




-
城市绿地是生态城市建设的基础和重要组成部分[1]。植物病害特别是土传病害已经对绿地生态系统健康造成了严重威胁[2-3],由真菌引起的植物病害占全部植物病害的2/3以上[4],其中土传植物病原真菌是造成植物病害的主要类群[5]。研究表明:立枯丝核菌Rhizoctonia solani和齐整小核菌Sclerotium rolfsii是上海绿地土壤中最常见的土传植物病原真菌[6]。立枯丝核菌属于无孢科Agonomycetaceae丝核菌属Rhizoctonia[7],在自然界中广泛存在,是引起植物根腐和茎腐症状的重要病原菌,被认为是最具破坏力的土传植物病原菌之一[8]。海桐Pittosporum tobira、八角金盘Fatsia japonica、高羊茅Festuca arundinacea和女贞Ligustrum lucidum等360个属的园林绿化植物均能被立枯丝核菌感染[9]。齐整小核菌属于无孢科小核菌属Sclerotium[6],是一种世界性传播的土传病原真菌,可以引起植物白绢病,致使植物腐烂死亡[10]。木棉属Bombax和紫檀属Pterocarpus植物以及樟树Cinnamomum camphora和夹竹桃Nerium oleander等园林绿化植物常被齐整小核菌侵染[11]。
土传病原真菌的传统检测方法主要是根据病原菌形态特征、选择性平板计数等方法进行[12-13],但耗时费力、效率较低,时效性较差,已不能满足实际工作需求。随着分子生物学技术的发展,实时荧光定量PCR (qPCR)因具有特异性强、灵敏度高等特点,已经越来越多地被应用于土传病原真菌的检测。目前,已经有应用该技术对农田土壤中立枯丝核菌和齐整小核菌进行定性、定量检测的报道[14-16]。但是,尚缺乏应用qPCR技术对城市绿地土壤进行立枯丝核菌和齐整小核菌定量检测的研究。与农田土壤存在明显差异,城市绿地土壤异质性高,普遍面临着重金属和有机污染物等的威胁[1, 17],因此,针对城市绿地土壤中立枯丝核菌和齐整小核菌,探索建立具有较强特异性、较高灵敏性和较好重复性的分子生物学检测方法,对于及时了解上海城市绿地土壤中立枯丝核菌和齐整小核菌的分布,保障植物和城市绿地生态系统健康具有重要意义。
-
立枯丝核菌和齐整小核菌标准菌株购买于中国农业微生物菌种保藏管理中心(Agricultural Culture Collection of China, ACCC),编号分别为ACCC37615和ACCC37946。菌株的培养参照《丝状真菌分子细胞生物学与实验技术》[18]。
-
2018年7月,采用多点混合采样法,在上海市随机采集绿地0~20 cm表层土壤样品。采样点信息见表1。每个采样点设置5个子样点,每个土壤样品由5个子样点样品混合而成。土壤样品充分混合后,去除石块、草根等杂物,−20 ℃保存,用于后续试验[17]。
表 1 绿地土壤样品信息
Table 1. Information of green space soil samples
样品编号 绿地类型 采样位置 行政区 绿地名称 纬度(N) 经度(E) 1 道路绿地 金山区 朱平公路中央绿化带内 30°49′ 121°08′ 2 公园绿地 闵行区 闵行文化公园 31°10′ 121°21′ 3 公园绿地 金山区 亭林公园 30°20′ 121°18′ 4 道路绿地 金山区 亭卫公路 30°20′ 121°20′ 5 公园绿地 奉贤区 奉浦四季生态园 30°56′ 121°27′ 6 公园绿地 崇明区 崇明新城公园 31°37′ 121°25′ 7 公园绿地 黄浦区 复兴公园 31°13′ 121°27′ 8 道路绿地 徐汇区 虹梅路 31°08′ 121°24′ 9 道路绿地 普陀区 真北路 31°16′ 121°23′ 10 道路绿地 浦东新区 杨高中路 31°13′ 121°32′ 11 道路绿地 浦东新区 杨高北路 31°15′ 121°35′ 12 道路绿地 浦东新区 济阳路上浦路 31°08′ 121°29′ 13 道路绿地 浦东新区 杨高南路 31°11′ 121°31′ 14 道路绿地 青浦区 外青松公路 31°12′ 121°07′ 15 道路绿地 浦东新区 唐镇路 31°12′ 121°38′ 16 公园绿地 宝山区 炮台湾湿地公园 31°23′ 121°30′ 17 公园绿地 宝山区 淞南公园 31°20′ 121°28′ 18 道路绿地 闵行区 华宁路剑川路 31°01′ 121°23′ 19 道路绿地 杨浦区 殷行路 31°19′ 121°31′ 20 公园绿地 浦东新区 名人苑公园北门东侧 31°14′ 121°33′ 21 公园绿地 嘉定区 汇龙潭公园 31°22′ 121°14′ -
采用AntGene DNA提取试剂盒 (武汉安特捷生物技术有限公司)提取立枯丝核菌和齐整小核菌标准菌株DNA。采用土壤基因组DNA提取试剂盒(FastDNA spin kit for soil, MP Biomedicals, 美国)提取绿地土壤样品DNA。使用NanoDrop 2000微量分光光度计(Thermo Fisher Scientific, 美国)测定DNA的吸光度比值D(260)/D(280)及D(260)/D(230),确定DNA质量及浓度,之后于−80 ℃超低温冰箱保存。
-
通过文献检索,根据已有报道选择了ST-RS1/ ITS4、ITS1/GMRS-3等引物进行扩增立枯丝核菌的特异性引物的筛选,选择SCR-F/SCR-R、SRITSF/SRITSR等引物进行扩增齐整小核菌的特异性引物的筛选(表2),引物由生工生物工程(上海)股份有限公司合成。通过美国国家生物技术信息中心(NCBI,
https://www.ncbi.nlm.nih.gov/blast/ )中Primer-BLAST功能将4对引物分别与Non-Redundant Protein Sequence (NR)数据库进行序列比对,在线筛选和验证引物的特异性。表 2 用于立枯丝核菌和齐整小核菌qPCR的引物信息
Table 2. qPCR primer sets of R. solani and S. rolfsii
病原菌 引物名称 引物序列 (5ʹ→3ʹ) 产物大小/bp 退火温度/℃ 参考文献 立枯丝核菌 ST-RS1 AGTGTTATGCTTGGTTCCACT 187 60 [19] ITS4 TCCTCCGCTTATTGATATGC ITS1 TCCGTAGGTGAACCTGCGG 564 65 [20] GMRS-3 AGTGGAACCAAGCATAACACT 齐整小核菌 SCR-F CGTAGGTGAACCTGCGGA 540 54 [21] SCR-R CATACAAGCTAGAATCCC SRITSF TACACCTGTGAACCAACTG 465 52 [22] SRITSR CATACAAGCTAGAATCCC -
以2种病原菌标准菌株DNA为模板,分别使用筛选的特异性引物对其进行PCR扩增。PCR产物经质量分数为2%琼脂糖凝胶电泳后切胶回收目的条带,使用通用型DNA纯化试剂盒[天根生化科技(北京) 有限公司,中国北京]对目的条带进行纯化回收。按照pGM-T Fast连接试剂盒[天根生化科技(北京)有限公司,中国北京]说明书将回收所得片段连接到pGEM-T Easy载体上(Promega, 美国),转化至JM109感受态细胞中(Promega, 美国),经蓝白斑筛选阳性克隆子,并进行菌落PCR验证后,按照质粒抽提试剂盒[天根生化科技(北京)有限公司,中国北京]说明书进行质粒抽提,提取所得重组质粒送至上海美吉生物医药科技有限公司测序。使用NanoDrop 2000 微量分光光度计(Thermo Fisher Scientific, 美国)测定质粒标准品浓度,计算质粒拷贝数[23]。
-
qPCR反应体系包含有10 μL GoTaq q-PCR Master Mix (Promega,美国),DNA模板用量分别设置为0.5、1.0、2.0、3.0、4.0 μL,上、下游引物(10 μmol·L−1)分别设置为0.1、0.3、0.5、0.7、0.9 μL,加无菌重蒸水补足至20 μL。阴性对照使用无菌重蒸水代替DNA模板。使用Roche Light Cycle 480Ⅱ型qPCR仪(罗氏医学仪器公司,瑞士)。qPCR反应程序:95 ℃ 预变性5 min;95 ℃变性 15 s,退火温度下退火40 s [退火温度在文献中引物Tm值(表2)上下浮动3 ℃范围内设置7个温度梯度],72 ℃延伸40 s,45个循环。循环结束后进入熔解曲线程序:65 ℃升温至95 ℃,隔1 ℃采集荧光信号。
-
将立枯丝核菌和齐整小核菌质粒标准品分别进行10倍梯度稀释,质粒标准品浓度范围为108~101 (×106拷贝·L−1),分别得到8个稀释浓度的质粒标准品模板,使用1.6.1中优化后的qPCR反应条件进行扩增,绘制标准曲线。
-
以质粒标准品和绿地土壤样品DNA为模板进行qPCR扩增,通过熔解曲线判断qPCR扩增产物是否单一。qPCR扩增产物利用质量分数为2%琼脂糖凝胶电泳(110 V, 40 min)检测,通过紫外凝胶成像系统对电泳条带进行观察和分析。将质粒标准品和土壤样品扩增产物送至上海美吉生物医药科技有限公司进行克隆测序,并利用NCBI数据库对质粒标准品和土壤样品PCR扩增产物测序结果进行序列比对。
-
分别将2种病原真菌的质粒标准品进行10倍梯度稀释,从108 ×106拷贝·L−1稀释至101×106拷贝·L−1,取稀释好的质粒标准品作为模板,按照1.6中优化后的qPCR条件进行扩增,根据试验结果确定方法的最低检测限,验证此方法的灵敏性。
-
随机选择5个不同类型绿地土壤样品DNA作为模板,按照1.6中优化后的方法进行立枯丝核菌和齐整小核菌qPCR扩增,每个反应重复2次。随后根据每个样品组内重复间拷贝数的变异系数,验证方法的重复性和稳定性。
-
采用1.6中优化后的qPCR方法对21份上海绿地土壤样品进行立枯丝核菌和齐整小核菌定量检测,每个反应设置3个平行试验,计算土壤样品中病原菌ITS基因的拷贝数,y=c×n×v/s。其中:y为基因拷贝数(拷贝·g−1),c为浓度(×106拷贝·L−1),n为模板稀释倍数,v为DNA洗脱体积(μL),s为土壤干质量(g)。
-
运用Excel 2016软件对数据进行分析处理。
-
使用NCBI中Primer-BLAST功能将表2的4对引物分别与NR数据库进行序列比对。结果显示:引物ST-RS1/ITS4和ITS1/GMRS-3均可有效匹配目的序列900余条,错配率分别为7.7%和6.3%,在理论上均可实现对立枯丝核菌种水平特异性扩增(表3)。与引物ITS1/GMRS-3相比,引物ST-RS1/ITS4扩增目的片段长度小于200 bp,更适用于在qPCR方法中的应用,因此,本研究仅保留引物ST-RS1/ITS4用于后续立枯丝核菌定量检测方法的建立。引物SRITSF/SRITSR可有效匹配目的序列429条,错配率为8.1%,理论上可以实现对齐整小核菌种水平特异性扩增,而引物SCR-F/SCR-R仅匹配30条目的序列,覆盖率相对低于引物SRITSF/SRITSR,且该引物错配率较高,因此,本研究仅保留引物SRITSF/SRITSR用于后续齐整小核菌定量检测方法的建立(表3)。
表 3 qPCR引物在线比对结果
Table 3. On-line blast results of qPCR primers
引物名称 目的序列
长度/bp匹配目的
序列数/条匹配非目的
序列数/条错配
率/%ST-RS1/ITS4 187 908 76 7.7 ITS1/GMRS-3 564 936 63 6.3 SRITSF/SRITSR 465 429 38 8.1 SCR-F/SCR-R 540 30 4 13.3 -
以2种病原菌质粒标准品为模板进行qPCR反应条件的优化。结果表明:引物ST-RS1/ITS4最佳退火温度为60 ℃,引物SRITSF/SRITSR最佳退火温度为52 ℃。在上述退火温度下,2种病原菌qPCR扩增效率最高。因此,最终确定立枯丝核菌qPCR扩增条件为:95 ℃预变性5 min;95 ℃变性15 s,60 ℃退火40 s,72 ℃延伸40 s,45个循环。齐整小核菌qPCR扩增条件为:95 ℃预变性5 min;95 ℃变性15 s,52 ℃退火40 s,72 ℃延伸40 s,45个循环。
2种病原菌qPCR反应体系中最适引物用量均为0.3 μL (10 μmol·L−1),DNA模板用量均为2 μL (10 mg·L−1),在该反应条件下荧光信号最强。因此,最终确定2种病原菌qPCR反应体系(20 μL)为:模板2 μL(10 mg·L−1),上下游引物(10 μmol·L−1)各0.3 μL,GoTaq®q-PCR Master Mix (Promega,美国)10 μL,无菌重蒸水7.4 μL。
-
以10倍梯度稀释的质粒标准品为模板,使用2.2.1优化后的qPCR反应条件进行扩增,绘制标准曲线。立枯丝核菌标准曲线线性关系表达式为y=−3.319x+33.98 (y为Ct值,x为模板量),决定系数(R2)为0.999 5,扩增效率为100.1%。齐整小核菌标准曲线线性关系表达式为y=−3.552x+33.65 (y为Ct值,x为模板量),决定系数(R2)为0.997 9,扩增效率为91.2%。结果表明:2种病原菌qPCR 的Ct值与质粒标准品浓度的对数均具有良好的线性关系,扩增效率符合qPCR定量检测方法的要求,表明本研究建立的2种病原菌定量检测方法具有较高的准确度和有效性,可作为立枯丝核菌和齐整小核菌快速定量检测的方法。对立枯丝核菌的线性检测范围为2.42×101~2.42×108 (×106拷贝·L−1),对齐整小核菌的线性检测范围为2.20×101~2.20×108 (×106拷贝·L−1)。
-
以质粒标准品和绿地土壤样品DNA为模板进行qPCR扩增。结果显示:熔解曲线均为单一熔解峰,无杂峰,表明qPCR扩增产物单一且未产生引物二聚体。qPCR扩增产物琼脂糖凝胶电泳结果显示:引物ST-RS1/ITS4扩增得到约200 bp的片段,引物SRITSF/SRITSR扩增得到约450 bp的片段,均与目的片段大小一致(图1)。利用NCBI数据库对PCR扩增产物克隆测序的结果进行序列比对,结果见表4。采用引物ST-RS1/ITS4从质粒标准品及土壤样品扩增到的产物分别与立枯丝核菌R. solani BTRFB1(登录号: MZ158299)和R. solani AG-F(登录号:JQ343829)序列相似度最高,相似度为99.4%。采用引物SRITSF/SRITSR从质粒标准品及土壤样品扩增到的产物分别与齐整小核菌S. rolfsii NTNM(登录号: MT126473)和S. rolfsii SR1(登录号: MG847186)序列相似度最高,分别为99.5%和98.8%。扩增产物测序结果说明:利用引物ST-RS1/ITS4和SRITSF/SRITSR扩增到的DNA序列均与目的片段序列一致,具有较强的特异性。
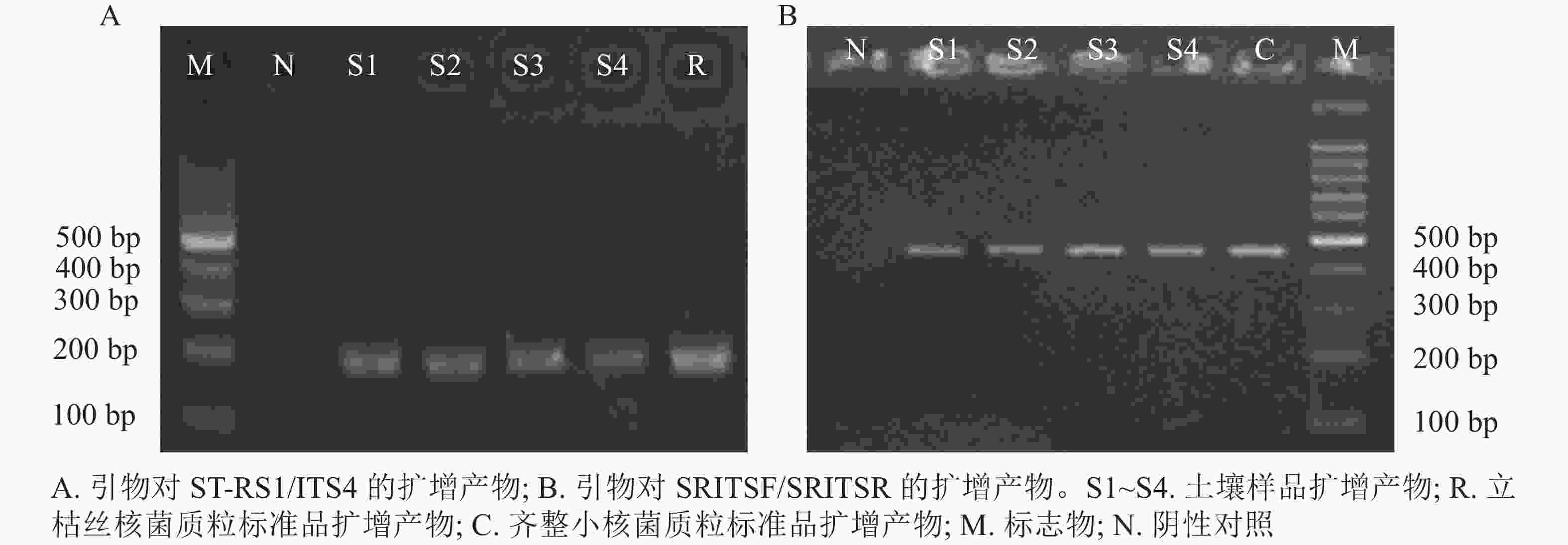
图 1 立枯丝核菌和齐整小核菌 DNA的PCR扩增
Figure 1. PCR amplification of R. solani and S. rolfsii DNA
-
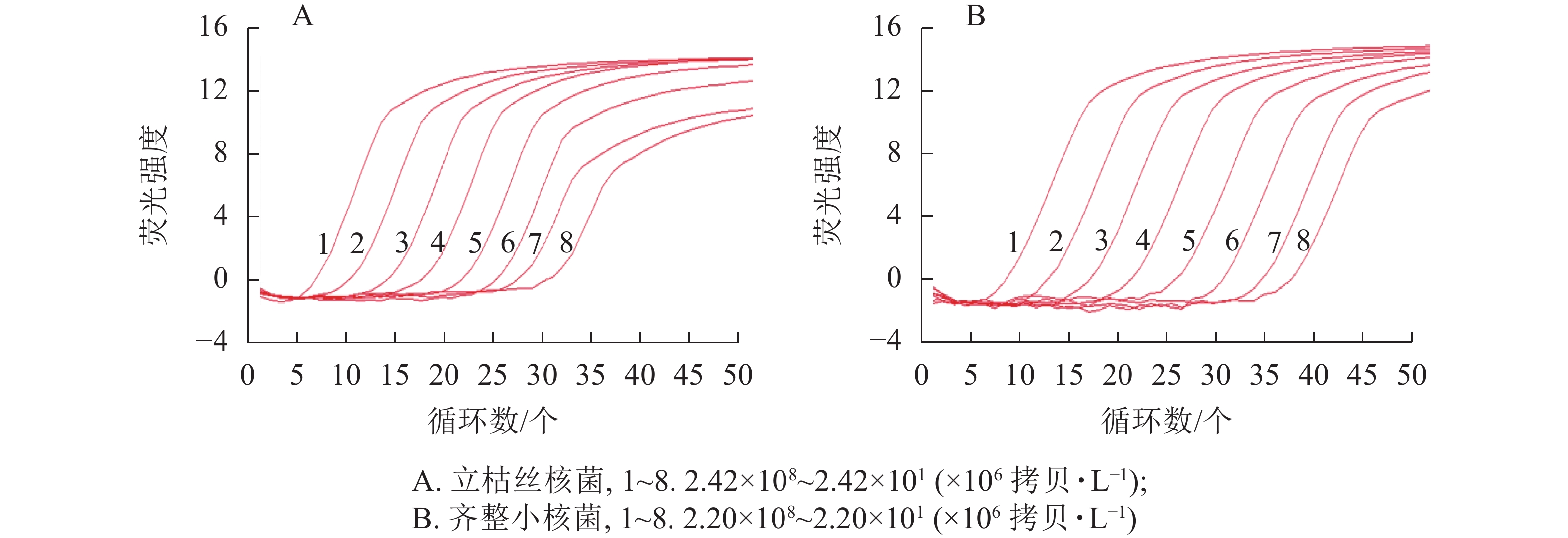
以10倍梯度稀释的质粒标准品为模板,使用2.2.1优化后的qPCR反应条件进行扩增,进行qPCR灵敏性验证。结果显示:本研究建立的qPCR定量检测方法对立枯丝核菌和齐整小核菌的检测限分别可达24×106和22×106 拷贝·L−1 (图2)。
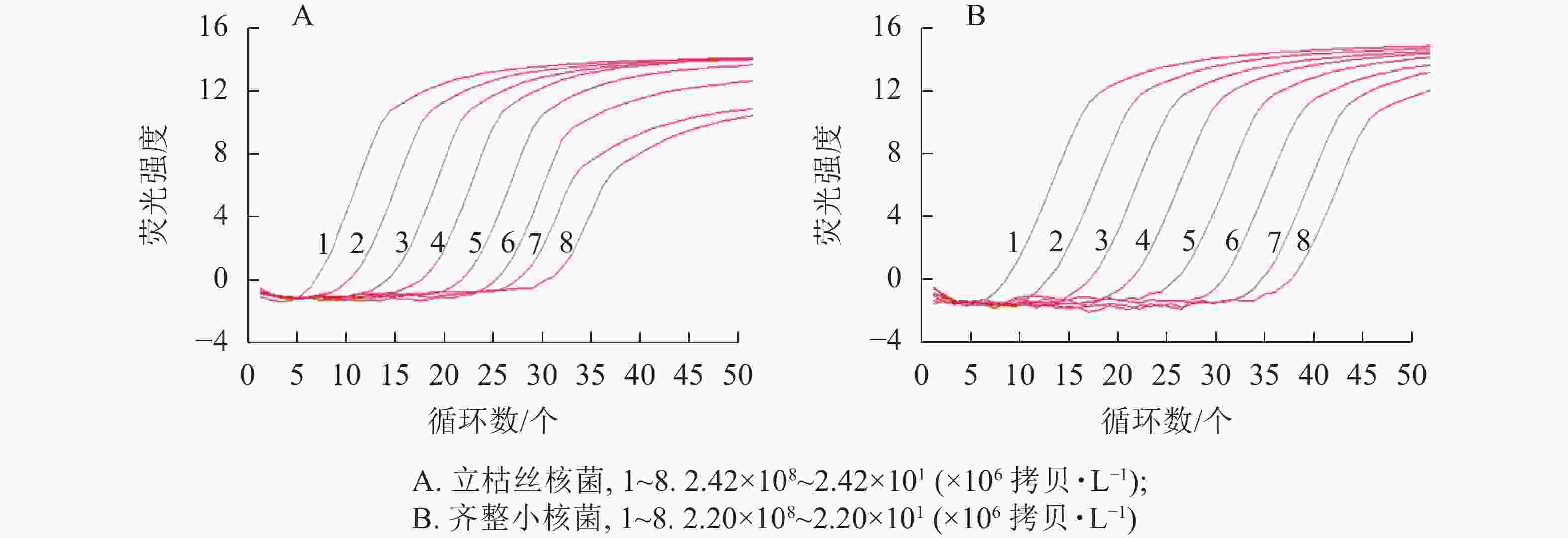
图 2 qPCR的灵敏性
Figure 2. Sensitivity test of qPCR
表 4 质粒标准品和土壤样品测序比对结果
Table 4. Sequencing alignment result of plasmid standard and soil sample
引物 样品 同源菌株 基因登录号 相似度/% ST-RS1/ITS4 质粒标准品 R. solani BTRFB1 MZ158299 99.4 土壤样品 R. solani AG-F JQ343829 99.4 SRITSF/SRITSR 质粒标准品 S. rolfsii NTNM MT126473 99.5 土壤样品 S. rolfsii SR1 MG847186 98.8 -
随机选择5个绿地土壤样品DNA作为模板,使用本研究建立的方法对样品中立枯丝核菌和齐整小核菌ITS基因拷贝数进行测定。结果表明:绿地土壤样品立枯丝核菌和齐整小核菌ITS基因拷贝数2次检测结果的变异系数范围分别为3.37%~4.61%和0.66%~8.61%(表5),变异系数均低于10%,表明本研究建立的立枯丝核菌和齐整小核菌定量检测方法具有较好的重复性和稳定性。
表 5 qPCR的重复性
Table 5. Repeatability of qPCR
病原菌 样品编号 第1次试验 第2次试验 变异系数/% Ct 拷贝数/(×106拷贝·L−1) Ct 拷贝数/(×106拷贝·L−1) 立枯丝核菌 4 31.74 3.09×101 31.60 3.25×101 3.57 5 30.54 6.45×101 30.56 6.15×101 3.37 6 29.49 1.23×102 29.51 1.17×102 3.54 10 30.98 4.91×101 31.03 4.60×101 4.61 12 29.71 1.07×102 29.74 1.01×102 4.08 齐整小核菌 3 33.76 2.16×102 33.53 2.44×102 8.61 6 35.15 1.02×102 35.09 1.05×102 2.05 14 35.04 1.08×102 35.05 1.07×102 0.66 16 36.55 4.77×101 36.63 4.57×101 3.03 18 35.72 7.46×101 35.67 7.66×101 1.87 -
将本研究建立的立枯丝核菌和齐整小核菌定量检测方法应用于上海绿地土壤样品的检测。检测结果表明:立枯丝核菌的检出率为100%, ITS基因拷贝数最高为2.61×106 拷贝·g−1;齐整小核菌的检出率为19%,检出样品的ITS基因拷贝数最高为2.94×105 拷贝·g−1。说明立枯丝核菌在上海绿地表层土壤中广泛存在。
-
引物的特异性是对土传病原真菌进行qPCR检测的关键[24]。真菌ITS序列目前已被广泛用于真菌的亲缘关系分析,为病原菌的分子检测提供了较为理想的分子标记[25]。本研究通过检索文献[19, 22]及数据库,筛选得到了基于真菌ITS区域设计的立枯丝核菌和齐整小核菌种水平特异性引物ST-RS1/ITS4和SRITSF/SRITSR。NCBI NR数据库在线比对结果显示:ST-RS1/ITS4和SRITSF/SRITSR目的片段错配率分别为7.7%和8.1%,理论上可实现对立枯丝核菌和齐整小核菌种水平特异性扩增;同时,qPCR熔解曲线均为单一熔解峰,扩增产物片段大小与序列均与目的片段一致。上述结果均表明:本研究建立的立枯丝核菌和齐整小核菌荧光定量检测方法具有较强的特异性。此外,qPCR检测方法的灵敏性是评价其是否可行的另一个关键因素,一般是指可以准确测量的样品最小拷贝数[26]。前人的研究表明:对农田土壤立枯丝核菌和大豆茎褐腐病菌Phialophora gregata等土传病原菌qPCR检测的灵敏性可以达26.7~100.0 (×106拷贝·L−1)[13, 27-28]。本研究针对立枯丝核菌和齐整小核菌检测限分别可达24×106和22×106拷贝·L−1,具有较高的灵敏性。因此,本研究所建立的qPCR检测方法可以用于城市绿地土壤中立枯丝核菌和齐整小核菌的定量检测。
立枯丝核菌和齐整小核菌是寄主范围广、危害性强的土传病原真菌,可侵染一些重要经济植物,例如水稻Oryza sativa、花生Arachis hypogaea、樟树和海桐等,造成植物根腐病和白绢病等病害的发生[7, 10]。鉴于这2种病原菌分布的广泛性,因此有必要了解其在城市绿地土壤中的分布情况,从而有效预防园林植物立枯病、根腐病和白绢病等病害的发生。本研究中上海绿地表层土壤立枯丝核菌的检出率达100%,表明立枯丝核菌在上海绿地土壤中广泛存在。齐整小核菌检出率较低,为19%。齐整小核菌检出率低,可能有两方面的原因。一方面,随着对城市绿地养护技术和措施的不断提高和加强,齐整小核菌在土壤中的存在范围可能缩小了;另一方面,也可能是由于采样点的局限性,还不足以反映上海绿地土壤中齐整小核菌整体的存在情况,后期需要通过扩大绿地土壤样品采集范围,更为全面地检测齐整小核菌在上海绿地土壤中的分布情况。同时应该看到,应用qPCR方法检测土壤中病原菌的数量时,并不能有效区分活体和死体病原菌的DNA,检测结果可能会过高估计土壤中病原菌基因的丰度[29]。因此,如何准确、快速测定城市绿地土壤中活体立枯丝核菌和齐整小核菌基因的丰度,尚需进一步研究。
综上所述,本研究通过立枯丝核菌和齐整小核菌qPCR引物的筛选和特异性验证、qPCR反应条件的建立及灵敏性和重复性评价,建立了针对立枯丝核菌和齐整小核菌的快速、特异且灵敏性较高的qPCR检测方法,可用于城市绿地土壤中这2种病原菌的定量检测。
Quantitative real-time PCR for rapid detection of Rhizoctonia solani and Sclerotium rolfsii in urban green space
-
摘要:
目的 土传病原真菌立枯丝核菌Rhizoctonia solani和齐整小核菌Sclerotium rolfsii严重威胁园林绿化植物正常生长。建立针对这2种土壤病原真菌的快速定量检测方法。 方法 通过筛选2种病原菌特异性引物,优化反应条件。 结果 初步建立了2种病原菌的实时荧光定量PCR(qPCR)检测方法。引物ST-RS1/ITS4和SRITSF/SRITSR可以分别用于立枯丝核菌和齐整小核菌的qPCR检测,其灵敏度分别达24×106和22×106拷贝·L−1,2次重复反应的变异系数分别为3.37%~4.61%和0.66%~8.61%。对上海绿地土壤样品的检测结果表明:立枯丝核菌和齐整小核菌的检出率分别为100%和19%。 结论 建立的qPCR检测方法具有较强特异性、较高灵敏度和较强重复性,可以用于上海城市绿地土壤中立枯丝核菌和齐整小核菌的快速、有效定量检测。图2表5参29 Abstract:Objective This study aims to establish a rapid quantitative detection method for Rhizoctonia solani and Sclerotium rolfsii, 2 soil-borne pathogenic fungi that seriously threaten the normal growth of landscape plants in Shanghai. Method The reaction conditions were optimized by screening 2 pathogen specific primers. Result A quantitative real-time PCR (qPCR) method was established for the detection of the two pathogens. The primers ST-RS1/ITS4 and SRITSF/SRITSR could be used for qPCR detection of R. solani and S. rolfsii, with sensitivity of 24×106 and 22×106 copies·L−1, respectively. The coefficients of variation of the 2 repeated reactions were 3.37%−4.61% and 0.66%−8.61%, respectively. The detection results of soil samples in Shanghai green space showed that the detection rates of R. solani and S. rolfsii were 100% and 19%, respectively. Conclusion The established qPCR method has high specificity, sensitivity and repeatability, and can be used for rapid and effective quantitative detection of R. solani and S. rolfsii in Shanghai urban green soil. [Ch, 2 fig. 5 tab. 29 ref.] -
Key words:
- urban green space soil /
- Rhizoctonia solani /
- Sclerotium rolfsii /
- qPCR
-
表 1 绿地土壤样品信息
Table 1. Information of green space soil samples
样品编号 绿地类型 采样位置 行政区 绿地名称 纬度(N) 经度(E) 1 道路绿地 金山区 朱平公路中央绿化带内 30°49′ 121°08′ 2 公园绿地 闵行区 闵行文化公园 31°10′ 121°21′ 3 公园绿地 金山区 亭林公园 30°20′ 121°18′ 4 道路绿地 金山区 亭卫公路 30°20′ 121°20′ 5 公园绿地 奉贤区 奉浦四季生态园 30°56′ 121°27′ 6 公园绿地 崇明区 崇明新城公园 31°37′ 121°25′ 7 公园绿地 黄浦区 复兴公园 31°13′ 121°27′ 8 道路绿地 徐汇区 虹梅路 31°08′ 121°24′ 9 道路绿地 普陀区 真北路 31°16′ 121°23′ 10 道路绿地 浦东新区 杨高中路 31°13′ 121°32′ 11 道路绿地 浦东新区 杨高北路 31°15′ 121°35′ 12 道路绿地 浦东新区 济阳路上浦路 31°08′ 121°29′ 13 道路绿地 浦东新区 杨高南路 31°11′ 121°31′ 14 道路绿地 青浦区 外青松公路 31°12′ 121°07′ 15 道路绿地 浦东新区 唐镇路 31°12′ 121°38′ 16 公园绿地 宝山区 炮台湾湿地公园 31°23′ 121°30′ 17 公园绿地 宝山区 淞南公园 31°20′ 121°28′ 18 道路绿地 闵行区 华宁路剑川路 31°01′ 121°23′ 19 道路绿地 杨浦区 殷行路 31°19′ 121°31′ 20 公园绿地 浦东新区 名人苑公园北门东侧 31°14′ 121°33′ 21 公园绿地 嘉定区 汇龙潭公园 31°22′ 121°14′  下载: 导出CSV
下载: 导出CSV
表 2 用于立枯丝核菌和齐整小核菌qPCR的引物信息
Table 2. qPCR primer sets of R. solani and S. rolfsii
病原菌 引物名称 引物序列 (5ʹ→3ʹ) 产物大小/bp 退火温度/℃ 参考文献 立枯丝核菌 ST-RS1 AGTGTTATGCTTGGTTCCACT 187 60 [19] ITS4 TCCTCCGCTTATTGATATGC ITS1 TCCGTAGGTGAACCTGCGG 564 65 [20] GMRS-3 AGTGGAACCAAGCATAACACT 齐整小核菌 SCR-F CGTAGGTGAACCTGCGGA 540 54 [21] SCR-R CATACAAGCTAGAATCCC SRITSF TACACCTGTGAACCAACTG 465 52 [22] SRITSR CATACAAGCTAGAATCCC
下载: 导出CSV
表 3 qPCR引物在线比对结果
Table 3. On-line blast results of qPCR primers
引物名称 目的序列
长度/bp匹配目的
序列数/条匹配非目的
序列数/条错配
率/%ST-RS1/ITS4 187 908 76 7.7 ITS1/GMRS-3 564 936 63 6.3 SRITSF/SRITSR 465 429 38 8.1 SCR-F/SCR-R 540 30 4 13.3
下载: 导出CSV
表 4 质粒标准品和土壤样品测序比对结果
Table 4. Sequencing alignment result of plasmid standard and soil sample
引物 样品 同源菌株 基因登录号 相似度/% ST-RS1/ITS4 质粒标准品 R. solani BTRFB1 MZ158299 99.4 土壤样品 R. solani AG-F JQ343829 99.4 SRITSF/SRITSR 质粒标准品 S. rolfsii NTNM MT126473 99.5 土壤样品 S. rolfsii SR1 MG847186 98.8
下载: 导出CSV
表 5 qPCR的重复性
Table 5. Repeatability of qPCR
病原菌 样品编号 第1次试验 第2次试验 变异系数/% Ct 拷贝数/(×106拷贝·L−1) Ct 拷贝数/(×106拷贝·L−1) 立枯丝核菌 4 31.74 3.09×101 31.60 3.25×101 3.57 5 30.54 6.45×101 30.56 6.15×101 3.37 6 29.49 1.23×102 29.51 1.17×102 3.54 10 30.98 4.91×101 31.03 4.60×101 4.61 12 29.71 1.07×102 29.74 1.01×102 4.08 齐整小核菌 3 33.76 2.16×102 33.53 2.44×102 8.61 6 35.15 1.02×102 35.09 1.05×102 2.05 14 35.04 1.08×102 35.05 1.07×102 0.66 16 36.55 4.77×101 36.63 4.57×101 3.03 18 35.72 7.46×101 35.67 7.66×101 1.87
下载: 导出CSV
-
[1] 马想, 张浪, 黄绍敏, 等. 上海城市绿地土壤肥力变化分析[J]. 中国园林, 2020, 36(5): 104 − 109. MA Xiang, ZHANG Lang, HUANG Shaomin, et al. Analysis on the change of soil fertility in Shanghai urban space [J]. Chin Landscape Archit, 2020, 36(5): 104 − 109. [2] 李世东, 缪作清, 高卫东. 我国农林园艺作物土传病害发生和防治现状及对策分析[J]. 中国生物防治学报, 2011, 27(4): 433 − 440. LI Shidong, LIAO Zuoqing, GAO Weidong. Challenges, opportunities and obligations in management of soilborne plant diseases in China [J]. ChinJ Biol Control, 2011, 27(4): 433 − 440. [3] 付先惠, 曹敏, 唐勇. 植物病原菌在森林动态中的作用[J]. 生态学杂志, 2003, 22(3): 59 − 64. FU Xianhui, CAO Min, TANG Yong. Effects of plant pathogens on forest dynamics [J]. Chin J Ecol, 2003, 22(3): 59 − 64. [4] 彭丹丹, 张源明, 舒灿伟, 等. 植物病原真菌分子检测技术的研究进展[J]. 基因组学与应用生物学, 2017, 36(5): 2015 − 2022. PENG Dandan, ZHANG Yuanming, SHU Canwei, et al. Research progress on the molecular detection techniques of plant pathogenic fungi [J]. Genomics Appl Biol, 2017, 36(5): 2015 − 2022. [5] 杨珍, 戴传超, 王兴祥, 等. 作物土传真菌病害发生的根际微生物机制研究进展[J]. 土壤学报, 2019, 56(1): 12 − 22. YANG Zhen, DAI Chuanchao, WANG Xingxiang, et al. Advance in research on rhizosphere microbial mechanisms of crop soil-borne fungal diseases [J]. Acta Pedol Sin, 2019, 56(1): 12 − 22. [6] 蒋杰贤, 严巍. 城市绿地有害生物预警及控制[M]. 上海: 上海科学技术出版社, 2007: 207 − 225. JIANG Jiexian, YAN Wei. Pest Warning and Control in Urban Green Space[M]. Shanghai: Shanghai Scientific and Technical Press, 2007: 207 − 225. [7] 骆丹, 田慧, 张彩霞, 等. 植物立枯丝核菌根腐病研究进展[J]. 中国植保导刊, 2020, 40(3): 23 − 31. LUO Dan, TIAN Hui, ZHANG Caixia, et al. Advances in the research on plant root rot caused by Rhizoctonia solani [J]. China Plant Prot, 2020, 40(3): 23 − 31. [8] ANDERSON N A. The genetics and pathology of Rhizoctonia solani [J]. Ann Rev Phytopathol, 2003, 20(1): 329 − 347. [9] FARR D F, BILLS G F, CHAMURIS G P, et al. Fungi on plants and plant products in the United States [J]. Mycologia, 1990, 82(3): 243 − 246. [10] BILLAH K M M, HOSSAIN M B, PRINCE M H, et al. Pathogenicity of Sclerotium rolfsii on different host, and its over wintering survival: a mini review [J]. Int J Adv Agric Sci, 2017, 2(7): 1 − 6. [11] FLORENCE E J M, SHARMA J K, SANKARAN K V, et al. Some diseases of forest tree seedlings in India caused by Sclerotium rolfsii and Rhizoctonia solani [J]. Eur J For Pathol, 1985, 15(3): 187 − 190. [12] 曹月霞. 尖孢镰刀菌西瓜专化型分子检测技术的建立及其效应子的初步鉴定[D]. 北京: 中国农业科学院, 2016. CAO Yuexia. The Establishment of the Molecular Detection Technique and Basic Identification of Effectors of Fusarium oxysporum f. sp. niveum[D]. Beijing: Chinese Academy of Agricultural Sciences, 2016. [13] 申永铭, 郭成瑾, 王喜刚, 等. 土壤中立枯丝核菌AG3菌核的荧光定量PCR快速检测[J]. 菌物学报, 2017, 36(10): 1383 − 1391. SHEN Yongming, GUO Chengjin, WANG Xigang, et al. Rapid detection of Rhizoctonia solani AG3 sclerotia in soil by quantitative real-time PCR [J]. Mycosystema, 2017, 36(10): 1383 − 1391. [14] OKUBARA P A, SCHROEDER K L, PAULITZ T C. Identification and quantification of Rhizoctonia solani and R. oryzae using real-time polymerase chain reaction [J]. Phytopathology, 2008, 98(7): 837 − 847. [15] MILNER H, JI Pingsheng, SABULA M, et al. Quantitative polymerase chain reaction (Q-PCR) and fluorescent in situ hybridization (FISH) detection of soilborne pathogen Sclerotium rolfsii [J]. Appl Soil Ecol, 2019, 136: 86 − 92. [16] GAO Yuhan, WU Miao, GUO Rongjun, et al. Real time PCR quantification of Sclerotium rolfsii in chilli tissue and soil [J]. Plant Prot Sci, 2015, 51(2): 61 − 66. [17] 骆玉珍, 张维维, 李雅颖, 等. 上海市公园绿地土壤肥力特征分析与综合评价[J]. 中国土壤与肥料, 2019(6): 86 − 93. LUO Yuzhen, ZHANG Weiwei, LI Yaying, et al. Analysis and comprehensive evaluation of soil fertility characteristics for the urban park in Shanghai [J]. Soil Fert Sci China, 2019(6): 86 − 93. [18] 林福呈, 王洪凯. 丝状真菌分子细胞生物学与实验技术[M]. 北京: 科学出版社, 2010. LIN Fucheng, WANG Hongkai. Molecular Cell Biology and Experimental Techniques of Filamentous Fungi [M]. Beijing: Science Press, 2010. [19] LIEVENS B, BROUWER M, VANACHTER A, et al. Real-time PCR for detection and quantification of fungal and oomycete tomato pathogens in plant and soil samples [J]. Plant Sci, 2006, 171(1): 155 − 165. [20] JOHANSON A, TURNER H C, MCKAY G J, et al. A PCR-based method to distinguish fungi of the rice sheath-blight complex, Rhizoctonia solani, R. oryzae and R. oryzae-sativae [J]. FEMS Microbiol Lett, 2010, 162(2): 289 − 294. [21] JEEVA M L, MISHRA A K, VIDYADHARAN P, et al. A species-specific polymerase chain reaction assay for rapid and sensitive detection of Sclerotium rolfsii [J]. Australas Plant Pathol, 2010, 39(6): 517 − 523. [22] PRAVI V, JEEVA M L, ARCHANA P V. Rapid and sensitive detection of Sclerotium rolfsii associated with collar rot disease of Amorphophallus paeoniifolius by species-specific polymerase chain reaction assay [J]. Mol Biotechnol, 2014, 56(9): 787 − 794. [23] 汪最, 李丽, 刘鹏, 等. ALV-J、REV和MDV多重荧光定量PCR检测方法的建立[J]. 中国动物传染病学报, 2020, 28(6): 73 − 78. WANG Zui, LI Li, LIU Peng, et al. Establishment of a triple real-time PCR detection ALV-J, REV and MDV [J]. Chin J Anim Infect Dis, 2020, 28(6): 73 − 78. [24] 张敏, 杨丹娇, 蓝岚, 等. 羊口疮病毒SYBR Green Ⅰ实时荧光定量PCR法的建立[J]. 四川畜牧兽医, 2021, 48(2): 25 − 28. ZHANG Min, YANG Danjiao, LAN Lan, et al. Establishment of SYBR GreenⅠreal-time fluorescence quantitative PCR method for sheep mouth virus [J]. Sichuan Anim Vet Sci, 2021, 48(2): 25 − 28. [25] 赵爽. 土壤病原真菌的检测及土壤中真菌群落多样性的研究 [D]. 南京: 南京农业大学, 2012. ZHAO Shuang. Soil Pathogens Detection and Soil Fungi Distribution in Soil[D]. Nanjing: Nanjing Agricultural University, 2012. [26] BUSTIN S A, BENES V, GARSON J A, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments [J]. Clin Chem, 2009, 55(4): 611 − 622. [27] CARRILLO J, MAYORQUIN J S, STAJICH J E, et al. Probe-based multiplex real-time PCR as a diagnostic tool to distinguish distinct fungal symbionts associated with Euwallacea kuroshio and E. whitfordiodendrus in California [J]. Plant Dis, 2019, 104(1): 227 − 238. [28] MALVICK D K, IMPULLITTI A E. Detection and quantification of Phialophora gregata in soybean and soil samples with a quantitative, real-time PCR assay [J]. Plant Dis, 2007, 91(6): 736 − 742. [29] 魏锋, 余真真, 商文静, 等. 土壤大丽轮枝菌微菌核的快速定量检测[J]. 植物病理学报, 2013, 43(5): 449 − 457. WEI Feng, YU Zhenzhen, SHANG Wenjing, et al. Rapid detection and quantification of Verticillium dahliae microsclerotia in soil [J]. Acta Phytopathol Sin, 2013, 43(5): 449 − 457. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210801
 点击查看大图
点击查看大图

计量
- 文章访问数: 1560
- HTML全文浏览量: 377
- PDF下载量: 73
- 被引次数: 0
