




 下载:
下载:


-
绿竹Bambusa oldhami是禾本科Gramineae竹亚科Bambusoideae绿竹属Bambusa的优质笋用竹植物,分布于华南丛生竹林亚区和华中亚热带混生竹林亚区南部,5−10月为产笋期[1-3]。绿竹笋形似马蹄,俗称“马蹄笋”,笋肉细嫩,味道鲜美,深受人们喜爱。由于出笋时期温度较高,加之出笋集中,绿竹笋采后容易发生变质、腐败、木质化,储藏难度较大,限制了产品的流通和销售[4-5]。草酸广泛存在于动植物和真菌体内,具有很强的抗氧化性[6]。适当浓度的草酸溶液可以作为抗氧化剂应用于果蔬的保鲜、储藏与运输等。有研究发现:草酸处理能够有效延缓鲜切香蕉Musa nana[7]、荔枝Litchi chinensis[8]、去皮荸荠Heleocharis dulcis[9]的变质腐败,延长保鲜期。关于外源草酸处理对于采后去壳绿竹笋的抗氧化和木质化的影响尚未被报道。因此,本研究以绿竹笋为对象,研究外源草酸对去壳绿竹笋抗氧化酶和木质化的影响,为竹笋的采后储藏保鲜提供依据。
-
供试材料绿竹笋于2017年7月下旬采自浙江省瑞安市,瑞安市地理坐标27°40′~28°00′N,120°10′~120°15′E,全年无严寒酷暑,冬短夏长,四季分明,雨水充沛。全年平均气温17.9 ℃,年平均降水量1 110~2 200 mm,历史年平均降水量1 527.2 mm。挑选外观完好且直径和长度相近的绿竹笋,放置于泡沫箱中6 h内运至实验室。绿竹笋样品切除基部2~3 cm不可食用部分,小心剥除笋壳,用自来水清洗干净沥干,处理组浸入5 mmol·L−1草酸溶液中10 min(该草酸浓度和处理时间为前期预实验获得的最佳处理参数),对照组浸入自来水保持同等时间,取出沥干后于阴凉通风处晾干,然后放于塑料筐中,套0.5 mm厚度的聚乙烯袋,不封口,置于(6±1) ℃的恒温恒湿箱(Sanyo,MIR-554)中储藏。处理当天取样为T0,随后每天随机取样,取样时间为Tn,n=1、2、3、4、5、6 d,取笋体基部切口位置往上2~4 cm的一段笋肉用于指标测定。每个处理每次取5根笋样,重复3次。
-
过氧化氢(H2O2)质量摩尔浓度采用苏州科铭生物技术有限公司过氧化氢试剂盒测定。超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性、苯丙氨酸解氨酶(PAL)活性、过氧化物酶(POD)活性参照曹建康等[10]的方法。木质素质量分数参照鞠志国等[11]的方法。4-香豆酸-辅酶A连接酶(4-CL)活性参照毕咏梅等[12]的方法并加以修改。羟基肉桂醇脱氢酶(CAD)活性参照CAI等[13]的方法并加以修改。基因表达量采用大连TaKaRa公司SYBR Premix Ex TapTM(TliRNaseH Plus)Kit测定。
-
用Excel进行数据统计分析及图表制作;用SPSS 22进行0.05水平差异显著性分析。
-
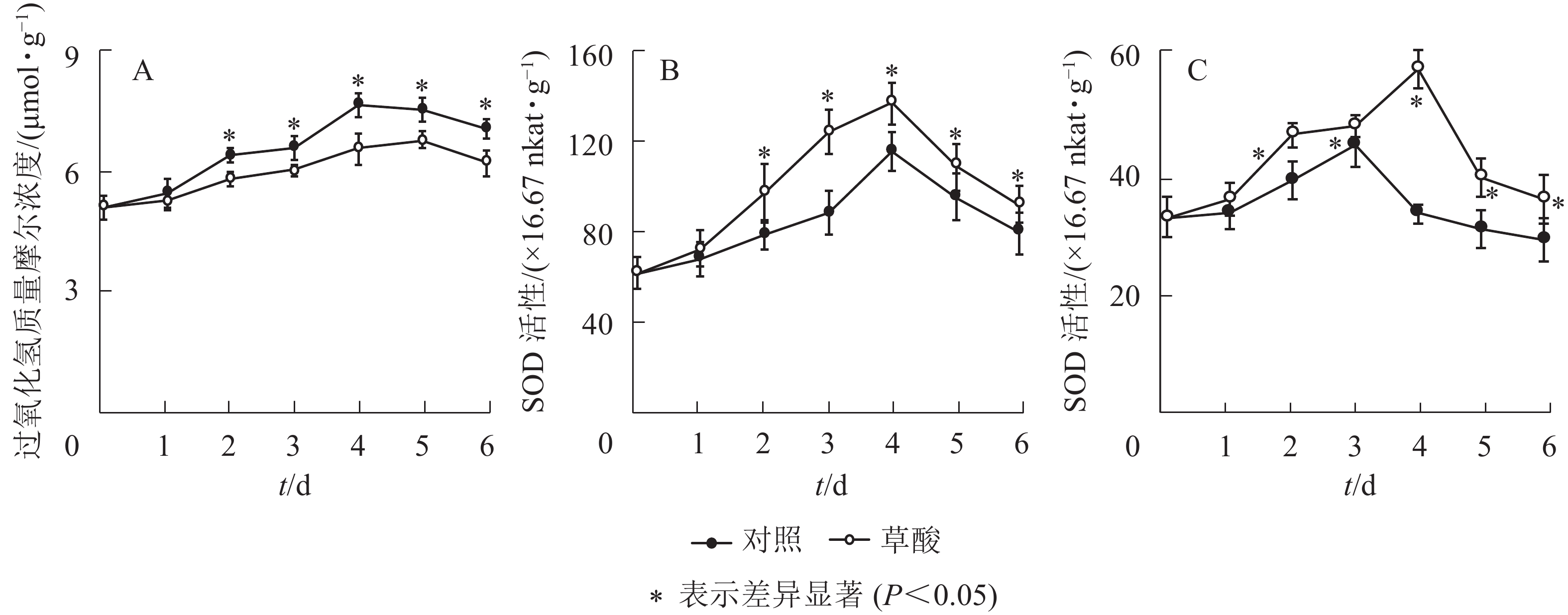
由图1A可知:草酸处理组与对照组的绿竹笋H2O2质量摩尔浓度在冷藏期间的变化趋势基本一致,总体上呈先上升后下降的趋势,草酸处理组的H2O2质量摩尔浓度在冷藏2~6 d时显著低于对照组(P<0.05),比对照组分别降低了9.2%、8.5%、14.3%、10.2%和12.2%。由图1B和C可知:草酸处理组与对照组的绿竹笋SOD和CAT活性在冷藏期间的变化趋势基本一致,总体上呈先上升后下降的趋势;草酸处理组的SOD活性在冷藏2~6 d时显著高于对照组的SOD活性(P<0.05),比对照组分别提高了23.4%、41.5%、18.1%、14.6%和16.4%;草酸处理组的CAT活性在冷藏2~6 d时显著高于对照组(P<0.05),比对照组分别提高了20.2%、7.1%、78.4%、29.7%和25.6%。由此表明:外源草酸处理可以提高绿竹笋体内抗氧化酶SOD、CAT的活性,并延缓H2O2的积累。
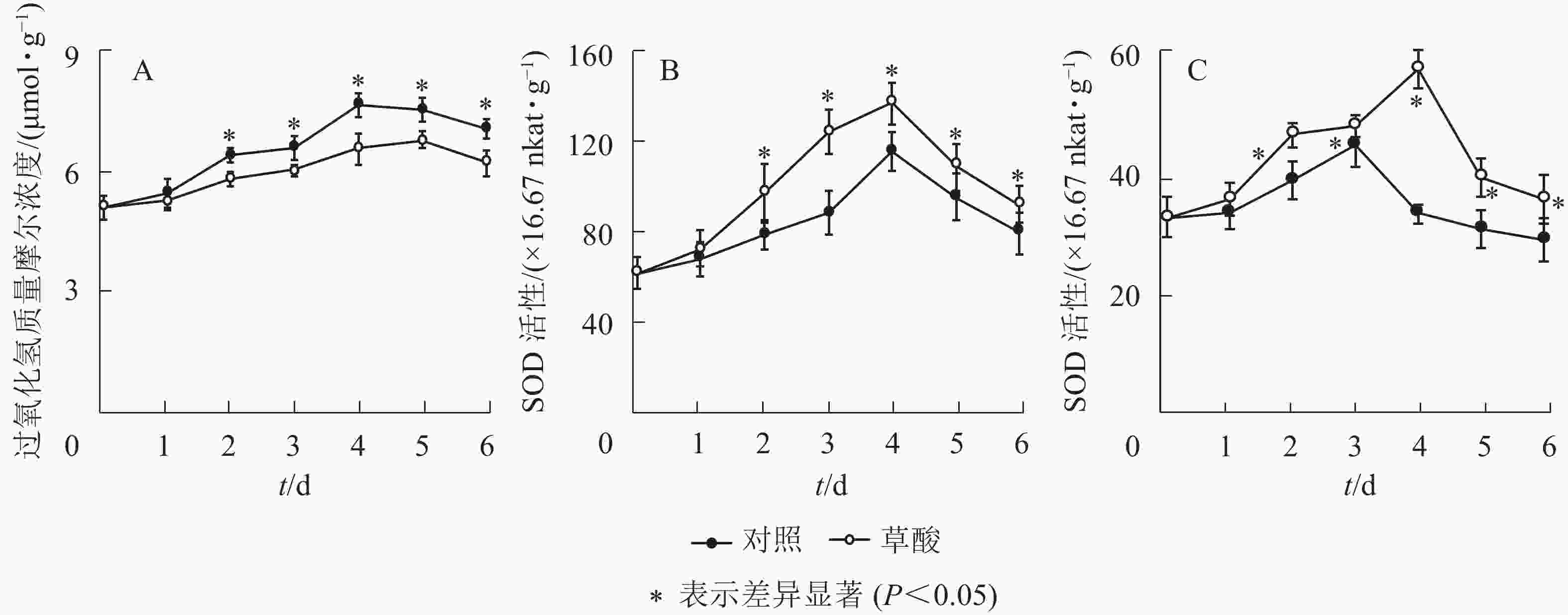
图 1 低温下草酸对绿竹笋H2O2质量摩尔浓度、SOD和CAT活性的影响
Figure 1. Effects of oxalic acid treatment on H2O2 content, SOD and CAT activities of bamboo shoots at low temperature
-
如图2可知:草酸处理组与对照组的绿竹笋木质素质量分数和硬度在冷藏期间的变化趋势基本一致,总体上呈上升趋势。由图2A可知:草酸处理组的木质素质量分数在冷藏的2、3和5 d时显著低于对照组(P<0.05),比对照组分别降低了25.3%、17.3%、12.7%。由图2B可知:草酸处理组的硬度在冷藏的3~5 d时显著低于对照组(P<0.05),比对照组分别降低了30.5%、33.8%、16.5%。由此说明:外源草酸处理可以延缓绿竹笋木质素的积累和硬度的上升。
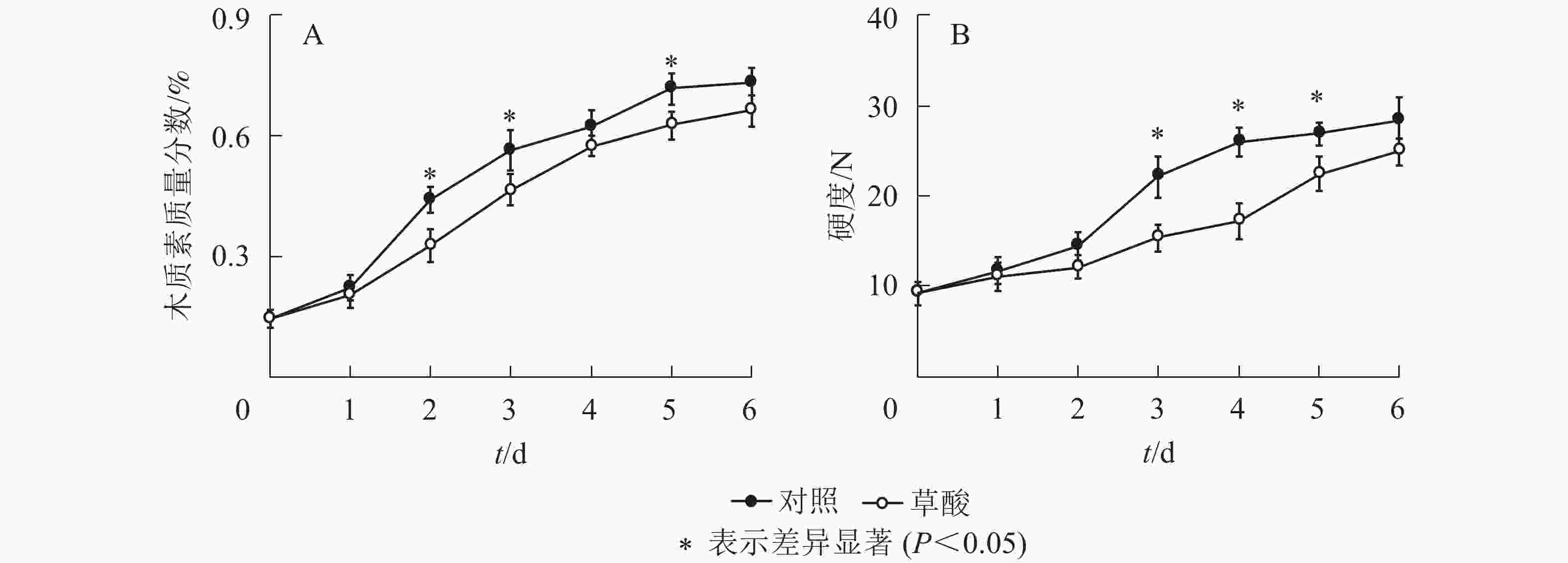
图 2 低温下草酸对绿竹笋木质素质量分数和硬度的影响
Figure 2. Effects of oxalic acid treatment on lignin content and firmness of bamboo shoots at low temperature
-
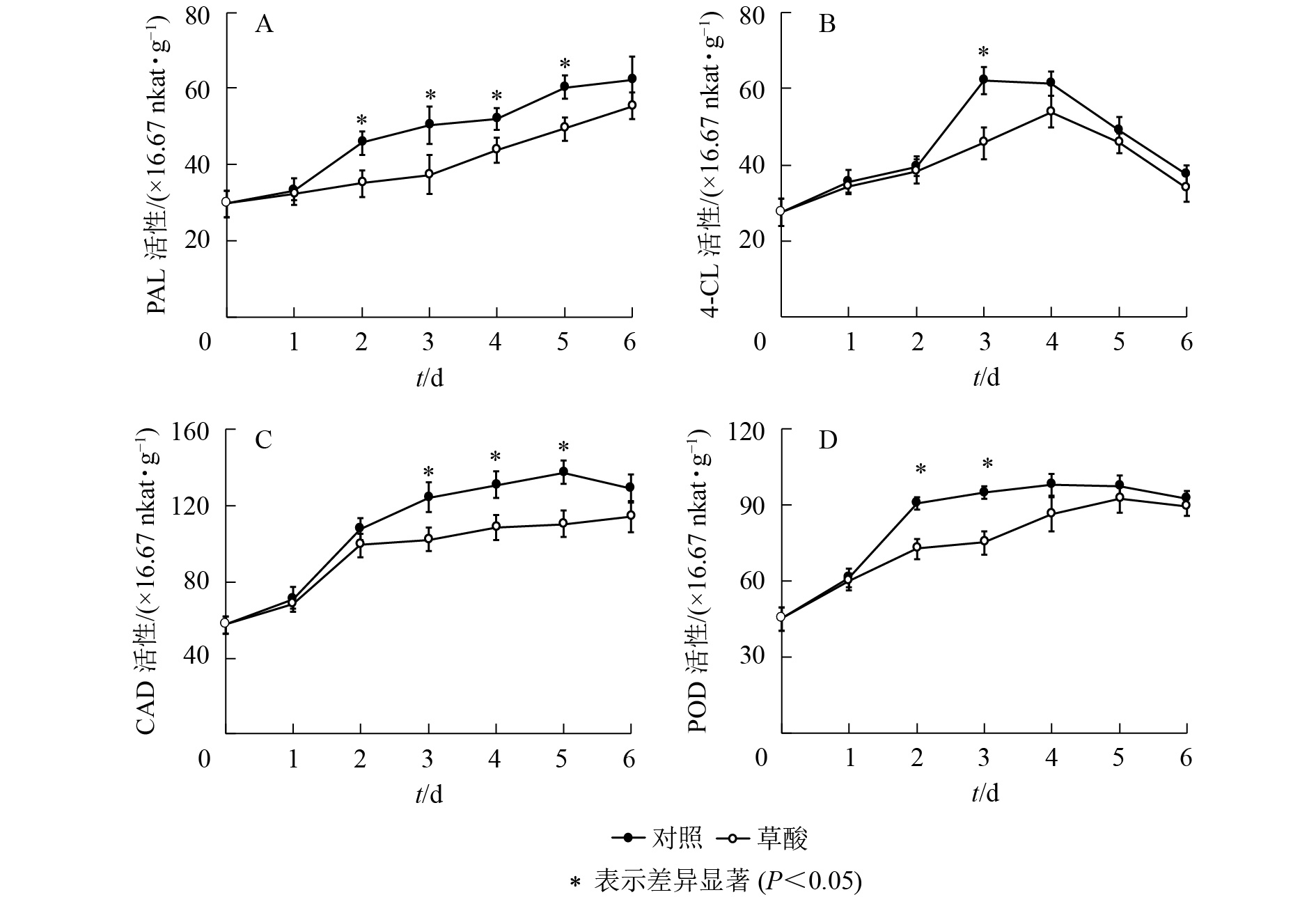
如图3可知:草酸处理组与对照组的绿竹笋PAL、4-CL、CAD、POD活性在冷藏期间的变化趋势基本一致。如图3A可知:PAL活性总体上呈上升的趋势,草酸处理组的PAL活性在冷藏2~5 d时显著低于对照组(P<0.05),比对照组分别降低了23.3%、25.8%、16.0%、18.2%。如图3B可知:4-CL活性总体上呈先上升后下降的趋势,草酸处理组的4-CL活性在冷藏3 d时显著低于对照组(P<0.05),比对照组降低了26.3%。如图3C可知:CAD活性总体上呈先上升后下降的趋势,草酸处理组的CAD活性在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了17.7%、17.0%、19.7%。如图3D可知:POD活性总体上呈先上升后下降的趋势,草酸处理组的POD活性在冷藏2~3 d时显著低于对照组(P<0.05),比对照组分别降低了19.9%、21.1%。由此表明:外源草酸处理可以抑制绿竹笋木质素合成关键酶PAL、4-CL、CAD、POD活性的上升。
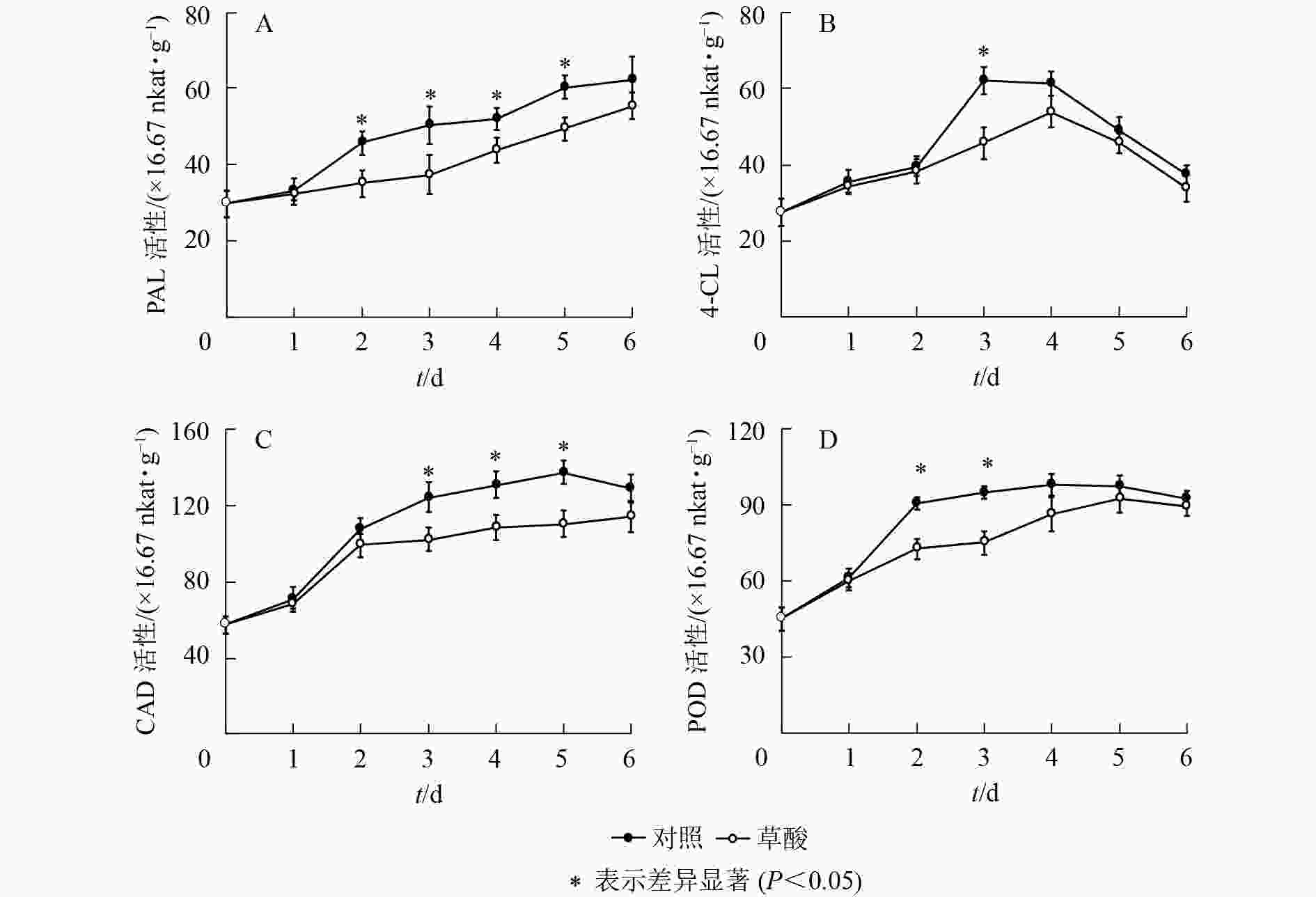
图 3 低温下草酸对绿竹笋PAL、4-CL、CAD和POD活性的影响
Figure 3. Effects of oxalic acid treatment on PAL, 4-CL, CAD and POD activities of bamboo shoots at low temperature
-
如图4可知:草酸处理组与对照组的绿竹笋PAL、4-CL、CAD、POD基因相对表达量在冷藏期间的变化趋势基本一致。由图4A可知:PAL基因相对表达量总体上呈先下降后上升再下降的趋势,草酸处理组的PAL基因相对表达量在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了37.4%、28.1%、45.4%。由图4B可知:4-CL基因相对表达量总体上呈上升的趋势,草酸处理组的4-CL基因相对表达量在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了45.0%、17.5%、28.1%。由图4C可知:CAD基因相对表达量总体上呈先上升后下降的趋势,草酸处理组的CAD基因相对表达量在冷藏2~3 d时显著低于对照组(P<0.05),比对照组分别降低了34.7%、48.2%。由图4D可知:POD基因相对表达量总体上呈下降的趋势,草酸处理组的POD基因相对表达量略低于对照组,差异不显著(P>0.05)。由此说明:外源草酸处理可以抑制PAL、4-CL、CAD和POD基因的表达。
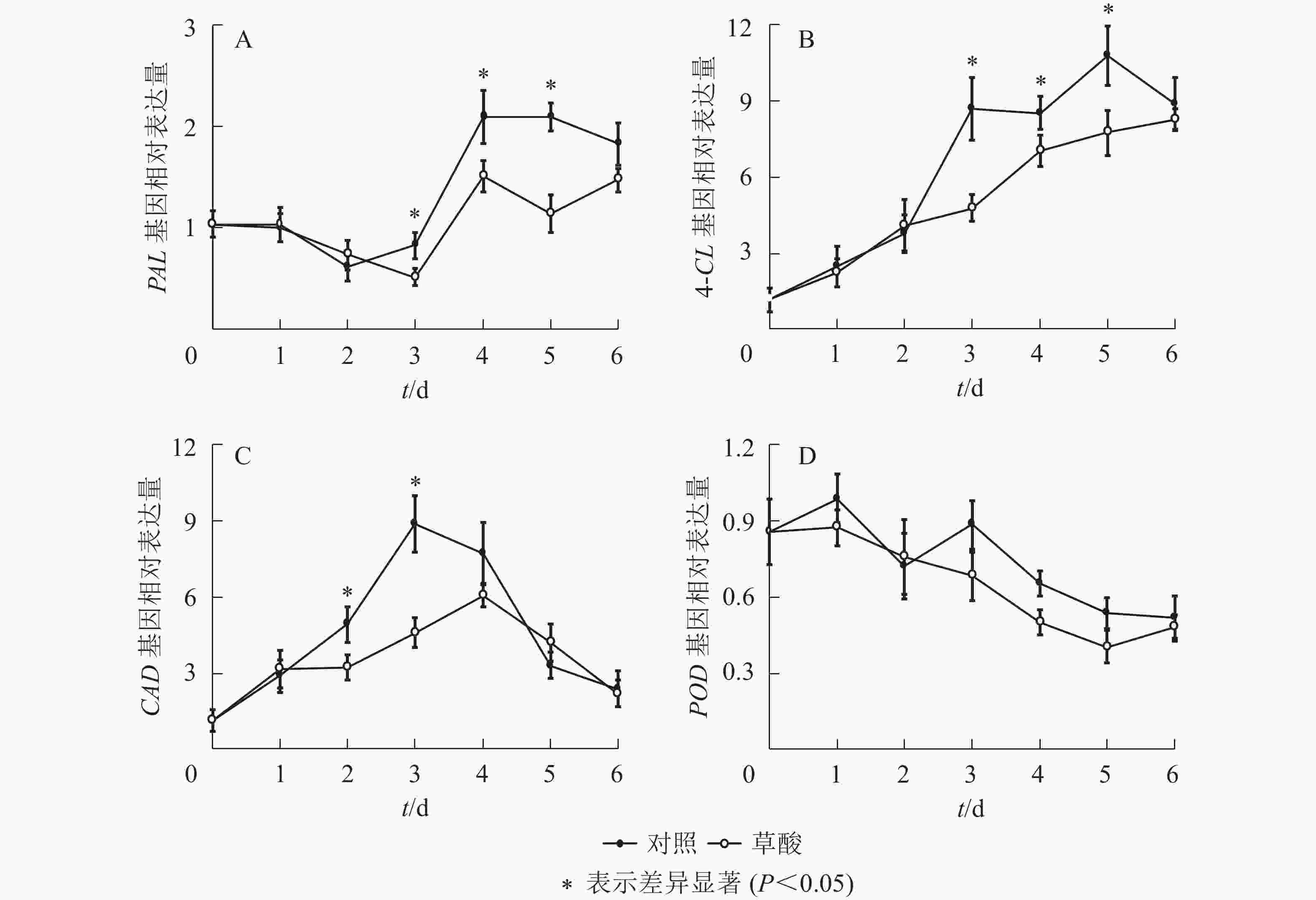
图 4 低温下草酸对绿竹笋PAL、4-CL、CAD和POD基因相对表达量的影响
Figure 4. Effects of oxalic acid treatment on PAL, 4-CL, CAD and POD relative expression of bamboo shoots at low temperature
-
本研究表明:外源草酸处理可以提高绿竹笋体内抗氧化酶SOD、CAT的活性并延缓H2O2质量摩尔浓度的积累;可以延缓木质素的积累和硬度的上升;既可以抑制绿竹笋木质素合成关键酶PAL、4-CL、CAD、POD活性的上升,又可以抑制PAL、
4-CL、CAD和POD基因的表达;可以减缓绿竹笋的木质化进程,延缓绿竹笋品质的下降,延长绿竹笋的保鲜期。 植物在受到外界逆境胁迫时会产生活性氧(ROS),导致细胞膜脂过氧化,破坏生物膜的结构和功能,严重则会致使细胞程序性死亡。为了抵御ROS的伤害,植物体会激活体内抗氧化防御系统,提高抗氧化酶的活性,以消除ROS[14−17]。本研究结果显示:在低温储藏下,草酸处理组的去壳绿竹笋笋肉组织中的H2O2质量摩尔浓度显著低于对照组(P<0.05),较低的H2O2质量摩尔浓度有助于减轻ROS对笋肉细胞的毒害。草酸处理组的去壳绿竹笋笋肉组织中的SOD和CAT活性显著高于对照组(P<0.05)。梁春强等[18]研究认为:适合浓度的草酸处理不但可以显著提高氧化酶SOD和CAT的活性,还可以有效降低O2-的生成速率和H2O2的含量。
木质素为植物次生代谢的产物,属酚类化合物,是构成细胞壁次生结构的主要成分。竹笋在采后非常容易老化变质,失去食用价值,其中一个重要的因素就是木质化。木质化主要表现为笋体组织硬度上升,木质素质量分数增加,笋体发生褐变等[19−21]。除了竹笋,茭白Zizania latifolia、芦笋Asparagus officinalis等蔬菜在采后储藏过程中均有木质化的发生[22−25]。本研究发现:低温条件下储藏,草酸处理不但可以抑制绿竹笋硬度的上升,还能够减缓木质素的生物合成,抑制采后去壳竹笋的木质化,维持较好的口感品质,延长竹笋可食用期限。与王琪等[26]研究的采用外源草酸抑制带壳雷竹Phyllostachys praecox笋
木质素的合成累积以及硬度的上升的结论相似。 PAL、4-CL、CAD和POD等4种酶与木质素的合成有密切的关系,其活性的升高都能促进木质素的合成,提高组织的木质化程度。本研究结果表明:低温条件下,经草酸处理的绿竹笋的PAL、4-CL、CAD和POD的活性均低于对照组,草酸处理在总体上明显抑制了去壳绿竹笋的木质素生物合成关键酶的活性。这与沈玫等[27]研究的利用外源草酸抑制带壳绿竹笋PAL和POD活性的结论类似。
果蔬的木质化进程受编码木质素代谢的关键酶基因的表达调控。LU等[28]研究发现:PpCAD1和PpCAD2这2个基因主要调控梨Pyrus pyrifolia的木质素代谢生物合成。不同种类的果蔬,调控木质素合成关键酶的基因也不同,SHAN等[29]研究表明,调控枇杷Eriobotrya japonica的木质素代谢基因主要是EjCAD1、EjPOD、EjPAL2这3个基因。此外,大多数研究人员认为是PAL、4-CL、CAD和POD的活性与其对应基因的表达在协同调控木质素的合成[30−31]。本研究结果表明:草酸处理在冷藏的中后期能够显著抑制去壳绿竹笋的木质素合成关键酶PAL、CAD和4-CL的基因表达水平,可以抑制POD的基因表达水平。结合硬度、木质素质量分数以及对应酶的活性的变化趋势分析可见,PAL、4-CL、CAD基因表达与硬度、木质素质量分数以及酶活性的变化趋势相一致,POD基因表达与硬度、木质素质量分数以及酶活性的变化趋势相反,说明绿竹笋木质素合成关键酶的基因表达和活性变化同步调控木质素的生物合成。LU等[28]研究发现:利用氯化钙可以抑制梨果肉中PpCAD1和PpCAD2基因表达,从而调控木质素的生物合成途径,最终达到抑制梨的硬化的效果,这与本研究结果类似。另外,SEWALT等[32]研究发现,烟草Nicotiana tabacum的PAL基因在受到抑制之后,不但木质素质量分数明显下降,PAL的活性也明显降低。本研究中草酸处理既抑制了木质素代谢关键酶活性,又抑制了对应基因表达水平,从2个方面抑制了去壳绿竹笋笋肉硬度的上升和木质素的累积,延缓了冷藏过程中竹笋食用品质的下降。
Effects of exogenous oxalic acid on antioxidant enzymes and lignification of Bambusa oldhami shoots
-
摘要:
目的 研究外源草酸处理对低温下去壳绿竹笋Bambusa oldhami的保鲜效果及其机制。 方法 用5 mmol·L−1草酸浸泡去壳绿竹笋10 min,置于(6±1) ℃下,定期测定去壳绿竹笋的过氧化氢(H2O2)质量摩尔浓度、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性、硬度、木质素质量分数和木质素代谢关键酶(PAL、4-CL、CAD、POD)的活性及其基因表达水平。 结果 5 mmol·L−1草酸溶液浸泡去壳绿竹笋10 min,能够提高去壳绿竹笋组织内SOD和CAT的活性并延缓H2O2的累积,抑制去壳绿竹笋木质素代谢关键酶(PAL、4-CL、CAD、POD)的基因表达水平和活性的上升,并显著降低木质素质量分数和硬度的上升。 结论 外源草酸处理通过抑制木质素代谢和提高抗氧化酶系统阻止去壳绿竹笋采后木质化进程,从而延缓了其冷藏期内品质的下降。图4参32 Abstract:Objective The purpose is to explore the preservation effect and mechanism of exogenous oxalic acid treatment on bamboo(Bambusa oldhami) shoots at low temperature. Method The peeled bamboo shoots were soaked in 5 mmol·L−1 oxalic acid solution for 10 minutes and stored at (6±1) ℃. The effects of oxalic acid treatment on H2O2 content, SOD activity, CAT activity, hardness, lignin content, key enzymes of lignin metabolism (PAL, 4-CL, CAD, POD) activities as well as their gene expression levels were measured regularly. Result Soaking the bamboo shoots in 5 mmol·L−1 oxalic acid solution for 10 minutes increased the activities of SOD and CAT, delayed the accumulation of H2O2, inhibited the increase of key enzymes of lignin metabolism (PAL, 4-CL, CAD, POD) activities and their gene expression level, and significantly reduced the accumulation of lignin and hardness increase in bamboo shoots without sheaths. Conclusion Exogenous oxalic acid treatment can retard the lignification process in bamboo shoots without sheaths by inhibiting lignin metabolism and improving antioxidant enzyme system. Thus, the quality decline during the cold storage could be postponed. [Ch, 4 fig. 32 ref.] -
Key words:
- botany /
- Bambusa oldhami shoots /
- exogenous oxalic acid /
- lignin metabolism /
- antioxidant system
-
图 1 低温下草酸对绿竹笋H2O2质量摩尔浓度、SOD和CAT活性的影响
Figure 1 Effects of oxalic acid treatment on H2O2 content, SOD and CAT activities of bamboo shoots at low temperature
图 2 低温下草酸对绿竹笋木质素质量分数和硬度的影响
Figure 2 Effects of oxalic acid treatment on lignin content and firmness of bamboo shoots at low temperature
图 3 低温下草酸对绿竹笋PAL、4-CL、CAD和POD活性的影响
Figure 3 Effects of oxalic acid treatment on PAL, 4-CL, CAD and POD activities of bamboo shoots at low temperature
-
[1] 郑蓉. 产地绿竹笋品质及土壤养分的主成分与典型相关分析[J]. 浙江农林大学学报, 2012, 29(5): 710 − 714. ZHENG Rong. Canonical correlation and principal components analysis of different production areas of Dendrocalamopsis oldhami with quality indicators of bamboo shoots and its soil nutrients [J]. J Zhejiang A&F Univ, 2012, 29(5): 710 − 714. [2] 郑蓉,郑维鹏,方伟,等. 绿竹笋形态性状与营养成分的产地差异分析[J]. 浙江农林大学学报, 2010, 27(6): 845 − 850. ZHENG Rong, ZHENG Weipeng, FANG Wei, et al. Shapes and nutrients of Dendrocalamopsis oldhami bamboo shoots in 12 production areas [J]. J Zhejiang A&F Univ, 2010, 27(6): 845 − 850. [3] 孙茂盛, 鄢波, 徐田, 等. 竹类植物资源与利用[M]. 北京: 科学出版社, 2015: 64-74. [4] 余学军,窦可,章兆福,等. 不同保鲜预处理对绿竹笋呼吸速率的影响[J]. 浙江农林大学学报, 2007, 24(4): 424 − 427. YU Xuejun, DOU Ke, ZHANG Zhaofu, et al. Effects of different fresh pretreatments on the respiration intensity of Dendrocalamopsis oldhami shoots [J]. J ZhejiangA&F Univ, 2007, 24(4): 424 − 427. [5] 余学军. 绿竹笋采后生理及鲜笋保鲜技术研究[D]. 南京: 南京林业大学, 2004. YU Xuejun. A Study on Postharvest Physiology and Storage Techniques of Dendrocalamopsis oldhami[D]. Nanjing: Nanjing Forestry University, 2004. [6] KAYASHIMA T, KATAYAMA T. Oxalic acid is available as a natural antioxidant in some systems [J]. Biochim Biophys Acta, 2002, 1573(1): 1 − 3. [7] 覃海元,杨昌鹏,陈智理,等. 草酸与柠檬酸抑制鲜切香蕉酶褐变的比较研究[J]. 食品工业科技, 2011, 32(4): 75 − 77. QIN Haiyuan, YANG Changpeng, CHEN Zhili, et al. Study on inhibition of enzymatic browning of fresh-cut banana by oxalic acid and citric acid [J]. Sci Technol Food Ind, 2011, 32(4): 75 − 77. [8] ZHENG Xiaolin, TIAN Shiping. Effect of oxalic acid on control of postharvest browning of litchi fruit [J]. Food Chem, 2006, 96(4): 519 − 523. [9] 杨莹. 草酸处理对去皮荸荠块茎的保鲜效果及其作用机制研究[D]. 杭州: 浙江工商大学, 2016. YANG Ying. Effects of Oxalic Acid Treatment on Peeled Chinese Water-Chestnut during Storage at Low Temperature[D]. Hangzhou: Zhejiang Gongshang University. 2016. [10] 曹建康, 姜微波, 赵玉梅. 果蔬采后生理生化实验指导[M]. 北京: 中国轻工业出版社, 2007: 101-144. [11] 鞠志国. 采期对莱阳茌梨酚类物质代谢和组织褐变的影响[J]. 中国农业科学, 1991, 24(2): 63 − 68. JU Zhiguo. Effects of harvest date on phenolics metabolism and tissue browning of Laiyang Chili (Pyrus bretschnederi Rehd) [J]. Sci Agric Sin, 1991, 24(2): 63 − 68. [12] 毕咏梅,欧阳光察. 稻瘟病菌诱导物对水稻苯丙烷类途径酶系和绿原酸的诱导作用[J]. 植物生理学报, 1990(3): 18 − 20. BI Yongmei, OUYANG Guangcha. Induction effect of elicitor from Piriculariaoryzae on enzymes of phenylpropane pathway and chlorogenic acid [J]. Plant Physiol Commun, 1990(3): 18 − 20. [13] CAI Chong, XU Changjie, LI Xian, et al. Accumulation of lignin in relation to change in activities of lignification enzymes in loquat fruit flesh after harvest [J]. Postharvest Biol Technol, 2006, 40(2): 163 − 169. [14] PALIYATH G. The mechanism of membrane deterioration and disassembly during senescence [J]. Plant Physiol, 1992, 30: 789 − 812. [15] JIMENEZ A, CREISSEN G, KULAR B, et al. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening [J]. Planta, 2002, 214(5): 751 − 758. [16] GNM R S, KNOWLES N R. Maturation and ripening of fruit of Amelanchier alnifolia Nutt. are accompanied by increasing oxidative stress [J]. Annals Bot, 1998, 81(2): 203 − 211. [17] LACAN D, BACCOU J C. High levels of antioxidant enzymes correlate with delayed senescence in nonnetted muskmelon fruits [J]. Planta, 1998, 204(3): 377 − 382. [18] 梁春强,吕茳,靳蜜静, 等. 草酸处理对采后猕猴桃冷害、抗氧化能力及能荷的影响[J]. 园艺学报, 2017, 40(2): 279 − 287. LIANG Chunqiang, LÜ Jiang, JIN Mijing, et al. Effects of oxalic acid treatment on chilling injury, antioxidant capacity and energy status in harvested kiwifruits under low temperature stress [J]. Acta Hortic Sin, 2017, 40(2): 279 − 287. [19] 罗自生. 采后竹笋木质化与内源激素的关系[J]. 中国农业科学, 2006, 39(4): 792 − 797. LUO Zisheng. The relationship between lignification and endogenous hormone content of excised bamboo shoots [J]. Sci Agric Sin, 2006, 39(4): 792 − 797. [20] 余学军,裘贤龙. 不同储藏条件对绿竹笋酶活性与纤维化的影响[J]. 浙江农林大学学报, 2011, 28(3): 380 − 385. YU Xuejun, QIU Xianlong. POD, PAL activities and fibrosis of Bambusa oldhami bamboo shoots with different storage conditions [J]. J Zhejiang A&F Univ, 2011, 28(3): 380 − 385. [21] 高贵宾,顾小平,张小平,等. 微波处理对绿竹笋老化生理的影响[J]. 浙江农林大学学报, 2008, 25(5): 675 − 678. GAO Guibin, GU Xiaoping, ZHANG Xiaoping, et al. Microwave treatments on the ageing physiology of Dendrocalamopsis oldhami bamboo shoots [J]. J Zhejiang A&F Univ, 2008, 25(5): 675 − 678. [22] 房祥军,郜海燕,宋丽丽,等. 减压贮藏保持茭白采后品质及调控细胞壁物质代谢[J]. 农业工程学报, 2013, 29(12): 257 − 263. FANG Xiangjun, GAO Haiyan, SONG Lili, et al. Hypobaric storage maintains postharvest quality and regulates cell wall metabolism in water bamboo shoot [J]. Transac Chin Soc Agric Eng, 2013, 29(12): 257 − 263. [23] 凌建刚,李娇,康孟利,等. 弱酸性电位水在茭白防腐保鲜中的应用[J]. 食品科学, 2015, 36(22): 250 − 254. LING Jiangang, LI Jiao, KANG Mengli, et al. Application of slightly acidic electrolyzed water (SAEW) in preservation of Zizania latifolia stems [J]. Food Sci, 2015, 36(22): 250 − 254. [24] 宋秀香,鲁晓翔,陈绍慧,等. 冰温结合纳他霉素对绿芦笋采后生理品质的影响[J]. 食品科学, 2013, 34(24): 294 − 298. SONG Xiuxiang, LU Xiaoxiang, CHEN Shaohui, et al. Effect of natamycin combined with ice-temperature storage on postharvest physiological quality of green asparagus [J]. Food Sci, 2013, 34(24): 294 − 298. [25] 龚佳惠,陈贝莉,董欢欢,等. L-精氨酸对绿芦笋贮藏品质及生理生化特性的影响[J]. 保鲜与加工, 2017, 17(1): 7 − 11. GONG Jianghui, CHEN Beili, DONG Huanhuan, et al. Effects of L-arginine on storage qualities and physio-biochemical characteristics of Asparagus officinalis L [J]. Storage Process, 2017, 17(1): 7 − 11. [26] 王琪,郑小林,励建荣,等. 草酸处理对冷藏雷竹笋保鲜效果的影响[J]. 中国食品学报, 2012, 12(11): 84 − 89. WANG Qi, ZHENG Xiaolin, LI Jianrong, et al. Effect of oxalic acid treatment on preservation of bamboo shoots during storage at lower temperature [J]. J Chin Inst Food Sci Technol, 2012, 12(11): 84 − 89. [27] 沈玫,王琪,赵宇瑛,等. 外源草酸对冷藏绿竹笋的保鲜效果及其生理基础[J]. 园艺学报, 2013, 40(2): 355 − 362. SHEN Mei, WANG Qi, ZHAO Yuying, et al. Physiological roles of exogenous oxalic acid in preservation of bamboo shoots during cold storage [J]. Acta Hortic Sin, 2013, 40(2): 355 − 362. [28] LU Guilong, LI Zhijun, ZHANG Xinfu, et al. Expression analysis of lignin-associated genes in hard end pear (Pyrus pyrifolia Whangkeumbae) and its response to calcium chloride treatment conditions [J]. J Plant Growth Regul, 2015, 34(2): 251 − 262. [29] SHAN Lanlan, LI Xian, WANG Ping, et al. Characterization of cDNAs associated with lignification and their expression profiles in loquat fruit with different lignin accumulation [J]. Planta, 2008, 227(6): 1243 − 1254. [30] ANTEROLA A M, LEWIS N G. Trends in lignin modification: a comprehensive analysis of the effects of genetic manipulations/mutations on lignification and vascular integrity [J]. Phytochemistry, 2002, 61(3): 221 − 294. [31] BAUCHER M, HALPIN C, PETIT-CONIL M, et al. Lignin: Genetic engineering and impact on pulping [J]. Crit Rev Biochem Mol Biol, 2003, 38(4): 305 − 350. [32] SEWALT V, NI W, BLOUNT J W, et al. Reduced lignin content and altered lignin composition in transgenic tobacco down-regulated in expression of l-phenylalanine ammonia-lyase or cinnamate 4-hydroxylase [J]. Plant Physiol, 1997, 115(1): 41 − 50. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190337
 点击查看大图
点击查看大图
计量
- 文章访问数: 3303
- HTML全文浏览量: 1113
- PDF下载量: 55
- 被引次数: 0
