




-
欧李Cerasus humilis为蔷薇科Rosaceae樱桃属Ceraras灌木类果树,是世界驯化最晚的珍稀灌木,属中国独有,栽培历史约3 000 a[1]。欧李的盛花期一般在4月,春可观花夏可观果,具有很高的观赏价值。欧李果实不仅可以观赏而且含钙量高,营养丰富,鲜果中含钙量是600 mg·kg−1[2],所以又称“钙果”,可以加工成果酒等食品[3-4]。欧李喜光,具有较强的抗寒、抗旱、抗瘠薄能力,是适应性极强的优良树种,能承受−40~−35 ℃的低温[5];在年降水量约200 mm、无霜期120 d的条件下可正常开花结果;根系发达,属强分蘖根系,根冠比达9∶1[2];欧李庞大的根系盘根错节、纵横交错,集中分布在20~40 cm的土层内,深可达1.5~2.0 m,形成表土密集的网状结构,将20 cm深土层中的土壤紧紧包住,显示出极强的固水保土作用,是沙漠化地区治沙造林的优良树种[6-8]。欧李作为防治荒漠化的优良树种已在中国西北地区广泛种植。2011年,宁夏从山西引进欧李,在退化压砂地试种成功后[9],短短几年时间种植区域已覆盖中卫市沙坡头区、海原县、同心县、盐池县等区域。随着欧李引种及种植面积的扩大,其害虫发生严重,尤其是果实食心虫[10-11],严重影响了欧李果实的品质和产量。目前,有关欧李园昆虫群落多样性及害虫研究较少,从而制约了欧李害虫的防控和产业的发展,因此,开展宁夏不同区域欧李园主要害虫和天敌田间发生动态、昆虫群落结构及多样性研究,将为欧李不同种植区确定害虫防治关键时期,制定防控措施和保护天敌提供技术依据,从而为欧李害虫可持续防控及欧李产业发展奠定基础。
-
本研究所选欧李园包括:①玉泉营欧李园位于宁夏回族自治区北部引黄灌区玉泉营苗木繁育中心,其周围为温棚,种植蔬菜;海拔900~1 500 m,年降水量200 mm,年日照时数2 800~3 100 h,无霜期149~161 d;土壤主要有灌淤土、灰钙土、潮土、盐碱土等。②施记圈子欧李园位于中部干旱区盐池县高沙窝镇施记圈子村,其周围为农田,种植玉米Zea mays;海拔1 300~1 500 m,年降水量200~350 mm,年日照时数2 850 h,无霜期141 d;土壤以灰钙土、风沙土为主。③候磨欧李园位于南部黄土丘陵区固原市原州区彭堡镇候磨村,其周围为农田,种植蔬菜和牧草;海拔1 248~2 942 m,年降水量350~500 mm,年日照时数2 200~2 700 h,无霜期113~149 d;土壤以黑垆土、黄绵土为主(表1)。本研究调查的3个欧李园在采样前已经喷洒过农药,统一田间管理。在研究开展期间,3个欧李园并未施过农药。
表 1 不同区域欧李园种植密度
Table 1. Planting density in C. humilis fields
欧李园 经度(E) 纬度(N) 树龄/
a面积/
hm2株行距/
(m×m)玉泉营 106°12′00″ 38°28′48″ 2 2.01 0.6×1.2 候磨 106°07′48″ 36°07′12″ 2 0.67 0.5×1.0 施记圈子 106°51′00″ 38°01′48″ 7 0.67 0.5×1.0 -
本研究采用梨小食心虫Grapholitha molesta诱捕器诱集法,于2019年5−9月进行了5次昆虫群落调查,隔30 d调查1次。根据欧李树体大小及种植密度,悬挂梨小食心虫诱捕器密度为60~75个·hm−2,诱捕器间距10~20 m,呈“W”型排列,性诱芯与粘虫板(北京中捷四方生物科技股份有限公司提供)组合为三角形诱捕器,先将白色钙塑板折成三角形状,再用细铁丝将性诱芯穿过诱捕器的中心,并将粘虫板的覆膜揭开,性诱芯与底面的垂直距离为1 cm。诱捕器用铁丝固定在竹竿上,挂在离地100 cm处,每月收集1次,并更换新的粘虫板。每个粘虫板的两边采用保鲜膜覆盖,防止相互粘连。所有粘虫板统一收集,带回实验室鉴定,并进行统计。试验期间,每日记录平均风速(m·s−1)、日平均气温(℃)、日最高气温(℃)、日最低气温(℃)、降水量(mm)等气象因子。
-
本研究数据通过Excel 2010、DPS(18.10)和SAS.v 8.2进行统计分析,运用生态学的分析方法主要分析以下参数[12-16]。① Margalef丰富度指数(d):d=(S−1)/lnN。其中:S为物种数;N为所有物种的个体数之和。② Simpson集中性概率指数(λ):
$\lambda = \displaystyle\sum\limits_{i=1}^3 {\frac{{{N_i}({N_i} - 1)}}{{N(N - 1)}}}$ 。其中:Ni为第i监测区物种个体数之和。λ越大,说明群落的集中性越高,多样性越低。③ Shannon-Wiener多样性指数(H′):$H' = - \displaystyle\sum\limits_{i = 1}^S {{P_i}} \ln {P_i}$ 。其中:S为物种数;Pi为i种个体占总个体数的比例。④ Pielou均匀度指数(E):E= H′/ H′max= H′/lnS。⑤群落Jaccard相似性系数(q):q=c/(a+b+c)。其中:c为2个群落共有物种数;a和b分别为群落A和群落B的物种数[17]。根据相似性系数原理,判断相似性程度。若0≤q<0.25,表示群落A和群落B极不相似;若0.25≤q<0.50,表示中等不相似;若0.50≤q<0.75,表示中等相似;若0.75≤q<1.00,表示极相似。⑥群落相对稳定性测定。采用群落物种数与个体数之比(St/Si)和天敌类群物种数与植食性类群物种数之比(Sn/Sp)表示;St表示群落物种数,Si表示群落个体数,St/Si反映种间数量上的制约作用,St/Si越大说明种间的制约作用越强;Sn表示天敌类群物种数,Sp植食性类群物种数,Sn/Sp反映食物网关系和天敌—害虫相互制约的复杂程度,Sn/Sp越大说明昆虫群落内部的食物网结构和相互制约关系越复杂[18]。 -
2019年5−9月在玉泉营、施记圈子、候磨欧李园采用诱捕器诱集法共获得标本4 743头,隶属于10目46科101种。主要昆虫群落组成及数量特征如表2所示(因篇所限,仅列出前10种)。由表2可见:主要昆虫群落的优势种有梨小食心虫、多异瓢虫Hippodamia variegata、小长蝽Nysius ericae、东方绢金龟Serica orientalis、黑点食蚜盲蝽Deraeocoris punctulatus、丽草蛉Chrysopa formosa、红翅伪叶甲Lagria rufipennis、牧草盲蝽Lygus pratensis、艾箭蚁Cataglyphis aenescens和双斑萤叶甲Monolepta hieroglyphica;个体数量分别占群落个体总数的27.05%、16.72%、7.95%、6.56%、3.98%、3.46%、3.16%、2.49%、1.90%和1.83%。其中,在玉泉营欧李园共获昆虫标本2289头,隶属于10目36科69种,优势种为梨小食心虫、多异瓢虫、小长蝽、红翅伪叶甲、黑点食蚜盲蝽、东方绢金龟、细胸锥尾叩甲Agriotes fusicollis、弱脊异丽金龟Anomala sucipennis和福婆鳃金龟Brahmina falderman,分别占个体总数的30.32%、17.21%、7.60%、6.20%、4.94%、4.54%、3.01%、2.36%和2.18%;施记圈子欧李园共获昆虫样本1104头,隶属于7目24科44种,其优势种为梨小食心虫、丽草蛉、东方绢金龟、艾箭蚁、小长蝽、多异瓢虫、黑点食蚜盲蝽,分别占个体总数的51.09%、10.87%、8.51%、5.53%、5.43%、4.98%和1.99%;候磨欧李园共获昆虫样本1350头,隶属于7目36科36种,其优势种为多异瓢虫、小长蝽、东方绢金龟、牧草盲蝽、双斑萤叶甲、中华布里隐翅虫Bledius chinensis、黑点食蚜盲蝽、淡色姬蝽Nabis palifer、牛虻Atylotus sp、黄鞘婪步甲Harpalus pallidipennis、丽草蛉和梨小食心虫,分别占个体总数的25.48%、10.59%、8.37%、7.48%、5.63%、4.22%、4.00%、3.11%、2.52%、2.44%、2.30%和1.85%。3个欧李园内,梨小食心虫比例均最大,从科数、物种数和个体数来看,玉泉营欧李园均最多。
表 2 不同区域欧李园主要昆虫群落组成及数量特征
Table 2. Species composition and quantity of main insect community in C. humilis fields in different growing areas
昆虫种类 玉泉营 施记圈子 候磨 合计 数量/头 比例/% 数量/头 比例/% 数量/头 比例/% 数量/头 比例/% 梨小食心虫 694 30.32 564 51.09 25 1.85 1283 27.05 多异瓢虫 394 17.21 55 4.98 344 25.48 793 16.72 小长蝽 174 7.60 60 5.43 143 10.59 377 7.95 东方绢金龟 104 4.54 94 8.51 113 8.37 311 6.56 黑点食蚜盲蝽 113 4.94 22 1.99 54 4.00 189 3.98 丽草蛉 13 0.57 120 10.87 31 2.30 164 3.46 红翅伪叶甲 142 6.20 5 0.45 3 0.22 150 3.16 牧草盲蝽 13 0.57 4 0.36 101 7.48 118 2.49 艾箭蚁 29 1.27 61 5.53 0 0.00 90 1.90 双斑萤叶甲 10 0.44 1 0.09 76 5.63 87 1.83 -
由图1可知:3个欧李园昆虫群落丰富度指数、多样性指数和均匀度指数由高到低依次为候磨欧李园、玉泉营欧李园、施记圈子欧李园;昆虫群落集中性概率指数由高到低依次为施记圈子欧李园、玉泉营欧李园、候磨欧李园。从丰富度指数和多样性指数看,候磨最高,施记圈子最低,且施记圈子与候磨、玉泉营的丰富度指数和多样性指数之间存在显著差异(P<0.05)。从均匀度指数上看,候磨最高,且候磨与施记圈子、玉泉营之间存在显著差异(P<0.05),表明候磨欧李园内昆虫物种分布相对均衡,没有突发性,种间具有一定的制约能力。从集中性概率指数上看,施记圈子的集中性概率指数显著高于其他2个欧李园(P<0.05),表明施记圈子优势类群的地位比较突出。

图 1 不同区域欧李园昆虫群落特征指数
Figure 1. Characteristic index of insect communities for C. humilis fields in different growing areas
-
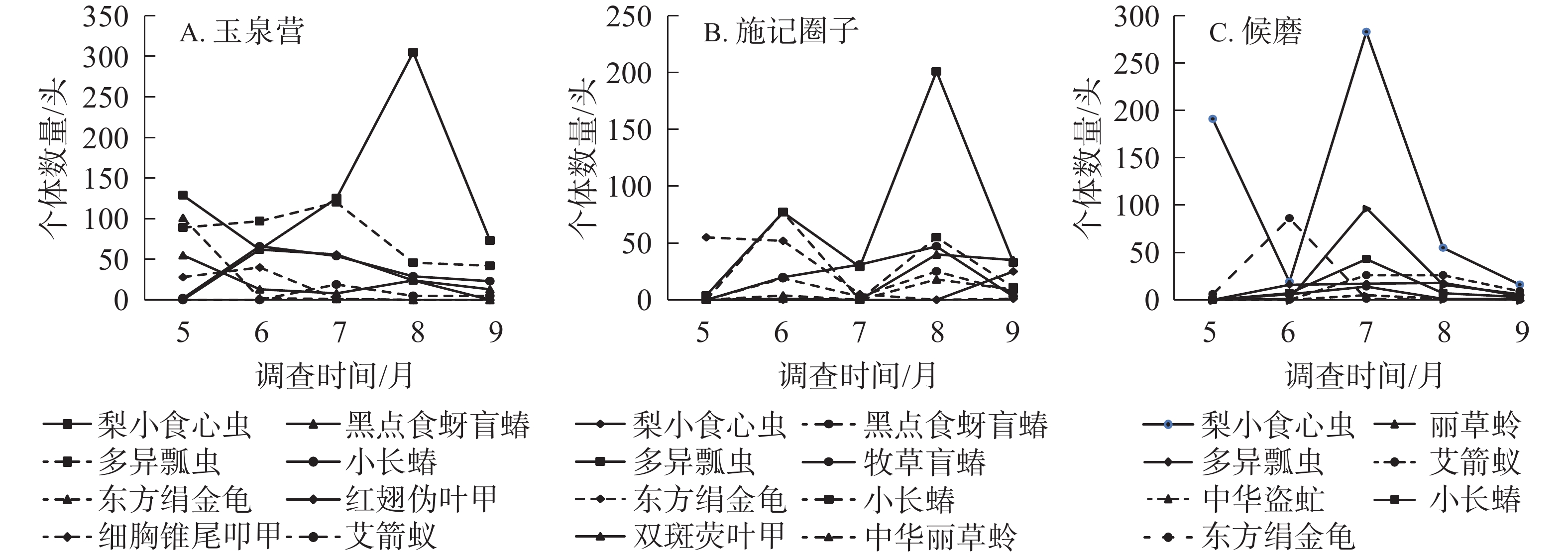
宁夏不同区域欧李园的主要类群动态变化如图2所示。玉泉营欧李园害虫类群主要有梨小食心虫、东方绢金龟、小长蝽、红翅伪叶甲、细胸锥尾叩甲;天敌类群主要有多异瓢虫、黑点食蚜盲蝽、艾箭蚁。整体来看,主要类群到达峰值时间不同,害虫类群的数量大于天敌类群的数量,且梨小食心虫的发生量在5、7、8、9月均比天敌类群的发生量多;果实收获后,主要类群个体数减少,到9月个体数达到最低。从昆虫种类来看,梨小食心虫的发生量上下波动幅度较大,5−8月呈直线上升趋势,8月每个诱集板平均数达到最大值,为305头;东方绢金龟主要发生在5月,每个诱集板平均数最多可达101头;小长蝽主要发生在6月,每个诱集板平均数最大可达66头;红翅伪叶甲主要发生在6月,每个诱集板平均数最大可达62头;细胸锥尾叩甲主要发生在6月,每个诱集板平均数最大可达40头;多异瓢虫主要发生在7月,每个诱集板平均数最多为120头;黑点食蚜盲蝽主要发生在5月,每个诱集板平均数可达55头;艾箭蚁主要发生在7月,每个诱集板平均数最多为19头(图2A)。
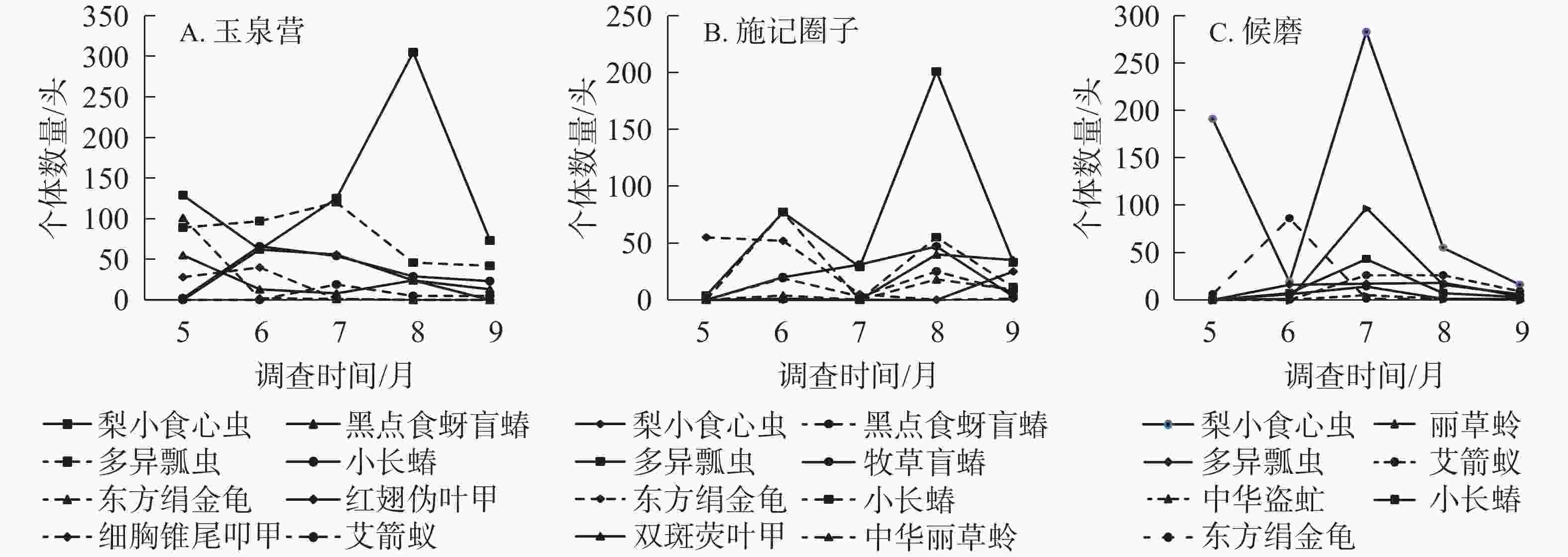
图 2 不同区域欧李园主要害虫及天敌昆虫的时间动态
Figure 2. Temporal dynamic of major insect in C. humilis fields in different growing areas
施记圈子欧李园的害虫种类主要有梨小食心虫、东方绢金龟和小长蝽;天敌种类主要有多异瓢虫、黑点食蚜盲蝽、中华盗虻 Cophinopoda chinensis、丽草蛉和艾箭蚁。整体来看,5月欧李园天敌类群发生量较少,而梨小食心虫的发生量较多,欧李主要害虫及天敌个体数量在7月达到高峰。9月欧李主要害虫与天敌个体数量减少到最低。从昆虫种类来看,梨小食心虫的发生量上下波动较大,5−6月下降,7月上升到最高峰,每个诱集板平均数最多达283头,8、9月逐渐下降到最低值。东方绢金龟主要发生在6月,每个诱集板平均数最多可达86头。丽草蛉主要发生在7月,每个诱集板平均数最多可达97头。小长蝽主要发生在7月,每个诱集板平均数最大可达43头。艾箭蚁主要发生在7、8月,每个诱集板最多,为26头。黑点食蚜盲蝽主要发生在7月,每个诱集板平均数可达14头。中华盗虻主要发生在7月,其每个诱集板平均数为5头。多异瓢虫主要发生在6、7、8月,其中8月的发生量最大,每个诱集板平均数为18头(图2B)。
候磨欧李园害虫类群主要有梨小食心虫、东方绢金龟、牧草盲蝽、小长蝽、双斑萤叶甲;天敌类群主要有多异瓢虫、黑点食蚜盲蝽、中华丽草蛉。整体来看,5月欧李园天敌类群发生量较少,而东方绢金龟的发生量较多,在2019年6月,主要类群数量达到第1个峰,在2019年8月达到第2个峰,且天敌类群的数量较多。从昆虫种类来看,梨小食心虫一直到9月才出现,其每个诱集板平均数为25头;东方绢金龟主要发生在5月,最多可达55头;小长蝽主要发生在6月,每个诱集板平均数最大可达77头;牧草盲蝽主要发生在8月,每个诱集板平均数最多可达47头;双斑萤叶甲主要发生在8月,每个诱集板平均数最大可达40头;多异瓢虫主要发生在8月,每个诱集板平均数最多为201头;黑点食蚜盲蝽主要发生在8月,每个诱集板平均数发生量可达25头;丽草蛉主要发生在8月,每个诱集板平均数为18头(图2C)。
-
从不同时间不同区域欧李园的相似性来看(表3):5月,3个欧李园的相似性系数均大于0,小于0.25,表明5月3个欧李园的昆虫群落结构极不相似;6月,3个欧李园的相似性系数均未达0.50,其中施记圈子欧李园与候磨欧李园的相似性系数为0.24(0≤q<0.25),说明施记圈子欧李园与候磨欧李园在6月的昆虫群落结构极不相似;其他地区欧李园的相似性系数均大于0.25,小于0.50,表明其他地区欧李园之间的昆虫群落结构在6月均为中等不相似。7月候磨欧李园与施记圈子欧李园、玉泉营欧李园的相似性系数为0.15、0.16(0≤q<0.25),说明候磨欧李园与施记圈子欧李园、玉泉营欧李园在7月的昆虫群落结构极不相似;玉泉营欧李园与施记圈子欧李园的相似系数大于0.25,小于0.50,表明玉泉营欧李园和施记圈子欧李园在7月昆虫群落结构均为中等不相似;玉泉营欧李园与施记圈子欧李园在8月的相似性系数较高,达0.60,表明玉泉营欧李园和施记圈子欧李园在8月的昆虫群落结构中等相似,其他地区欧李园的相似性系数均大于0.25,小于0.50,表明其他地区欧李园之间的昆虫群落结构在8月均为中等不相似;9月,3个欧李园的相似性系数均大于0.25,小于0.50,表明9月3个地区欧李园的昆虫群落结构中等不相似。由此可知,不同时间同一空间或同一时间不同空间具有不同的昆虫群落组成特点及变化程度,说明不同的昆虫种类可以利用不同的生态位资源,出现时间或空间差异使其可以尽量减少竞争。
表 3 欧李园各月昆虫群落相似性分析
Table 3. Similarity analysis of insect community in C. humilis fields in different months
欧李园 月份 施记圈子 候磨 5 6 7 8 9月 5 6 7 8 9月 玉泉营 5 0.12 0.23 6 0.30 0.28 7 0.41 0.15 8 0.60 0.29 9 0.38 0.40 施记圈子 5 0.08 6 0.24 7 0.16 8 0.29 9 0.26 -
由表4可见:3个欧李园之间的St/Si从大到小依次为候磨欧李园、施记圈子欧李园、玉泉营欧李园,说明候磨欧李园昆虫的物种种类数相对于个体数较多,种间的制约作用较强,个别物种的变动产生的影响比另外2个欧李园小,昆虫种类分布相对均衡。Sn/Sp从大到小依次为候磨欧李园、玉泉营欧李园、施记圈子欧李园,说明候磨欧李园昆虫群落内部的食物网结构和相互制约关系较另外2个欧李园复杂,所以遇到外界的干扰时,昆虫群落的抗干扰性比较高,天敌类群相对于其本身的植食类群来说较多。结合这2点分析可以看出:候磨欧李园昆虫群落的稳定性比其他2个欧李园高。
表 4 不同区域欧李园昆虫群落相对稳定性数值
Table 4. Relative stability values of insect communities in different C. humilis fields
指标 玉泉营 施记圈子 候磨 St/Si 0.030 1 0.040 8 0.052 6 Sn/Sp 0.565 5 0.551 7 0.666 7 -
欧李园主要害虫与天敌及环境因子的相关分析表明(表5):昆虫群落的物种数与平均风速、日平均气温、日最高气温和日最低气温均呈现相关性,其中与平均风速表现为极显著负相关(P<0.001),说明风速越大,昆虫群落的物种数就越少;与日最高气温和日最低气温表现为极显著正相关(P<0.01);与日平均气温表现为显著正相关(P<0.05)。小长蝽与日平均气温、日最高气温和日最低气温均呈现相关性,其中与日最低气温表现为极显著正相关(P<0.01),与日平均气温和日最高气温表现为显著正相关(P<0.05)。红翅伪叶甲与日平均气温、日最高气温和日最低气温均呈显著正相关(P<0.05)。双斑萤叶甲与降水量表现为极显著正相关(P<0.01)。
表 5 欧李园主要害虫与天敌及环境因子的相关性分析
Table 5. Analysis of the correlation between major insect and environmental factors in C. humilis fields
项目 个体数 物种数 梨小食心虫数量 小长蝽数量 红翅伪叶甲数量 东方绢金龟数量 双斑萤叶甲数量 平均风速 0.091 3 −0.858 0*** 0.282 1 −0.273 4 −0.348 7 0.501 8 −0.376 1 日平均气温 0.487 4 0.622 0* 0.274 7 0.606 1* 0.629 0* −0.050 8 −0.241 9 日最高气温 0.470 7 0.650 4** 0.282 5 0.570 8* 0.615 5* −0.108 0 −0.260 0 日最低气温 0.473 5 0.669 3** 0.220 1 0.659 7** 0.624 0* −0.076 9 −0.129 7 降水量 −0.120 4 0.305 5 −0.375 4 0.092 5 −0.119 7 −0.162 6 0.726 2** 丽草蛉数量 −0.132 9 0.482 0 −0.355 0 0.217 4 0.105 3 −0.277 6 0.887 2*** 艾箭蚁数量 −0.042 5 0.451 6 −0.339 0 0.128 8 0.602 4* −0.204 2 −0.053 1 多异瓢虫数量 0.879 6*** 0.456 5 0.570 7* 0.827 6** 0.170 5 −0.060 1 0.227 7 黑点食蚜盲蝽数量 0.561 6* 0.275 7 0.324 5 0.244 1 −0.073 4 0.402 4 −0.019 4 说明:*表示在0.05水平上差异显著 (P<0.05);**表示在0.01水平上差异显著(P<0.01);***表示在0.001水平上差异显著(P<0.001) 昆虫群落个体数与多异瓢虫、黑点食蚜盲蝽均呈现相关性,其中与多异瓢虫表现为极显著正相关(P<0.001),说明天敌多异瓢虫数量越多,昆虫群落的个体数就越多;与黑点食蚜盲蝽表现为显著正相关(P<0.05)。从害虫与天敌的相关性可以看出:梨小食心虫与多异瓢虫表现为显著正相关(P<0.05)。小长蝽与多异瓢虫表现为极显著正相关(P<0.01)。红翅伪叶甲与艾箭蚁表现为显著正相关(P<0.05)。双斑萤叶甲与丽草蛉表现为极显著正相关(P<0.001)。说明欧李园内害虫的数量越多,天敌的数量也随之增多。
-
宁夏3个不同区域欧李园共获得标本4 743头,隶属于10目46科101种。昆虫群落丰富,结构复杂,其中为害欧李的主要害虫为梨小食心虫、东方绢金龟、红翅伪叶甲、双斑萤叶甲、小长蝽和牧草盲蝽。天敌资源丰富,主要天敌种类有多异瓢虫、龟纹瓢虫Propylaea japonica、七星瓢虫Coccinella septmpunctata、二星瓢虫Adalia bipunctata、双七瓢虫Coccinula quatuordecimpustulata、小花蝽Orius similis、黑点食蚜盲蝽、丽草蛉和艾箭蚁。梨小食心虫捕食性天敌主要有蜘蛛、蚂蚁、草蛉、瓢虫、花蝽等,寄生性天敌主要有松毛虫赤眼蜂Trichogramma dendrolimi、广赤眼蜂Trichogramma evnescens和玉米螟赤眼蜂Trichogramma ostriniae[19-20]。这与本研究结果相似,其中,丽草蛉、艾箭蚁、多异瓢虫及黑点食蚜盲蝽等为优势种。多异瓢虫是本研究3个不同区域欧李园中的优势天敌,且多异瓢虫密度与梨小食心虫和小长蝽的密度呈显著正相关,艾箭蚁与红翅伪叶甲呈显著正相关(P<0.05),丽草蛉与双斑萤叶甲呈极显著正相关(P<0.001),表明天敌群落对害虫群落有着控制作用,这对控制欧李园虫害暴发及稳定欧李园生态系统平衡有重要作用[21]。梨小食心虫在候磨、施记圈子、玉泉营欧李园的发生高峰期分别为9、7和8月;王志梅等[11]对山西钙果园中梨小食心虫发生规律研究发现:梨小食心虫在6月发生量最多;刘显臣等[22]对长白山欧李蛀果害虫发生规律研究发现:梨小食心虫在7月下旬羽化峰值最高。由此可见:在中国不同欧李种植区,梨小食心虫发生规律不同,其主要原因是害虫发生与环境及气候变化因子密切相关[23],因此,中国不同欧李种植区的梨小食心虫的防治关键期也不同。此外,不同的时间段,欧李园的主要害虫和天敌的优势种种类均不相同。确定主要害虫和天敌在时间上的同步性以及在空间上的同域性[24],可以合理地保护和利用自然天敌,从而进行害虫的综合防治。
对宁夏3个不同区域欧李园的昆虫群落多样性指数、均匀度指数和群落稳定性分析得出:各昆虫群落稳定性从大到小依次为候磨欧李园、玉泉营欧李园、施记圈子欧李园。候磨欧李园的昆虫群落在丰富度指数、多样性指数和均匀度指数上显著高于其他2个欧李园,说明候磨欧李园昆虫群落物种分布最均匀,群落组成更为丰富;在候磨欧李园,群落物种数与个体数的St/Si和天敌种数与害虫种数的Sn/Sp均偏高。在不同欧李园中天敌是影响昆虫群落结构的主要因子,从侯磨欧李园主要类群的发生动态可以看出:捕食性天敌类群对植食性昆虫的控制作用较强。这些指标都表明:候磨欧李园昆虫群落稳定性相对较高,结构复杂,对于外界的适应调节能力最强。另外2个欧李园的昆虫群落,从均匀度指数和多样性指数的一致性来看,玉泉营欧李园的稳定性要强于施记圈子欧李园,而St/Si则显示施记圈子欧李园大于玉泉营欧李园,但数值相差较小。综合比较推断:玉泉营欧李园稳定性要高于施记圈子欧李园。群落相似性分析显示:不同欧李园在5、6、7、9月的昆虫群落结构不相似,玉泉营欧李园与施记圈子欧李园在8月的相似性系数较高(0.60),说明不同昆虫种类可以利用不同的生态位资源,出现时间的差异可以尽量减少竞争[25]。
Insect community diversity of Cerasus humilis orchards in different regions of Ningxia
-
摘要:
目的 研究宁夏不同区域欧李Cerasus humilis园昆虫群落结构及多样性特征,为制定有效的害虫防治策略奠定基础。 方法 2019年5−9月采用诱捕器诱集法对宁夏3个不同区域欧李园昆虫群落进行调查。 结果 共获得标本4 743头,隶属于10目46科101种。梨小食心虫Grapholitha molesta、东方绢金龟Serica orientalis、小长蝽Nysius ericae、牧草盲蝽Lygus pratensis、双斑萤叶甲Monolepta hieroglyphica、红翅伪叶甲Lagria rufipennis、多异瓢虫Hippodamia variegata、丽草蛉Chrysopa formosa、艾箭蚁Cataglyphis aenescens和黑点食蚜盲蝽Deraeocoris punctulatus为优势种。宁夏不同区域欧李园昆虫群落多样性指数、均匀度指数及丰富度指数由高到低依次为候磨欧李园、玉泉营欧李园、施记圈子欧李园;集中性概率指数由高到低依次为施记圈子欧李园、玉泉营欧李园、候磨欧李园。相似性分析显示:玉泉营欧李园与施记圈子欧李园在8月的昆虫群落结构中等相似(0.60)。害虫与天敌的相关性分析显示:梨小食心虫与多异瓢虫呈显著正相关(P<0.05);小长蝽与多异瓢虫呈极显著正相关(P<0.01);红翅伪叶甲与艾箭蚁呈显著正相关(P<0.05);双斑萤叶甲与丽草蛉呈极显著正相关(P<0.01)。 结论 相对于玉泉营欧李园和施记圈子欧李园,候磨欧李园的昆虫群落稳定性最高。图2表5参25 Abstract:Objective This study aims to clarify the structure and diversity characteristics of insect community of Cerasus humilis orchards in different regions of Ningxia, so as to develop effective pest control strategies. Method The insect community of C. humilis in three different regions of Ningxia was investigated using trap trapping method from May to September in 2019. Result A total of 4743 insect specimens belonging to 10 orders, 46 families and 101 species were obtained. The dominant species were Grapholitha molesta, Serica orientalis, Nysius ericae, Lygus pratensis, Monolepta hieroglyphica, Lagria rufipennis, Hippodamia variegate, Chrysopa formosa, Cataglyphis aenescens and Deraeocoris punctulatus. The descending order of diversity index, evenness index and richness index of insect community of C. humilis orchards was Houmo, Yuquanying, and Shijijuanzi. The concentration probability index from high to low was Shijijuanzi, Yuquanying and Houmo. Similarity analysis showed that the insect community structure in Yuquanying and Shijijuanzi C. humilis orchards was moderately similar in August(0.60). The correlation analysis between pests and natural enemies showed that there was a significant positive correlation between G. molesta and H. variegata (P<0.05). There existed a very significant positive correlation between N. ericae and H. variegata (P<0.01), a positive correlation between L. rufipennis and C. aenescens (P<0.05), and a significant positive correlation between M. hieroglyphica and C. formosa (P<0.01). Conclusion Compared with orchards in Yuquanying and Shijijuanzi, the C. humilis orchard in Houmo has highest stability of insect community. [Ch, 2 fig. 5 tab. 25 ref.] -
Key words:
- Cerasus humilis /
- planting area /
- insect /
- community structure /
- diversity
-
图 1 不同区域欧李园昆虫群落特征指数
不同字母表示不同欧李园间存在显著差异(P<0.05)
Figure 1 Characteristic index of insect communities for C. humilis fields in different growing areas
图 2 不同区域欧李园主要害虫及天敌昆虫的时间动态
Figure 2 Temporal dynamic of major insect in C. humilis fields in different growing areas
表 1 不同区域欧李园种植密度
Table 1. Planting density in C. humilis fields
欧李园 经度(E) 纬度(N) 树龄/
a面积/
hm2株行距/
(m×m)玉泉营 106°12′00″ 38°28′48″ 2 2.01 0.6×1.2 候磨 106°07′48″ 36°07′12″ 2 0.67 0.5×1.0 施记圈子 106°51′00″ 38°01′48″ 7 0.67 0.5×1.0  下载: 导出CSV
下载: 导出CSV
表 2 不同区域欧李园主要昆虫群落组成及数量特征
Table 2. Species composition and quantity of main insect community in C. humilis fields in different growing areas
昆虫种类 玉泉营 施记圈子 候磨 合计 数量/头 比例/% 数量/头 比例/% 数量/头 比例/% 数量/头 比例/% 梨小食心虫 694 30.32 564 51.09 25 1.85 1283 27.05 多异瓢虫 394 17.21 55 4.98 344 25.48 793 16.72 小长蝽 174 7.60 60 5.43 143 10.59 377 7.95 东方绢金龟 104 4.54 94 8.51 113 8.37 311 6.56 黑点食蚜盲蝽 113 4.94 22 1.99 54 4.00 189 3.98 丽草蛉 13 0.57 120 10.87 31 2.30 164 3.46 红翅伪叶甲 142 6.20 5 0.45 3 0.22 150 3.16 牧草盲蝽 13 0.57 4 0.36 101 7.48 118 2.49 艾箭蚁 29 1.27 61 5.53 0 0.00 90 1.90 双斑萤叶甲 10 0.44 1 0.09 76 5.63 87 1.83
下载: 导出CSV
表 3 欧李园各月昆虫群落相似性分析
Table 3. Similarity analysis of insect community in C. humilis fields in different months
欧李园 月份 施记圈子 候磨 5 6 7 8 9月 5 6 7 8 9月 玉泉营 5 0.12 0.23 6 0.30 0.28 7 0.41 0.15 8 0.60 0.29 9 0.38 0.40 施记圈子 5 0.08 6 0.24 7 0.16 8 0.29 9 0.26
下载: 导出CSV
表 4 不同区域欧李园昆虫群落相对稳定性数值
Table 4. Relative stability values of insect communities in different C. humilis fields
指标 玉泉营 施记圈子 候磨 St/Si 0.030 1 0.040 8 0.052 6 Sn/Sp 0.565 5 0.551 7 0.666 7
下载: 导出CSV
表 5 欧李园主要害虫与天敌及环境因子的相关性分析
Table 5. Analysis of the correlation between major insect and environmental factors in C. humilis fields
项目 个体数 物种数 梨小食心虫数量 小长蝽数量 红翅伪叶甲数量 东方绢金龟数量 双斑萤叶甲数量 平均风速 0.091 3 −0.858 0*** 0.282 1 −0.273 4 −0.348 7 0.501 8 −0.376 1 日平均气温 0.487 4 0.622 0* 0.274 7 0.606 1* 0.629 0* −0.050 8 −0.241 9 日最高气温 0.470 7 0.650 4** 0.282 5 0.570 8* 0.615 5* −0.108 0 −0.260 0 日最低气温 0.473 5 0.669 3** 0.220 1 0.659 7** 0.624 0* −0.076 9 −0.129 7 降水量 −0.120 4 0.305 5 −0.375 4 0.092 5 −0.119 7 −0.162 6 0.726 2** 丽草蛉数量 −0.132 9 0.482 0 −0.355 0 0.217 4 0.105 3 −0.277 6 0.887 2*** 艾箭蚁数量 −0.042 5 0.451 6 −0.339 0 0.128 8 0.602 4* −0.204 2 −0.053 1 多异瓢虫数量 0.879 6*** 0.456 5 0.570 7* 0.827 6** 0.170 5 −0.060 1 0.227 7 黑点食蚜盲蝽数量 0.561 6* 0.275 7 0.324 5 0.244 1 −0.073 4 0.402 4 −0.019 4 说明:*表示在0.05水平上差异显著 (P<0.05);**表示在0.01水平上差异显著(P<0.01);***表示在0.001水平上差异显著(P<0.001)
下载: 导出CSV
-
[1] 曹琴, 张玉娥, 岳文英. 欧李名与实研究[J]. 山西农业大学学报(社会科学版), 2002, 1(3): 247 − 249. CAO Qin, ZHANG Yu’e, YUE Wenying. A study of name and entity of chinese dwarf cherry [J]. J Shanxi Agric Univ Soc Sci Ed, 2002, 1(3): 247 − 249. [2] 薛晓芳, 杜俊杰. 浅谈欧李的开发利用价值[J]. 落叶果树, 2012, 44(3): 23 − 26. XUE Xiaofang, DU Junjie. On the value of Cerasus humilis development and utilization [J]. Deciduous Fruits, 2012, 44(3): 23 − 26. [3] 李卫东, 李欧, 和银霞, 等. 基于TXRF法的欧李果肉中营养元素特征分析[J]. 食品科学, 2015, 36(4): 164 − 167. LI Weidong, LI Ou, HE Yinxia, et al. Analysis of nutrient element characteristics of flesh of Chinese dwarf cherry based on total reflection X-Ray fluorescence method [J]. Food Sci, 2015, 36(4): 164 − 167. [4] 刘婷婷, 马岩石, 李娜, 等. 全果发酵与果渣发酵对欧李果酒香气特征的影响[J]. 食品科学, 2016, 37(12): 99 − 104. LIU Tingting, MA Yanshi, LI Na, et al. Comparison of aroma characteristics of Cerasus humilis wines fermented from whole fruits and pomace [J]. Food Sci, 2016, 37(12): 99 − 104. [5] 刘保东, 孙井山, 杜鹃, 等. 西部特有野生树种欧李优良类型选择研究[J]. 内蒙古林业科技, 2008, 34(4): 33 − 35. LIU Baodong, SUN Jingshan, DU Juan, et al. Choice of good variety of unique wildCerasus humilis west China [J]. J Inn Mong For Sci Technol, 2008, 34(4): 33 − 35. [6] 张东为, 贾天会, 舒乔生. 水保优良树种欧李的研究进展及今后研究方向[J]. 中国水土保持, 2012(1): 45 − 47. ZHANG Dongwei, JIA Tianhui, SHU Qiaosheng. Research progress and future research directions of the excellent soil and water conservation tree species [J]. Soil Water Conserv China, 2012(1): 45 − 47. [7] 毕庆玲, 杜宏志, 王庆茹, 等. 沙地珍贵经济树种的筛选[J]. 防护林科技, 2015(6): 70 − 71. BI Qingling, DU Hongzhi, WANG Qingru, et al. Screening on valuable economic species on sandy land [J]. Prot For Sci Technol, 2015(6): 70 − 71. [8] 张琴, 宋经元, 邵飞, 等. 防风固沙优良树种欧李的潜在适生区及生态特征[J]. 北京林业大学学报, 2018, 40(3): 66 − 74. ZHANG Qin, SONG Jingyuan, SHAO Fei, et al. Potential suitable distribution area and ecological characteristics of Cerasus humilis, an excellent tree species for windproof and sand fixation [J]. J Beijing For Univ, 2018, 40(3): 66 − 74. [9] 张璐瑶. 压砂地欧李品种筛选及水肥耦合试验研究[D]. 银川: 宁夏大学, 2018. ZHANG Luyao. Study on the Varieties Selection and Water-fertilizer Coupling Effects of Cerasus Humilis in Gravel-mulched Field[D]. Yinchuan: Ningxia University, 2018. [10] 李捷, 薛艳花, 王怡, 等. 钙果园桃小食心虫田间消长动态[J]. 山西农业科学, 2015, 43(10): 1308 − 1310. LI Jie, XUE Yanhua, WANG Yi, et al. Population dynamic of Carposina sasakii matsumura in Cerasus humilis (bunge) sok orchard [J]. J Shanxi Agric Sci, 2015, 43(10): 1308 − 1310. [11] 王志梅, 杜俊杰. 钙果园梨小食心虫成虫预测预报与防治研究[J]. 山西果树, 2012(5): 3 − 5. WANG Zhimei, DU Junjie. Forecast and prevention research of oriental fruit moth adult in Cerasus humilis (bunge) sok orchard [J]. J Fruit Resour, 2012(5): 3 − 5. [12] 姜洋, 皮兵. 我国昆虫物种多样性研究现状[J]. 湖南林业科技, 2004(3): 47 − 49. JIANG Yang, PI Bing. Research status of insect species diversity in my country [J]. Hunan For Sci Technol, 2004(3): 47 − 49. [13] 吴亮亮. 油菜田节肢动物群落研究[D]. 杨凌: 西北农林科技大学, 2014. WU Liangliang. Arthropod Community Research on Canola[D]. Yangling: Northwest A&F University, 2014. [14] 韦柳妮. 靖西县烟田节肢动物群落结构及其动态研究[D]. 南宁: 广西大学, 2014. WEI Liuni. Study on the Arthropod Community Structure and Dynamics in Tobacco Field of Jingxi County[D]. Nanning: Guangxi University, 2014. [15] 赵志模, 郭依泉. 群落生态学原理及方法[M]. 重庆: 科学技术出版社重庆分社, 1990: 1 − 48. [16] SCHOENER T W. Resource partitioning in ecological communities [J]. Science, 1974, 185(4145): 27 − 38. [17] 丁岩钦. 昆虫数学生态学[M]. 北京: 科学出版社, 1994. [18] 蒋杰贤, 万年峰, 季香云, 等. 桃园生草对桃树节肢动物群落多样性与稳定性的影响[J]. 应用生态学报, 2001, 22(9): 2303 − 2308. JIANG Jiexian, WAN Nianfeng, JI Xiangyun, et al. Diversity and stability of arthropod community in peach orchard under effects of ground cover vegetation [J]. Chin J Appl Ecol, 2001, 22(9): 2303 − 2308. [19] 穆宝锋, 董兆克, 刘士峰, 等. 4种庇护植物对苹果园梨小食心虫及其主要天敌的种群动态影响[J]. 北京农学院学报, 2019, 34(1): 39 − 44. MU Baofeng, DONG Zhaoke, LIU Shifeng, et al. Effects of four species of refuge plants oriental fruit moth, Grapholita molest on population dynamics of and its major natural enemies [J]. J Beijing Univ Agric, 2019, 34(1): 39 − 44. [20] 孟豪, 赵利鼎, 李先伟, 等. 梨小食心虫的天敌资源[J]. 山西农业科学, 2011, 39(8): 858 − 861. MENG Hao, ZHAO Liding, LI Xianwei, et al. The investigation into natural enemies of Grapholitha molesta (Lepidoptera: Tortricoidea) [J]. J Shanxi Agric Sci, 2011, 39(8): 858 − 861. [21] 张晓明, 李强, 陈国华, 等. 不同种植模式花椒园昆虫群落的结构及稳定[J]. 应用生态学报, 2009, 20(8): 1986 − 1991. ZHANG Xiaoming, LI Qiang, CHEN Guohua, et al. Insect community structure and its stability in a Zanthoxylum bungeanum garden with different planting pattern [J]. Chin J Appl Ecol, 2009, 20(8): 1986 − 1991. [22] 刘显臣, 陈丽, 徐霞, 等. 长白山钙果蛀果害虫发生规律及药剂防治[J]. 吉林农业科学, 2014, 39(1): 58 − 60. LIU Xianchen, CHEN Li, XU Xia, et al. Occurrence and chemical control of fruit borer ofPrunus humilis bunge [J]. J Jilin Agric Sci, 2014, 39(1): 58 − 60. [23] 史普想, 于国庆, 于洪波, 等. 辽宁阜新花生产区昆虫群落结构及多样性分析[J]. 花生学报, 2019, 48(1): 40 − 47. SHI Puxiang, YU Guoqing, YU Hongbo, et al. Analysis of insect community structure and diversity in peanut fields in Fuxin, Liaoning Province [J]. J Peanut Sci, 2019, 48(1): 40 − 47. [24] 王东. 桃园昆虫群落结构特征及主要害虫防控技术的研究[D]. 泰安: 山东农业大学, 2010. WANG Dong. Study on the Structure Characteristics and Control Technology of Insect Community in Peach Orchard[D]. Tai’an: Shandong Agricultural University, 2010. [25] 张勇. 北京八达岭林场油松林昆虫群落多样性研究[D]. 北京: 北京林业大学, 2007. ZHANG Yong. Diversity of Insect Community in Chinese Pine Forest from Badaling Forest Center in Beijing[D]. Beijing: Beijing Forestry University, 2007. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200774
 点击查看大图
点击查看大图
计量
- 文章访问数: 2419
- HTML全文浏览量: 641
- PDF下载量: 40
- 被引次数: 0


