





-
在当前中国水土流失严重、生态平衡危机频发、栽培药材品质备受质疑的大背景下,尤其是2020年初新冠病毒的突然来袭,金银花Lonicera japonica、内蒙黄芪Astragalus membranaceus var. mongholicus等传统中药材的品质和产量需求大增。发展药用植物的生态种植在合理利用土地资源、防治病虫害、改良土壤肥力、增强作物对自然灾害的抗逆能力等方面都有重要的作用。传统药用植物多为单一栽培模式,土地生产力和使用率低,对产业持续性发展不利[1]。相比而言,混播栽培是提高作物产量的有效措施。研究表明[2-3]:混播有助于防止水土流失,有利于病虫害治理和增强作物对自然灾害的抵抗力,在调节土壤生物养分有效性及生态系统资源合理配置等方面也至关重要。目前,混播多在豆科Leguminosae植物(如紫花苜蓿Medicago sativa、豌豆Pisum sativum等)-禾本科Gramineae植物(如燕麦Avena sativa、小麦Triticum aestivum等)间采用[4-5]。豆科植物根系的固氮能力不仅可以维持自身的氮素营养需求,还可通过根系间的相互作用为与其混播的作物提供养分,满足后者生长过程中的生理需求。近年来药用植物的研究主要聚焦在土壤施用肥料数量和不同质地土壤对药用植物生产量的影响[6-9]上,对不同播种模式下,土壤中有机碳、氮质量分数及土壤酶活性的研究较少;少有豆科植物与与药用植物的混播研究。本研究在不施氮磷钾肥的情况下,以忍冬科Caprifoliaceae金银花、菊科Compositae杭白菊Chrysanthemum morifolium和豆科内蒙黄芪为研究对象,采用大田实验随机区组设计,以沙化裸地(ck)为对照,研究金银花单播(LJT)、杭白菊单播(CMR)、黄芪单播(AMB)、金银花/内蒙黄芪混播(LJT/AMB)、杭白菊/内蒙黄芪混播(CMR/AMB) 5种模式下土壤碳、氮质量分数和酶活性的变化,旨在为人工药用草地建植及土壤治理保护提供科学依据。
-
研究区位于河南省新乡市洪门镇(35°16′N,113°57′E),海拔120 m,大陆性季风气候,四季分明,昼夜温差大,年平均气温为14.2 ℃,全年平均日光辐射时数约2 400 h,年平均降水量为573.4 mm,年际间降水分布不均,主要集中在7、8月,年平均相对湿度为68.0%,无霜期220 d[10]。供试土壤为砂壤土,供试土壤0~40 cm土层基本养分状况见表1。
表 1 供试土壤(0~40 cm)养分特性
Table 1. Soil nutrient characteristics (0−40 cm)
土层/cm 全氮/(g·kg−1) 全磷/(g·kg−1) 全钾/(g·kg−1) 碱解氮/(mg·kg−1) 速效磷/(mg·kg−1) 速效钾/(mg·kg−1) 有机碳/(g·kg−1) pH 0~20 1.28 0.77 13.13 73.75 15.39 107.69 9.44 7.87 20~40 0.69 0.59 11.25 42.68 10.60 66.28 6.10 7.83 -
2016−2019年连续4 a进行随机区组试验。共设6个处理,即:以沙化裸地为对照(ck),设LJT、CMR、AMB、LJT/AMB、CMR/AMB等5种不同的播种模式,各处理重复3次,共18个小区,各小区面积为30 m2 (5 m×6 m)。杭白菊、金银花均为南北行种植,株距60 cm,行距60 cm。内蒙黄芪为条播,行距为33 cm,播种量为15 kg·hm−2,混播模式下金银花与内蒙黄芪的行间距为40 cm,杭白菊与内蒙黄芪的行间距为40 cm。不施用任何肥料,采用免耕处理,定期浇水及除草。
-
2019年10月,清除土壤表面的枯枝落叶及地表碎石,五点法取样采集土壤样品装入自封袋带回实验室。自然风干后研磨,过筛,检测土壤各指标。采用有机碳分析仪(Elementar,德国)测定土壤有机碳质量分数(g·kg−1),采用碱解扩散法测定土壤碱解氮质量分数(g·kg−1)[11]。采用常规测定方法检测分析土壤酶活性[12]。采用靛酚蓝比色法测定脲酶活性,以24 h后1.0 g土样中铵态氮质量分数(mg·g−1)表示;采用3,5-二硝基水杨酸比色法测定蔗糖酶活性,以24 h后1.0 g土样中葡萄糖质量分数(mg·g−1)表示;采用磷酸苯二钠比色法测定碱性磷酸酶活性,以24 h后1.0 g土样中释放的酚的质量分数(mg·g−1)表示;采用邻苯二酚比色法测定多酚氧化酶活性,以2 h后1.0 g土样中紫色没食子素的质量(mg)表示。
-
数据以3次平行测定的平均值±标准误表示,经Excel 2007和SPSS 19.0软件统计分析。采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)进行分析,Pearson相关系数法进行相关性分析。
-
由图1可知:相比于ck,5种播种模式下不同土层土壤有机碳质量分数均显著增加(P<0.05),且同一土壤深度土壤有机碳质量分数从大到小均依次为CMR/AMB、LJT/AMB、AMB、CMR、LJT、ck。与3种单播模式相比,2种混播模式下土壤有机碳质量分数均显著增加(P<0.05),其中CMR/AMB模式有机碳质量分数略高于LJT/AMB(P>0.05)。LJT和CMR模式下土壤表层(0~20 cm)有机碳质量分数较ck显著增加(P<0.05),而在土壤亚表层(20~40 cm)无显著性差异(P>0.05)。此外,不同种植模式下土壤有机碳质量分数均呈表聚性现象,即土壤有机碳质量分数随着土壤垂直剖面加深而逐渐降低。
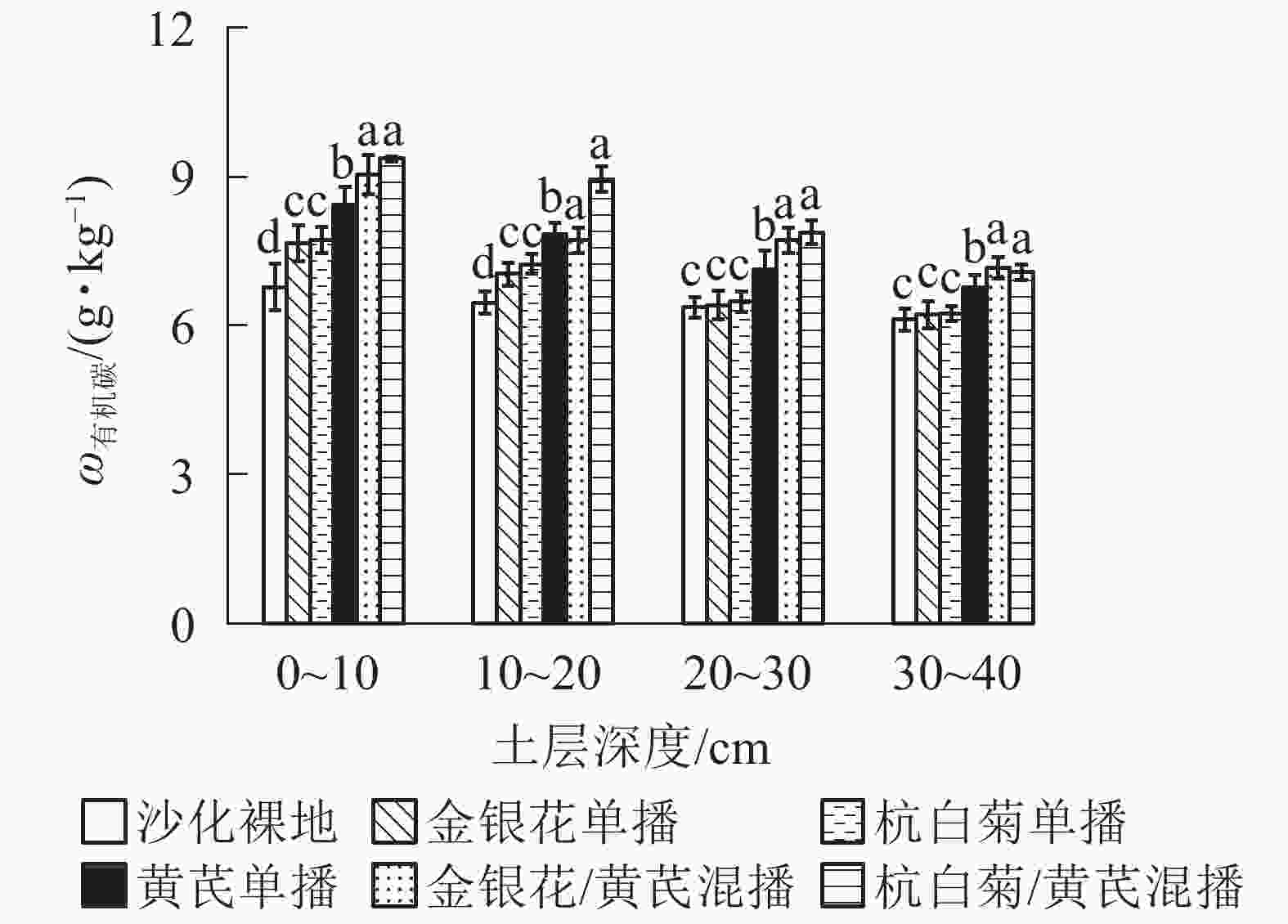
图 1 不同播种模式下土壤各土层有机碳质量分数
Figure 1. Soil organic carbon content under different planting patterns
-
由图2可知:AMB、CMR/AMB和LJT/AMB播种模式下土壤碱解氮质量分数最高,分别为63.89,62.87和61.66 mg·kg−1,3种播种模式间无显著差异(P>0.05);但显著高于LJT和CMR模式(P<0.05)。在土壤表层(0~20 cm)5种播种模式下碱解氮质量分数均显著高于ck (P<0.05),但在土壤亚表层(20~40 cm)无显著差异(P>0.05)。说明不同播种模式下土壤碱解氮质量分数也呈现表聚性现象。
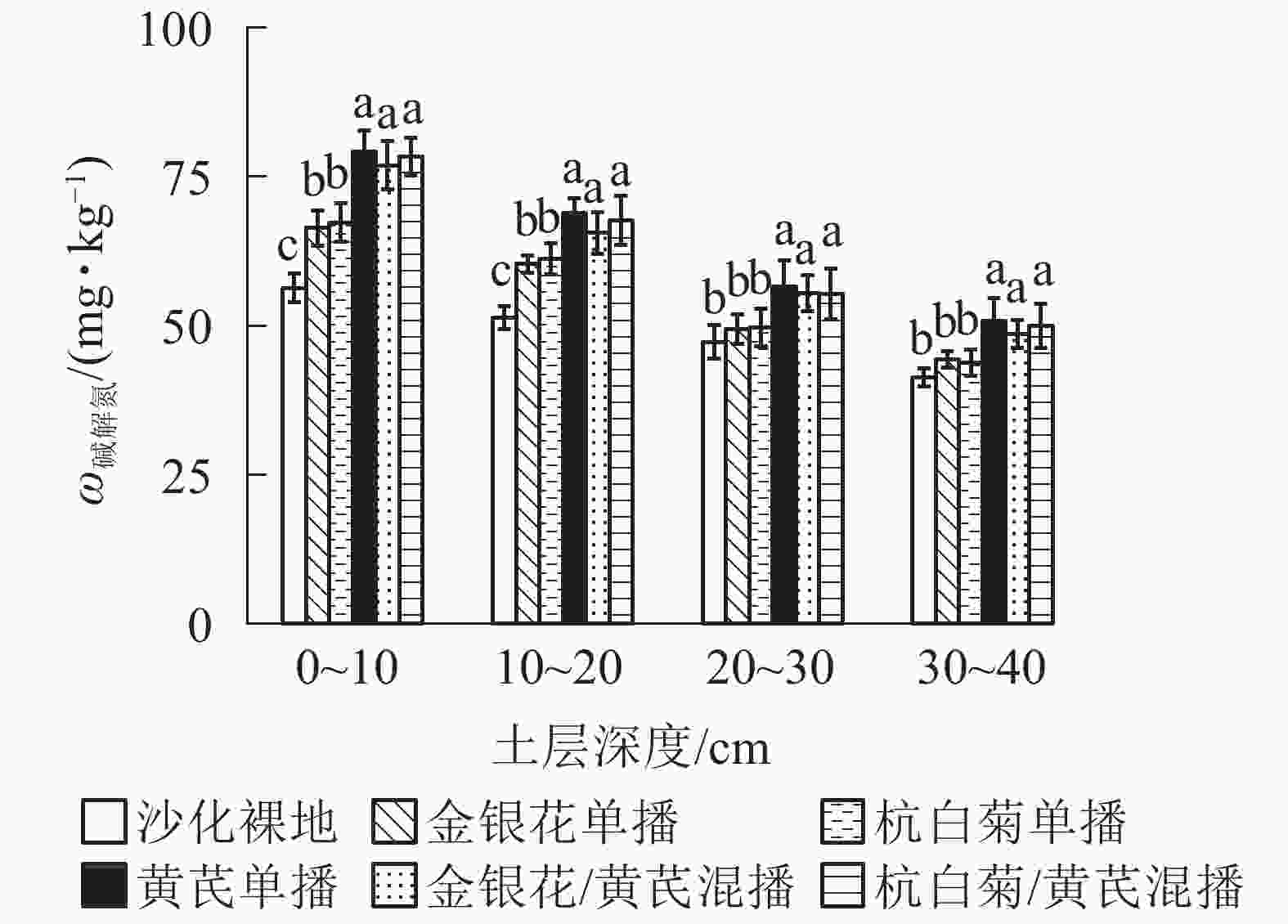
图 2 不同播种模式下土壤各土层碱解氮质量分数
Figure 2. Soil available nitrogen content under different planting patterns
-
与ck相比(表2),5种播种模式下4种土壤酶(脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶)活性均显著增加(P<0.05)。20~40 cm土层,单播和混播模式之间蔗糖酶和碱性磷酸酶活性差异显著(P<0.05),5种播种模式下土壤脲酶活性差异显著(P<0.05),土壤氧化酶活性差异不显著(P>0.05)。从土壤酶活性的各土层平均值来看,各个播种模式下土壤脲酶活性较ck的增幅最大。LJT/AMB的脲酶活性(0.789 9 mg·g−1)和多酚氧化酶活性(1.077 6 mg·g−1)最高,CMR/AMB下的蔗糖酶活性(8.289 6 mg·g−1)和碱性磷酸酶活性(0.644 8 mg·g−1)最高。LJT/AMB下脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶活性对比ck分别增加201.82%、56.95%、77.05%和70.85%,CMR/AMB下脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶活性对比ck分别增加192.63%、58.37%、78.75%和59.36%,AMB模式下脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶活性对比ck分别增加140.62%、44.57%、52.86%和57.90%,CMR模式下脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶活性对比ck分别增加106.74%、38.42%、49.06%和55.25%,LJT模式下脲酶、蔗糖酶、碱性磷酸酶和多酚氧化酶活性对比ck分别增加67.69%、33.80%、23.63%和55.81%。此外,从土壤的垂直分布来看,不同播种模式下土壤酶活性也呈表聚性特征。
表 2 不同播种模式下各土层土壤酶活性
Table 2. Soil enzyme activities in each soil layer under different planting patterns
土层/cm 种植方式 脲酶/(mg·g−1) 蔗糖酶/(mg·g−1) 碱性磷酸酶/(mg·g−1) 多酚氧化酶/(mg·g−1) 0~10 ck 0.342 5±0.021 8 d 6.358 2±0.265 0 d 0.417 6±0.020 6 d 0.754 8±0.046 9 b LJT 0.624 2±0.056 4 c 7.947 8±0.645 6 c 0.617 6±0.043 2 c 1.116 4±0.074 6 a CMR 0.606 7±0.046 5 c 7.847 8±0.241 8 c 0.655 6±0.034 5 c 1.103 1±0.072 6 a AMB 0.844 7±0.062 6 b 8.720 9±0.468 9 b 0.732 5±0.052 6 b 1.127 3±0.089 5 a LJT/AMB 1.108 3±0.061 3 a 9.466 5±0.532 5 a 0.843 2±0.067 8 a 1.275 3±0.084 2 a CMR/AMB 1.036 7±0.055 8 a 9.217 4±0.204 1 a 0.800 4±0.057 2 a 1.134 3±0.067 0 a 10~20 ck 0.310 3±0.018 7 d 5.575 0±0.344 7 d 0.387 5±0.025 3 d 0.697 9±0.037 0 b LJT 0.473 0±0.057 5 c 7.443 8±0.245 9 c 0.490 3±0.053 7 c 1.075 2±0.063 5 a CMR 0.578 1±0.032 1 c 7.143 8±0.443 0 c 0.587 9±0.047 6 c 1.052 2±0.068 7 a AMB 0.639 3±0.052 4 b 8.039 8±0.352 6 b 0.585 8±0.036 2 b 1.068 2±0.070 1 a LJT/AMB 0.843 2±0.064 7 a 8.732 1±0.231 9 a 0.665 4±0.054 7 a 1.182 6±0.096 4 a CMR/AMB 0.798 2±0.045 1 a 8.798 4±0.234 7 a 0.696 3±0.047 3 a 1.093 6±0.074 3 a 20~30 ck 0.222 5±0.015 3 d 4.754 6±0.125 4 c 0.332 7±0.022 3 c 0.563 7±0.026 4 b LJT 0.357 2±0.043 6 c 6.764 5±0.538 6 b 0.355 0±0.023 1 b 0.931 6±0.065 3 a CMR 0.489 7±0.030 7 c 6.994 5±0.364 8 b 0.453 6±0.036 1 b 0.880 7±0.057 0 a AMB 0.553 6±0.038 7 b 7.184 6±0.342 1 b 0.472 2±0.032 7 b 0.952 5±0.085 4 a LJT/AMB 0.667 1±0.059 8 a 7.648 7±0.452 3 a 0.551 2±0.166 0 a 0.994 3±0.073 2 a CMR/AMB 0.663 3±0.043 7 a 7.957 2±0.102 5 a 0.575 4±0.048 9 a 0.926 6±0.088 5 a 30~40 ck 0.171 6±0.013 4 d 4.248 9±0.247 5 c 0.304 9±0.019 7 c 0.506 4±0.022 7 b LJT 0.301 0±0.016 7 c 5.856 4±0.448 7 b 0.320 8±0.003 8 b 0.807 5±0.042 0 a CMR 0.489 7±0.030 7 c 6.994 5±0.364 8 b 0.453 6±0.036 1 b 0.8807±0.057 0 a AMB 0.481 2±0.034 2 b 6.323 1±0.476 8 b 0.415 0±0.025 4 b 0.835 4±0.067 3 a LJT/AMB 0.540 9±0.043 4 a 7.013 2±0.189 7 a 0.494 7±0.113 7 a 0.858 0±0.067 6 a CMR/AMB 0.565 0±0.038 1 a 7.185 4±0.114 6 a 0.506 9±0.036 1 a 0.865 9±0.064 0 a 说明:不同字母表示同一土层不同播种模式间差异显著(P<0.05) -
由Pearson相关系数(表3)可知:土壤有机碳与碱解氮、4种酶呈极显著相关(P<0.01),土壤碱解氮与4种酶呈极显著相关(P<0.01)。4种土壤酶彼此之间呈极显著相关(P<0.01)。其中脲酶与碱性磷酸酶之间相关系数达0.965,有机碳与碱性磷酸酶之间相关系数达0.949,蔗糖酶与多酚氧化酶之间相关系数达0.948,说明土壤碳、氮与酶之间关系极为紧密。
表 3 土壤碳、氮质量分数与酶活性的相关性分析
Table 3. Correlations analysis of soil carbon, available nitrogen and enzyme activities
参数 有机碳 碱解氮 脲酶 蔗糖酶 碱性磷酸酶 多酚氧化酶 有机碳 1 0.893** 0.943** 0.897** 0.949** 0.808** 碱解氮 1 0.848** 0.844** 0.917** 0.835** 脲酶 1 0.949** 0.965** 0.871** 蔗糖酶 1 0.931** 0.948** 碱性磷酸酶 1 0.875** 多酚氧化酶 1 说明:**表示在0.01水平(双侧)上显著相关 -
与单播相比,混播不仅可以增加土壤碳、氮总量,更利于土壤有机碳库的稳定[13-14],还能增加土壤速效养分含量,提高土壤养分的有效性。本研究发现:LJT/AMB模式下,土壤有机碳和碱解氮质量分数(0~40 cm土层平均值)较LJT分别提高了19.41%和11.80%,CMR/AMB模式下,土壤有机碳和碱解氮质量分数较CMR分别提高了20.16%和13.24%,这与包兴国等[15]、郑伟等[16]发现混播更有助于改善土壤有机碳、碱解氮的结论一致。作为生物固氮植物,混播模式下内蒙黄芪在向金银花/杭白菊转移土壤氮素的同时,也能通过氮素根际沉积为金银花/杭白菊提供一定的氮素,减缓土壤氮素消耗,土壤速效氮质量分数明显高于LJT和CMR。土壤脲酶通过水解土壤中的尿素形成铵态氮提供氮源,其活性直接表征氮素转化率[17-19];土壤蔗糖酶通过水解土壤有机质生成葡萄糖提供碳源,表征有机碳的积累转化和土壤熟化程度[20-21];土壤碱性磷酸酶通过加速有机磷的脱磷速度直接参与土壤磷素循环,表征磷元素生物转化方向和强弱[22];土壤多酚氧化酶参与土壤芳香族化合物循环,通过分子氧氧化酚或多酚形成醌,醌与土壤中蛋白质、糖、矿质元素等反应生成有机质,分解凋落物木质素获取碳、氮元素[23-24]。蔺芳[18]发现:土壤脲酶、蔗糖酶对土壤碳循环有直接贡献,脲酶、蔗糖酶和碱性磷酸酶对土壤氮循环有直接贡献,与本研究一致,说明土壤中不同种类酶之间、酶和土壤养分之间具有协同交互作用[25]。本研究混播模式下的4种土壤酶活性均显著均高于3种单播和ck (P<0.05),且相关性分析表明:土壤有机碳和碱解氮均与4种酶活性极显著相关(P<0.01)。说明在混播体系中,豆科植物的生物固氮作用及种间竞争作用促进了植物根系生长,大大增加根系种类、生物量和根系纵横分布,增加土壤孔隙度,提高蓄水保水能力,从而改变土壤理化性质。豆科植物根系周围的根瘤菌能促进根系分泌物、细根周转,改善菌根代谢,使得土壤微生物代谢旺盛,呼吸强度增强,促进蔗糖酶、脲酶活性,提高碳素、氮素转化效率,促进碱性磷酸酶和多酚氧化酶活性,对提高土壤肥力有重要作用[26-27]。不同种植模式下土壤有机碳、碱解氮和酶活性均在土壤剖面呈表聚性,表明土壤表层更有利于酶促反应形成有机质和有效氮。与TU等[25]、来幸樑等[28]结果一致。土壤表层凋落物、腐殖质容易积累,微生物代谢旺盛,植物根系集中,有利于物质积累和酶促反应的进行;随着土壤深度的加深,土壤中的水、汽、热状况变差,微生物生境变差,数量急剧减少,酶促反应底物也不如表层,碳氮积累和酶活均随着土层剖面下降。
混播模式充分利用豆科植物内蒙黄芪的生物固氮作用,使金银花/杭白菊在没有氮肥的条件下获取氮源,提高植物氮素吸收利用效率,减少土壤无机氮的累积,降低种植地氮素污染的风险。杭白菊/黄芪混播和金银花/黄芪混播模式下土壤有机碳、碱解氮质量分数、土壤脲酶、蔗糖酶和碱性磷酸酶活性均显著高于杭白菊或金银花单播(P<0.05),而彼此之间差异不显著(P>0.05)。相较于单播,2种混播模式对土壤有机碳、碱解氮的积累和土壤酶活性的促进作用更强,更有利于改良土壤肥力,是适合在豫北地区推广的可持续发展的生态农业模式。
Effects of single/mixed sowing of three medicinal plants on the contents of carbon, nitrogen and enzymes activities of sandy soil
-
摘要:
目的 探究3种药用植物单/混播对土壤品质的影响。 方法 以金银花Lonicera japonica (LJT)、杭白菊Chrysanthemum morifolium (CMR)、内蒙黄芪Astragalus membranaceus (AMB)为材料,以沙化裸地为对照(ck),在豫北地区连续4 a进行定位试验。分析单/混播种植模式对不同土层(0~40 cm)中有机碳、碱解氮和土壤酶活性的影响,并分析其相关性。 结果 杭白菊/内蒙黄芪(CMR/AMB)和金银花/内蒙黄芪(LJT/AMB)混播模式下土壤有机碳、碱解氮质量分数、土壤脲酶、蔗糖酶和碱性磷酸酶活性均显著高于杭白菊(CMR)和金银花(LJT)单播模式(P<0.05)。CMR/AMB模式下土壤有机碳质量分数较ck、CMR和AMB分别提高了29.32%、20.16%和10.25%,碱解氮质量分数较ck和CMR分别提高了28.02%和13.24%;LJT/AMB模式下土壤有机碳质量分数较ck、LJT和AMB分别提高了25.46%、18.09%和6.96%,碱解氮质量分数较ck和LJT分别提高了25.56%和11.80%。土壤有机碳、碱解氮和4种土壤酶彼此之间均呈极显著相关(P<0.01)。从土壤剖面看,不同种植模式下土壤有机碳、碱解氮含量及4种酶活性均呈表聚性特征。 结论 LJT/AMB和CMR/AMB混播模式能通过内蒙黄芪的生物固氮作用提高土壤养分有效性,是适合在当地推广的可持续发展的生态种植模式。图2表3参28 Abstract:Objective This study aims to explore the effects of single and mixed sowing of three medicinal plants on soil quality. Method Taking Lonicera japonica (LJT), Chrysanthemum morifolium (CMR) and Astragalus membranaceus var. mongholicus (AMB) as experimental materials and sandy bare land as control(ck), the effects of single and mixed sowing patterns on organic carbon, alkaline hydrolyzable nitrogen and soil enzyme activity in different soil layers (0−40 cm) were studied in the location test conducted in northern Henan Province for 4 consecutive years, and their correlation was analyzed. Result The content of soil organic carbon, alkali hydrolyzable nitrogen and soil enzyme activity of urease, sucrase and alkaline phosphatase under the mixed sowing patterns of CMR/AMB and LJT/AMB were significantly higher than those under the single patten of CMR or LJT (P<0.05). Soil organic carbon content in CMR/AMB pattern increased by 29.32%, 20.16% and 10.25% respectively, compared with ck, CMR and AMB. Alkaline hydrolyzable nitrogen content increased by 28.02% and 13.24% respectively, compared with ck and CMR. Soil organic carbon content of LJT/AMB pattern increased by 25.46%, 18.09% and 6.96%, respectively, compared with ck, LJT and AMB. Alkaline hydrolyzable nitrogen content increased by 25.56% and 11.80% respectively, compared with ck and LJT. Soil organic carbon, alkali hydrolyzable nitrogen, and four soil enzymes were all significantly correlated with each other (P<0.01). From the view of soil profile, the soil organic carbon, alkali hydrolyzable nitrogen and the activities of 4 species of soil enzymes showed the characteristics of surface aggregation under different planting patterns. Conclusion CMR/AMB and LJT/AMB mixed patterns can improve the effectiveness of soil nutrients through the biological nitrogen fixation of A. membranaceus var. mongholicus. They are sustainable ecological patterns suitable for local promotion. [Ch, 2 fig. 3 tab. 28 ref.] -
图 1 不同播种模式下土壤各土层有机碳质量分数
不同字母表示同一土层不同播种模式间差异显著(P<0.05)
Figure 1 Soil organic carbon content under different planting patterns
图 2 不同播种模式下土壤各土层碱解氮质量分数
不同字母表示同一土层不同播种模式间差异显著(P<0.05)
Figure 2 Soil available nitrogen content under different planting patterns
表 1 供试土壤(0~40 cm)养分特性
Table 1. Soil nutrient characteristics (0−40 cm)
土层/cm 全氮/(g·kg−1) 全磷/(g·kg−1) 全钾/(g·kg−1) 碱解氮/(mg·kg−1) 速效磷/(mg·kg−1) 速效钾/(mg·kg−1) 有机碳/(g·kg−1) pH 0~20 1.28 0.77 13.13 73.75 15.39 107.69 9.44 7.87 20~40 0.69 0.59 11.25 42.68 10.60 66.28 6.10 7.83  下载: 导出CSV
下载: 导出CSV
表 2 不同播种模式下各土层土壤酶活性
Table 2. Soil enzyme activities in each soil layer under different planting patterns
土层/cm 种植方式 脲酶/(mg·g−1) 蔗糖酶/(mg·g−1) 碱性磷酸酶/(mg·g−1) 多酚氧化酶/(mg·g−1) 0~10 ck 0.342 5±0.021 8 d 6.358 2±0.265 0 d 0.417 6±0.020 6 d 0.754 8±0.046 9 b LJT 0.624 2±0.056 4 c 7.947 8±0.645 6 c 0.617 6±0.043 2 c 1.116 4±0.074 6 a CMR 0.606 7±0.046 5 c 7.847 8±0.241 8 c 0.655 6±0.034 5 c 1.103 1±0.072 6 a AMB 0.844 7±0.062 6 b 8.720 9±0.468 9 b 0.732 5±0.052 6 b 1.127 3±0.089 5 a LJT/AMB 1.108 3±0.061 3 a 9.466 5±0.532 5 a 0.843 2±0.067 8 a 1.275 3±0.084 2 a CMR/AMB 1.036 7±0.055 8 a 9.217 4±0.204 1 a 0.800 4±0.057 2 a 1.134 3±0.067 0 a 10~20 ck 0.310 3±0.018 7 d 5.575 0±0.344 7 d 0.387 5±0.025 3 d 0.697 9±0.037 0 b LJT 0.473 0±0.057 5 c 7.443 8±0.245 9 c 0.490 3±0.053 7 c 1.075 2±0.063 5 a CMR 0.578 1±0.032 1 c 7.143 8±0.443 0 c 0.587 9±0.047 6 c 1.052 2±0.068 7 a AMB 0.639 3±0.052 4 b 8.039 8±0.352 6 b 0.585 8±0.036 2 b 1.068 2±0.070 1 a LJT/AMB 0.843 2±0.064 7 a 8.732 1±0.231 9 a 0.665 4±0.054 7 a 1.182 6±0.096 4 a CMR/AMB 0.798 2±0.045 1 a 8.798 4±0.234 7 a 0.696 3±0.047 3 a 1.093 6±0.074 3 a 20~30 ck 0.222 5±0.015 3 d 4.754 6±0.125 4 c 0.332 7±0.022 3 c 0.563 7±0.026 4 b LJT 0.357 2±0.043 6 c 6.764 5±0.538 6 b 0.355 0±0.023 1 b 0.931 6±0.065 3 a CMR 0.489 7±0.030 7 c 6.994 5±0.364 8 b 0.453 6±0.036 1 b 0.880 7±0.057 0 a AMB 0.553 6±0.038 7 b 7.184 6±0.342 1 b 0.472 2±0.032 7 b 0.952 5±0.085 4 a LJT/AMB 0.667 1±0.059 8 a 7.648 7±0.452 3 a 0.551 2±0.166 0 a 0.994 3±0.073 2 a CMR/AMB 0.663 3±0.043 7 a 7.957 2±0.102 5 a 0.575 4±0.048 9 a 0.926 6±0.088 5 a 30~40 ck 0.171 6±0.013 4 d 4.248 9±0.247 5 c 0.304 9±0.019 7 c 0.506 4±0.022 7 b LJT 0.301 0±0.016 7 c 5.856 4±0.448 7 b 0.320 8±0.003 8 b 0.807 5±0.042 0 a CMR 0.489 7±0.030 7 c 6.994 5±0.364 8 b 0.453 6±0.036 1 b 0.8807±0.057 0 a AMB 0.481 2±0.034 2 b 6.323 1±0.476 8 b 0.415 0±0.025 4 b 0.835 4±0.067 3 a LJT/AMB 0.540 9±0.043 4 a 7.013 2±0.189 7 a 0.494 7±0.113 7 a 0.858 0±0.067 6 a CMR/AMB 0.565 0±0.038 1 a 7.185 4±0.114 6 a 0.506 9±0.036 1 a 0.865 9±0.064 0 a 说明:不同字母表示同一土层不同播种模式间差异显著(P<0.05)
下载: 导出CSV
表 3 土壤碳、氮质量分数与酶活性的相关性分析
Table 3. Correlations analysis of soil carbon, available nitrogen and enzyme activities
参数 有机碳 碱解氮 脲酶 蔗糖酶 碱性磷酸酶 多酚氧化酶 有机碳 1 0.893** 0.943** 0.897** 0.949** 0.808** 碱解氮 1 0.848** 0.844** 0.917** 0.835** 脲酶 1 0.949** 0.965** 0.871** 蔗糖酶 1 0.931** 0.948** 碱性磷酸酶 1 0.875** 多酚氧化酶 1 说明:**表示在0.01水平(双侧)上显著相关
下载: 导出CSV
-
[1] 郭秀芝, 彭政, 王铁霖, 等. 间套作体系下种间互作对药用植物影响的研究进展[J]. 中国中药杂志, 2020, 45(9): 2017 − 2022. GUO Xiuzhi, PENG Zheng, WANG Tielin, et al. Research progress in effects of interspecific interaction on medicinal plants in intercropping system [J]. China J Chin Mater Medica, 2020, 45(9): 2017 − 2022. [2] 谢开云, 李向林, 何峰, 等. 单播与混播下紫花苜蓿与无芒雀麦生物量对氮肥的响应[J]. 草业学报, 2014, 23(6): 148 − 156. XIE Kaiyun, LI Xianglin, HE Feng, et al. Response of alfalfa and smooth brome to nitrogen fertilizer in monoculture and mixed grasslands [J]. Acta Prataculturae Sin, 2014, 23(6): 148 − 156. [3] 欧阳铖人, 吴伯志. 水土保持耕作措施的研究进展及展望[J]. 云南农业大学学报(自然科学), 2017, 32(4): 718 − 726. OUYANG Chengren, WU Bozhi. A review of soil conservation practices on uplands [J]. J Yunnan Agric Univ Nat Sci, 2017, 32(4): 718 − 726. [4] 蔺芳, 刘晓静, 张家洋. 紫花苜蓿与多年生黑麦草不同种植模式下沙化土壤碳、氮含量和酶活性研究[J]. 草原与草坪, 2019, 39(3): 43 − 49. LIN Fang, LIU Xiaojing, ZHANG Jiayang. Study of the contents of carbon, nitrogen and enzymes activities of sandy soil grown alfalfa and perennial ryegrass with different planting patterns [J]. Grassland Turf, 2019, 39(3): 43 − 49. [5] 李昂, 李昌明, 尹卓忻, 等. 甘肃秦王川灌区种植豆禾混播牧草的农田生态保育效应[J]. 水土保持学报, 2020, 34(2): 239 − 244, 268. LI Ang, LI Changming, YIN Zhuoxin, et al. Ecological conservation effect of mixed planing legume and grass in Qinwangchuan irrigation area of Gansu Province [J]. J Soil Water Conserv, 2020, 34(2): 239 − 244, 268. [6] 哈文秀, 周金星, 庞丹波, 等. 岩溶区不同恢复方式下土壤有机碳组分及酶活性研究[J]. 北京林业大学学报, 2019, 41(2): 1 − 11. HA Wenxiu, ZHOU Jinxing, PANG Danbo, et al. Soil organic carbon fraction and enzyme activities under different restoration methods in karst area [J]. J Beijing For Univ, 2019, 41(2): 1 − 11. [7] 张家春, 刘婧, 林绍霞, 等. 不同种植方式下贵州玄参种植土壤碳氮磷特征[J]. 河南农业科学, 2015, 44(6): 64 − 67. ZHANG Jiachun, LIU Jing, LIN Shaoxia, et al. Features of carbon, nitrogen and phosphorus in Scrophulariaceae growing soil under different planting patterns in Guizhou [J]. J Henan Agric Sci, 2015, 44(6): 64 − 67. [8] 王文锋, 李春花, 黄绍文, 等. 不同施肥模式对设施秋冬茬芹菜生育期间土壤酶活性的影响[J]. 植物营养与肥料学报, 2016, 22(3): 676 − 686. WANG Wenfeng, LI Chunhua, HUANG Shaowen, et al. Effects of different fertilization patterns on soil enzyme activities during growing period of autumn-winter season celery in greenhouse [J]. J Plant Nutr Fert, 2016, 22(3): 676 − 686. [9] 陈立明, 丁雷, 满秀玲. 云冷杉林土壤酶活性与植物多样性[J]. 东北林业大学学报, 2009, 37(3): 58 − 61. CHEN Liming, DING Lei, MAN Xiuling. Soil enzyme activities and plant diversity in spruce-fir forest [J]. J Northeast For Univ, 2009, 37(3): 58 − 61. [10] 张家洋, 蔺芳, 詹乃才, 等. 紫花苜蓿与无芒雀麦不同栽培模式下土壤团聚体形态结构、组成及有机碳特征[J]. 浙江农林大学学报, 2019, 36(6): 1077 − 1086. ZHANG Jiayang, LIN Fang, ZHAN Naicai, et al. Morphological structure, composition, and organic carbon characteristics of soil agglomerations for alfalfa and ryegrass planting patterns [J]. J Zhejiang A&F Univ, 2019, 36(6): 1077 − 1086. [11] 于天一, 逄焕成, 唐海明, 等. 不同母质类型水稻土酶活性及其与理化性质的关系[J]. 土壤学报, 2013, 50(5): 1043 − 1047. YU Tianyi, PANG Huancheng, TANG Haiming, et al. Soil enzyme activities and their relationships with soil physicochemical properties in paddy soils derived from different parent materials under double-rice cropping system in South China [J]. Acta Pedol Sin, 2013, 50(5): 1043 − 1047. [12] 关松荫. 土壤酶及其研究方法[M]. 北京: 农业出版社, 1986: 260 − 339. [13] 关正翾, 娜尔克孜, 朱亚琼, 等. 不同混播方式下燕麦+箭筈豌豆混播草地的生产性能及土壤养分特征[J]. 草业科学, 2019, 36(3): 772 − 784. GUAN Zhengxuan, Naerkezi, ZHU Yaqiong, et al. Effect of different sowing patterns on production performance and soil nutrients in Avena sativa+Vicia sativa mixtures [J]. Pratacultural Sci, 2019, 36(3): 772 − 784. [14] 邰继承, 杨恒山, 范富, 等. 播种方式对紫花苜蓿+无芒雀麦草地土壤碳密度和组分的影响[J]. 草业科学, 2010, 27(6): 102 − 107. TAI Jicheng, YANG Hengshan, FAN Fu, et al. Effects of sowing methods on soil carbon density and composition in the alfalfa and Bromus inermis pasture [J]. Pratacultural Sci, 2010, 27(6): 102 − 107. [15] 包兴国, 杨文玉, 曹卫东, 等. 豆科与禾本科绿肥饲草作物混播增肥及改土效果研究[J]. 中国草地学报, 2012, 34(1): 43 − 47. BAO Xingguo, YANG Wenyu, CAO Weidong, et al. Soil fertility improvement by mixed planting of Leguminous and Gramineous green manure crops [J]. Chin J Grassland, 2012, 34(1): 43 − 47. [16] 郑伟, 加娜尔古丽, 唐高溶, 等. 混播种类与混播比例对豆禾混播草地浅层土壤养分的影响[J]. 草业科学, 2015, 32(3): 329 − 339. ZHENG Wei, Jianaerguli, TANG Gaorong, et al. Effects of mixed species, mixed ratios of legume to grass on soil nutrients in surface soils of legume-grass mixture pasture [J]. Pratacultural Sci, 2015, 32(3): 329 − 339. [17] 王理德, 王方琳, 郭春秀, 等. 土壤酶学硏究进展[J]. 土壤, 2016, 48(1): 12 − 21. WANG Lide, WANG Fanglin, GUO Chunxiu, et al. Review: progress of soil enzymology [J]. Soils, 2016, 48(1): 12 − 21. [18] 蔺芳. 不同栽培模式下豫北沙化土壤微生物量和酶活性[J]. 浙江农林大学学报, 2019, 36(3): 590 − 597. LIN Fang. Microbial biomass and enzymatic activity in sandy soils of northern Henan Province with different alfalfa and awnless brome cultivating patterns [J]. J Zhejiang A&F Univ, 2019, 36(3): 590 − 597. [19] LI Qiang, LIANG Jianhong, HE Y Y, et al. Effect of land use on soil enzyme activities at karst area in Nanchuan, Chongqing, Southwest China [J]. Plant Soil Environ, 2014, 60(1): 15 − 20. [20] 刘文辉, 张英俊, 师尚礼, 等. 高寒区施肥和豆科混播水平对燕麦人工草地土壤酶活性的影响[J]. 草业学报, 2017, 26(1): 23 − 33. LIU Wenhui, ZHANG Yingjun, SHI Shangli, et al. Soil enzyme activities in alpine naked oat-artificial grassland in reponse to fertilizer and legume mix levels [J]. Acta Prataculturae Sin, 2017, 26(1): 23 − 33. [21] 黄玙璠, 舒英格, 肖盛杨, 等. 喀斯特山区不同草地土壤养分与酶活性特征[J]. 草业学报, 2020, 29(6): 93 − 104. HUANG Yufan, SHU Yingge, XIAO Shengyang, et al. Quantification of soil nutrient levels and enzyme activities in different grassland categories in karst mountains [J]. Acta Prataculturae Sin, 2020, 29(6): 93 − 104. [22] 张丽琼, 郝明德, 臧逸飞, 等. 苜蓿和小麦长期连作对土壤酶活性及养分的影响[J]. 应用生态学报, 2014, 25(11): 3191 − 3196. ZHANG Liqiong, HAO Mingde, ZANG Yifei, et al. Effects of continuous cropping of wheat and alfalfa on soil enzyme activities and nutrients [J]. Chin J Appl Ecol, 2014, 25(11): 3191 − 3196. [23] 郝建朝, 吴沿友, 连宾, 等. 土壤多酚氧化酶性质研究及意义[J]. 土壤通报, 2006, 37(3): 470 − 474. HAO Jianchao, WU Yanyou, LIAN Bin, et al. Properties of polyphenol oxidase in soil and its significance [J]. Chin J Soil Sci, 2006, 37(3): 470 − 474. [24] TOSCANO G, COLARIETI M L, Jr GRECO G. Oxidative polymerisation of phenols by a phenoloxidase from green olives [J]. Enzyme Microb Technol, 2003, 33(1): 47 − 54. [25] TU Cheng, TENG Ying, LUO Yongming, et al. PCB removal, soil enzyme activities and microbial community structures during the phytoremediation by alfalfa in field soils [J]. J Soils Sediments, 2011, 11(4): 649 − 656. [26] 盛亚萍, 赵成章, 张静, 等. 高寒山区混播草地燕麦和毛苕子根长密度分布格局[J]. 生态学杂志, 2013, 32(2): 279 − 284. SHENG Yaping, ZHAO Chengzhang, ZHANG Jing, et al. Spatial distribution patterns of root length density of Avena sativa and Vicia villosa in their mixed-sowing grassland in alpine region [J]. Chin J Ecol, 2013, 32(2): 279 − 284. [27] 徐炳成, 山仑, 黄占斌, 等. 沙打旺与柳枝稷单、混播种苗期水分利用和根冠生长的比较[J]. 应用与环境生物学报, 2004, 10(5): 577 − 580. XU Bingcheng, SHAN Lun, HUANG Zhanbin, et al. Comparative study of water use efficiency and growth of Astrafalus adsurgens and Panicum virgatum seedings under separated and mixed plantation [J]. Chin J Appl Environ Biol, 2004, 10(5): 577 − 580. [28] 来幸樑, 师尚礼, 吴芳, 等. 紫花苜蓿与3种多年生禾草混播草地的土壤养分特征[J]. 草业科学, 2020, 37(1): 52 − 64. LAI Xingliang, SHI Shangli, WU Fang, et al. Nutrient characteristics of soil sowed with different combinations of alfalfa and three perennial grasses [J]. Acta Prataculturae Sin, 2020, 37(1): 52 − 64. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200486
 点击查看大图
点击查看大图
计量
- 文章访问数: 1932
- HTML全文浏览量: 514
- PDF下载量: 42
- 被引次数: 0
