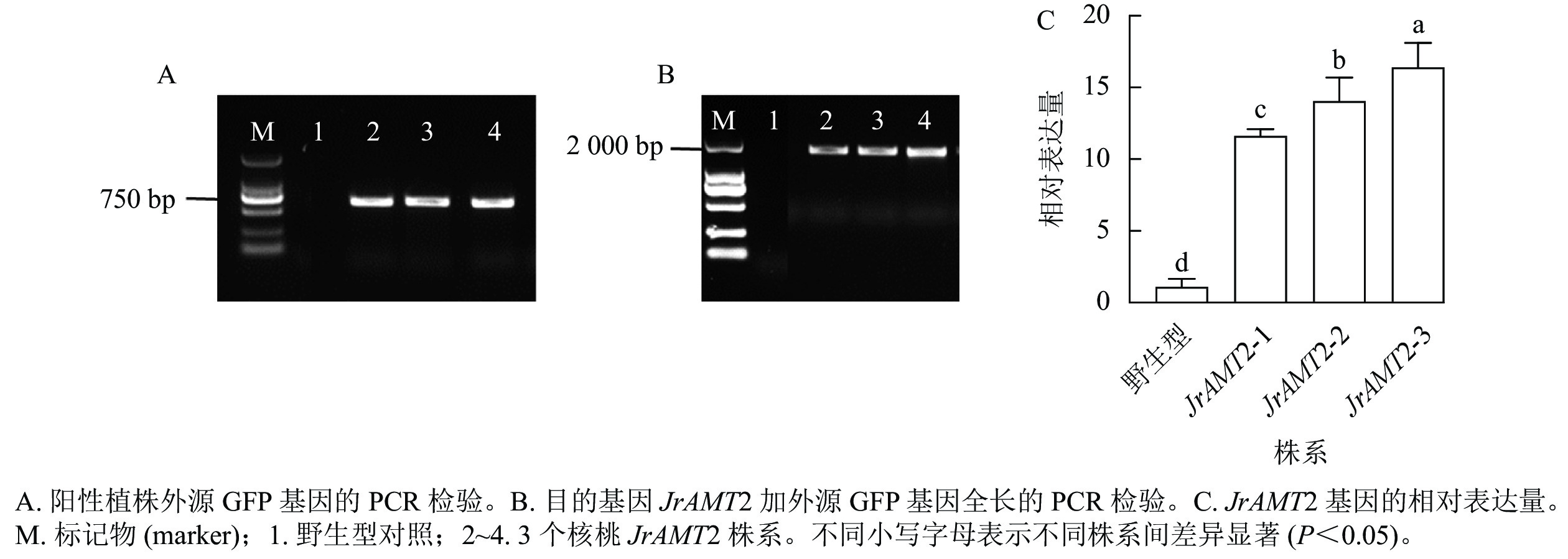 图 3 PCR 和RT-qPCR 对核桃组培苗的JrAMT2基因的检测Figure 3 Detection of JrAMT2 gene in J. regia tissue culture seedlings by PCR and qRT-PCR
图 3 PCR 和RT-qPCR 对核桃组培苗的JrAMT2基因的检测Figure 3 Detection of JrAMT2 gene in J. regia tissue culture seedlings by PCR and qRT-PCR对获得的过表达株系进行GFP荧光阳性鉴定,分别将核桃过表达及野生型幼苗(图4A)的小叶、茎、根置于荧光体视显微镜下在波长为488 nm蓝光激发下拍摄。结果显示:过表达植株的小叶、茎、根表面呈现均匀的绿色荧光,其中腋芽处荧光更明亮(图4B1~6),野生型核桃幼苗的小叶、茎、根在荧光下拍摄无绿色荧光激发(图4B7~12),说明JrAMT2蛋白在腋芽处积累。为进一步研究JrAMT2基因在核桃幼苗中的表达,对过表达株系及野生型组培苗的茎段进行横切,根进行纵切后,进行GFP荧光检测,结果显示:核桃JrAMT2过表达植株茎段横切中呈现均匀的绿色荧光,其中在维管组织中荧光更明亮,野生型植株茎段横切面无绿色荧面光(图4B1~12);核桃JrAMT2过表达植株根段纵切面中呈现均匀的绿色荧光,且在维管组织中荧光更明亮,野生型植株根段纵切面无绿色荧光(图4C1~8)。表明JrAMT2基因在核桃幼苗的根和茎中均可稳定表达,其中JrAMT2蛋白在根和茎的维管束组织中积累。
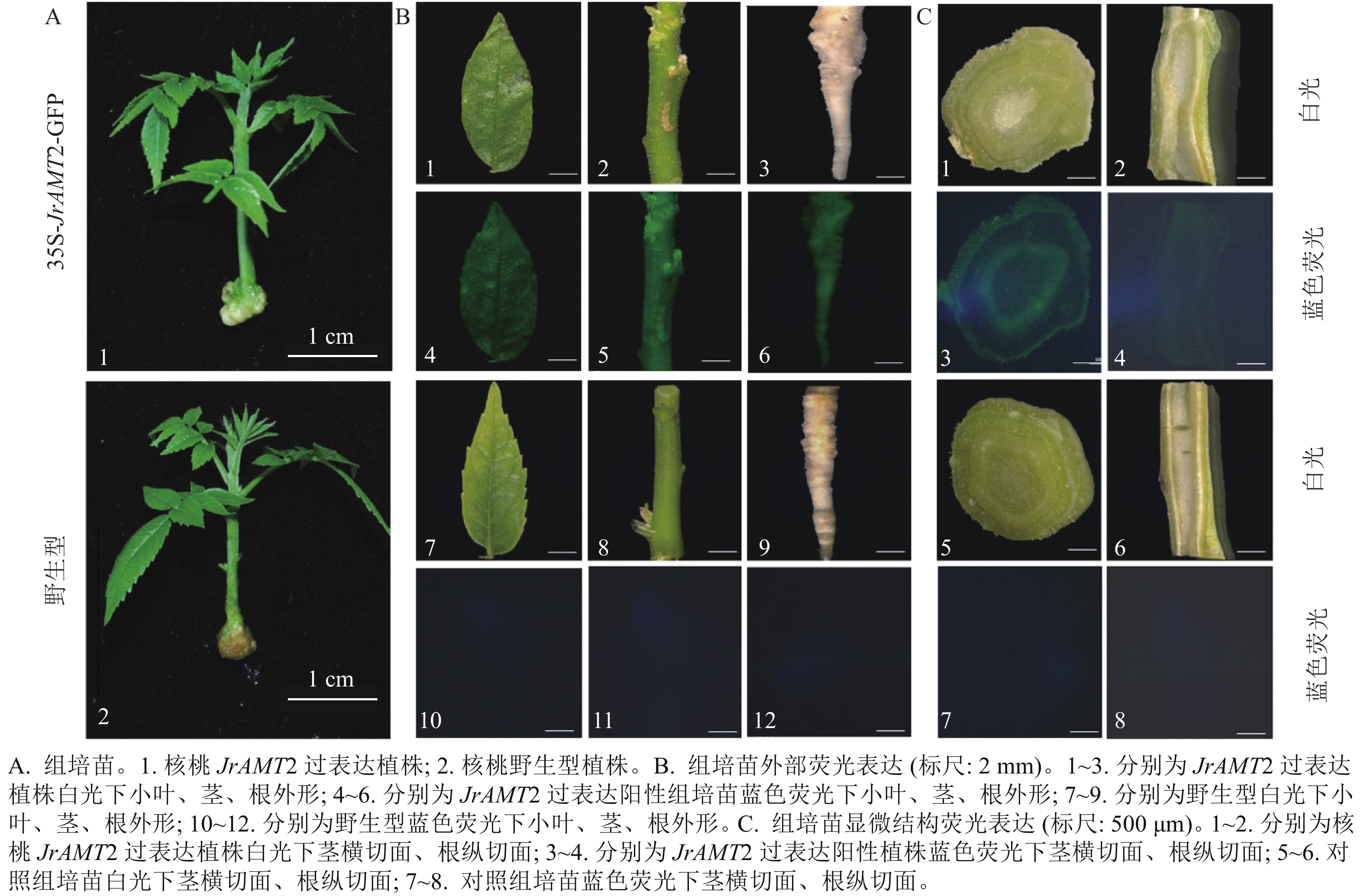 图 4 铵态氮转蛋白基因JrAMT2在核桃苗中稳定表达Figure 4 Stable expression of ammonium nitrogen transfer protein gene JrAMT2 in J. regia
图 4 铵态氮转蛋白基因JrAMT2在核桃苗中稳定表达Figure 4 Stable expression of ammonium nitrogen transfer protein gene JrAMT2 in J. regia2.3 核桃JrAMT2基因的生物学功能分析
2.3.1 核桃JrAMT2过表达植株生长表型分析
为了进一步研究核桃JrAMT2基因的生物学功能,将3个JrAMT2过表达植株与野生型核桃植株培养14 d。结果表明:与野生型相比,核桃JrAMT2过表达植株均生长旺盛,株高显著增加,节间显著增长(P<0.05,图5A)。3个JrAMT2过表达株系株高分别为3.87、4.23和4.12 cm,节间长分别为0.33、0.35和0.34 cm,野生型植株株高为3.30 cm,节间长为0.28 cm,与野生型相比,3个JrAMT2过表达株系株高分别增加了17.0%、28.0%和29.0% (图5B),节间长分别增加了19.0%、25.0%和24.0% (图5C)。综上所述,JrAMT2过表达对核桃幼苗的生长有显著提高作用。
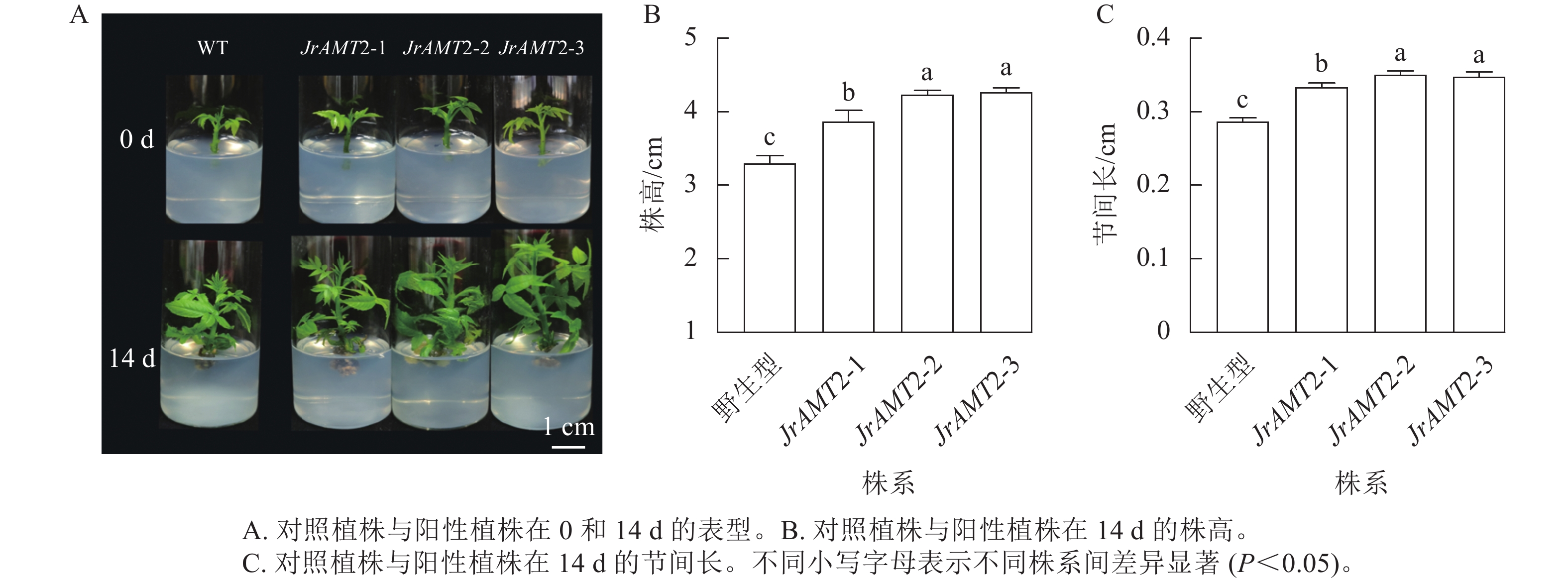 图 5 核桃JrAMT2过表达离体培养植株表型分析Figure 5 Phenotypic analysis of J. regia JrAMT2 overexpression plant in vitro culture
图 5 核桃JrAMT2过表达离体培养植株表型分析Figure 5 Phenotypic analysis of J. regia JrAMT2 overexpression plant in vitro culture对核桃JrAMT2过表达植株进一步驯化培养,成功获得核桃JrAMT2过表达温室苗。对其生长表型进行分析(图6A),定植后培养40 d,核桃JrAMT2过表达温室苗的株高、节间长、叶片长度、叶片宽度增加显著高于野生型(P<0.05)。培养至第20天时,与野生型相比,核桃JrAMT2过表达温室苗3个株系株高分别增加21.9%、26.0%和24.6% (图6B),节间长分别增加41.2%、27.7%和26.3% (图6C),叶片长度分别增加18.7%、16%和30.7% (图6D),叶片宽度分别增加41.3%、48.2%和55.1% (图6E);培养至第40天时,与野生型相比,JrAMT2过表达温室苗3个株系株高分别增加53.6%、68.2%和62.1%,节间长分别增加37.2%、50.3%和44.8%,叶片长度分别增加了27.8%、34.1%和49.3%,叶片宽度分别增加24.3%、19.5%和29.2%。综上所述,核桃JrAMT2过表达温室苗与野生型相比,株高、节间长、叶片大小均显著增加,进一步证明JrAMT2基因过表达加快了核桃的生长速度。
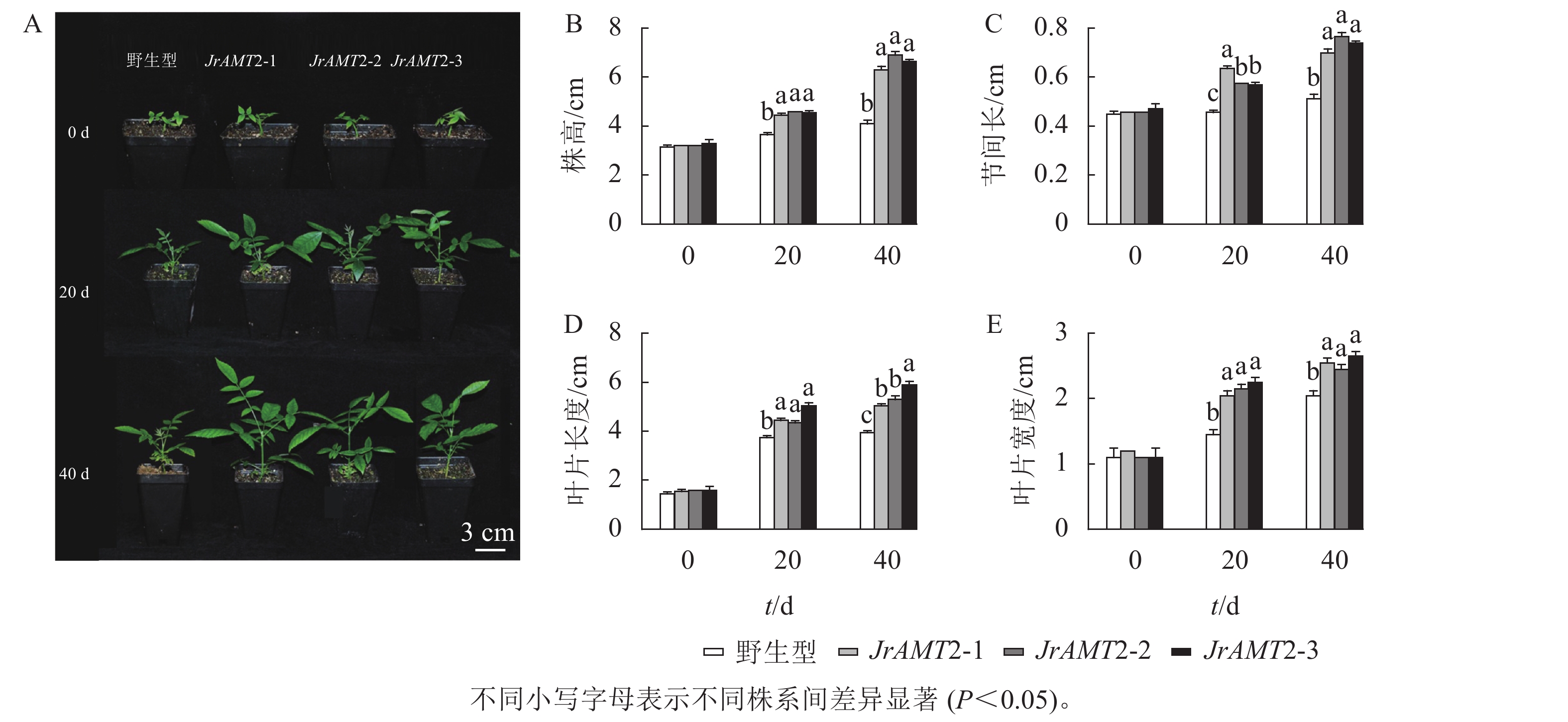 图 6 核桃JrAMT2过表达阳性温室苗性状分析Figure 6 Character analysis of J. regia JrAMT2 overexpression positive greenhouse seedlings
图 6 核桃JrAMT2过表达阳性温室苗性状分析Figure 6 Character analysis of J. regia JrAMT2 overexpression positive greenhouse seedlings2.3.2 核桃JrAMT2过表达植株对氮素吸收分析
为探究JrAMT2基因在核桃中是否存在差异表达,将核桃野生型及核桃JrAMT2过表达植株进行2步生根,获得核桃JrAMT2过表达生根植株,将其分为地上部分与地下部分。与野生型相比,核桃JrAMT2过表达生根植株地下部分不定根生长旺盛(图7A)。对核桃JrAMT2过表达植株地上部分及地下部分差异分析,分别提取3个核桃JrAMT2过表达株系生根植株地上部分及地下部分RNA,利用实时定量PCR技术,分析地上部分及地下部分JrAMT2基因表达的差异。结果表明:3个JrAMT2过表达株系地上部分JrAMT2基因表达量分别是野生型的11.94、12.70和15.06倍,地下部分JrAMT2基因表达量分别是野生型的19.07、23.34和24.00倍(图7B)。对核桃野生型及3个JrAMT2过表达株系地上部分及地下部分进行干质量和鲜质量测定。结果表明:3个JrAMT2过表达株系地上部分鲜质量和干质量显著高于野生型(P<0.05),与野生型相比,鲜质量分别增加23.45%、33.67%和56.26%,干质量分别增加23.45%、43.89%和56.26%;3个JrAMT2过表达株系地下部分干质量和鲜质量显著高于野生型(P<0.05),与野生型相比,鲜质量分别增加222.65%、199.24%和344.38%,干质量分别增加222.65%、199.24%和354.33%(图7C)。
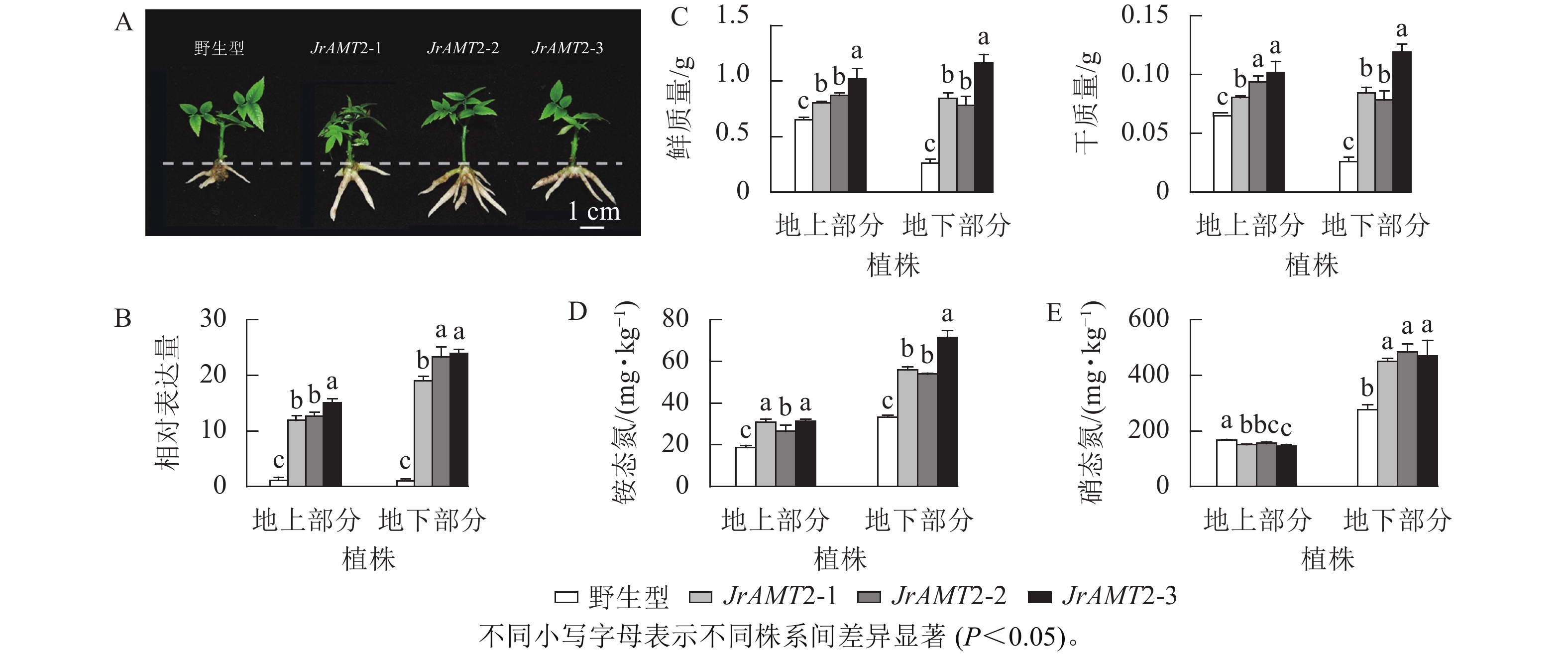 图 7 核桃JrAMT2过表达植株氮素吸收分析Figure 7 Character analysis of the regenerated plants with expression of JrAMT2 in J. regia
图 7 核桃JrAMT2过表达植株氮素吸收分析Figure 7 Character analysis of the regenerated plants with expression of JrAMT2 in J. regia基于核桃JrAMT2过表达植株表型变化,本研究进一步测定核桃JrAMT2过表达植株对硝态氮及铵态氮吸收。结果表明:与野生型相比,核桃3个JrAMT2过表达株系地上部分及地下部分铵态氮质量分数显著增加(P<0.05),地上部分分别增加59.1%、32.3%和55.1%,地下部分别增加68.1%、61.9%和114.1% (图7D);核桃3个JrAMT2过表达株系地上部分硝态氮质量分数显著降低(P<0.05),地下部分硝态氮质量分数显著增加(P<0.05),地上部分分别降低10.1%、7.1%和13.6%,地下部分别增加63.5%、75.7%和70.3% (图7E)。综上所述,JrAMT2基因主要作用于核桃地下部分,且JrAMT2基因过表达促进植株地下部分对铵态氮和硝态氮的吸收,并介导了铵态氮从地下部到地上部的运输。
2.3.3 核桃JrAMT2过表达植株叶绿素质量分数及叶绿素荧光特性分析
对核桃JrAMT2过表达植株叶色与叶片细胞叶绿体进行观察。核桃JrAMT2过表达植株叶色与野生型相比颜色较深(图8A),在显微镜下观察3个核桃JrAMT2过表达植株株系及野生型叶绿体特征发现,3个核桃JrAMT2过表达株系扩张的栅栏叶肉细胞中的叶绿体更致密(图8B)。进一步分析发现:对3个阳性株系进行总叶绿素质量分数测定,3个核桃JrAMT2过表达株系叶绿素质量分数显著高于野生型(P<0.05),野生型总叶绿素质量分数为3.53 mg·g−1,核桃JrAMT2过表达阳性植株3个株系总叶绿素质量分数分别为4.64、4.69和6.10 mg·g−1,与野生型相比分别增加了32.6%、34.0%和74.3% (图8C);JrAMT2过表达株系叶绿体表面积在单层细胞表面积的占比显著高于野生型(P<0.05),野生型叶绿体表面积与单层细胞表面积的比率为0.56,3个核桃JrAMT2过表达株系叶绿体表面积与单层细胞表面积的比值分别为0.67、0.68和0.69,与野生型相比分别增加了19.41%、21.18%和22.94% (图8D)。综上所述,JrAMT2基因过表达提高了叶绿体表面积与单层细胞表面积的比率及核桃叶肉细胞内叶绿素的积累。
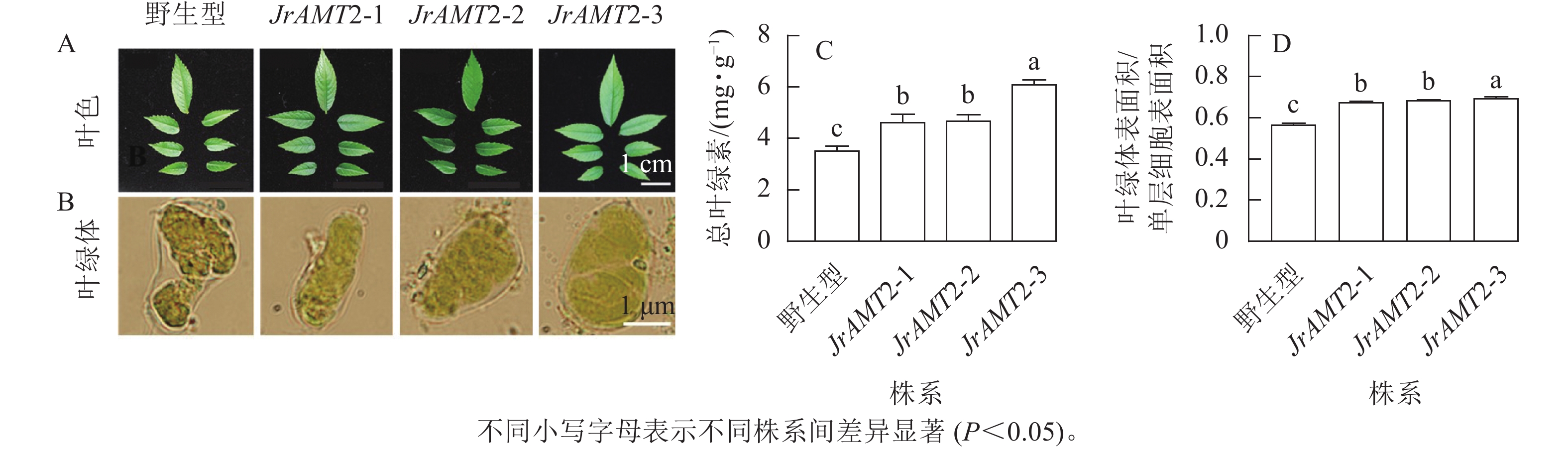 图 8 核桃JrAMT2过表达植株叶绿体及叶绿素质量分数分析Figure 8 Analysis of chloroplast and chlorophyll content in J. regia with JrAMT2 overexpression
图 8 核桃JrAMT2过表达植株叶绿体及叶绿素质量分数分析Figure 8 Analysis of chloroplast and chlorophyll content in J. regia with JrAMT2 overexpression对3个核桃JrAMT2过表达株系进行快速叶绿素荧光诱导动力学曲线测定。结果显示:与野生型相比,3个核桃JrAMT2过表达株系的O (Fo点)相、K相降低,J点、I点、P (Fm)及J~P点振幅均提高,OJIP曲线较陡,表明JrAMT2基因一定程度上提高了叶片的活性(图9A)。与野生型相比,3个核桃JrAMT2过表达株系暗适应后的最大荧光强度(Fm)、最大量子产额(Fv/Fm)均提高,Fm分别提高了47.1%、45.9%、50.2%,Fv/Fm分别提高了15.6%、13.4%、14.7%,表明JrAMT2基因一定程度上提高了光系统Ⅱ(PSⅡ)的光化学效率(图9B)。计算归一化处理的OJIP曲线(Vt)[Vt =(Ft−Fo /(Fm−Fo),Ft为任意时刻t的荧光值],并计算相对荧光差异ΔVt[ΔVt = Vt处理−Vt对照,用ΔK、ΔJ值分别显示在300 μs和20 ms处的ΔVt值,表示放氧复活体的活性(OEC)]。结果显示:与野生型相比,3个核桃JrAMT2过表达株系ΔK、ΔJ值下降且<0 (图9C),表明JrAMT2基因一定程度上提升了核桃叶片PSⅡ供体侧及受体侧电子传递效率及放氧复活体的活性。
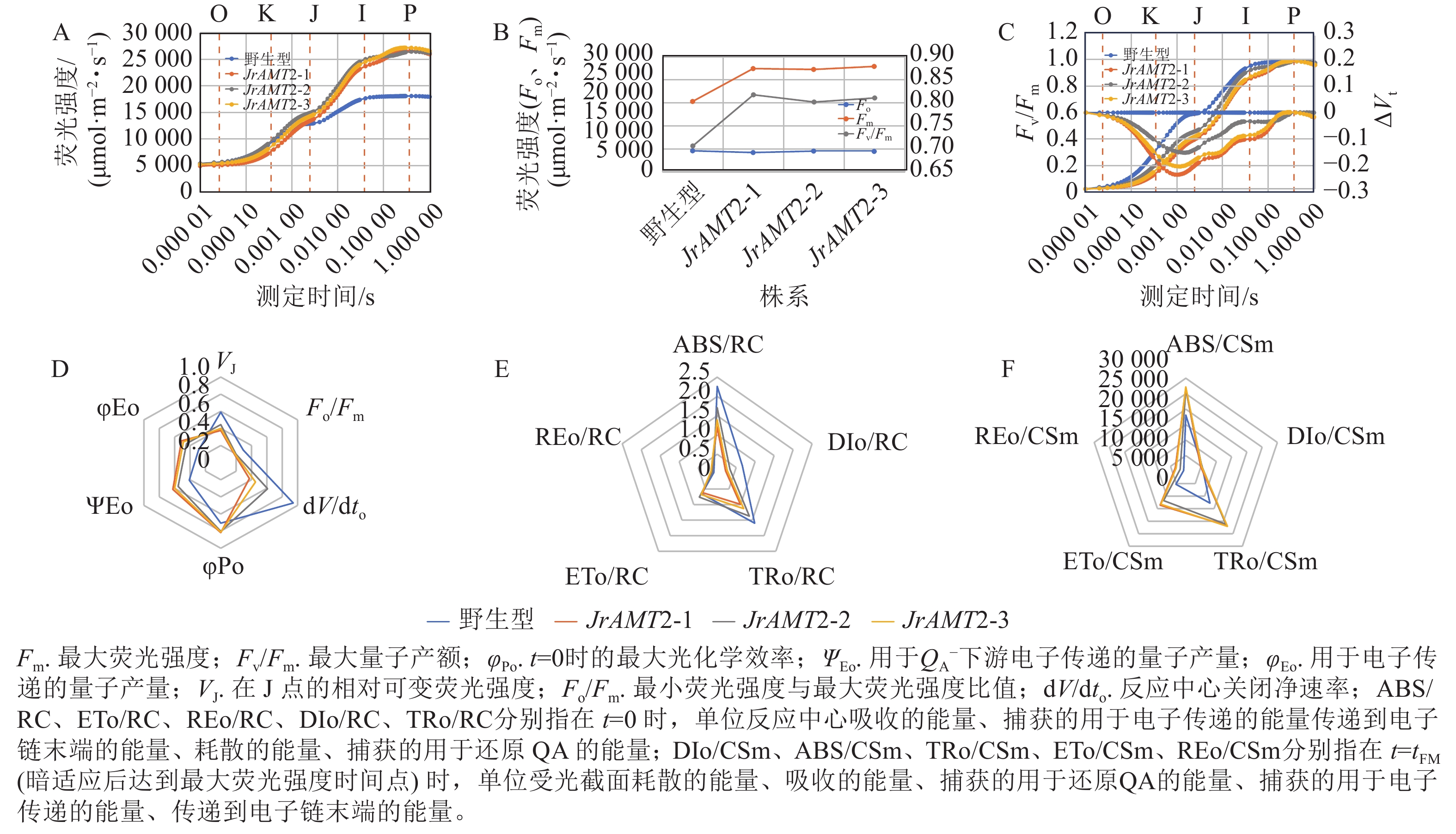 图 9 铵态氮转运蛋白JrAMT2基因表达对核桃叶绿素荧光的影响Figure 9 Effect of ammonium nitrogen transporter JrAMT2 gene on chlorophyll fluorescence of J. regia
图 9 铵态氮转运蛋白JrAMT2基因表达对核桃叶绿素荧光的影响Figure 9 Effect of ammonium nitrogen transporter JrAMT2 gene on chlorophyll fluorescence of J. regia用JIP-test参数对OJIP曲线进行定量分析,结果显示:核桃3个JrAMT2过表达株系在t=0时的最大光化学效率(φPo)、反应中心捕获的光能用于${\rm{Q}}_{\rm{A}}^- $下游电子传递的量子产量(ΨEo)、吸收的能量用于电子传递的量子产量(φEo)上升,分别上升了14.62%、44.59%、65.80%;在J点的相对可变荧光强度(VJ)、最小荧光强度与最大荧光强度比值(Fo/Fm)、反应中心关闭净速率(dV/dto)下降,分别下降了31.03%、35.29%、49.34% (图9D),表明JrAMT2基因一定程度上提高了PS反应中心的量子比率及产额;对单位PS反应中心比活性参数分析,结果显示:与野生型相比,3个核桃JrAMT2过表达株系在t=0 时的单位反应中心捕获的用于电子传递的能量(ETo/RC)、在t=0 时的单位反应中心传递到电子链末端的能量(REo/RC)无明显变化,在t=0 时单位反应中心吸收的能量(ABS/RC)、在t=0 时单位反应中心耗散的能量(DIo/RC)、在 t=0 时单位反应中心捕获的用于还原QA的能量(TRo/RC)显著下降,分别为36.43%、58.67%、27.22%,表明JrAMT2基因一定程度提高了核桃叶片反应中心活性和用于电子传递的能量份额,增强了电子传递能力(图9E);对单位受光截面比活性参数分析,结果显示:与野生型相比,3个核桃JrAMT2过表达株系在 t=tFM(暗适应后达到最大荧光强度时间点)时的单位受光截面耗散的能量(DIo/CSm)无明显变化,在 t=tFM时单位受光截面吸收的能量(ABS/CSm)、在t=tFM时单位受光截面捕获的用于还原QA的能量(TRo/CSm)、在 t=tFM时单位受光截面捕获的用于电子传递的能量(ETo/CSm)、在t=tFM时单位受光截面传递到电子链末端的能量(REo/CSm)上升,分别上升了 47.79%、69.39%、145.10%、303.62%(图9F),表明JrAMT2基因一定程度提高核桃叶片单位受光截面的电子传递的份额及电子传递效率。综上所述,JrAMT2基因过表达促进核桃的光合作用。
3. 讨论
氮素作为植物生长发育必不可少的营养元素,是核酸、蛋白质、酶、叶绿素、植物激素等的重要组成部分[30]。自然界中可供利用的氮素资源有限,作物高产就需要化肥的投入。中国的化肥投入总量逐渐升高,但化肥利用率很低,给自然环境带来极大的负担 [31−32],因此合理使用化肥,提高作物对氮素的吸收效率是促进农业生态化发展的重要途径。铵转运蛋白基因AMTs是广泛存在于动物、植物、微生物中用于运输${\rm{NH}}_4^+ $的载体蛋白,从分子层面提高植物对氮素的利用具有重要意义。前人研究发现:在拟南芥中,氮饥饿能诱导AtAMT1.1、AtAMT2.1基因上调表达,并且AtAMT2基因的表达水平随着氮饥饿时间的延长而增加[33-34]。在充足或高氮条件下,观察到拟南芥中AtAMT2.1及水稻根中OsAMT1.2基因表达水平仍上调 [35-36],其中氮素形态对AMT基因转录水平的调控也取决于AMT基因个体和植物物种。
本研究对核桃JrAMT2过表达植株进行初步的功能验证,对JrAMT2基因进行生物信息学分析表明:JrAMT2蛋白中含有29个酸性氨基酸残基(Asp+Glu)和29个碱性氨基酸残基(Arg+Lys)。该蛋白N端在膜外,C端存在膜内,共含有11个跨膜螺旋结构域,与李畅[36]预测的OsAMT2.1蛋白结构域特点相同,且与夏金泽等[37]研究的木薯Manihot esculenta MeAMT2.6基因的蛋白结构相似。AMT2 型蛋白通常在植物的各种组织中表达,包括根、芽和叶。前人研究发现:杜梨 Pyrus betulifolia PbAMT2基因在所有器官中均有表达,但在根部表达最高[38]。在拟南芥中发现:AtAMT2.1基因主要在维管组织中表达,在芽中的表达高于根[15]。本研究对核桃JrAMT2过表达植株进行绿色荧光观察发现:JrAMT2蛋白在核桃苗整株均有表达,且在芽及根茎的维管束中荧光更明亮,说明与拟南芥相同,核桃JrAMT2蛋白在所有器官中表达,且主要在维管组织中表达。
在植物生长发育重要阶段充足的氮素营养供给可以促进其生长发育,增加产量,对植物外施氮素能增加植株株高及叶面积[39]。本研究对核桃JrAMT2过表达植株生长性状分析发现:JrAMT2基因在核桃中过表达对植株生长发育有调控作用,主要表现在过表达植株株高、节间长显著增加。对过表达植株生根驯化结果显示:过表达植株生物量显著增加,根系发达。对核桃JrAMT2过表达植株移栽驯化,定植后植株生长速率显著高于野生型,主要表现在节间伸长快,叶面积增加。AMT2是具有铵吸收功能的铵转运蛋白,且在维管组织表达,暗示着该基因可能参与铵向木质部的装载,介导铵在植物中的长距离运输。研究发现:拟南芥AtAMT2.1除了对根吸收氮素有一定贡献外,主要作用于${\rm{NH}}_4^+ $从根部到茎部的运输[40]。本研究对核桃JrAMT2过表达植株地上部分和地下部分基因表达测定发现:植株地下部分JrAMT2基因表达量显著高于地上部分,且植株地下部分生物量的增加显著高于地下部分,说明核桃JrAMT2基因主要在地下部分表达。对核桃JrAMT2过表达植株对铵态氮和硝态氮吸收测定结果表明:核桃JrAMT2过表达植株地上部分仅对铵态氮的吸收显著上调,地下部分对铵态氮与硝态氮的吸收均显著上调,说明JrAMT2基因促进植株对铵态氮和硝态氮的吸收,并介导了铵态氮从地下部分到地上部分的运输。氮素营养还会通过影响叶绿素合成和叶绿素荧光参数的变化来参与光能的利用和调控[41]。有研究表明:水稻OsAMT2.1基因敲除株系的光合特性与野生型相比出现下降趋势,同样说明了AMT2基因对植物光合作用有调控作用[36]。本研究对核桃JrAMT2过表达植株叶绿素质量分数及叶绿素荧光参数变化的测定结果显示:JrAMT2基因显著提高了核桃的叶绿体表面积与单层细胞表面积比率、叶绿素质量分数及叶绿素荧光参数中叶片放氧复活体活性、量子产额、电子传递效率。
4. 结论
JrAMT2作为铵态氮转运基因,促进核桃对铵态氮和硝态氮的吸收,且介导铵态氮从根部到茎部的运输,对核桃生长发育、光合作用等有积极作用,对研究核桃高效利用氮素及良种的筛选有重要意义。
-
图 1 2020年吴江南片区土地利用现状与模拟
Figure 1 Landuse status and simulation of southern Wujiang District in 2020
图 2 吴江南片区蓝绿空间土地利用变化
Figure 2 Landuse change of blue and green space in southern Wujiang District
图 3 吴江南片区蓝绿空间形态学空间格局分析图
Figure 3 Analysis of morphological spatial pattern of blue and green space in southern Wujiang District
图 4 吴江南片区蓝绿空间景观连通性变化
Figure 4 Change of landscape connectivity of blue and green space in the southern Wujiang District
表 1 2000—2010年吴江南片区土地利用类型转移概率矩阵
Table 1. Probability matrix of land use type transfer in southern Wujiang District from 2000 to 2010
2000年土地利用类型 2010年土地类型转移概率/% 耕地 林地 草地 水域 建设用地 耕地 63.74 0.03 0.02 4.58 31.63 林地 6.02 68.85 0.19 14.30 10.64 草地 0.01 0.73 72.42 26.83 0.01 水域 13.93 0.05 0.01 76.64 9.37 建设用地 19.95 0.06 0.01 4.21 75.77  下载: 导出CSV
下载: 导出CSV
表 2 2010—2020年吴江南片区土地利用类型转移概率矩阵
Table 2. Probability matrix of land use type transfer in southern Wujiang District from 2010 to 2020
2010年土地利用类型 2020年土地利用类型转移概率/% 耕地 林地 草地 水域 建设用地 耕地 65.13 0.01 0.01 3.44 31.41 林地 0.90 75.78 0.13 13.45 9.74 草地 15.42 0.01 84.55 0.01 0.01 水域 13.37 0.01 0.01 75.57 11.04 建设用地 14.27 0.01 0.01 5.13 80.58
下载: 导出CSV
表 3 2020—2030年吴江南片区蓝绿空间土地利用数据
Table 3. Land use data of blue and green space in southern Wujiang District from 2020 to 2030
类型 2020年现状 2030年自然发展情景 2030年生态保护优先情景 2030年城镇发展优先情景 面积/hm2 占比/% 面积/hm2 占比/% 面积/hm2 占比/% 面积/hm2 占比/% 耕地 33 837.03 55.84 31 826.79 52.52 33 058.35 54.36 28 946.79 47.84 林地 142.29 0.23 129.78 0.21 148.59 0.24 30.41 0.05 草地 19.44 0.03 17.64 0.03 22.14 0.04 17.64 0.03 水域 9 838.98 16.24 8 237.16 13.59 9 886.59 16.26 7 697.16 12.72 建设用地 16 763.22 27.66 20 390.76 33.65 17 693.10 29.10 23 810.58 39.36
下载: 导出CSV
表 4 2020—2030年吴江南片区蓝绿空间MSPA景观类型变化
Table 4. Changes of MSPA landscape types in the blue and green space of southern Wujiang District from 2020 to 2030
类别 2020年实际
面积/hm2自然发展情景 生态保护优先情景 城镇优先发展情景 2030年预测
面积/hm2面积变化/
hm2变化率/
%2030年预测
面积/hm2面积变化/
hm2变化率/
%2030年预测
面积/hm2面积变化/
hm2变化率/
%核心区 31 312.51 29 258.13 −2 054.37 −6.56 31 803.37 490.87 1.57 23 804.05 −7 508.46 −23.98 孤岛 54.54 30.30 −24.24 −44.44 42.42 −12.12 −22.22 96.96 42.42 77.78 孔隙 4 005.72 3 508.79 −496.93 −12.41 2 817.94 −1 187.18 −29.65 4 496.59 490.87 12.25 边缘区 6 708.52 6 211.60 −496.93 −7.41 5 987.37 −721.15 −10.75 6 787.31 78.78 1.17 环道区 351.49 309.06 −42.42 −12.07 290.88 −60.60 −17.24 103.02 −248.46 −70.69 桥接区 454.51 327.25 −127.26 −28.00 230.28 −224.22 −49.33 696.91 242.40 53.33 支线 963.55 569.65 −393.91 −40.88 569.65 −393.91 −40.88 612.07 −351.49 −36.48
下载: 导出CSV
-
[1] 杨朝斌, 张亭, 胡长涛, 等. 蓝绿空间冷岛效应时空变化及其影响因素——以苏州市为例[J]. 长江流域资源与环境, 2021, 30(3): 677 − 688. YANG Chaobin, ZHANG Ting, HU Changtao, et al. Spatial-temporal characteristics of the cooling island for blue-green space and its driving factors in Suzhou, China [J]. Resources and Environment in the Yangtze Basin, 2021, 30(3): 677 − 688. [2] 张月朋, 常青, 郭旭东. 面向实践的生态用地内涵、多维度分类体系[J]. 生态学报, 2016, 36(12): 3655 − 3665. ZHANG Yuepeng, CHANG Qing, GUO Xudong. Management-oriented ecological land's conception and multi-dimensional classification system in China [J]. Acta Ecologica Sinica, 2016, 36(12): 3655 − 3665. [3] 支林蛟, 王锦, 刘敏, 等. 滇中城市群绿色空间格局动态变化多尺度研究[J]. 西南林业大学学报(自然科学), 2021, 41(5): 88 − 97. ZHI Linjiao, WANG Jin, LIU Min, et al. Multiscale dynamic change of green spatial pattern of urban agglomeration in central Yunnan [J]. Journal of Southwest Forestry University, 2021, 41(5): 88 − 97. [4] 崔洁, 许浩, 刘伟. 近20年徐州都市圈蓝绿空间景观格局演变分析[J]. 园林, 2022, 39(7): 75 − 81. CUI Jie, XU Hao, LIU Wei. Analysis on the evolution of blue-green spatial landscape pattern in Xuzhou metropolitan area in the past 20 years [J]. Landscape Architecture Academic Journal, 2022, 39(7): 75 − 81. [5] 许浩, 金婷, 刘伟. 苏锡常都市圈蓝绿空间规模与格局演变特征[J]. 南京林业大学学报(自然科学版), 2022, 46(1): 219 − 226. XU Hao, JIN Ting, LIU Wei. Study on the scale and landscape pattern evolution characteristics of blue-green space in Suzhou-Wuxi-Changzhou metropolitan area, China [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2022, 46(1): 219 − 226. [6] IOJĂ I, OSACI-COSTACHE G, BREUSTE J, et al. Integrating urban blue and green areas based on historical evidence [J]. Urban Forestry &Urban Greening, 2018, 34: 217 − 225. [7] 曹靖. 全域一张蓝图导向的城乡蓝绿空间营建策略——以安徽省界首市为例[J]. 规划师, 2021, 37(9): 26 − 32. CAO Jing. Construction strategy of urban and rural blue-green space on one blueprint: a case study of Jieshou in Anhui Province [J]. Planners, 2021, 37(9): 26 − 32. [8] 吴岩, 贺旭生, 杨玲. 国土空间规划体系背景下市县级蓝绿空间系统专项规划的编制构想[J]. 风景园林, 2020, 27(1): 30 − 34. WU Yan, HE Xusheng, YANG Ling. Compilation conception of city and county level blue-green space system specialized planning under national territory spatial planning system [J]. Landscape Architecture, 2020, 27(1): 30 − 34. [9] CRUZ J A, BLANCO A C, GARCIA J J, et al. Evaluation of the cooling effect of green and blue spaces on urban microclimate through numerical simulation: a case study of Iloilo River Esplanade, Philippines [J]. Sustainable Cities and Society, 2021, 74: 103 − 184. [10] FINLAY J, FRANKE T, MCKAY H, et al. Therapeutic landscapes and wellbeing in later life: impacts of blue and green spaces for older adults [J]. Health &Place, 2015, 34: 97 − 106. [11] TRIGUERO-MAS M, DONAIRE-GONZALEZ D, SETO E, et al. Natural outdoor environments and mental health: stress as a possible mechanism [J]. Environmental Research, 2017, 159: 629 − 638. [12] WATERS D P. von Neumann’s theory of self-reproducing automata: a useful framework for biosemiotics? [J]. Biosemiotics, 2012, 5: 5 − 15. [13] GUAN Dongjie, LI Haifeng, INOHAE T, et al. Modeling urban land use change by the integration of cellular automaton and Markov model [J]. Ecological Modelling, 2011, 222: 3761 − 3772. [14] 高星, 杨刘婉青, 李晨曦, 等. 模拟多情景下白洋淀流域土地利用变化及生态系统服务价值的空间响应[J]. 生态学报, 2021, 41(20): 7974 − 7988. GAO Xing, YANG Liuwanqing, LI Chenxi, et al. Land use change and ecosystem service value measurement in Baiyangdian Basin under the simulated multiple scenarios [J]. Acta Ecologica Sinica, 2021, 41(20): 7974 − 7988. [15] 林鸿煜, 钱晶, 严力蛟, 等. 基于形态学空间格局分析与 CA-Markov 模型的武义县绿色基础设施时空格局变化及情景模拟[J]. 浙江农业学报, 2019, 31(7): 1193 − 1204. LIN Hongyu, QIAN Jing, YAN Lijiao, et al. Analysis of spatial-temporal pattern and scenario simulation of green infrastructure in Wuyi County based on morphological spatial pattern analysis and CA-Markov model [J]. Acta Agriculturae Zhejiangensis, 2019, 31(7): 1193 − 1204. [16] 汪辉, 刘媛, 时艳, 等. 基于 PLUS 模型的溱湖国家湿地公园情景模拟与预测[J]. 浙江农林大学学报, 2023, 40(6): 1311 − 1321. WANG Hui, LIU Yuan, SHI Yan, et al. Scenario simulation and prediction of Qinhu National Wetland Park based on PLUS Model [J]. Journal of Zhejiang A&F University, 2023, 40(6): 1311 − 1321. [17] 杨清可, 段学军, 金志丰, 等. 南通市建设用地扩张情景模拟与景观生态效应[J]. 地理科学, 2017, 37(4): 528 − 536. YANG Qingke, DUAN Xuejun, JIN Zhifeng, et al. Spatial scenario simulation and landscape ecological effect based on construction land expansion of Nantong City [J]. Scientia Georgaphica Sinica, 2017, 37(4): 528 − 536. [18] 李空明, 李春林, 曹建军, 等. 基于景观生态学的辽宁中部城市群绿色基础设施20年时空格局演变[J]. 生态学报, 2021, 41(21): 8408 − 8420. LI Kongming, LI Chunlin, CAO Jianjun, et al. Spatiotemporal evolution characteristics of urban green infrastructure in central Liaoning urban agglomeration during the past 20 years based on landscape ecology and morphology [J]. Acta Ecologica Sinica, 2021, 41(21): 8408 − 8420. [19] 陈利顶, 傅伯杰. 景观连接度的生态学意义及其应用[J]. 生态学杂志, 1996, 15(4): 37 − 42, 73. CHEN Liding, FU Bojie. The ecological significance and application of landscape connectivity [J]. Chinese Journal of Ecology, 1996, 15(4): 37 − 42, 73. [20] 谢婧, 李文, 贾佳, 等. 齐齐哈尔市主城区生态网络构建与优化[J]. 水土保持研究, 2021, 28(6): 308 − 315. XIE Jing, LI Wen, JIA Jia, et al. Construction and optimization of ecological network in the main urban area of Qiqihar City [J]. Research of Soil and Water Conservation, 2021, 28(6): 308 − 315. [21] 陈德超, 施祝凯, 王祖静, 等. 苏州环太湖地区生态网络构建与空间冲突识别[J]. 生态与农村环境学报, 2020, 36(6): 778 − 787. CHEN Dechao, SHI Zhukai, WANG Zujing, et al. Ecological network construction and spatial conflict identification around Taihu Lake Area in Suzhou City [J]. Journal of Ecology and Rural Environment, 2020, 36(6): 778 − 787. [22] 李国煜, 林丽群, 伍世代, 等. 生态源地识别与生态安全格局构建研究——以福建省福清市为例[J]. 地域研究与开发, 2018, 37(3): 120 − 125. LI Guoyu, LIN Liqun, WU Shidai, et al. Recognition of ecological source and ecological security pattern construction: a case study of Fuqing City [J]. Areal Research and Development, 2018, 37(3): 120 − 125. [23] 蒙吉军, 王雅, 王晓东, 等. 基于最小累积阻力模型的贵阳市景观生态安全格局构建[J]. 长江流域资源与环境, 2016, 25(7): 1052 − 1061. MENG Jijun, WANG Ya, WANG Xiaodong, et al. Construction of landscape ecological security pattern in Guiyang based on MCR model [J]. Resources and Environment in the Yangtza Basin, 2016, 25(7): 1052 − 1061. 期刊类型引用(0)
其他类型引用(13)
-

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220741
 点击查看大图
点击查看大图
计量
- 文章访问数: 419
- HTML全文浏览量: 181
- PDF下载量: 17
- 被引次数: 13
