




-
蒙古栎Quercus mongolica为壳斗科Fagaceae栎属Quercus植物,广泛分布于寒温带、温带和暖温带地区[1]。距今2 000~7 000 a,以栎为主的栎类阔叶林和草甸是河北省平原地区的原生植被[2]。蒙古栎是重要的用材[3]、柞蚕[1, 4]、染料、栲胶[5]、食品、药用[6]、酿酒原料和景观树种,也是营造防风林、水源涵养林及防火林的优良树种,具有十分重要的经济、文化和生态功能。到目前为止,蒙古栎大多数处于野生状态,良种水平较为低下,良种选育研究仅限于对天然群体变异选种或以用材等为目的的种源试验研究阶段,而以结实为目的的良种选育研究尚未开始。因此,加强良种选育工作,提高蒙古栎的良种化水平迫在眉睫。
种子表型性状作为重要的经济性状[7],是遗传多样性的重要基础[8]。因此,对种子遗传变异进行分析显得尤为重要[9]。通过表型性状分析种质资源的遗传多样性直观易行,能够快速了解植物的遗传变异水平[10]。目前,针对东北三省(黑龙江省、辽宁省、吉林省)[11-12]及其他个别地区[13-14]蒙古栎不同种源的苗木生长情况、种子性状的分析研究较多,但对于全国蒙古栎不同种源的种子特性及环境因子的分析较少。本研究对全国蒙古栎天然分布区进行调查收集,了解现有蒙古栎的自然分布状况,采集种子作为种质资源保存,并对各种源收集的蒙古栎种子进行表型性状测定和统计分析,以期为蒙古栎种质资源收集、杂交育种及品种改良提供基础数据。
-
于2020年8—11月蒙古栎种子成熟期,在全国蒙古栎天然分布区调查并收集8省12个种源的蒙古栎种质资源,种源确定依据河北省洪崖山国有林场2018—2019年的全国蒙古栎种质资源踏查结果,结合厉月桥[15]关于蒙古栎分布的研究,选择蒙古栎主要天然分布区且具有优良林分的蒙古栎天然林,所选种源基本涵盖了蒙古栎主要天然分布区,可代表全国蒙古栎分布情况。所选采种林分为当地分布的天然林分,林龄>30 a,每个种源选取30株以上采种母树,母树间距50 m以上,收集的种子均已成熟饱满,无病虫害。蒙古栎不同种源的主要生态因子见表1。
表 1 不同种源蒙古栎主要生态因子
Table 1. Main ecological factors of Q. mongolica from different provenances
种源编号 种源地 北纬(N) 东经(E) 海拔/m 年均气温/℃ 年降水量/mm 无霜期/d 1 黑龙江省黑河市 49°49′18″ 127°16′48″ 294 −1.0 525 110 2 辽宁省抚顺市 41°56′33″ 125°14′24″ 673 5.1 863 130 3 辽宁省本溪市 40°40′05″ 124°00′55″ 487 6.5 926 170 4 河北省张家口市 39°53′36″ 114°57′17″ 1 573 6.0 400 152 5 河北省承德市围场县 42°10′21″ 117°15′31″ 1 330 3.3 370 80 6 河北省承德市兴隆县 40°33′34″ 117°30′00″ 1 092 7.6 763 130 7 河北省秦皇岛市 40°08′33″ 119°25′28″ 1 110 8.9 741 180 8 北京市门头沟区 39°59′01″ 115°26′28″ 1 270 6.4 420 150 9 吉林省延边市 43°11′32″ 129°32′20″ 370 3.9 580 146 10 内蒙古赤峰市 42°06′57″ 118°19′36″ 1 175 7.7 559 157 11 山东省泰安市 53°27′05″ 117°16′20″ 868 13.0 750 200 -
8—9月,选取健壮植株,挂牌标记,拍摄植株照片,进行蒙古栎种质资源踏查,并准确记录采种地的经纬度、海拔及年均气温等;9—11月,收集挂牌标记且具代表性的蒙古栎植株种子。
-
分别对每个种源所采集的蒙古栎单株种子等量混合后,采用四分法随机选取300粒种子进行种长、种宽及种子单粒重、千粒重的测定,用电子游标卡尺进行蒙古栎种宽和种长的测定,精确到0.01 mm,用电子天平进行种子单粒重的测定,用百粒法测量种子千粒重,精确到0.01 g。
-
用Excel对测量的基础数据进行整理和统计;种形指数为种子的宽(直径)和长(纵径)之比[16];运用SPSS 11.5进行方差分析、相关分析(Pearson相关)、主成分分析;同时运用SPSS 11.5将原始数据进行标准化转换,采用欧式距离法进行聚类分析。
-
如表2所示:不同种源蒙古栎种长、种宽差异较大,种长为11.38~20.96 mm,种宽为11.07~17.80 mm。其中辽宁省抚顺市蒙古栎种子的种长和种宽最长,均值分别为20.96、17.80 mm,其种宽、单粒重显著高于其他地区(P<0.05);内蒙古赤峰市最低,种长和种宽均值分别为11.38、11.07 mm,显著低于其他地区(P<0.05)。河北省承德市围场县与兴隆县蒙古栎种子的种长、种宽无显著差异。种形指数均存在一定差异,但差异较小,其中河北省秦皇岛市最高,山东省泰安市最低,均值分别为1.28、1.02。
表 2 不同种源蒙古栎种子表型性状差异性分析
Table 2. Difference analysis of seeds phenotypic characters in Q. mongolica from different provenances
种源地 种长/mm 种宽/mm 种形指数 单粒重/g 千粒重/g 黑龙江省黑河市 20.94±0.38 a 16.58±0.44 bc 1.26±0.04 ab 2.93±0.07 ab 2 949.41±9.65 b 辽宁省抚顺市 20.96±0.73 a 17.80±0.71 a 1.18±0.01 ab 3.05±0.16 a 3 027.39±7.74 a 辽宁省本溪市 18.37±0.46 bc 15.46±0.64 d 1.20±0.02 ab 2.83±0.02 b 2 878.82±7.03 c 河北省张家口市 16.47±0.51 de 13.11±0.68 f 1.26±0.10 ab 1.82±0.10 e 1 824.11±9.20 h 河北省承德市围场县 16.49±0.50 de 13.94±0.23 ef 1.17±0.12 ab 1.41±0.07 f 1 482.71±1.36 j 河北省承德市兴隆县 16.18±0.22 e 14.01±0.49 ef 1.15±0.01 abc 1.79±0.07 e 1 765.37±4.14 i 河北省秦皇岛市 18.22±0.88 c 14.29±0.23 e 1.28±0.04 a 2.07±0.06 d 2 075.77±2.53 f 北京市门头沟区 19.38±1.19 b 16.02±0.97 cd 1.21±0.10 ab 2.31±0.11 c 2 310.59±7.58 e 吉林省延边市 19.39±0.23 b 17.26±0.19 ab 1.12±0.02 bcd 2.45±0.07 c 2 421.51±2.44 d 内蒙古赤峰市 11.38±0.38 f 11.07±0.81 g 1.03±0.09 cd 1.22±0.09 g 1 244.40±5.51 k 山东省泰安市 17.40±1.04 cd 17.10±0.42 abc 1.02±0.09 d 1.92±0.03 de 1 926.28±1.40 g 说明:同列不同字母表示不同种源地之间差异显著(P<0.05) 单粒重和千粒重差异较大,其中各种源千粒重之间均存在显著差异(P<0.05),最高为辽宁省抚顺市(3 027.39 g),最低为内蒙古赤峰市(1 244.40 g),可以看出千粒重的变异幅度较大。
-
如表3所示:不同种源蒙古栎种子表型性状存在一定的相关性。其中蒙古栎种子的种长、种宽、单粒重、千粒重相互之间均存在极显著正相关(P<0.01),即种长越长,种宽越宽,单粒重和千粒重随着种长、种宽的增大而增加,表明这4个表型性状是相互影响的。种形指数与种长、种宽、单粒重和千粒重的相关性呈不显著正相关。
表 3 不同种源蒙古栎种子表型性状的相关性
Table 3. Correlation of seeds phenotypic characters in Q. mongolica from different provenances
性状 种长 种宽 种形指数 单粒重 千粒重 种长 1.00 0.88** 0.52 0.88** 0.88** 种宽 1.00 0.06 0.79** 0.78** 种形指数 1.00 0.40 0.41 单粒重 1.00 1.00** 说明:**表示极显著相关(P<0.01) -
从不同种源蒙古栎种子表型性状与生态因子之间的相关性分析(表4)可知:海拔与种宽呈显著负相关(P<0.05),与单粒重、千粒重呈极显著负相关(P<0.01),相关系数达−0.67、−0.75、−0.75,即海拔越高,种宽、单粒重和千粒重越小。东经与千粒重、单粒重呈显著正相关(P<0.05),相关系数均达0.72,即随着经度的增加,单粒重、千粒重增大。无霜期与单粒重呈正相关,即无霜期越长,单粒重越大。种长、种形指数与生态因子之间也存在一定的相关性,但差异不显著,说明蒙古栎种子表型性状的差异与种源生态环境因子有一定的相关性,即存在地理变异特性。
表 4 不同种源蒙古栎种子表型性状与生态因子之间的相关性
Table 4. Correlation between seeds phenotypic characters in Q. mongolica and ecological factors from different provenances
生态因子 种长 种宽 种形指数 单粒重 千粒重 东经 0.56 0.58 0.06 0.72* 0.72* 北纬 0.18 0.44 −0.41 0.13 0.13 海拔 −0.57 −0.67* 0.07 −0.75** −0.75** 年均气温 −0.41 −0.15 −0.50 −0.40 −0.41 年降水量 0.20 0.34 −0.17 0.45 0.45 无霜期 −0.13 0.05 −0.29 0.01 −0.01 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
以11个种源间蒙古栎种子相关特性为样本,将种子5个表型性状和6个生态因子数据进行主成分分析。由表5可知:特征值大于1有3个主成分,贡献率分别为43.78%、22.92%、12.57%,累计贡献率达79.27%。其中,第1主成分特征值为5.37,对应的特征向量千粒重和单粒重最大,均为0.41。可以把第1主成分称为种子质量因子。第2主成分特征值为2.52,对应的特征向量无霜期最大,为0.51,可以把第2主成分称为环境因子;第3主成分特征值为1.38,对应的特征向量北纬最小,可以把第3主成分称为地理因子。综上可知,影响蒙古栎种子特性的因素从大到小为单粒重、千粒重、无霜期、北纬。
表 5 不同种源蒙古栎种子相关特性的主成分分析
Table 5. Principal component analysis of seeds related characteristics in Q. mongolica from different provenances
主成分 特征向量 特征值 贡献率/% 累计贡献率/% 种长 种宽 种形指数 单粒重 千粒重 东经 北纬 海拔 年均气温 年降水量 无霜期 1 0.38 0.37 0.13 0.41 0.41 0.36 0.12 −0.37 −0.21 0.18 −0.04 5.37 43.78 43.78 2 −0.09 0.14 −0.42 0.00 0.00 0.00 0.29 −0.15 0.50 0.42 0.51 2.52 22.92 66.70 3 0.16 −0.04 0.47 0.19 0.18 −0.24 −0.55 0.26 0.26 0.28 0.32 1.38 12.57 79.27 -
由聚类分析结果(图1)可以看出:根据单粒重和千粒重,不同种源蒙古栎共分为4类,分别为Ⅰ、Ⅱ、Ⅲ和Ⅳ类种群,其中黑龙江省黑河市、辽宁省本溪市、辽宁省抚顺市为Ⅰ类,种子的单粒重和千粒重远高于所有种源的均值,为最优种源;北京市门头沟区、吉林省延边市为Ⅱ类,单粒重和千粒重略高于均值;河北省张家口市、河北省承德市兴隆县、山东省泰安市、河北省秦皇岛市为Ⅲ类,单粒重和千粒重略低于均值;河北省承德市围场县、内蒙古赤峰市为Ⅳ类,单粒重和千粒重最小。从聚类分析结果看,蒙古栎种子表型性状表现出了一定的区域性,天然种源基本上分布于东北部地区,Ⅰ类分布于东部偏冷地区,Ⅱ类分布于东部和北部地区,Ⅲ类分布于中部地区(华北和华东),Ⅳ类分布于北部地区,表现出从东北向西南的走向,种源质量逐渐降低,这与相关性分析结果相吻合。
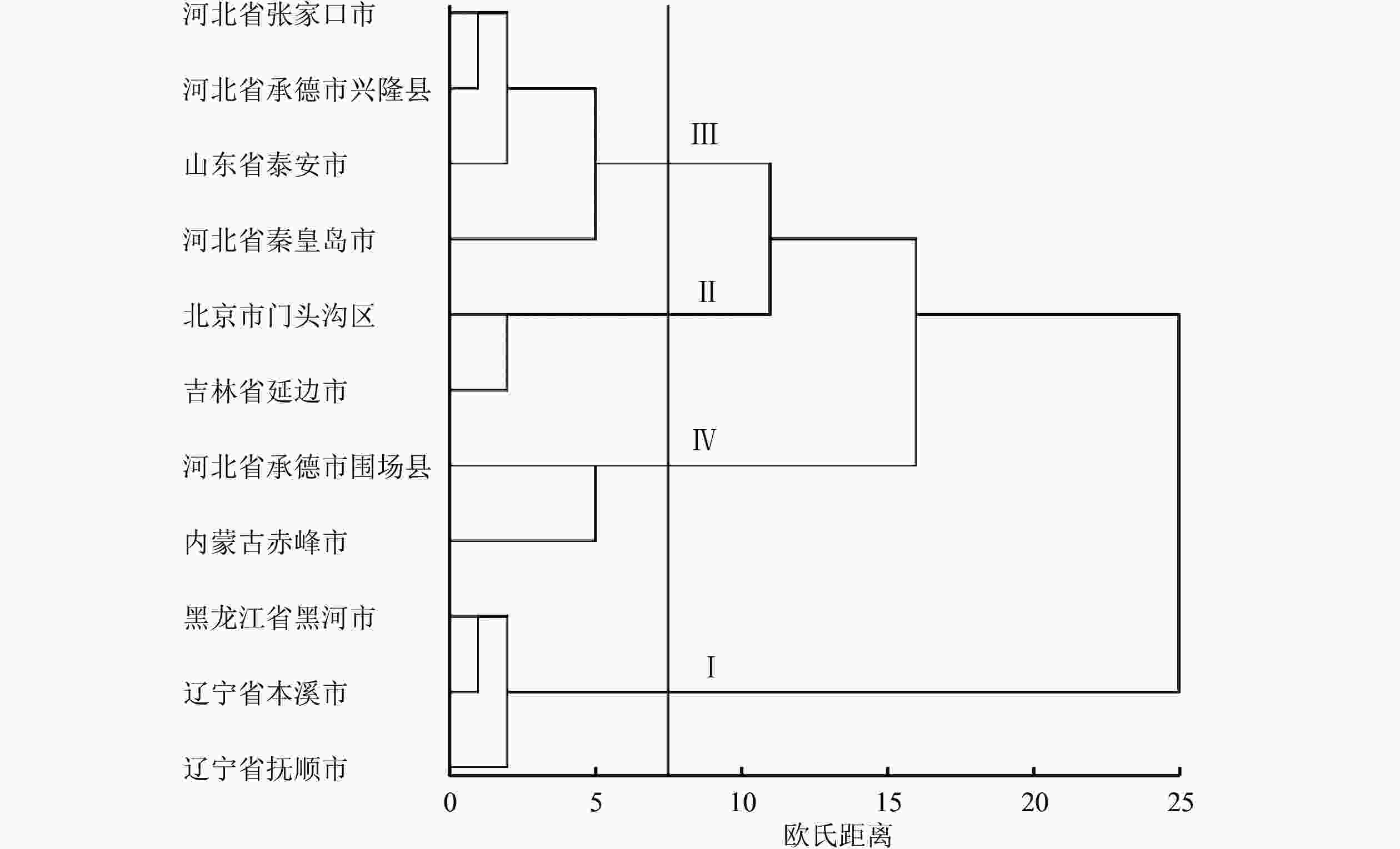
图 1 不同种源蒙古栎种子的聚类分析
Figure 1. Cluster analysis for Q. mongolica seeds from different provenances
-
种质资源的调查和评价是研究品种起源、演化和驯化的基础,对优异资源筛选、品种选育具有重要意义[17]。本研究通过对蒙古栎种子5种表型性状的测定和分析,得出不同种源蒙古栎种实表型性状之间存在差异,其中种长、千粒重均存在显著差异(P<0.05),这与厉月桥等[18]对蒙古栎、李迎超[19]对栓皮栎Q. variabilis、麻栎Q. acutissima的研究结果一致,且种长与种宽、单粒重、千粒重之间均存在极显著正相关(P<0.01),表明蒙古栎种质资源丰富,各生长性状在种源间存在较大的遗传改良潜力。
植物与其生存环境的关系一直是关注的热点[20],处于不同生态环境的同种物种,经过长期对当地生境的适应,发生了地理变异,形成了不同的地理种源[21]。蒙古栎地理分布较为广泛,生存环境存在较大差异。气候条件,如气温、水分等对植物的有性繁殖有着重要的影响[22],海拔等对种子、果实的品质也具有影响[23-27],而气候条件的波动及其他环境条件的改变会影响物种更新和种群动态[28-30]。本研究表明:蒙古栎不同种源种子表型特征与生态因子之间均存在相关性,其中海拔与种宽存在显著负相关,这与厉月桥等[18]对蒙古栎研究结果相同,单粒重和千粒重受分布区海拔的影响存在垂直变异,说明蒙古栎具有广泛的适应性,随着环境条件的变化,出现遗传变异。这与李斌等[31]对白皮松Pinus bungeana及张清等[32]对铁橡栎Quercus cocciferoides的研究结果一致,即变异越大,适应环境的能力就越强。单粒重与东经存在显著正相关,随着经度的降低,单粒重和千粒重逐渐降低,种形指数无显著变化,种源质量相对降低,可能由于种子储藏营养物质的量直接影响幼苗建植,大种子有利于幼苗建植早期根系的发育和生物量的积累[33],从而具有更强的生长势、逆境耐受能力和对资源的竞争能力[34]。种子的形状与种长和年均气温的相关性较大,说明蒙古栎种子形状受自身遗传物质控制,同时也受周边环境因子的影响,存在地理变异,由于长期生长在生态条件明显不同的地区,蒙古栎种源间可能产生遗传分化,这与相关的研究结果[15, 35]一致。
-
本研究表明:影响种子特性的因素按照重要性从大到小依次为单粒重、千粒重、无霜期、北纬。主成分分析表明:贡献率最大的第1主成分表征的特征向量为单粒重和千粒重,因此可将单粒重和千粒重作为快速筛选种源优良与否的优先筛选指标,这与常恩福等[36]对铁橡栎的研究结果一致。第2、3主成分主要为环境因子和地理因子,这与常恩福等[36]、张清等[32]的研究结果存在差异。常恩福等[36]研究表明:第2、3、4主成分是种形指数、纬度和海拔,即形状因子和生态环境因子;张清等[32]研究表明:第2主成分为种形指数,可能与树种特性及分布范围存在一定关系。
根据种子性状的综合聚类分析结果,将11个种源按单粒重和千粒重聚为4类,地理分布相近的种源并没有聚在一起,说明了种源间表型性状变异不具有连续性。Ⅰ类种群中,黑河同辽宁地理位置距离较大,但聚为同一类群,可能由于其经度相似,经度和千粒重、单粒重呈显著正相关有关,这与李文文[37]对蒙古栎种源变异的研究结果一致。Ⅱ类和Ⅲ类种群中,河北和山东相隔距离较大,可能由于同一类群在不同种源之间地形起伏较大,形成了一定程度上的地理隔离,从而造成同一类群的种源,地理距离反而较大,这与李文文[37]的研究结果一致。Ⅲ类种群除山东泰安外,均位于河北地区,地理距离相对集中,且与Ⅳ类种群相隔地理距离较小,单粒重和千粒重均低于均值,种源质量相对较差。
其中Ⅰ类种群在种源质量方面优于Ⅱ、Ⅲ和Ⅳ类,Ⅰ类分布区均为蒙古栎集中分布区[15],同时根据蒙古栎适生分布区的研究可知:不同种源间地理梯度变异明显,呈东北—西南走向,这与李斌等[31]对白皮松的研究结果相同。
只通过5种表型性状特征确定各种源之间的表型差异存在一定的局限性,因此,还需进一步连续多年收集测定或继续开展分子水平方面的研究,深入分析蒙古栎不同种源之间的遗传变异。
Diversity of phenotypic characters of Quercus mongolica seeds from different provenances
-
摘要:
目的 研究不同种源蒙古栎Quercus mongolica种子表型性状多样性,为蒙古栎种质资源收集、评价和利用提供科学依据。 方法 在中国蒙古栎天然分布区选取11个种源采集种子,分别测量种子的种长、种宽、种形指数、单粒重、千粒重5个表型性状。并利用SPSS软件对不同种源的蒙古栎种子进行性状差异性、相关性及主成分分析。 结果 ①不同种源蒙古栎种子的种长、种宽、种形指数、单粒重差异较大,各种源千粒重之间均存在显著差异(P<0.05);②不同种源蒙古栎种子的种长与种宽、单粒重、千粒重均存在极显著正相关(P<0.01);③不同种源海拔与种宽存在显著负相关(P<0.05),与单粒重、千粒重存在极显著负相关(P<0.01);④影响种子性状的因素按照重要性从高到低依次为单粒重、千粒重、无霜期、北纬;根据聚类分析结果,可将11个种源划分为4类,分别为Ⅰ、Ⅱ、Ⅲ和Ⅳ类,其中,黑龙江省黑河市、辽宁省本溪市、辽宁省抚顺市聚为Ⅰ类,单粒重和千粒重显著高于其他类别种源(P<0.05),为最优种源。 结论 蒙古栎种子表型性状随地理梯度变异明显,整体呈从东北—西南种源质量逐渐下降的趋势,单粒重和千粒重可以作为快速筛选种源优良与否的优先筛选指标。图1表5参37 Abstract:Objective The objective is to study the diversity of phenotypic characters of Quercus mongolica seeds from different provenances, so as to provide scientific basis for collection, evaluation and utilization of Q. mongolica germplasm resources. Method Seeds were collected from 11 provenances in natural distribution areas of Q. mongolica in China and 5 phenotypic characters were measured, including seed length, seed width, seed shape index, single grain weight, and thousand grain weight. SPSS software was used to analyze the character difference, correlation and principal component of Q. mongolica seeds from different provenances. Result (1) Seed length, seed width, seed shape index and single seed weight varied greatly among different provenances, and there were significant differences (P< 0.05) in thousand grain weight among different provenances. (2) There was a significant positive correlation (P<0.01) between seed length and seed width, single grain weight and thousand grain weight among different provenances. (3) There was a significant negative correlation (P< 0.05) between altitude and seed width, and a very significant negative correlation (P<0.01) with single grain weight and thousand grain weight. (4) According to the importance, the factors affecting seed characteristics in descending order were single grain weight, thousand grain weight, frost-free period and north latitude. According to the results of cluster analysis, the 11 provenances can be divided into 4 categories: Ⅰ, Ⅱ, Ⅲ and Ⅳ respectively. Among them, the provenances from Heihe City in Heilongjiang Province, Benxi City and Fushun City in Liaoning Province were grouped into Class Ⅰ, whose single grain weight and thousand grain weight were significantly higher (P<0.05) than those of other provenances, and they were the best provenances. Conclusion The phenotypic characters of Q. mongolica seeds vary significantly with geographical gradient, and the quality of provenances decreases gradually from northeast to southwest. Single grain weight and thousand grain weight can be used as the priority screening index for rapid screening of provenances. [Ch, 1 fig. 5 tab. 37 ref.] -
Key words:
- Quercus mongolica /
- provenances /
- phenotypic characters /
- diversity /
- ecological factors
-
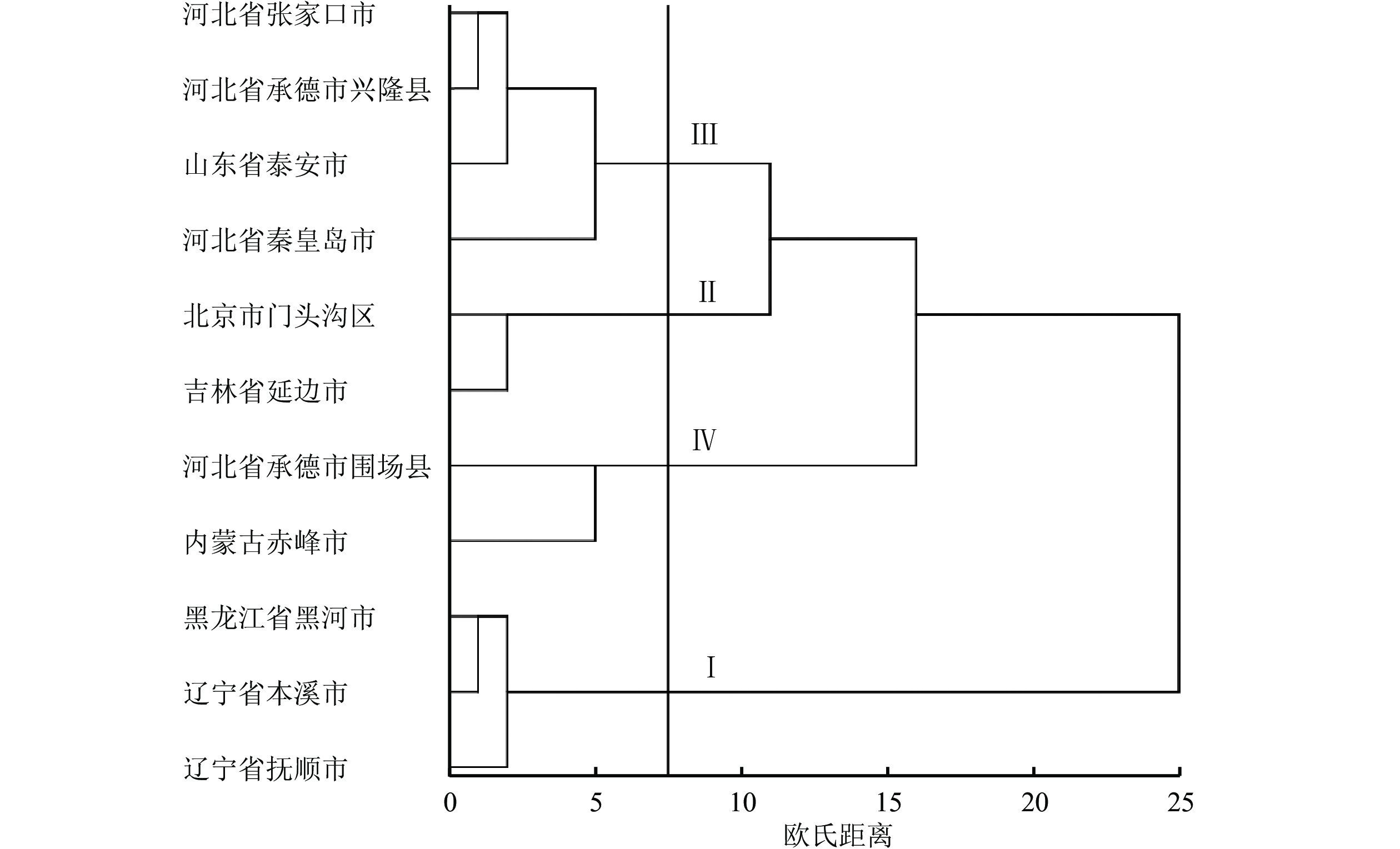
图 1 不同种源蒙古栎种子的聚类分析
Figure 1 Cluster analysis for Q. mongolica seeds from different provenances
表 1 不同种源蒙古栎主要生态因子
Table 1. Main ecological factors of Q. mongolica from different provenances
种源编号 种源地 北纬(N) 东经(E) 海拔/m 年均气温/℃ 年降水量/mm 无霜期/d 1 黑龙江省黑河市 49°49′18″ 127°16′48″ 294 −1.0 525 110 2 辽宁省抚顺市 41°56′33″ 125°14′24″ 673 5.1 863 130 3 辽宁省本溪市 40°40′05″ 124°00′55″ 487 6.5 926 170 4 河北省张家口市 39°53′36″ 114°57′17″ 1 573 6.0 400 152 5 河北省承德市围场县 42°10′21″ 117°15′31″ 1 330 3.3 370 80 6 河北省承德市兴隆县 40°33′34″ 117°30′00″ 1 092 7.6 763 130 7 河北省秦皇岛市 40°08′33″ 119°25′28″ 1 110 8.9 741 180 8 北京市门头沟区 39°59′01″ 115°26′28″ 1 270 6.4 420 150 9 吉林省延边市 43°11′32″ 129°32′20″ 370 3.9 580 146 10 内蒙古赤峰市 42°06′57″ 118°19′36″ 1 175 7.7 559 157 11 山东省泰安市 53°27′05″ 117°16′20″ 868 13.0 750 200  下载: 导出CSV
下载: 导出CSV
表 2 不同种源蒙古栎种子表型性状差异性分析
Table 2. Difference analysis of seeds phenotypic characters in Q. mongolica from different provenances
种源地 种长/mm 种宽/mm 种形指数 单粒重/g 千粒重/g 黑龙江省黑河市 20.94±0.38 a 16.58±0.44 bc 1.26±0.04 ab 2.93±0.07 ab 2 949.41±9.65 b 辽宁省抚顺市 20.96±0.73 a 17.80±0.71 a 1.18±0.01 ab 3.05±0.16 a 3 027.39±7.74 a 辽宁省本溪市 18.37±0.46 bc 15.46±0.64 d 1.20±0.02 ab 2.83±0.02 b 2 878.82±7.03 c 河北省张家口市 16.47±0.51 de 13.11±0.68 f 1.26±0.10 ab 1.82±0.10 e 1 824.11±9.20 h 河北省承德市围场县 16.49±0.50 de 13.94±0.23 ef 1.17±0.12 ab 1.41±0.07 f 1 482.71±1.36 j 河北省承德市兴隆县 16.18±0.22 e 14.01±0.49 ef 1.15±0.01 abc 1.79±0.07 e 1 765.37±4.14 i 河北省秦皇岛市 18.22±0.88 c 14.29±0.23 e 1.28±0.04 a 2.07±0.06 d 2 075.77±2.53 f 北京市门头沟区 19.38±1.19 b 16.02±0.97 cd 1.21±0.10 ab 2.31±0.11 c 2 310.59±7.58 e 吉林省延边市 19.39±0.23 b 17.26±0.19 ab 1.12±0.02 bcd 2.45±0.07 c 2 421.51±2.44 d 内蒙古赤峰市 11.38±0.38 f 11.07±0.81 g 1.03±0.09 cd 1.22±0.09 g 1 244.40±5.51 k 山东省泰安市 17.40±1.04 cd 17.10±0.42 abc 1.02±0.09 d 1.92±0.03 de 1 926.28±1.40 g 说明:同列不同字母表示不同种源地之间差异显著(P<0.05)
下载: 导出CSV
表 3 不同种源蒙古栎种子表型性状的相关性
Table 3. Correlation of seeds phenotypic characters in Q. mongolica from different provenances
性状 种长 种宽 种形指数 单粒重 千粒重 种长 1.00 0.88** 0.52 0.88** 0.88** 种宽 1.00 0.06 0.79** 0.78** 种形指数 1.00 0.40 0.41 单粒重 1.00 1.00** 说明:**表示极显著相关(P<0.01)
下载: 导出CSV
表 4 不同种源蒙古栎种子表型性状与生态因子之间的相关性
Table 4. Correlation between seeds phenotypic characters in Q. mongolica and ecological factors from different provenances
生态因子 种长 种宽 种形指数 单粒重 千粒重 东经 0.56 0.58 0.06 0.72* 0.72* 北纬 0.18 0.44 −0.41 0.13 0.13 海拔 −0.57 −0.67* 0.07 −0.75** −0.75** 年均气温 −0.41 −0.15 −0.50 −0.40 −0.41 年降水量 0.20 0.34 −0.17 0.45 0.45 无霜期 −0.13 0.05 −0.29 0.01 −0.01 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 5 不同种源蒙古栎种子相关特性的主成分分析
Table 5. Principal component analysis of seeds related characteristics in Q. mongolica from different provenances
主成分 特征向量 特征值 贡献率/% 累计贡献率/% 种长 种宽 种形指数 单粒重 千粒重 东经 北纬 海拔 年均气温 年降水量 无霜期 1 0.38 0.37 0.13 0.41 0.41 0.36 0.12 −0.37 −0.21 0.18 −0.04 5.37 43.78 43.78 2 −0.09 0.14 −0.42 0.00 0.00 0.00 0.29 −0.15 0.50 0.42 0.51 2.52 22.92 66.70 3 0.16 −0.04 0.47 0.19 0.18 −0.24 −0.55 0.26 0.26 0.28 0.32 1.38 12.57 79.27
下载: 导出CSV
-
[1] 申艳梅, 郭平平, 刘淑玲, 等. 蒙古栎的加工利用研究进展[J]. 森林工程, 2014, 30(5): 58 − 60. SHEN Yanmei, GUO Pingping, LIU Shuling, et al. Review of advances in progress and utilization of Mongolian oak [J]. For Eng, 2014, 30(5): 58 − 60. [2] 河北植被编辑委员会, 河北省农业区划委员会办公室. 河北植被[M]. 北京: 科学出版社, 1996: 1 − 15. Hebei Vegetation Editorial Committee, Office of Hebei Agricultural Zoning Commission. Hebei Vegetation[M]. Beijing: Science Press, 1996: 1 − 15. [3] 孙佳庆. 珍贵树种大径级用材林培育技术研究[J]. 吉林林业科技, 2012, 41(6): 14 − 18. SUN Jiaqing. Study on the cultivation technique for arge sized timber of rare tree species [J]. J Jilin For Sci Technol, 2012, 41(6): 14 − 18. [4] 王荣芳, 张子言, 李德海. 酶解法对蒙古栎实壳提取物活性成分及抗氧化活性的影响[J/OL]. 北京林业大学学报, 2022[2022-05-09]. doi:10.12171/j.1000-1522.20210352. WANG Rongfang, ZHANG Ziyan, LI Dehai. Effects of extraction methods on extraction components and antioxidant activity of Quercus mongolica shell[J/OL]. J Beijing For Univ, 2022[2022-05-09]. doi: 10.12171/j.1000-1522.20210352. [5] 姚大地, 于海洪. 蒙古栎叶、果实成分分析[J]. 吉林林学院学报, 1998, 14(4): 205 − 207. YAO Dadi, YU Haihong. Analysis of composition of leave and fruits on Quercus mongolica [J]. J Jilin For Univ, 1998, 14(4): 205 − 207. [6] 李佳宁. 蒙古栎生殖生物学特性的初步研究[D]. 沈阳: 沈阳农业大学, 2020. LI Jianing. A Preliminary Study on the Characteristics of Reproductive Biology of Quercus mongolica[D]. Shenyang: Shenyang Agricultural University, 2020. [7] GREIPSSON S, DAVY A J. Seed mass and germination behaviour in populations of the dune-building grass Leymus arenarius [J]. Ann Bot, 1995, 76(5): 493 − 501. [8] 火艳, 招雪晴, 黄厚毅, 等. 观赏石榴表型遗传多样性分析[J]. 浙江农林大学学报, 2020, 37(5): 939 − 949. HUO Yan, ZHAO Xueqing, HUANG Houyi, et al. Phenotypic genetic diversity of ornamental pomegranate cultivars [J]. J Zhejiang A&F Univ, 2020, 37(5): 939 − 949. [9] 张振, 张含国, 周宇, 等. 红松多无性系群体的种实性状变异研究[J]. 北京林业大学学报, 2015, 37(2): 67 − 78. ZHANG Zhen, ZHANG Hanguo, ZHOU Yu, et al. Variation of seed characters in Korean pine (Pinus koraiensis) multiclonal populations [J]. J Beijing For Univ, 2015, 37(2): 67 − 78. [10] 颉刚刚, 欧阳丽婷, 谢军, 等. 新疆地区欧洲李叶片表型性状多样性及亲缘关系分析[J]. 植物资源与环境学报, 2018, 27(3): 72 − 78. XIE Ganggang, OUYANG Liting, XIE Jun, et al. Analyses on diversity of leaf phenotypic traits and genetic relationships of Prunus domestica in Xinjiang region [J]. J Plant Resour Environ, 2018, 27(3): 72 − 78. [11] 颜冰, 刘刚, 陈爱华, 等. 东北三省不同种源蒙古栎种子表型性状和淀粉含量对比分析[J]. 安徽农业科学, 2015, 43(30): 121 − 123. YAN Bing, LIU Gang, CHEN Aihua, et al. Quercus mongolica seeds’ character and starch content comparisons of different provenance in the three northeastern Provinces [J]. J Anhui Agric Sci, 2015, 43(30): 121 − 123. [12] 梁德洋, 蒋路平, 张秦徽, 等. 辽宁省11个蒙古栎种源及家系种子性状变异[J]. 东北林业大学学报, 2019, 47(11): 1 − 5. LIANG Deyang, JIANG Luping, ZHANG Qinhui, et al. Variance analysis of seed traits in provenances and families on Quercus mongolica [J]. J Northeast For Univ, 2019, 47(11): 1 − 5. [13] 李亚男, 李东胜, 许中旗, 等. 栎属不同种源树种的出苗及生长过程[J]. 西北林学院学报, 2014, 29(4): 139 − 144. LI Ya’nan, LI Dongsheng, XU Zhongqi, et al. The process of emergence and height growth of Quercus species in different provenances [J]. J Northwest For Univ, 2014, 29(4): 139 − 144. [14] 胡玉珠. 4个不同种源蒙古栎种实形态分析[J]. 吉林林业科技, 2020, 49(1): 1 − 3. HU Yuzhu. Morphological analysis of 4 different provenances of Quercus mongolica seeds [J]. J Jilin For Sci Technol, 2020, 49(1): 1 − 3. [15] 厉月桥. 木本能源植物蒙古栎与辽东栎资源调查与优良种质资源筛选[D]. 北京: 中国林业科学研究院, 2011: 17 − 20. LI Yueqiao. Resource Investigation and Superior Germplasm Resources Selection of Woody Energy Plants Quercus mongolica Fisch. and Quercus liaotungensis Koidz[D]. Beijing: Chinese Academy of Forestry, 2011: 17 − 20. [16] 包松莲, 李志国, 张建云, 等. 云南塔拉种子形态变异的初步研究[J]. 氨基酸和生物资源, 2011, 33(2): 17 − 19, 38. BAO Songlian, LI Zhiguo, ZHANG Jianyun, et al. A preliminary study on morphological variation of seeds of Tara in Yunnan [J]. Amino Acids Biotic Resour, 2011, 33(2): 17 − 19, 38. [17] DENWAR N N, AWUKU F J, DIERS B, et al. Genetic diversity, population structure and key phenotypic traits driving variation within soyabean (Glycine max) collection in Ghana [J]. Plant Breed, 2019, 138(5): 577 − 587. [18] 厉月桥, 李迎超, 吴志庄. 不同种源蒙古栎种子表型性状与淀粉含量的变异分析[J]. 林业科学研究, 2013, 26(4): 528 − 532. LI Yueqiao, LI Yingchao, WU Zhizhuang. Variation in phenotype characters and starch content of Quercus mongolica Fisch. seed from different provenances [J]. For Res, 2013, 26(4): 528 − 532. [19] 李迎超. 木本淀粉能源植物栓皮栎与麻栎资源调查及地理种源变异分析[D]. 北京: 中国林业科学研究院, 2013: 51 − 71. LI Yingchao. Resource Investigation and Provenance Analysis of Woody Starch Engergy Plant Quercus variabilis BL. and Quercus acutissima Carr.[D]. Beijing: Chinese Academy of Forestry, 2013: 51 − 71. [20] SIEFERT A, FRIDLEY J D, RITCHIE M E. Community functional responses to soil and climate at multiple spatial scales: when does intraspecific variation matter? [J/OL]. PLoS One, 2014, 9(10): e111189[2021-01-11]. doi: 10.1371/journal.pone.0111189. [21] 安海龙, 谢乾瑾, 刘超, 等. 水分胁迫和种源对黄柳叶功能性状的影响[J]. 林业科学, 2015, 51(10): 75 − 84. AN Hailong, XIE Qianjin, LIU Chao, et al. Effects of water stress and provenance on leaf functional traits of Salix gordejevii [J]. Sci Silv Sin, 2015, 51(10): 75 − 84. [22] ADLER P B, LAMBERS J H R. The influence of climate and species composition on the population dynamics of ten prairie forbs [J]. Ecology, 2008, 89(11): 3049 − 3060. [23] 梁小玉, 张新全. 不同海拔对宝兴鸭茅种子生产的影响初探[J]. 草业科学, 2007, 24(12): 64 − 66. LIANG Xiaoyu, ZHANG Xinquan. Effect of elevation on seed production of Dactlis glomerata cv. Baoxing [J]. Pratacult Sci, 2007, 24(12): 64 − 66. [24] 王志恒, 陈安平, 朴世龙, 等. 高黎贡山种子植物物种丰富度沿海拔梯度的变化[J]. 生物多样性, 2004, 12(1): 82 − 88. WANG Zhiheng, CHEN Anping, PIAO Shilong, et al. Pattern of species richness a long an altitudinal gradient on Gaoligong Mountains, Southwest China [J]. Biodiversity Sci, 2004, 12(1): 82 − 88. [25] 郭淑青, 齐威, 王玉林, 等. 青藏高原东缘海拔对植物种子大小的影响[J]. 草业学报, 2010, 19(1): 50 − 58. GUO Shuqing, QI Wei, WANG Yulin, et al. Effects of altitude on seed size on the eastern Qinghai-Tibetan Plateau [J]. Acta Pratacult Sin, 2010, 19(1): 50 − 58. [26] 李丹, 彭少麟. 3个不同海拔梯度马尾松种群的遗传多样性及其与生态因子的相关性[J]. 生态学报, 2001, 21(3): 415 − 440. LI Dan, PENG Shaolin. Genetic diversity in three Pinnus massoniana populations in different elevations and its relation with ecological factors [J]. Acta Ecol Sin, 2001, 21(3): 415 − 440. [27] PLUESS A R, SCHÜTZ W, STCKLIN J. Seed weight increases with altitude in the Swiss Alps between related species but not among populations of individual species [J]. Oecologia, 2005, 144(1): 55 − 61. [28] ARFT A M, WALKER M D, GUREVITCH J, et al. Responses of tundra plants to experimental warming: metal analysis of the international tundra experiment [J]. Ecol Monographs, 1999, 69(4): 491 − 511. [29] DIEMER M. Population stasis in a high-elevation herbaceous plant under moderate climate warming [J]. Basic Appl Ecol, 2002, 3(1): 77 − 83. [30] FITCH E A, WALCK J L, HIDAYATI S N. Agroecosystem management for rare species of Paysonia (Brassicaceae): integrating their seed ecology and life cycle with cropping regimens in a changing climate [J]. Am J Bot, 2007, 94(1): 101 − 110. [31] 李斌, 顾万春, 卢宝明. 白皮松天然群体种实性状表型多样性研究[J]. 生物多样性, 2002, 10(2): 181 − 188. LI Bin, GU Wanchun, LU Baoming. A study on phenotypic of seeds and cones characteristics in Pinus bungeana [J]. Biodiversity Sci, 2002, 10(2): 181 − 188. [32] 张清, 肖桂英, 李品荣, 等. 同一种源铁橡栎种子的形态特征及变异[J]. 黑龙江农业科学, 2019(8): 26 − 31. ZHANG Qing, XIAO Guiying, LI Pinrong, et al. Seed morphological characteristics and variation of Quercus cocciferoides from the same provence [J]. Heilongjiang Agric Sci, 2019(8): 26 − 31. [33] KENNEDY P G, HAUSMANN N J, WENK E H, et al. The importance of seed reserves for seedling performance: an integrated approach using morphological, physiological, and stable isotope techniques [J]. Oecologia, 2004, 141: 547 − 554. [34] LEBRIJA-TREJOS E, REICH P B, HERNNDEZ A, et al. Species with greater seed mass are more tolerant of conspecific neighbours: a key driver of early survival and future abundances in a tropical forest [J]. Ecol Lett, 2016, 19: 1071 − 1080. [35] 张俊, 尚家辉, 程广有, 等. 蒙古栎生长性状种内变异规律初步研究[J]. 北华大学学报(自然科学版), 2020, 21(4): 447 − 451. ZHANG Jun, SHANG Jiahui, CHENG Guangyou, et al. Preliminary study on intraspecific variation of growth traits of Quercus mongolica Fisch. [J]. J Beihua Univ Nat Sci, 2020, 21(4): 447 − 451. [36] 常恩福, 张清, 肖桂英, 等. 铁橡栎不同种源及家系种子的形态特征及变异[J]. 种子, 2020, 39(6): 53 − 58. CHANG Enfu, ZHANG Qing, XIAO Guiying, et al. Morphological characteristics and variation analysis of seeds from different provenances and families of Quercus cocciferoides [J]. Seed, 2020, 39(6): 53 − 58. [37] 李文文. 蒙古栎(Quercus mongolica Fisch. )种源变异及无性繁殖研究[D]. 北京: 中国林业科学研究院, 2010: 13 − 36. LI Wenwen. Study on Variation of Provence and Vegetative Propagation in Quercus mongolica Fisch.[D]. Beijing: Chinese Academy of Forestry, 2010: 13 − 36. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220133
 点击查看大图
点击查看大图
计量
- 文章访问数: 1800
- HTML全文浏览量: 277
- PDF下载量: 67
- 被引次数: 0
