





-
木材染色技术就是将染料或染料与其他改性剂复配后,通过常压热浸渍、减压或加压的方式固定在木材纤维上,从而改变木材或单板的材色[1-3]。染色后的木材或单板既可以模拟珍贵树种材色,也可满足产品室内设计需求,木材染色产品已被广泛应用在家具、地板、室内外装饰和运动器材等领域[4-5]。目前,木材工业生产中主要应用酸性染料进行染色,但酸性染料仅对木质素染色效果良好,存在上染率低、易流失和染色单板耐水性差等问题[6-7]。活性染料分子上的活性基能与木材纤维素、半纤维素和木质素上的活性基团发生共价键反应,上染率高,且染色单板具有较好的耐水性、耐光性、表面胶合性能和满意的染色效果,因此,活性染料必将广泛应用于木材工业[8-13]。在活性染料之中,具有异双活性基的M型染料染色木材的固色率较高,一般在染色温度70~80 ℃、硫酸钠添加量40 g·L−1、碳酸钠添加量20 g·L−1的工艺下可以获得较佳的上染率[14-16]。由此可见,为了提高活性染料染色木材的上染率和固色率,大量的电解质盐和固色碱被应用于染色工艺中,带来淡水盐化和污水处理难度增加等问题;强碱固色会导致染料易水解,木片烘干时易发生氧化损伤,从而影响染色效果[17]。在纺织工业中,采用低盐、低碱染色或无盐、中性染色工艺已成为重要的研究内容。一是对染料分子进行修饰和组装,例如适当减少活性染料结构中的磺酸基团的数目、提高染料母体分子的芳环共平面性、改变活性基等;二是对纤维改性,提高对染料的吸附能力,例如纤维表面阳离子化或酰胺化[18];三是开发新型交联剂和盐、碱替代物,提高染料上染率,取代传统无机盐和碱[19-20]。相比于其他2种方法,应用低盐和低碱活性染料染色织物工艺简单、成本低廉、染色效果较好,是主要的技术方法之一。与常规活性染料相比,使用少量无机盐获得较高上染率的活性染料被称为低盐活性染料,其对电解质盐敏感度低,但固色碱与常规染料用量相同;低碱活性染料则是在较少固色碱用量下能获得较高上染率,但其电解质盐用量与常规活性染料一致。低盐活性染料产品的盐用量是传统活性染料的25%~50%,低碱活性染料产品的固色碱用量是传统活性染料的10%~20%[18, 21]。目前,在木材染色领域,低盐、碱染色工艺尚未见相关报道。本研究选用低盐和低碱活性红染料染色渗透性较差的柞木Xylosma japonicum单板,以对木材染色性能好的活性红染料M-3BE作对照,通过染料上染性能和固色机理的对比分析,研究低盐、低碱染料染色木材的适用性,从工艺源头减少废水中染色助剂用量,为单板染色工业绿色节能生产提供新思路。
-
柞木单板购买自黑龙江省牡丹江市,单板尺寸1 000 mm×110 mm×1 mm(长×宽×厚),制备成50 mm×50 mm试件,单板平均相对含水率为4.92%。
试剂包括无水硫酸钠(分析纯)、无水碳酸钠(分析纯)、皂洗剂(德桑化工)。活性红(M-3BE)、低盐活性大红(SNE)、低碱活性红(LA)染料购买自江苏锦鸡染料有限公司。SNE染料分子式为C26H21N5Na4O19S6,分子量为991.82,具有2个β-羟乙基砜硫酸酯活性基;LA染料分子式为C48H42Cl2N14Na6O27S8,分子量1709.89,具有2个一氯均三嗪和2个β-羟乙基砜硫酸酯活性基。
-
将木片置于浴比1∶40、活性红染料质量分数1.0%(染料占木片质量百分比)、硫酸钠添加量40 g·L−1的染液中,利用恒温水浴锅(上海一恒HWS-26)加热染液,30 ℃入染,70 ℃染色2.5 h后,加入20 g·L−1碳酸钠固色1 h。固色结束后,置于80 ℃、浴比1∶40、质量浓度1 g·L−1的皂洗液中10 min,之后水洗。染色后的试件先气干,而后置于铺垫铝箔纸的不锈钢压板中放入鼓风干燥箱内70 ℃干燥。
-
SNE染料染色仅改变硫酸钠添加量,分别为4、8、12、16和20 g·L−1,其他条件与M-3BE染料染色方法相同,做低盐活性大红染料染色柞木单板单因素试验。LA染料染色仅改变碳酸钠添加量,分别为1.0、2.5、5.0、7.5和10.0 g·L−1,其他条件与M-3BE染料染色方法相同,做低碱活性红染料染色柞木单板单因素试验。
-
在染色柞木单板上染率最大的工艺条件下,测试SNE和LA染料的上染性能(直接性、反应性和固色率);M-3BE染料上染性能按照1.2.1的方法。一般采用木片入染30 min上染率表示染料直接性;用固色、皂洗和水洗完成后的上染率表示固色率;用固色10 min的上染率表示反应性[14]。颜色、红外光谱、热重分析及扫描电镜测试所用试件均为该试验中固色完成后的试件。为降低单板个体差异对试验结果影响,盐梯度、碱梯度试验所用试件均含有同一单板相近位置的木片,上染性能试验所用试件均含有同一单板相近位置的木片。每个试验重复3次,每次放入5块木片。
-
利用紫外可见分光光度计(Agilent UV-Cary 100)分别测定空白染液和染色后混合液的吸光度,按照式(1)计算上染率E。
$$ E=(1-\frac{{A}_{\mathrm{i}}{N}_{\mathrm{i}}}{{A}_{0}{N}_{0}})\times 100\%{\text{。}} $$ (1) 式(1)中:A0为空白染液的吸光度;N0为空白染液的稀释倍数;Ai为染色后混合染液的吸光度;Ni为混合染液的稀释倍数。
-
采用CIE(1976)L*、a*、b*系统,利用色差仪(日本柯尼卡美能达公司CM-2300d)测量试件染色前后表面固定3点的L*、a*、b*值,按式(2)计算色差
$ \Delta $ E。$$ \Delta E=\sqrt{{({\Delta L}^{*})}^{2}{+{({\Delta a}^{*})}^{2}+({\Delta b}^{*})}^{2}}{\text{。}} $$ (2) 式(2)中:L*为明度;a*为色饱和度红绿值,正值表示红色,负值表示绿色;b*为色饱和度黄蓝值,正值表示黄色,负值表示蓝色;
$ \Delta $ L*、$ \Delta $ a*、$ \Delta $ b*为染色后的$L_2^* $ 、$a_2^* $ 、$b_2^* $ 与染色前$L_1^* $ 、$a_1^* $ 、$b_1^* $ 的差值。 -
将3种染料粉末与溴化钾粉末研磨然后压片制得参照试件,利用傅里叶变换红外光谱仪(Bruker Optics Tensor-27)进行测试,扫描32次,谱图分辨率为2 cm−1;在距单板横向端面4 cm、纵向端面4 cm处选取5 mm×5 mm×1 mm(长×宽×厚)的样品制成染色柞木和未处理柞木红外光谱测试样,在试样表面扫描64次,谱图分辨率为2 cm−1,扫描范围为500~4 000 cm−1。
-
将染色柞木和未处理柞木片粉碎、研磨后过40目筛,取4~6 mg木粉在热重分析仪(Perkin-Elmer TGA-7)上测定,在氮气(N2)下以10 ℃·min−1的速率从室温加热到400 ℃。
-
为减小木材试件各位置染色效果差异对测试结果的影响,在距单板横向端面4 cm、纵向端面4 cm处取样,制备成尺寸5 mm×5 mm×1 mm(长×宽×厚)的试样,镀金处理后,放入场发射扫描电子显微镜(FEI XL-30)电镜室内对试件的横、断面进行观察。
-
由表1可以看出:盐添加量对SNE染料的上染率有一定影响,但影响较弱;随着盐添加量的增加,上染率增大,但盐添加量提高5倍时,上染率仅提高了1.12倍,可见低盐染料对盐的用量敏感度较低。电解质盐电离产生的阳离子能够中和SNE染料的阴离子基团和木材纤维表面的负电荷,降低染料与木材纤维之间的库伦斥力,提高上染率;SNE染料阴离子基团较少,当质量浓度过高时,钠离子沉积在木材表面,阻碍染料分子与木材结合,染料上染率提升不明显。随着碳酸钠添加量增加,LA染料染色柞木单板的上染率呈先升高后快速降低的趋势,碳酸钠添加量为2.5 g·L−1时,染料上染率最大。碳酸钠用量少,则降低其催化作用;碳酸钠用量高,则染液不稳定,染料加速水解,与木材纤维反应性降低。
表 1 SNE和LA染料的上染率
Table 1. Dye-uptake of low-salt reactive dyes(SNE) and low-alkali reactive dyes(LA)
染料名称 编号 硫酸钠添加量/
(g·L−1)碳酸钠添加量/
(g·L−1)上染率/% 平均值 标准差 SNE 1 4 20 36.91 3.35 2 8 20 37.03 2.99 3 12 20 40.12 3.08 4 16 20 40.53 3.25 5 20 20 41.20 2.82 LA 1 40 1.0 9.06 1.39 2 40 2.5 21.47 1.64 3 40 5.0 16.92 0.84 4 40 7.5 8.03 0.77 5 40 10.0 7.83 0.97 -
利用表1结果,在上染率最大工艺下,即SNE染料硫酸钠添加量为20 g·L−1,LA染料碳酸钠添加量为2.5 g·L−1,测试2种染料的上染性能,以M-3BE染料作对照。利用紫外可见光分光光度计测定3种染料的最大吸收波长及吸光度,计算3种活性染料染色柞木单板的染色性能(表2)。
表 2 活性染料的最大吸收波长和染色性能
Table 2. Maximum absorption wavelength and dyeing performances of reactive dyes
染料名称 最大吸收
波长/nm直接性/% 反应性/% 固色率/% M-3BE 542 7.76±1.05 18.58±2.01 24.96±2.46 SNE 499 8.23±1.28 23.56±3.06 40.29±1.98 LA 545 12.70±1.02 16.38±3.21 21.59±1.93 从表2可知:直接性从大到小依次为LA、SNE、M-3BE,反应性和固色率从大到小依次为SNE、M-3BE、LA;当SNE染料硫酸钠用量为M-3BE染料的1/2时,固色率比M-3BE染料提高了15.33%,当LA染料的碳酸钠用量为M-3BE染料的1/8时,固色率降低了3.37%。染料染色性能与染料结构直接相关。活性染料的分子结构由母体、连接母体与活性基的桥基、活性基组成。从分子量来看,染料的分子量从大到小依次为LA、M-3BE、SNE,3种染料的最大吸收波长也反映分子结构的大小。从母体结构来看,LA染料母体为双偶氮类、结构复杂、芳环共平面性好,且引入了亲核性较强的烷胺基;SNE染料母体为双偶氮类,芳环共平面性好,且磺酸基数目最少;M-3BE染料母体为单偶氮类,芳环共平面性低于前2种染料。从活性基来看,SNE染料有2个β-羟乙基砜硫酸酯活性基,M-3BE染料有1个一氯均三嗪和1个β-羟乙基砜硫酸酯异双活性基,LA染料有4个可反应的活性基。
活性染料的直接性与染料的母体结构有关,通过适当增大染料分子,提高染料母体分子结构中的芳环共平面性,减少阴离子基数目,则可以提高染料的直接性[22-23]。分子结构大、芳环共平面性好的LA染料的直接性最好;SNE染料的芳环共平面性好,且与木材纤维表面负电荷产生斥力作用阴离子基磺酸基的数目少,即使在无机盐用量低时其直接性仍高于M-3BE染料。染料的固色率与染料的母体结构、活性基种类及数目、染料的扩散性有关。母体引入高亲核性结构、增加活性基数量,可以提高固色率[24],LA染料在纯碱用量较低时,反应率和固色率接近M-3BE染料,通过适当调整工艺可以实现单板的低碱固色。然而,染料分子结构越复杂、分子量越大、芳环共平面性越强,染料分子间的氢键和范德华力越大,越发生聚集,从而会引起染料的水溶性、扩散性和渗透性降低,不易向木材内部渗透[9]。分子量较小的SNE染料更易于向木材内部渗透,不仅能够附着在木材表面,也能迁移至木材内部,与木材内部纤维进行反应;另一方面,该染料具有双β-羟乙基砜硫酸酯,在上染过程中转变为双乙烯砜结构染料,该结构因不具有阴离子性的硫酸酯基,在电解质盐较低的工艺下,也具有较高的上染率,适合低盐和无盐染色[22]。
-
表3为3种染料对柞木单板的染色效果,色差从大到小依次为LA染色单板、M-3BE染色单板、SNE染色单板。染色后木材的亮度均降低,红绿指数增大,说明单板染色处理后颜色偏红;M-3BE和LA染色单板的黄蓝指数降低,说明染色处理后颜色由黄向蓝转变;SNE染色单板颜色则向黄色转变。与M-3BE相比,LA的颜色提升性好,通过合理的工艺优化,提高染料利用率,可对木材深色染色;SNE的上染率最高,但颜色提升性略低,可以用来模拟材色柔和的木材。
表 3 不同染料染色柞木单板的颜色
Table 3. Color change of X. japonicum veneers treated with different reactive red dyes
染料名称 染色后 染色前后差值 ∆E ${L_2^* }$ ${a_2^*} $ ${b_2^*} $ ∆L* ∆a* ∆b* M-3BE 47.48±3.25 32.50±4.70 5.50±2.53 − 23.29±4.09 25.59±5.10 − 15.32±2.17 38.03±5.61 SNE 46.31±1.77 34.78±3.49 24.21±1.55 − 24.15±2.71 27.41±3.95 2.72±1.53 36.68±4.68 LA 48.67±3.15 33.88±3.23 2.37±2.05 − 23.14±4.26 26.83±3.60 − 18.69±1.37 40.06±4.98 -
从图1A可见:3种染料的红外光谱图中,3 320~3 450 cm−1为N−H、O−H的伸缩振动峰[25],峰强度从大到小依次为LA染料、M-3BE染料、SNE染料,说明LA染料的N−H、O−H含量高于另外2种染料;LA染料在2 970 cm−1处特征峰为亚甲基的反对称伸缩振动峰,是由脂肪胺产生的;LA和M-3BE染料的红外谱图上,在1 750和1 502 cm−1处出现明显吸收峰,为三嗪基团的骨架振动,且LA染料的峰强度更高[26],在SNE染料谱图上没有吸收峰产生,说明SNE染料没有均三嗪基团;1 100 cm−1附近为硫酸盐S=O的振动峰。从图1B可见:柞木染色前后的红外光谱图基本形态一致,在图1A的特征吸收峰几乎被木材组分的吸收峰覆盖。3 340 cm−1为羟基的吸收峰,染色处理后木材表面羟基含量降低,一方面活性染料分子中的羟基(−OH)、氨基(−NH2)、亚氨基(−NH)与木材纤维上的−OH形成氢键,另一方面活性染料的活性基团与木材纤维发生反应(亲核加成或亲核取代)产生共价键,具有卤代均三嗪为活性基的染料与木材纤维直接通过亲核取代反应生成酯键,具有乙烯砜硫酸酯钠盐为活性基的染料与木材纤维发生亲核加成反应生成醚键[22, 27];强度从大到小依次为柞木、LA染色柞木、M-3BE染色柞木、SNE染色柞木,说明SNE染料与木材羟基的反应率最高,M-3BE染料次之,LA染料最低,与固色率试验结果一致。在1 026 cm−1处的吸收峰为醚键伸缩振动峰,波数1 738 cm−1处为木质素上醛基(−C=O)的特征峰,染色处理后,2个吸收峰的强度降低,分析是染料覆盖在木材表面,导致木纤维的醚键和醛基对红外光吸收强度降低。通过这2处吸收峰强度变化,也可以反映3种染料在木材表面附着量。3种染料在1 100 cm−1附近均出现很弱的吸收峰,而柞木的谱图中未出现此吸收峰,这是染料分子硫酸盐S=O吸收峰,说明染料分子在木材表面着色,无新官能团产生。

图 1 染料、柞木和染色柞木的红外光谱图
Figure 1. Infrared spectrum analysis diagram of dyes, X. japonicum veneer and X. japonicum veneers dyed by different reactive red dyes
-
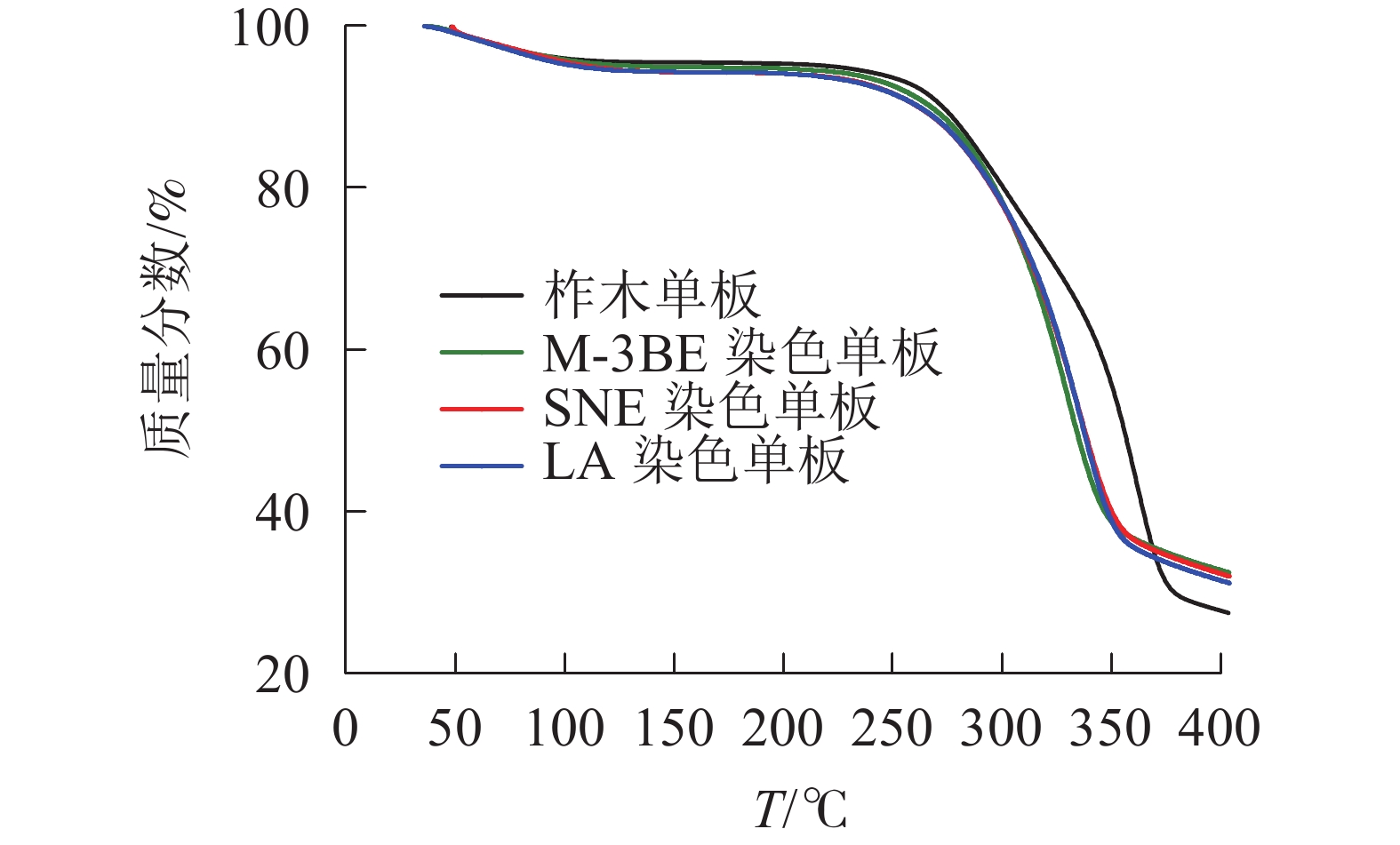
从图2可见:50~250 ℃为失水干燥阶段,主要是柞木粉中的水分受热蒸发和能量储存,无热解裂解反应发生;250~375 ℃为木粉的快速失重阶段,此时纤维素和半纤维素中某些化学键吸收足够的能量克服活化能快速断裂,形成小分子挥发分和气相的焦油;375~400 ℃为缓慢失重阶段,这时析出的挥发分较少,形成难以热解的残炭。
图 2 柞木染色前后木粉TG曲线
Figure 2. TG curves of X. japonicum veneers before and after treatment with different reactive red dyes
经过染色处理后,柞木粉的TG曲线发生了变化,主要发生在快速失重和缓慢失重阶段。在快速失重阶段,染色柞木木粉失重速率高于柞木木粉,热解温度低于柞木木粉,说明染色处理会加速木材的分解。分析原因是活性染料的引入降低了木粉中化学键断裂所需活化能,从而使纤维素、半纤维素中的化学键在低温区便能断裂。在缓慢失重阶段,柞木粉与染色柞木粉的转折温度不同,染色柞木粉在350 ℃进入缓慢失重阶段,且失重率更低,说明3种活性染料染色柞木的残炭量增多。由此可见,3种活性染料染色能降低柞木的热解活化能,降低热解温度,使热解始终曲线向低温偏移,促进结炭反应,说明活性染料与木材发生了化学反应,从而影响了木材组分的热分解特性。3种染料染色木材的热重曲线变化趋势一致,但在各阶段存在轻微差异,表明3种活性染料与木材的反应机理和生成的官能团结构相似,但因染料分子结构差异较大,导致对3种染色柞木热分解曲线略有不同。
-
图3A为未染色柞木单板的横切面图,可以观察到导管内含有侵填体,经过染色处理后,在图3B未见侵填体存在。柞木具有发达的木射线,薄壁组织或射线组织的原生质侵入接邻的界壁纹孔,形成侵填体,且可储存单宁、树脂等物质,从而充塞于导管内部[28],降低导管的渗透性。在木材染色过程中,木材经过水热处理和碱处理,部分木材导管通道被打开,从而有利于染料分子通过木射线和导管扩散到木材内部。图3C为未染色处理的柞木单板径切面图,经过染色处理后,在图3D可见较大的LA染料颗粒;在图3E可观察到大量的SNE染料颗粒堆积在木材导管内壁,覆盖了部分纹孔;在图3F发现少量的M-3BE染料颗粒存在于木射线表面,未明显观察到其附着于导管内壁。说明3种染料分子均可从木材表面扩散到木材内部,扩散程度从大到小依次为SNE、M-3BE、LA,这与染料固色率试验结果一致。与M-3BE染料相比,LA染料由于分子结构大,活性基多,且引入亲核性高的母体结构,导致染料分子易聚集,从而扩散性能降低,较难穿过细胞壁和纹孔等结构进入到细胞和导管内部;SNE染料分子量较小,渗透性强,可以良好地扩散到柞木导管内,固色率提高。

图 3 柞木染色前后的电镜图
Figure 3. SEM pictures of X. japonicum veneers before and after treatment with different reactive red dyes
-
与常规活性红染料相比,低盐活性大红染料染色柞木单板具有较高的上染率,低碱活性红染料染色柞木单板具有优良的染色效果,通过适当调整工艺,可以作为木材常用染料,降低硫酸钠和碳酸钠的用量。3种活性染料均与木材表面羟基发生化学反应,反应机理相似,但反应活性因其母体结构和活性基差异而异,因此了解染料结构与木材结合机理有助于开发木材专用活性染色剂。3种染料均可向木材内部扩散,附着于木材内部纤维上。与M-3BE染料相比,LA染料因母体结构的增大和活性基数量的增多,易聚集,导致其向柞木单板内部扩散能力降低,而SNE染料由于分子小,更易向木材内部扩散。
Application of low-alkali and low-salt reactive red dyes in Xylosma japonicum veneer dyeing
-
摘要:
目的 明确低盐和低碱活性染料应用于单板染色的适用性,可降低染液废液污染水平和生产成本。 方法 选择新型低盐活性大红(SNE)染料和低碱活性红(LA)染料对渗透性较差的柞木Xylosma japonicum单板进行染色,以应用较广的活性红(M-3BE)染料作为对照,测试3种染料染色柞木单板的直接性、反应性、固色率和染色效果,利用傅里叶红外光谱(FTIR)、热重分析仪(TG)和扫描电镜(SEM)分析其官能团、组分和细观结构的变化。 结果 与M-3BE染料相比,SNE染料硫酸钠用量降至1/2和LA染料碳酸钠用量降至1/8时,SNE染料固色率提高了15.33%、色差降低了1.35%,LA染料固色率降低了3.37%、色差提高了2.03%。染色处理后,木材表面羟基含量减少,其中低盐染料染色柞木单板羟基含量最低,除出现较弱的硫酸盐S=O吸收峰外,无新吸收峰产生。3种活性染料与木材的结合机理和产生的官能团结构相似,但因染料母体结构差异较大,导致热分解曲线略有不同。染料分子可从木材表面扩散到木材内部,扩散程度从大到小依次为SNE、M-3BE、LA。 结论 SNE染料具有较高的上染率,LA染料具有优异的颜色提升性,可用来染色木材单板,从而降低电解质盐和碱的排放。图3表3参28 Abstract:Objective This study is to analyze the applicability of low-alkali and low-salt reactive red dyes in veneer dyeing, with the purpose of reducing the pollution of dyeing wastewater and the cost of production. Method A new low-salt reactive red dye(SNE) and low-alkali reactive red dye(LA) were selected to dye Xylosma japonicum veneers with poor permeability. The widely used reactive red dye (M-3BE) was used as control. The substantivity, reactivity, fixation rate and dyeing effects were tested. Fourier transform infrared spectroscopy(FTIR), thermal gravimetric(TG) analyzer and scanning electron microscopy(SEM) were used to analyze the changes of functional groups, wood components and microstructure of specimens. Result Compared with M-3BE, when the dosages of Na2SO4 and Na2CO3 were reduced to 1/2 and 1/8 respectively, the fixation rate of SNE increased by 15.33% and color difference decreased by 1.35%, while the fixation rate of LA decreased by 3.37% and color difference increased by 2.03%. After dyeing, the hydroxyl content on the wood surface decreased, and the lowest hydroxyl content was detected on the X. japonicum wood surface dyed with SNE dyes. No new absorption peak was observed on the dyed X. japonicum veneers except weak sulfate S=O absorption peak. The mechanism and the functional group structure between three reactive dyes and wood were similar. The thermal decomposition curves were slightly different due to differences in the parent structures of the three reactive dyes. All dye molecules could diffuse to the wood interior from the wood surface, and the order of diffusion degree from large to small was SNE, M-3BE, and LA. Conclusion SNE has high dye uptake and LA has excellent color effect, which can greatly reduce the discharge of electrolyte salt and alkali. [Ch, 3 fig. 3 tab. 28 ref.] -
Key words:
- wood science /
- Xylosma japonicum veneer /
- dyeing /
- reactive dyes /
- low-salt /
- low-alkali
-
图 1 染料、柞木和染色柞木的红外光谱图
Figure 1 Infrared spectrum analysis diagram of dyes, X. japonicum veneer and X. japonicum veneers dyed by different reactive red dyes
图 2 柞木染色前后木粉TG曲线
Figure 2 TG curves of X. japonicum veneers before and after treatment with different reactive red dyes
图 3 柞木染色前后的电镜图
Figure 3 SEM pictures of X. japonicum veneers before and after treatment with different reactive red dyes
表 1 SNE和LA染料的上染率
Table 1. Dye-uptake of low-salt reactive dyes(SNE) and low-alkali reactive dyes(LA)
染料名称 编号 硫酸钠添加量/
(g·L−1)碳酸钠添加量/
(g·L−1)上染率/% 平均值 标准差 SNE 1 4 20 36.91 3.35 2 8 20 37.03 2.99 3 12 20 40.12 3.08 4 16 20 40.53 3.25 5 20 20 41.20 2.82 LA 1 40 1.0 9.06 1.39 2 40 2.5 21.47 1.64 3 40 5.0 16.92 0.84 4 40 7.5 8.03 0.77 5 40 10.0 7.83 0.97  下载: 导出CSV
下载: 导出CSV
表 2 活性染料的最大吸收波长和染色性能
Table 2. Maximum absorption wavelength and dyeing performances of reactive dyes
染料名称 最大吸收
波长/nm直接性/% 反应性/% 固色率/% M-3BE 542 7.76±1.05 18.58±2.01 24.96±2.46 SNE 499 8.23±1.28 23.56±3.06 40.29±1.98 LA 545 12.70±1.02 16.38±3.21 21.59±1.93
下载: 导出CSV
表 3 不同染料染色柞木单板的颜色
Table 3. Color change of X. japonicum veneers treated with different reactive red dyes
染料名称 染色后 染色前后差值 ∆E ${L_2^* }$ ${a_2^*} $ ${b_2^*} $ ∆L* ∆a* ∆b* M-3BE 47.48±3.25 32.50±4.70 5.50±2.53 − 23.29±4.09 25.59±5.10 − 15.32±2.17 38.03±5.61 SNE 46.31±1.77 34.78±3.49 24.21±1.55 − 24.15±2.71 27.41±3.95 2.72±1.53 36.68±4.68 LA 48.67±3.15 33.88±3.23 2.37±2.05 − 23.14±4.26 26.83±3.60 − 18.69±1.37 40.06±4.98
下载: 导出CSV
-
[1] 许茂松, 吕文华, 王雪玉. 人工林杨木增强-染色复合改性材的性能[J]. 林业科学, 2017, 53(1): 82 − 87. XU Maosong, LÜ Wenhua, WANG Xueyu. Properties of strengthening-dyeing-combined modified plantation poplar wood [J]. Sci Silv Sin, 2017, 53(1): 82 − 87. [2] WANG Xiaoqian, WANG Fang, YU Zhiming, et al. Surface free energy and dynamic wettability of wood simultaneously treated with acidic dye and flame retardant [J]. J Wood Sci, 2017, 63: 271 − 280. [3] 吴晓凤, 于志明, 张扬. 青杨生材纵向染色性能研究[J]. 北京林业大学学报, 2014, 36(3): 125 − 129. WU Xiaofeng, YU Zhiming, ZHANG Yang. Dyeing property of xylem utilizing the axil direction of freshly-felled Populous cathayana [J]. J Beijing For Univ, 2014, 36(3): 125 − 129. [4] WANG Xiaoqian. ZHANG Yang, YU Zhiming, et al. Properties of fast-growing poplar wood simultaneously treated with dye and flame retardant [J]. Eur J Wood Wood Prod, 2017, 75: 325 − 333. [5] NGUYEN N, OZARSKA B, FERGUSSON M, et al. Comparison of two dye uptake measurement methods for dyed wood veneer assessment [J]. Eur J Wood Wood Prod, 2018, 76: 1757 − 1759. [6] 张颖, 于志明, 张扬, 等. 载银壳聚糖的固色机理及其处理染色薄木性能的研究[J]. 北京林业大学学报, 2015, 37(6): 107 − 111. ZHANG Ying, YU Zhiming, ZHANG Yang, et al. Color fixation mechanism of CTS-Ag and its effect on the properties of dyed veneer [J]. J Beijing For Univ, 2015, 37(6): 107 − 111. [7] 张宏玉, 黄彦慧, 黄琼涛. 基于组子细工的珍贵树种余料再利用设计研究——以小径柚木为例[J]. 林产工业, 2020, 57(2): 37 − 40. ZHANG Hongyu, HUANG Yanhui, HUANG Qiongtao. Research on reuse design of the rare wood species residual material based on Kumiko: take small-diameter teak for example [J]. China For Prod Ind, 2020, 57(2): 37 − 40. [8] 胡极航, 范文苗, 李黎, 等. 北美糖槭单板染色工艺的优化[J]. 东北林业大学学报, 2016, 44(8): 68 − 72. HU Jihang, FAN Wenmiao, LI Li, et al. Optimal dyeing process for maple veneer [J]. J Northeast For Univ, 2016, 44(8): 68 − 72. [9] 李俊玲, 颜超, 余志成. 杨木活性染料染色性能及机理研究[J]. 浙江理工大学学报(自然科学版), 2010, 27(4): 535 − 539. LI Junling, YAN Chao, YU Zhicheng. Study on dyeing performance and mechanism of poplar with reactive dyes [J]. J Zhejiang Sci-Tech Univ Nat Sci Ed, 2010, 27(4): 535 − 539. [10] 刘强强, 吕文华, 石媛, 等. 木材染色研究现状及功能化展望[J]. 中国人造板, 2019, 26(9): 1 − 5. LIU Qiangqiang, LÜ Wenhua, SHI Yuan, et al. Advances in wood dyeing and its functionalizing prospects [J]. China Wood-Based Panels, 2019, 26(9): 1 − 5. [11] 赵泰, 郭明辉, 曹茜, 等. 活性染料对杨木单板染色耐光性影响的研究[J]. 林业机械与木工设备, 2011, 39(10): 29 − 31. ZHAO Tai, GUO Minghui, CAO Xi, et al. Study on the influence of reactive dyes on dyeing light fastness of poplar veneer [J]. For Mach Woodwork Equip, 2011, 39(10): 29 − 31. [12] 郭明辉, 曹龙. 活性染料对杨木单板染色后胶合性能的影响[J]. 木材工业, 2009, 23(1): 18 − 20. GUO Minghui, CAO Long. Effect of reactive dye on bonding performance of dyed poplar veneers [J]. China Wood Ind, 2009, 23(1): 18 − 20. [13] 李任, 黄荣凤, 高志强, 等. 预热温度对毛白杨压缩木材颜色及其回弹的影响[J]. 林产工业, 2019, 46(5): 17 − 21. LI Ren, HUANG Rongfeng, GAO Zhiqiang, et al. Effects of preheating temperatures on color and spring-back of Chinese white poplar compressed solid wood [J]. China For Prod Ind, 2019, 46(5): 17 − 21. [14] 喻胜飞, 刘元, 李贤军, 等. 活性红染料结构与杨木单板染色性能的关系[J]. 中南林业科技大学学报, 2015, 35(4): 114 − 118. YU Shengfei, LIU Yuan, LI Xianjun, et al. Relation on reactive red dyes structure and poplar veneer dyeing properties [J]. J Cent South For Univ, 2015, 35(4): 114 − 118. [15] 喻胜飞, 刘元, 李贤军, 等. 活性蓝染料结构与杨木单板染色性能的关系[J]. 中南林业科技大学学报, 2015, 35(2): 96 − 108. YU Shengfei, LIU Yuan, LI Xianjun, et al. Relation among different structure of reactive blue dyes and poplar dyeing properties [J]. J Cent South For Univ, 2015, 35(2): 96 − 108. [16] 赵璐, 薛静, 贾文彤, 等. 竹材的氟硼酸钠/DDAC改性与性能表征[J]. 林产工业, 2019, 56(12): 24 − 29, 51. ZHAO Lu, XUE Jing, JIA Wentong, et al. Properties characterization of bamboo modified with sodium fluoroborate/DDAC [J]. China For Prod Ind, 2019, 56(12): 24 − 29, 51. [17] 宋心远. 活性染料近代染色技术及助剂[J]. 印染助剂, 2008, 25(1): 1 − 8. SONG Xinyuan. Modern dyeing technologies and auxiliaries for reactive dyes [J]. Text Aux, 2008, 25(1): 1 − 8. [18] 刘影. 活性染料无盐低盐染色研究进展[J]. 印染助剂, 2014, 31(7): 8 − 11. LIU Ying. Research development of salt-free and low-salt dyeing for reactive dyes [J]. Text Aux, 2014, 31(7): 8 − 11. [19] 房宽峻, 刘曰兴, 舒大武, 等. 活性染料电中性无盐染色理论与应用[J]. 染整技术, 2017, 39(12): 50 − 54. FANG Kuanjun, LIU Yuexing, SHU Dawu, et al. Theory and application of reactive dye neutral salt free dyeing [J]. Text Dyeing Finish J, 2017, 39(12): 50 − 54. [20] 张鹏, 顾旦悦, 顾栋方, 等. 低碱型活性染料染色工艺[J]. 印染, 2013(17): 21 − 24. ZHANG Peng, GU Danyue, GU Dongfang, et al. Dyeing process with low alkali reactive dyes [J]. Textile, 2013(17): 21 − 24. [21] 彭志忠. 新型低碱活性染料染色实践研究[J]. 印整技术, 2015, 37(5): 11 − 14. PENG Zhizhong. Practice research of dyeing with novel low-alkaline reactive dyes [J]. Text Dyeing Finish J, 2015, 37(5): 11 − 14. [22] 宋心远. 活性染料高固色、高提升和高牢度染色技术[J]. 印染助剂, 2010, 27(3): 1 − 7. SONG Xinyuan. The high fixation, building up and fastness dyeing technology of reactive dyes [J]. Text Aux, 2010, 27(3): 1 − 7. [23] 宋心远. 活性染料低盐和无盐染色[J]. 印染助剂, 2006, 23(12): 1 − 3, 8. SONG Xinyuan. Low-salt and salt-free dyeing with reactive dyes [J]. Text Aux, 2006, 23(12): 1 − 3, 8. [24] 齐祥旭. 棉织物活性染料催化染色工艺研究[D]. 上海: 东华大学, 2016. QI Xiangxu. Catalytic Dyeing Technology of Reactive Dyes on Cotton Fabric[D]. Shanghai: Donghua University, 2016. [25] 顾园园. 双活性基蓝色谱活性染料结构设计及应用研究[D]. 上海: 东华大学, 2014. GU Yuanyuan. Synthesis and Application of Blue Reactive Dyes Containing Bifunctional Groups [D]. Shanghai: Donghua University, 2014. [26] 黄子宸, 侯婷婷, 叶凯, 等. 含羧基和磺酸基一氯均三嗪衍生物的合成及表征[J]. 广州化工, 2017, 45(5): 57 − 58. HUANG Zichen, HOU Tingting, YE Kai, et al. Synthesis and characterization of monochloro-s-triazine derivative containing carboxyl and sulfonate groups [J]. Guangzhou Chem Ind, 2017, 45(5): 57 − 58. [27] 李红, 于志明. 染料与木材结合机理的研究[J]. 北京林业大学学报, 2005, 27(4): 78 − 81. LI Hong, YU Zhiming. Combinative mechanism between dyestuff and wood [J]. J Beijing For Univ, 2005, 27(4): 78 − 81. [28] 苗平, 薛伟, 徐柏森. 蒸汽爆破预处理对柞木微观结构的影响[J]. 林业科技开发, 2007, 21(4): 51 − 53. MIAO Ping, XUE Wei, XU Bosen. Effects of steam explosion treatment on the microstructures of Mongolian oak Lumber [J]. J For Eng, 2007, 21(4): 51 − 53. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200391
 点击查看大图
点击查看大图

计量
- 文章访问数: 2099
- HTML全文浏览量: 405
- PDF下载量: 21
- 被引次数: 0












