




-
植被数量分类、排序是用于研究森林[1-4]、草地[5-8]、湿地[9]等群落及环境关系的重要生态学手段,是利用数量生态学研究群落的主流方法之一,它通过数量分析手段客观准确地揭示了个体间、群落间及群体与周围环境的生态学关系[10-14]。干热河谷气候是一类干热少雨的典型非地带性气候[15],成为科研的一个重点方向[16]。对干热河谷地区生物多样性资源保护[17-18]、生境恢复[19-20]、造林[21-22]等领域的研究发现:当前相关成果都集中于干热河谷这一特定小气候区域内[17, 23],割裂了干热河谷区域在同一地理单元中与其他垂直气候区的联系,在生境梯度上进行环境解释、探索形成机制的报道较少。本研究考察了绿汁江流域灌草丛,研究同一地理单元中天然灌草丛群落分布格局及形成机制、变化趋势,在考虑自然地理单元完整性的前提下,探索生态系统的关联性,提出系统修复和综合治理办法,对相互关联的自然生态要素进行整体性保护,实现以山上山下同治带动流域上下游同治,一域一策,以期最终达到流域生态系统全面综合治理。
-
研究区分布于云南省玉溪市峨山县境内的绿汁江流域,海拔为780.0~2 583.7 m;境内河流切割,小地形变化明显,垂直高差大,最低海拔点位于绿汁江边的丫勒河与新平县、双柏县交界处,属典型的干热河谷气候,最高海拔点位于高鲁山主峰西侧的火石山,属南温带气候,区域内年极端最低气温为−8.0 ℃,极端最高气温为33.6 ℃。由于特殊的地形因素和气候因素共同作用,形成了研究区典型的垂直气候分区,具有较丰富的生境多样性,因而生态系统多样性研究意义重大[24-26]。研究区详情见表1。
表 1 研究区主要气候类型及指标
Table 1. Typical climate types and factors
气候类型 海拔/m 年均气温/℃ 年均降
水量/mm年均蒸
发量/mm干燥度
指数分布 干热河谷带 ≤1 500 17~21 828.2 1 908.8 1.4 亚尼村、美党村、江边村等35个村,约5.34 万 hm2 高原面亚热带 1 500~2 000 14~16 934.6 1 869.6 1.2 牛白甸村、棚租村、厂上村等76个村,约12.51 万 hm2 中山南温带 >2 000 12~14 1 000.1 1 815.0 1.1 总果村、高平村、以他斗村等42个村,约1.42 万 hm2 研究区具体灌丛分布按行政区划统计情况见表2。
表 2 研究区灌丛分布统计表
Table 2. Disyribution of shrub communities
行政区划 灌丛图斑/个 面积/hm2 优势灌丛 岔河乡 16 87.0 清香木 Pistacia weinmannifolia 大龙潭乡 92 1 055.9 山矾 Symplocos sumuntia、珍珠花 Lyonia ovalifolia、清香木 甸中镇 118 601.2 华西小石积 Osteomeles schwerinae、清香木、杜鹃 Rhododendron simsii 富良棚乡 64 543.4 萌生栎 Quercus spp.、清香木 化念镇 274 2 307.7 余甘子 Phyllanthus emblica、清香木 双江镇 149 1 308.8 萌生栎、珍珠花 塔甸镇 37 103.3 华西小石积、清香木、牛筋条 Dichotomanthus tristaniaecarpa 小街镇 104 739.8 清香木、萌生栎 -
运用双向指示种分析法(TWINSPAN)和主成分分析法(PCA)对研究区内天然灌草丛进行分类,确定分布格局及环境关系分析。TWINSPAN是一种可以同时完成样地分类和物种分类的双向分类方法,通过把物种和样地排列在同一矩阵中反映物种和样地的相互关系,并且把类型相同(即主要物种和物种重要值相同)的图斑分为一类,便于统计分析。PCA是将多个变量通过线性变换选出较少个数重要变量的一种多元统计分析方法。研究区各类灌草丛的分布情况一般由1个或多个主要因子主导共同作用而形成,涉及多个解释变量(环境因子),因此PCA法可找到主要解释变量,对研究的准确性和科学性至关重要。
-
研究区天然灌草丛图斑数据通过2017年全省森林资源调查获得,调查因子包括共17类响应变量和解释变量[27];其中,响应变量包括灌木种、灌木高度、灌木盖度、草本种、草本高度、草本盖度,解释变量包括郁闭度、坡位、坡向、坡度、岩裸率、土壤亚类、土壤厚度、腐殖质厚度、土壤含石量、灌木分布、海拔[4]。
-
重要值是TWINSPAN植物群落分类的基础数据,被广泛用于植被分类研究[28-30]。陈仲新等[31]和上官铁梁等[32]通过TWINSPAN法对灌草丛群落进行分类。本研究根据森林资源调查数据中各物种的盖度、高度及频度计算出各物种的重要值,计算方法为:
$$ \normalsize{物种重要值=(相对盖度+相对频度+相对高度)/300。} $$ -
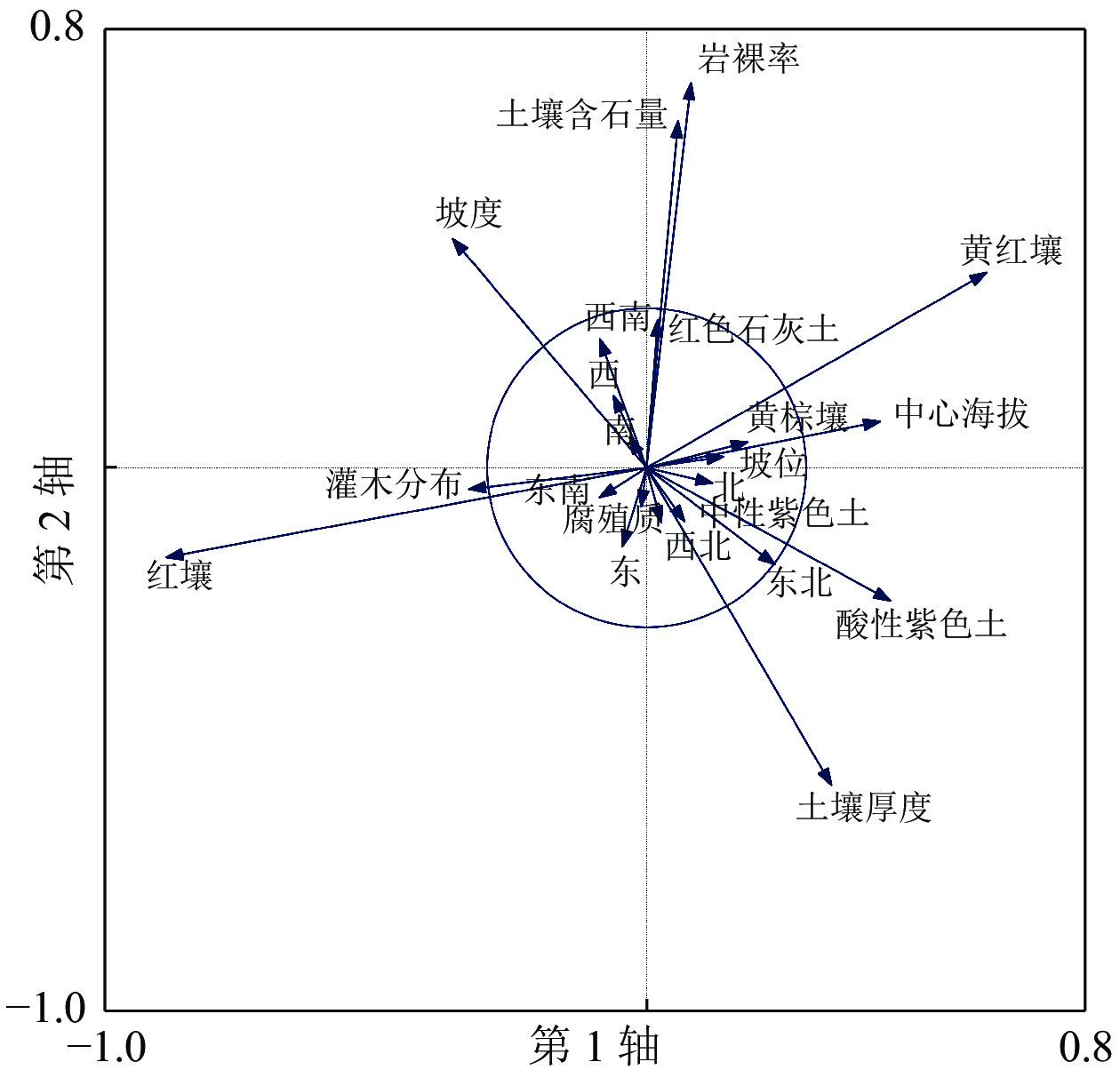
采用PCA获得各解释变量对应前4轴的解释值。从表3可知:当全部解释变量(郁闭度、坡位、坡向、坡度、岩裸率、土壤亚类、土壤厚度、腐殖质厚度、土壤含石量、灌木分布、海拔)代入PCA分析模型时,4轴的解释值仅有30.88%,说明有部分解释变量并不能够很好地解释环境图斑分布与环境关系。借助PCA分析法中的均等贡献(EC)模块对解释变量进行降噪处理[27, 33],以选出最佳解释变量。去除对4轴贡献度较小的值(圈内的变量),及贡献方向相同的较小值(在圈外但贡献方向相同且较短的变量),挑选出坡度、土壤含石量、土壤厚度、海拔作为图斑与环境分布关系的最优解释变量(图1)。降噪处理后的解释变量4轴的解释值达99.98%,即降噪处理后选择的解释变量可以较好地解释环境与图斑的分布关系。
表 3 解释变量的PCA分析
Table 3. Analysis of PCA with explanatory variables
分类轴 全解释变量主成分 优化解释变量主成分 特征值 累积贡献率/% 特征值 累积贡献率/% 轴1 0.095 9 9.59 0.372 9 37.29 轴2 0.081 5 17.74 0.243 7 61.66 轴3 0.069 9 24.93 0.228 7 84.53 轴4 0.061 6 30.88 0.154 5 99.98 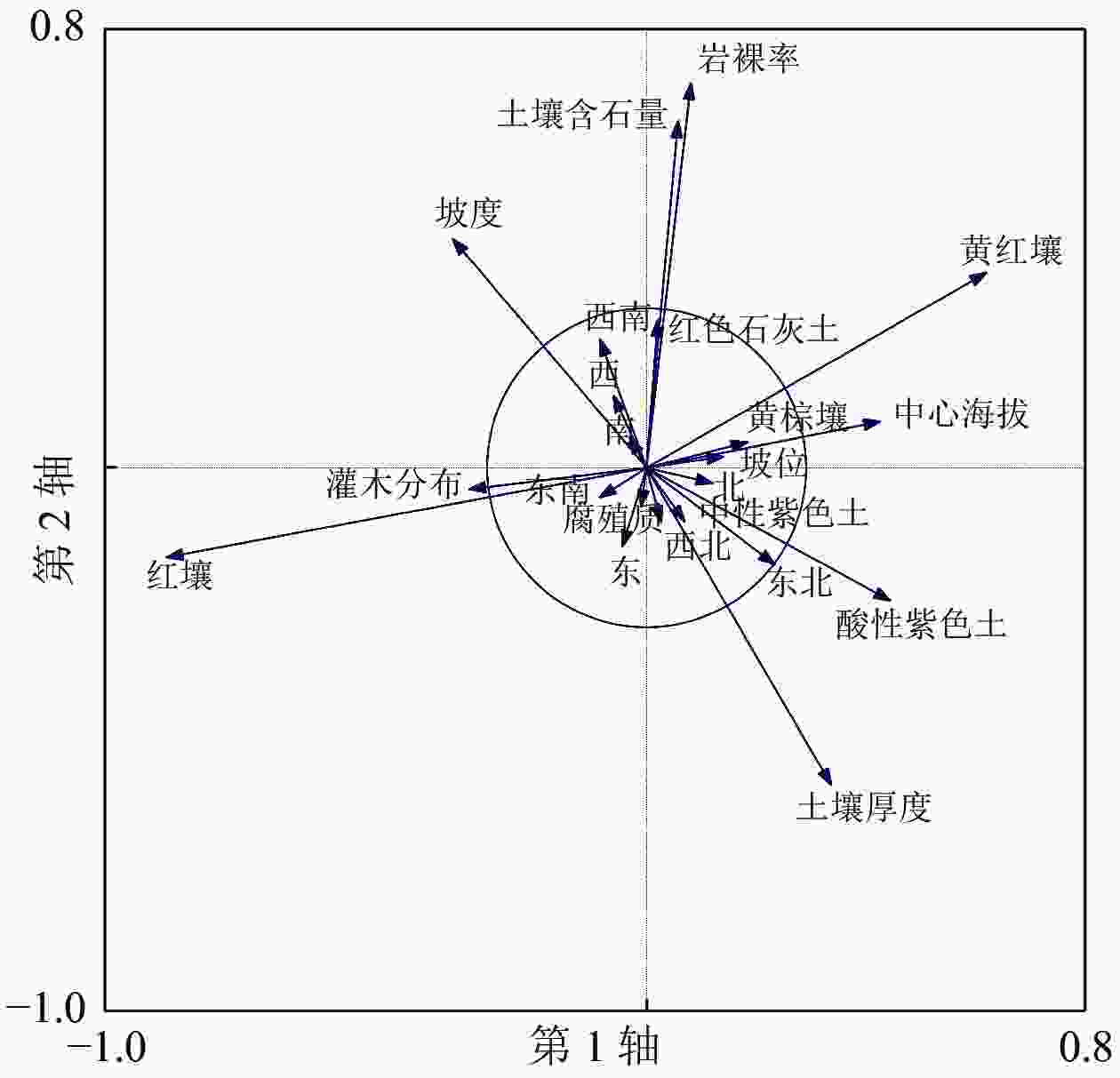
图 1 PCA 全解释变量降噪分析
Figure 1. Noise reduction analysis of PCA with fully explanatory variables
-
采用PC-ORD 5.0和 CANOCO 5.0软件对数据进行处理及分析。
-
利用TWINSPAN将灌木层数据首先进行一次分类,达到划分标准的定义为一个群丛,否则再运用草本数据进行下一级细分。研究区共得到854个灌木图斑,TWINSPAN等级分类从群落外貌尺度出发,将群落自上而下分层分类,并最终定义到群丛[29]。灌草丛群落中灌木层建群种或优势种对群落的结构、功能、外貌特征起到决定性作用,并结合《中国植被》[34]群落命名原则,将具有不同灌木建群种或优势种的图斑直接定义为不同的群丛组,并通过草本层继续细分到群丛。
854个图斑初步可分为云南杨梅Myrica nana、车桑子Dodonaea viscosa、杜鹃、窄叶火棘Pyracantha angustifolia、金丝桃Hypericum monogynum、苦刺Solanum deflexicarpum、山矾、马桑Coriaria nepalensis、萌生栎、珍珠花、牛筋条、清香木、水红木Viburnum cylindricum、算盘子Glochidion puberum、乌鸦果Vaccinium fragile、虾子花Woodfordia fruticosa、华西小石积、红毛悬钩子Rubus pinfaensis、余甘子等19个群丛组,其中云南杨梅、窄叶火棘、金丝桃、算盘子、虾子花等5个群丛组草本层相同,可直接定义到群丛,其他14个群从组根据物种—图斑关系建立数据矩阵,进行下一级划分。以剩余未分类图斑草本物种重要值为基础,按不同类型下的最小样本数(3、4、5、6个),当分类最小样本数为5个、分类的最大水平为4次时群丛分类最为合适,研究区植被共区分出19个群从组65个群丛(表4)。
表 4 灌木群落TWINSPAN分类结果
Table 4. Classification of shrub communities by TWINSPAN
编号 群丛组 群丛类型 小班数/个 1 云南杨梅 云南杨梅-刺芒野古草 Arundinella setosa 3 2 窄叶火棘 窄叶火棘-旱茅 Eremopogon delavayi 1 3 算盘子 算盘子-刺芒野古草 1 4 虾子花 虾子花-鬼针草 Bidens pilosa 1 5 金丝桃 金丝桃-刺芒野古草-莎草 Carex spp. 2 6 车桑子 车桑子-阿拉伯黄背草 Themeda triandra、车桑子-黄茅 Heteropogon contortus-紫荆泽兰 Eupatorium
adenophorum、车桑子-刺芒野古草11 7 苦刺 苦刺-阿拉伯黄背草、苦刺-金茅 Eulalia speciosa、苦刺-刺芒野古草-黄茅 17 8 山矾 山矾-旱茅-莎草、山矾-刺芒野古草-黄茅、山矾-细柄草 Capillipedium parviflorum、山矾-金发草
Pogonatherum paniceum28 9 马桑 马桑-蕨 Pteridium aquilinum、马桑-旱茅-黄茅、马桑-刺芒野古草、马桑-斑茅 Saccharum
arundinaceum35 10 萌生栎 萌生栎-蕨、萌生栎-紫荆泽兰、萌生栎-黄茅、萌生栎-刺芒野古草-旱茅、萌生栎-间型沿阶草
Ophiopogon intermedius-金发草、萌生栎-阿拉伯黄背草、萌生栎-刺芒野古草117 11 牛筋条 牛筋条-阿拉伯黄背草、牛筋条-紫荆泽兰、牛筋条-刺芒野古草-黄茅、牛筋条-刺芒野古草-西南菅
草 Themeda hookeri、牛筋条-香薷 Elsholtzia ciliata17 12 水红木 水红木-刺芒野古草、水红木-蕨、水红木-金发草-阿拉伯黄背草 12 13 乌鸦果 乌鸦果-斑茅、乌鸦果-刺芒野古草-旱茅 13 14 红毛悬钩子 红毛悬钩子-刺芒野古草、红毛悬钩子-蕨 8 15 杜鹃 杜鹃-蕨、杜鹃-刺芒野古草-旱茅、杜鹃-香薷-刺芒野古草、杜鹃-金发草-白茅 Imperata
cylindrica、杜鹃-金发草-紫荆泽兰、杜鹃-细柄草-紫荆泽兰、杜鹃-紫茎泽兰-蒿60 16 珍珠花 珍珠花-黄茅-间型沿阶草、珍珠花-蕨、珍珠花-刺芒野古草、珍珠花-白茅-细柄草、珍珠花-旱茅 90 17 华西小石积 华西小石积-刺芒野古草、华西小石积-黄茅、华西小石积-阿拉伯黄背草、华西小石积-旱茅-金茅 70 18 清香木 清香木-鬼针草、清香木-阿拉伯黄背草、清香木-间型沿阶草-蒿 Artemisia spp.、清香木-金茅-白
茅、清香木-金发草、清香木-蕨、清香木-紫荆泽兰、清香木-刺芒野古草-黄茅281 19 余甘子 余甘子-刺芒野古草-黄茅、余甘子-旱茅-金发草、余甘子-黄茅 87 结合环境因子对群丛组分类结果进行环境解释。
-
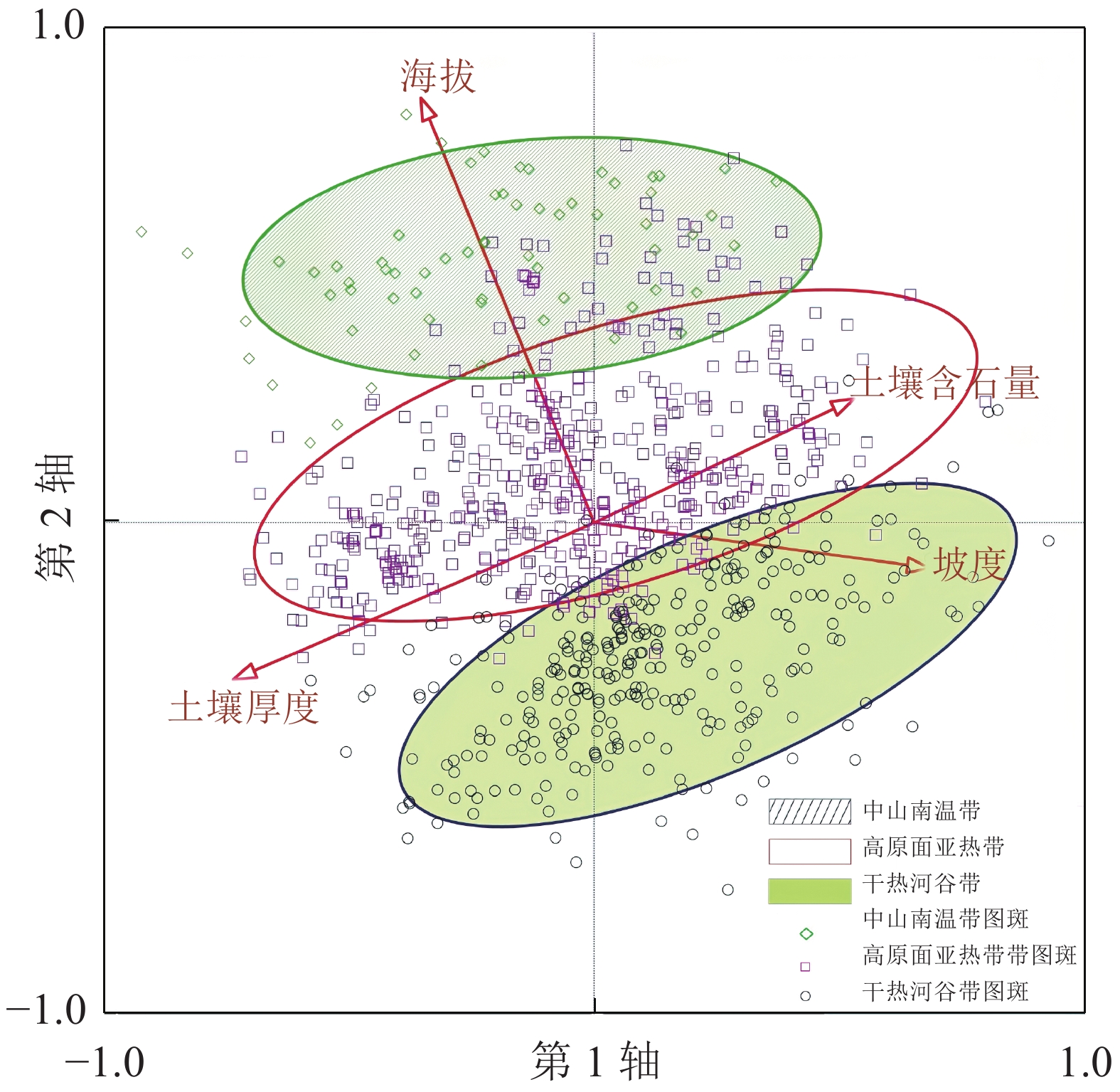
根据PCA对样方的排序,运用envelope工具结合项目区垂直气候情况,将研究区分为3个区,即干热河谷灌草丛区、高原面亚热带灌草丛区、中山南温带灌草丛区(图2)。通过各图斑所分布的海拔区间确定所处垂直气候分区,并通过某灌草丛在该区域出现的次数占总数的百分比确定各分区灌草丛分布频率。研究发现(图3):干热河谷灌草丛区共有307个图斑,涉及13种灌草丛;分布最多的灌草丛类型是清香木灌草丛,占总量的56.02%;较多的是余甘子灌草丛,占总量的17.26%;次多的有萌生栎灌草丛以及山矾灌草丛,共占总量的12.04%;稀少的灌草丛类型包括车桑子灌草丛、虾子花灌草丛等9种灌草丛,共占总量的14.68%。结合表1可知:这些图斑基本分布在绿汁江边,具有典型的干热河谷气候特征,体现了以清香木灌草丛及余甘子灌草丛这类干热河谷指示种占优势的分布格局。高原面亚热带灌草丛区共有481个图斑,涉及18种灌草丛;分布最多的灌草丛类型是清香木灌草丛,占总量的29.73%;较多的是萌生栎灌草丛、珍珠花灌草丛和华西小石积灌草丛,共占总量的40.76%;次多的灌草丛类型是杜鹃灌草丛、马桑灌草丛,共占总量的12.47%;稀少的灌草丛类型包括车桑子灌草丛、余甘子灌草丛等12种灌草丛,共占总量的17.04%。与干热河谷相比,该地区海拔较高,受干热气候影响较小,降水较多;适宜的降水和热量造成了这一地区灌草丛在不同生境轴中的分布也更加均匀,对环境的适应能力有所增强,因此灌草丛种类最为丰富,形成多种灌草丛相对平均的灌草丛分布格局。中山南温带灌草丛区共有66个图斑,涉及8种灌草丛;分布最多的是杜鹃灌草丛,占总量的50.00%;较多的是萌生栎灌草丛、珍珠花灌草丛,共占总量的37.87%;稀少的灌草丛类型包括清香木灌草丛、马桑灌草丛等5类灌草丛,共占总量的12.13%。该地区地处垂直气候带中的南温带,相对亚热带地区更加寒冷,形成了以杜鹃灌草丛、萌生栎灌草丛、珍珠花灌草丛这类相对耐寒的灌草丛景观为主的自然景观格局,同时由于热量降低,灌草丛类型丰富度也有所减少。
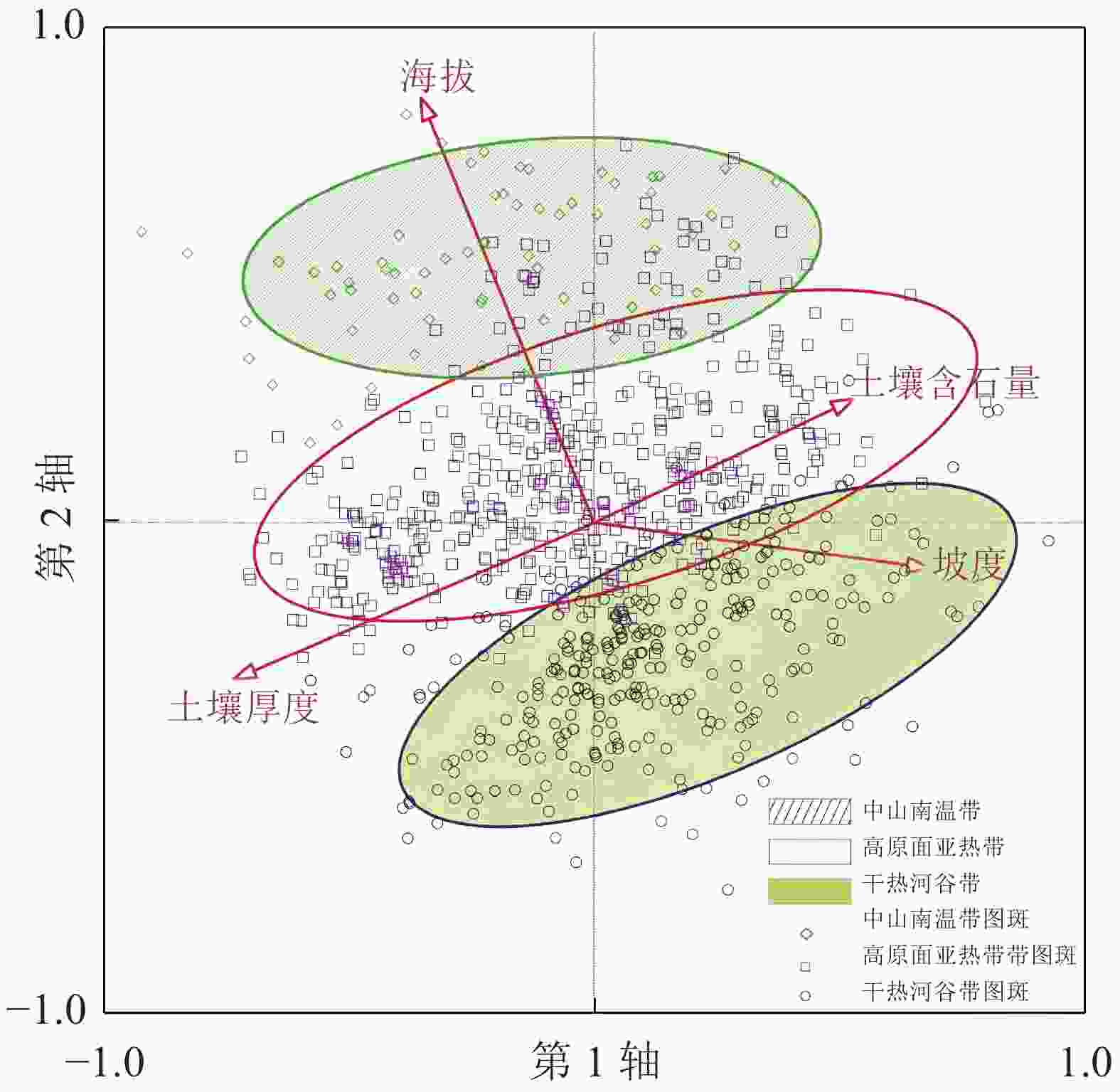
图 2 854个图斑垂直海拔带PCA排序图
Figure 2. PCA ordination of the 854 squares in altitude aspect
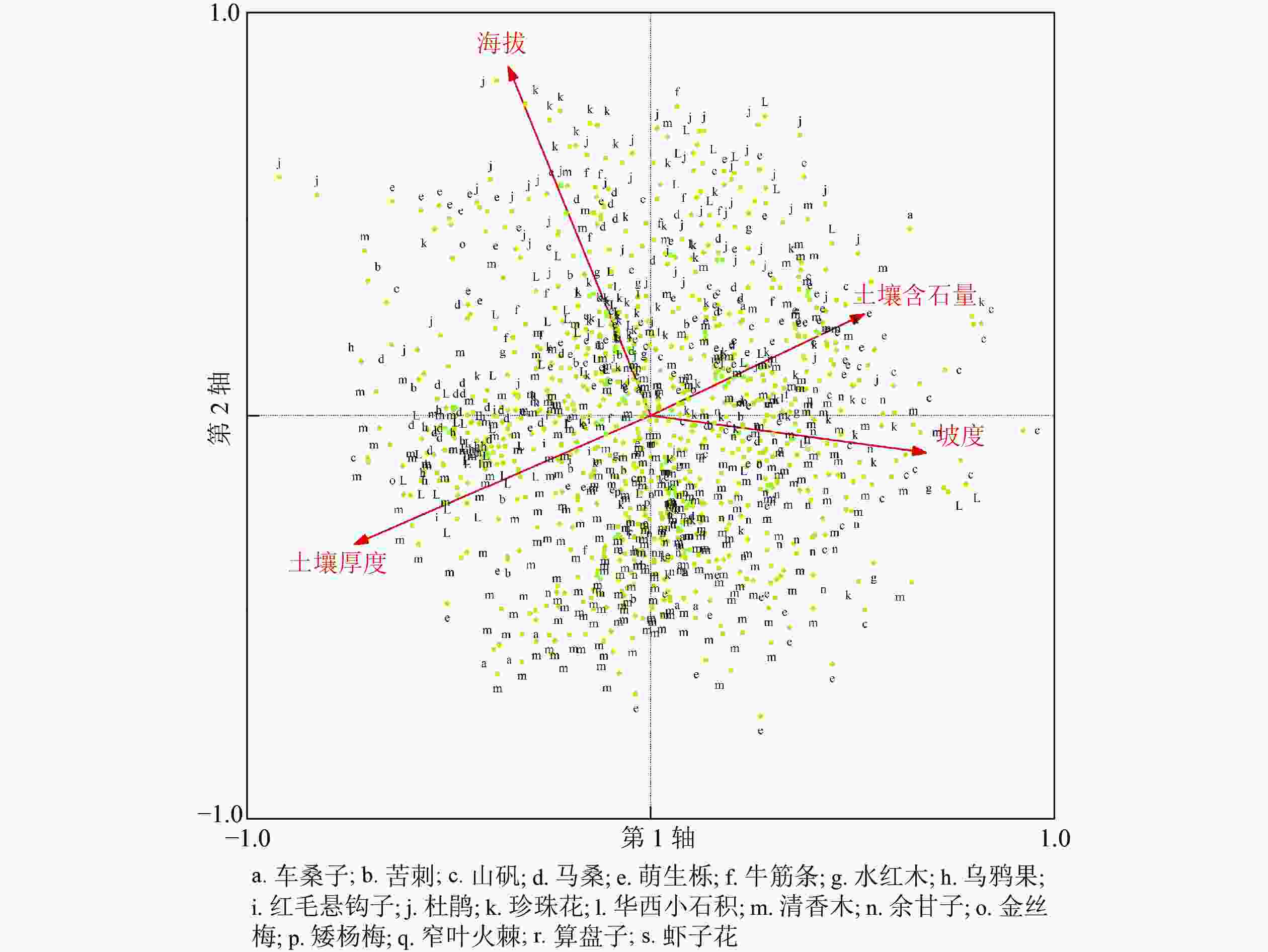
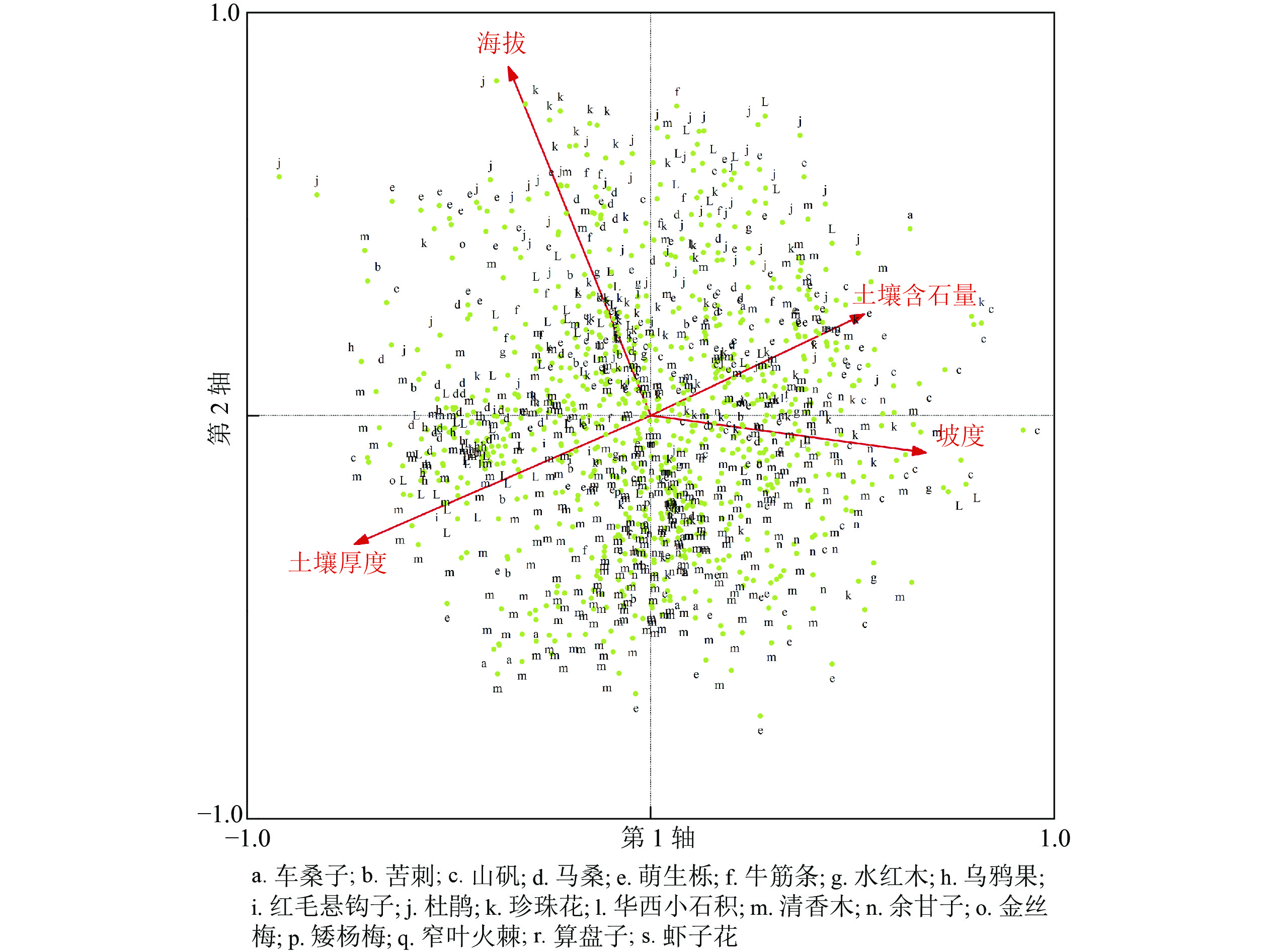
图 3 854个图斑灌丛PCA排序图
Figure 3. PCA ordination of the 854 squares in shrub aspect
-
根据不同类型灌草丛分布频率(表5),结合PCA排序图(图2和图3)可以确定一些灌草丛的分布范围及最适生存区。对出现频次较高的灌草丛的分析可见:华西小石积灌草丛、珍珠花灌草丛、萌生栎灌草丛的适应范围为亚热带和南温带,清香木灌草丛的适应范围为亚热带,余甘子灌草丛的适应范围为干热河谷带,杜鹃灌草丛的适应范围为南温带。坡陡、土壤含石量高、土层较薄的生境条件下,山矾灌草丛、萌生栎灌草丛、华西小石积灌草丛、杜鹃灌草丛出现频率相较其他灌丛更高,提示生境过滤是形成这种分布格局的主要驱动力。乌鸦果灌草丛、红毛悬钩子灌草丛、金丝桃灌草丛、云南杨梅灌草丛、窄叶火棘灌草丛、算盘子灌草丛、虾子花灌草丛属于研究区较少分布的灌草丛。
表 5 不同气候带的灌草丛分布频率
Table 5. Distribution frequency of shrub community
灌草丛 干热河谷带/% 高原面亚热带/% 中山南温带/% 灌草丛 干热河谷带/% 高原面亚热带/% 中山南温带/% 车桑子 2.61 0.63 珍珠花 4.89 10.19 18.18 苦刺 1.63 2.29 1.52 华西小石积 1.95 12.69 3.03 山矾 6.18 1.87 清香木 56.02 29.73 4.55 马桑 0.33 6.86 1.51 余甘子 17.26 2.49 萌生栎 5.86 17.88 19.70 金丝桃 0.21 1.51 牛筋条 0.98 2.91 云南杨梅 0.62 水红木 1.63 1.46 窄叶火棘 0.21 乌鸦果 0.33 2.49 算盘子 0.21 红毛悬钩子 1.66 虾子花 0.33 杜鹃 5.61 50.00 -
利用PCA将TWINSPAN划分的灌草丛类型在二维排序图上进行排序,排序结果较好地揭示了绿汁江下段灌草丛群落的分布格局与环境梯度的关系。第1轴表达了坡度、土壤含石量和土壤厚度方向的综合梯度,主要反映了土层厚度、雨水截留量、保水能力的变化;第2轴表达了海拔的垂直梯度变化,主要反映了干热方向的差异。两者综合作用可表达该区灌草丛群落的空间梯度。结合区域群落空间数据,了解种群生态幅度、最适生存区,为干热河谷地区生态修复,提供本土树种的选择支撑。
在保证完整的地理单元情况下,采用TWINSPAN等级分类,将绿汁江下段灌草丛群落854个图斑划分为19个群丛组65个群丛;由于生境过滤,形成了以绿汁江流域峨山县段为完整地理单元的独特灌草丛生态系统本地参照,以及各类群丛组不同的分布格局和环境间的关联关系。通过参照群丛组环境关联关系可判断生态系统中物种所受到的主要生态胁迫,为系统修复和综合治理提供依据。
由不同灌草丛与环境因子的对应关系发现:土壤瘠薄、含石量高的区域,华西小石积灌草丛和萌生栎灌草丛分布较多。华西小石积灌草丛能够在这些区域集中出现,是因为这些适旱植物具有叶片小、叶片革质、枝条短、根茎比大等形态特征,后期可以选择克土、封育的方式对这些区域进行修复。而萌生栎灌草丛的出现则是因为人为过度干扰,森林遭到破坏后形成次生林,又被再次砍伐,造成森林无法正常发育,从而处于不断萌新状态;加上林地植被减少、长期无法封闭、没有正常的物质循环过程,造成所处立地进一步变差、土壤瘠薄,后期需对这些区域进行封育,减少人类活动的干扰。可见,运用不同植被类型结合关键立地条件因子分析不同的类型形成与修复方案,做到“适地适案”精准施策,可为亚热带干旱森林区的生态修复、保存提供借鉴。
研究区属于亚热带半湿润气候,但由于地处绿汁江河谷,受焚风效应影响较大,植被垂直带谱明显;不同垂直气候带都形成了与环境相对应的典型灌草丛,其中干热河谷带因干热的生境条件形成了以清香木、余甘子为主的灌草丛外貌特征,高原面亚热带灌草丛群落多样性丰富,存在多个共优灌草丛群落,形成了均匀型灌草丛外貌特征,中山南温带由于相对寒冷,形成了以杜鹃为主的灌草丛外貌特征。不同垂直气候带的灌草丛镶嵌分布形成了峨山县绿汁江为中心地区的整体灌草丛景观。后续研究可以根据以绿汁江谷为中心的地理单元的垂直灌草丛分布情况,探索区域内物种分布边界,掌握区流域典型灌草丛外貌分布格局,为绿汁江流域植被的总体保护和小流域恢复提供自然植被本底参考。
Analysis on the distribution pattern and the environmental relationship of natural shrub-grassland communities in the lower section of Lüzhi River
-
摘要:
目的 研究绿汁江下游区域天然灌草丛群落在同一地理单元中的分布格局与环境变化规律,探索群落分布形成机制,找到干热河谷地区生态系统整体性修复的重要挟点,提出系统修复和综合治理办法。 方法 运用双向指示种分析法(TWINSPAN)和主成分分析(PCA),以峨山县绿汁江下段灌草丛群落森林资源调查数据为依据,研究植被分类并分析环境关系。 结果 TWINSPAN数量分类方法显示:854个图斑可划分为19个群丛组65个群丛。PCA排序结果显示:第1轴呈现了土层厚度、雨水截留量、保水能力的变化,第2轴呈现了干热方向的变化。垂直气候梯度灌草丛的丰富度从大到小依次为高原面亚热带、干热河谷带、中山南温带。各垂直气候带中都具有典型指示类型。干扰强度大的地区形成了以华西小石积Osteomeles schwerinae、山矾Symplocos sumuntia、萌生栎Quercus spp.、杜鹃Rhododendron simsii灌草丛为主的分布格局。 结论 生境过滤和人为干扰是灌草丛群落分布格局的主要形成机制。通过分析同一地理单元的天然灌草丛群落分布,可了解种群生态幅度和最适生存区。参照群丛组与环境关联关系可判断生态系统中物种所受到的主要生态胁迫。图3表5参34 -
关键词:
- 灌草丛群落 /
- 分布格局 /
- 双向指示种法(TWINSPAN) /
- 主成分分析(PCA) /
- 干热河谷
Abstract:Objective With an investigation of the distribution patterns of shrub-grassland communities as well as their relationships with the environment in the same geographical unit in the lower section of Lüzhi basin, this study is aimed to explore the formation mechanism of community distribution and then to find out the important pinch point of ecosystem restoration in dry-hot valley area so as to propose methods of systematical restoration and comprehensive management. Method Two-way indicator species analysis (TWINSPAN) and Principle component analysis (PCA) were employed to classify shrub-grassland communities before a proper analysis was conducted of the relationship between the shrub-grassland communities and the environment on the basis of the forest resources investigation data of the shrub-grassland community in the lower section of Lüzhi River in Eshan County. Result The 854 patches were divided into 19 association groups and 65 associations using the TWINSPAN method. In terms of the PCA ordination, the first axis demonstrated the changes in soil depth, rainfall interception and water holding capacity, while the second axis demonstrated the changes in dry, and heat ranges. Under the vertical climate gradient, the richness of shrub-grassland communities was presented in the following pattern: subtropical plateau zone>dry-hot valley zone>south temperate middle mountain zone. Each vertical climate zone was featured with respective typical indicator types of shrub-grassland community and. In the areas with high disturbance intensity, the main distribution patterns were Osteomeles schwerinae, Symplocos sumuntia, Quercus spp., and Rhododendron simsii. Conclusion Habitat filtering and human disturbance are the main formation mechanisms of shrub-grassland community distribution and with an analysis of the distribution of natural shrub-grassland communities in the same geographical unit, the ecological amplitude and the optimal living areas of species populations can be found, which can provide the selections of native tree species for ecological restoration in the dry-hot valley area. Also, the main ecological stresses on the species in the ecosystem can be confirmed in accordance with the relationship between association groups and the environment, thus providing basis and support for systematical restoration and comprehensive management. [Ch, 3 fig. 5 tab. 34 ref.] -
图 1 PCA 全解释变量降噪分析
Figure 1 Noise reduction analysis of PCA with fully explanatory variables
表 1 研究区主要气候类型及指标
Table 1. Typical climate types and factors
气候类型 海拔/m 年均气温/℃ 年均降
水量/mm年均蒸
发量/mm干燥度
指数分布 干热河谷带 ≤1 500 17~21 828.2 1 908.8 1.4 亚尼村、美党村、江边村等35个村,约5.34 万 hm2 高原面亚热带 1 500~2 000 14~16 934.6 1 869.6 1.2 牛白甸村、棚租村、厂上村等76个村,约12.51 万 hm2 中山南温带 >2 000 12~14 1 000.1 1 815.0 1.1 总果村、高平村、以他斗村等42个村,约1.42 万 hm2  下载: 导出CSV
下载: 导出CSV
表 2 研究区灌丛分布统计表
Table 2. Disyribution of shrub communities
行政区划 灌丛图斑/个 面积/hm2 优势灌丛 岔河乡 16 87.0 清香木 Pistacia weinmannifolia 大龙潭乡 92 1 055.9 山矾 Symplocos sumuntia、珍珠花 Lyonia ovalifolia、清香木 甸中镇 118 601.2 华西小石积 Osteomeles schwerinae、清香木、杜鹃 Rhododendron simsii 富良棚乡 64 543.4 萌生栎 Quercus spp.、清香木 化念镇 274 2 307.7 余甘子 Phyllanthus emblica、清香木 双江镇 149 1 308.8 萌生栎、珍珠花 塔甸镇 37 103.3 华西小石积、清香木、牛筋条 Dichotomanthus tristaniaecarpa 小街镇 104 739.8 清香木、萌生栎
下载: 导出CSV
表 3 解释变量的PCA分析
Table 3. Analysis of PCA with explanatory variables
分类轴 全解释变量主成分 优化解释变量主成分 特征值 累积贡献率/% 特征值 累积贡献率/% 轴1 0.095 9 9.59 0.372 9 37.29 轴2 0.081 5 17.74 0.243 7 61.66 轴3 0.069 9 24.93 0.228 7 84.53 轴4 0.061 6 30.88 0.154 5 99.98
下载: 导出CSV
表 4 灌木群落TWINSPAN分类结果
Table 4. Classification of shrub communities by TWINSPAN
编号 群丛组 群丛类型 小班数/个 1 云南杨梅 云南杨梅-刺芒野古草 Arundinella setosa 3 2 窄叶火棘 窄叶火棘-旱茅 Eremopogon delavayi 1 3 算盘子 算盘子-刺芒野古草 1 4 虾子花 虾子花-鬼针草 Bidens pilosa 1 5 金丝桃 金丝桃-刺芒野古草-莎草 Carex spp. 2 6 车桑子 车桑子-阿拉伯黄背草 Themeda triandra、车桑子-黄茅 Heteropogon contortus-紫荆泽兰 Eupatorium
adenophorum、车桑子-刺芒野古草11 7 苦刺 苦刺-阿拉伯黄背草、苦刺-金茅 Eulalia speciosa、苦刺-刺芒野古草-黄茅 17 8 山矾 山矾-旱茅-莎草、山矾-刺芒野古草-黄茅、山矾-细柄草 Capillipedium parviflorum、山矾-金发草
Pogonatherum paniceum28 9 马桑 马桑-蕨 Pteridium aquilinum、马桑-旱茅-黄茅、马桑-刺芒野古草、马桑-斑茅 Saccharum
arundinaceum35 10 萌生栎 萌生栎-蕨、萌生栎-紫荆泽兰、萌生栎-黄茅、萌生栎-刺芒野古草-旱茅、萌生栎-间型沿阶草
Ophiopogon intermedius-金发草、萌生栎-阿拉伯黄背草、萌生栎-刺芒野古草117 11 牛筋条 牛筋条-阿拉伯黄背草、牛筋条-紫荆泽兰、牛筋条-刺芒野古草-黄茅、牛筋条-刺芒野古草-西南菅
草 Themeda hookeri、牛筋条-香薷 Elsholtzia ciliata17 12 水红木 水红木-刺芒野古草、水红木-蕨、水红木-金发草-阿拉伯黄背草 12 13 乌鸦果 乌鸦果-斑茅、乌鸦果-刺芒野古草-旱茅 13 14 红毛悬钩子 红毛悬钩子-刺芒野古草、红毛悬钩子-蕨 8 15 杜鹃 杜鹃-蕨、杜鹃-刺芒野古草-旱茅、杜鹃-香薷-刺芒野古草、杜鹃-金发草-白茅 Imperata
cylindrica、杜鹃-金发草-紫荆泽兰、杜鹃-细柄草-紫荆泽兰、杜鹃-紫茎泽兰-蒿60 16 珍珠花 珍珠花-黄茅-间型沿阶草、珍珠花-蕨、珍珠花-刺芒野古草、珍珠花-白茅-细柄草、珍珠花-旱茅 90 17 华西小石积 华西小石积-刺芒野古草、华西小石积-黄茅、华西小石积-阿拉伯黄背草、华西小石积-旱茅-金茅 70 18 清香木 清香木-鬼针草、清香木-阿拉伯黄背草、清香木-间型沿阶草-蒿 Artemisia spp.、清香木-金茅-白
茅、清香木-金发草、清香木-蕨、清香木-紫荆泽兰、清香木-刺芒野古草-黄茅281 19 余甘子 余甘子-刺芒野古草-黄茅、余甘子-旱茅-金发草、余甘子-黄茅 87
下载: 导出CSV
表 5 不同气候带的灌草丛分布频率
Table 5. Distribution frequency of shrub community
灌草丛 干热河谷带/% 高原面亚热带/% 中山南温带/% 灌草丛 干热河谷带/% 高原面亚热带/% 中山南温带/% 车桑子 2.61 0.63 珍珠花 4.89 10.19 18.18 苦刺 1.63 2.29 1.52 华西小石积 1.95 12.69 3.03 山矾 6.18 1.87 清香木 56.02 29.73 4.55 马桑 0.33 6.86 1.51 余甘子 17.26 2.49 萌生栎 5.86 17.88 19.70 金丝桃 0.21 1.51 牛筋条 0.98 2.91 云南杨梅 0.62 水红木 1.63 1.46 窄叶火棘 0.21 乌鸦果 0.33 2.49 算盘子 0.21 红毛悬钩子 1.66 虾子花 0.33 杜鹃 5.61 50.00
下载: 导出CSV
-
[1] 张峰, 张金屯, 张峰. 历山自然保护区猪尾沟森林群落植被格局及环境解释[J]. 生态学报, 2003, 23(3): 421 − 427. ZHANG Feng, ZHANG Jintun, ZHANG Feng. Pattern of forest vegetation and its environmental interpretation in Zhuweigou, Lishan Mountain Nature Reserve [J]. Acta Ecol Sin, 2003, 23(3): 421 − 427. [2] 刘梦婷, 王振锡, 王雅佩, 等. 新疆天山云杉林群落分布格局及环境解释[J]. 林业科学研究, 2019, 32(6): 90 − 98. LIU Mengting, WANG Zhenxi, WANG Yapei, et al. Plant communities pattern of Picea tianschanica forest and their interrelations with environmental factors in Tianshan Area [J]. For Res, 2019, 32(6): 90 − 98. [3] 杨立荣, 杨小波, 李东海, 等. 海南中部山区什运乡小斑块分布次生林的数量分类[J]. 福建林业科技, 2010, 37(3): 92 − 98. YANG Lirong, YANG Xiaobo, LI Donghai, et al. Classification and ordination of secondary forests Zhayun township in the interior mountainous area of Hainan [J]. J Fujian For Sci Technol, 2010, 37(3): 92 − 98. [4] 王敏, 周才平. 山地植物群落数量分类和排序研究进展[J]. 南京林业大学学报(自然科学版), 2011, 35(4): 126 − 130. WANG Min, ZHOU Caiping. Research progress on quantitative classification and ordination of mountain plant communities [J]. J Nanjing For Univ Nat Sci Ed, 2011, 35(4): 126 − 130. [5] 宋爱云, 刘世荣, 史作民, 等. 卧龙自然保护区亚高山草甸的数量分类与排序[J]. 应用生态学报, 2006, 17(7): 1174 − 1178. SONG Aiyun, LIU Shirong, SHI Zuomin, et al. Quantitative classification and ordination of subalpine meadow in Wolong Nature Reserve [J]. Chin J Appl Ecol, 2006, 17(7): 1174 − 1178. [6] 李素清, 张金屯, 上官铁梁. 芦芽山亚高山草甸的数量分类与排序研究[J]. 西北植物学报, 2005, 25(10): 2062 − 2067. LI Suqing, ZHANG Jintun, SHANGGUAN Tieliang. Quantitative classification and ordination of the subalpine meadows in Luyashan Mountain [J]. Acta Bot Boreali-Occident Sin, 2005, 25(10): 2062 − 2067. [7] 李素清, 张金屯. 山西云顶山亚高山草甸群落生态分析[J]. 地理研究, 2007, 26(1): 83 − 90. LI Suqing, ZHANG Jintun. Ecological analysis of subalpine meadow in Yunding Mountain, Shanxi [J]. Geogr Res, 2007, 26(1): 83 − 90. [8] 席跃翔, 张金屯, 李军玲. 关帝山亚高山灌草丛草甸群落的数量分类与排序研究[J]. 草业学报, 2004, 13(1): 15 − 20. XI Yuexiang, ZHANG Jintun, LI Junling. Quantitative classification and sorting of subalpine shrub and meadow land communities on Guandi Mountain [J]. Acta Pratacult Sin, 2004, 13(1): 15 − 20. [9] 吴华, 张建利, 范怡雯, 等. 草海流域植物群落结构数量分类与排序[J]. 南京林业大学学报(自然科学版), 2013, 37(3): 47 − 52. WU Hua, ZHANG Jianli, FAN Yiwen, et al. Numerical classification and ordination of forest communities in Caohai basin [J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(3): 47 − 52. [10] BURKE A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia [J]. J Veg Sci, 2001, 12(1): 53 − 60. [11] DIEKMANN M, EILERTSEN O, FREMSTAD E, et al. Beech forest communities in the Nordic countries: a multivariate analysis [J]. Plant Ecol, 1999, 140(2): 203 − 220. [12] JONES C C, del MORAL R. Patterns of primary succession on the foreland of Coleman Glacier, Washington, USA [J]. Plant Ecol, 2005, 180(1): 105 − 116. [13] EL-GHANI M M A. Vegetation composition of egyptian inland salt marshes [J]. Bot Bullet Acad Sin, 2000, 41: 305 − 314. [14] 许彬, 张金屯, 杨洪晓, 等. 京西百花山植物群落数量分析[J]. 北京师范大学学报(自然科学版), 2006, 42(1): 90 − 94. XU Bin, ZHANG Jintun, YANG Hongxiao, et al. Quantitative analysis of plant communities in Baihua Mountains, western Beijing [J]. J Beijing Norm Univ Nat Sci, 2006, 42(1): 90 − 94. [15] 曹敏, 金振洲. 云南巧家金沙江干热河谷的植被分类[J]. 云南植物研究, 1989, 11(3): 324 − 336. CAO Min, JIN Zhenzhou. Classification of vegetation in Qiaojia dry-hot river valley of Jinsha River, Yunnan [J]. Acta Bot Yunnan, 1989, 11(3): 324 − 336. [16] 杨济达, 张志明, 沈泽昊, 等. 云南干热河谷植被与环境研究进展[J]. 生物多样性, 2016, 24(4): 462 − 474. YANG Jida, ZHANG Zhiming, SHEN Zehao, et al. Review of research on the vegetation and environment of dry-hot valleys in Yunnan [J]. Biodiversity Sci, 2016, 24(4): 462 − 474. [17] 李新辉, 刘延虹, 刘晔, 等. 地理距离及环境差异对云南元江干热河谷植物群落beta多样性的影响[J]. 生物多样性, 2016, 24(4): 399 − 406. LI Xinhui, LIU Yanhong, LIU Ye, et al. Impacts of geographical distances and environmental differences on the beta diversity of plant communities in the hot-dry valley of the Yuanjiang River [J]. Biodiversity Sci, 2016, 24(4): 399 − 406. [18] 刘晔, 许玥, 石松林, 等. 金沙江干旱河谷植物群落的数量分类及其结构分异的环境解释[J]. 生物多样性, 2016, 24(4): 407 − 420. LIU Ye, XU Yue, SHI Songlin, et al. Quantitative classification and environmental interpretations for the structural differentiation of the plant communities in the dry valley of Jinshajiang River [J]. Biodiversity Sci, 2016, 24(4): 407 − 420. [19] 马焕成, 曾小红. 干旱和干热河谷及其植被恢复[J]. 西南林学院学报, 2005, 25(4): 52 − 55. MA Huancheng, ZENG Xiaohong. Vegetation restoration in the dry and dry-hot valley areas [J]. J Southwest For Coll, 2005, 25(4): 52 − 55. [20] 张金盈, 徐云, 苏春江, 等. 金沙江干热河谷植被恢复研究进展[J]. 水土保持研究, 2005, 12(6): 101 − 104. ZHANG Jinying, XU Yun, SU Chunjiang, et al. Research progress on vegetation restoration in the dry-hot valleys of the Jinsha River [J]. Res Soil Water Conserv, 2005, 12(6): 101 − 104. [21] 李昆, 曾觉民. 元谋干热河谷地区不同造林树种对土壤的改良作用研究[J]. 西南林学院学报, 1999, 19(3): 161 − 164. LI Kun, ZENG Juemin. Effects of some tree species on soil in Yuanmou hot and dry valley [J]. J Southwest For Coll, 1999, 19(3): 161 − 164. [22] 李昆, 张春华, 崔永忠, 等. 金沙江干热河谷区退耕还林适宜造林树种筛选研究[J]. 林业科学研究, 2004, 17(5): 555 − 563. LI Kun, ZHANG Chunhua, CUI Yongzhong, et al. A study on the fitting afforestation tree species during coverting the land for forestry in hot and arid valley of Jinsha River [J]. For Res, 2004, 17(5): 555 − 563. [23] 杨阳, 韩杰, 刘晔, 等. 三江并流地区干旱河谷植物物种多样性海拔梯度格局比较[J]. 生物多样性, 2016, 24(4): 440 − 452. YANG Yang, HAN Jie, LIU Ye, et al. A comparison of the altitudinal patterns in plant species diversity within the dry valleys of the Three Parallel Rivers region, northwestern Yunnan [J]. Biodiversity Sci, 2016, 24(4): 440 − 452. [24] 薛建辉. 森林生态学[M]. 北京: 中国林业出版社, 2006. XUE Jianhui. Forest Ecology[M]. Beijing: China Forestry Press, 2006. [25] 罗建. 色季拉山植物群落数量分析[D]. 拉萨: 西藏大学, 2008. LUO Jian. Quantitative Analysis of Plant Communities in Sejila Mountain, Xizang [D]. Lhasa: Xizang University, 2008. [26] 桂伟峰. 峨山县云南松天然林地位指数表的编制[J]. 林业调查规划, 2019, 44(6): 11 − 14, 33. GEI Weifeng. Establishment of site index table for Pinus yunnanensis natural forest in Eshan County [J]. For Invent Plann, 2019, 44(6): 11 − 14, 33. [27] ŠMILAUER P, LEPŠ J. Multivariate Analysis of Ecological Data using Canoco 5 [M]. Cambridge: Cambridge University Press, 2003. [28] 欧光龙, 彭明春, 和兆荣, 等. 高黎贡山北段植物群落TWINSPAN数量分类研究[J]. 云南植物研究, 2008, 30(6): 679 − 687. OU Guanglong, PENG Mingchun, HE Zhaorong, et al. Quantitative classification of plant communities on the northern part of Gaoligong Mountains by TWINSPAN [J]. Acta Bot Yunnan, 2008, 30(6): 679 − 687. [29] 于梦凡. 植物群丛的数量分类方法及对比研究: 以辽宁青龙河自然保护区为例[D]. 北京: 北京林业大学, 2014. YU Mengfan. Comparative Study on Numerical Classification of Plant Association: A Case Study in Liaoning Qinglong River Natural Reserve [D]. Beijing: Beijing Forestry University, 2014. [30] 刘润红, 涂洪润, 李娇凤, 等. 桂林岩溶石山青冈群落数量分类与排序[J]. 生态学报, 2019, 39(22): 8595 − 8605. LIU Runhong, XU Hongrun, LI Jiaofeng, et al. Numerical classification and ordination of Cyclobalanopsis glauca communities in karst hills of Guilin, Southwest China [J]. Acta Ecol Sin, 2019, 39(22): 8595 − 8605. [31] 陈仲新, 张新时. 毛乌素沙化草地景观生态分类与排序的研究[J]. 植物生态学报, 1996, 20(5): 423 − 437. CHEN Zhongxin, ZHANG Xinshi. The classification and ordination of ecological elements in Maowusu sandy grassland landscape [J]. Acta Phytoecol Sin, 1996, 20(5): 423 − 437. [32] 上官铁梁, 李晶, 李秀兰. 山西北部地区沙棘群落的数量分类和排序研究[J]. 西北植物学报, 2004, 24(8): 1452 − 1456. SHANGGUAN Tieliang, LI Jing, LI Xiulan. Study on classification and ordination of Hippophae rhamnoides subsp. sinensis community in the north of Shanxi [J]. Acta Bot Boreali-Occident Sin, 2004, 24(8): 1452 − 1456. [33] SEABY R, HENDERSON P. Community Analysis Package 4.0 Searching for Structure in Community Data [M]. Lymington: PISCES Conservation Ltd. , 2007. [34] 吴征镒. 中国植被[M]. 北京: 中国科学技术出版社, 1980. WU Zhengyi. Vegetation in China[M]. Beijing: China Science and Technology Press, 1980. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210197
 点击查看大图
点击查看大图
计量
- 文章访问数: 1760
- HTML全文浏览量: 405
- PDF下载量: 47
- 被引次数: 0

