





-
黄土高原是干旱半干旱地区,降水量少,降水分布不均,生态环境脆弱。自20世纪50年代以来,该地区采取了一系列的植被恢复措施,并取得了显著成效,但由于不合理的植被建设,导致林水矛盾激化,部分林地出现“小老头树”和土壤水分亏缺等现象[1-3]。基于此,研究黄土高原地区林地树种耗水规律尤为重要。99.8%以上的植物蒸腾耗水来自树干液流,液流速率可以较为准确地描述单木蒸腾耗水过程[4-5]。早期对植物蒸腾研究多集中于昼间,忽略夜间液流的存在。随着观测技术的发展与研究时间尺度的精细化,越来越多的研究发现整日树干液流可划分为昼间与夜间,昼夜液流变化特征具有差异性[6]。昼间液流量占整日液流量的80%~96%,是植物生长的主要环节,研究昼间液流的变化特征有助于理解植物的生长策略[7];夜间液流量占比为4%~20%[8],占比较少,但目前研究普遍认为[9]:夜间液流有助于植物适应干旱环境,在补充水分、传输营养、供应氧气等方面发挥着重要的作用,因此在研究植物蒸腾耗水时,夜间液流不可被忽视。
已经有大量的研究表明:植物液流速率受自身生理结构和气象因子的影响。任启文等[10]研究发现:冀北山地油松Pinus tabulaeformis液流速率与气温、太阳辐射、水汽压亏缺、风速呈正相关,与空气湿度呈负相关;贾国栋等[11]通过研究北方土石山区的油松、刺槐Robinia pseudoacacia发现:影响植物蒸腾速率的主要气象因子为太阳辐射、水汽压亏缺和风速;也有研究认为[12]:不同时间尺度下树干液流与气象因子的响应不同。上述研究关注于整日树干液流与气象因子关系研究,然而昼夜树干液流对环境因子响应机制不同,若不考虑昼夜液流变化特征及其对环境因子响应差异性,可能会降低植物耗水估算精度,影响林地水资源管理。目前,关于晋西黄土区生长季刺槐昼夜树干液流速率的研究较少,不同月份气象因子对昼夜树干液流速率的影响有待进一步研究。基于此,本研究以山西省吉县蔡家川流域的主要造林树种刺槐作为研究对象,定位观测其树干液流和气象因子。主要研究目的是①分析刺槐昼夜树干液流变化特征;②分析各月夜间树干液流对总液流的贡献率;③探究各月昼夜树干液流变化对气象因子的响应;④建立各月昼夜树干液流与气象因子关系模型,估算样地林分耗水量。
-
研究区位于晋西黄土残塬沟壑区山西省吉县蔡家川流域(36°14′27″~36°18′23″N,110°39′45″~110°47′45″E),属于黄河支流,流域面积为40.10 km2。该地区海拔为900~1500 m,属暖温带大陆性季风气候,年平均气温为10.0 ℃,年平均降水量为575.9 mm,降水分布不均,4—10月降水量约占全年的85% (489.5 mm),年平均蒸发量为1 723.9 mm。研究区土壤为褐土,黄土母质。
研究样地为人工刺槐纯林,林分密度为1625株·hm−2,平均树高为9.4 m,平均胸径为18.4 cm。林地灌木稀少,草本植物以萝藦Metaplexis japonica、鼠尾粟Sporobolus fertilis、艾蒿Artemisia lavandulaefolia、茜草Rubia cordifolia、短尾铁线莲Clematis brevicaudata、龙芽草Agrimonia pilosa等为主。
-
选取生长良好,树干通直,无病虫害的刺槐标准木8株。样木基本情况如表1。各样树树干液流采用Granier热扩散探针(TDP)观测,观测时间为2021年5月1日至9月30日。在大约1.3 m胸高处选择观测点,并刮去树干死皮,用配套工具沿垂直方向钻2个小孔,将热源探针插入上方孔中,感应探针插在下方孔中,使用固体胶将树体与探针的空隙涂抹密封。将电缆线固定在树干上并与电源连接,用铝箔纸将安装部位整个树干包裹起来,防止阳光直射和雨水造成数据误差。将数据采集器设置为30 s采集1次数据,隔15 min计算平均值并记录。
表 1 测定样树主要参数
Table 1. Main parameters of sample trees
样树编号 胸径/cm 树高/m 边材面积/cm2 样树编号 胸径/cm 树高/m 边材面积/cm2 1 22.5 10.2 142.2 5 16.8 9.5 85.2 2 21.9 11.2 141.5 6 18.0 11.0 93.4 3 20.6 9.8 138.7 7 17.4 10.3 90.2 4 21.3 11.2 140.2 8 18.2 10.8 92.6 树干液流速率采用Granier经验公式获得。计算公式为:
$$ {F}_{\mathrm{d}}=0.011\;9\times {\left(\frac{{\Delta T}_{{\rm{m}}}-\Delta T}{\Delta T}\right)}^{1.231}\times 3\;600 。 $$ (1) 式(1)中:ΔTm为无液流时加热探针与参考探针的最大温差(℃);ΔT为瞬时温差值(℃);Fd为树干液流速率(cm3·h−1·cm−2)
-
研究区刺槐样地里安装有全自动气象站,可全天观测林冠上方(距地面20 m)气象因子。气温(T,℃)和空气相对湿度(HR,%)采用HMP155A空气温湿度传感器观测,太阳辐射(Rs,W·m−2)采用CMP-3总辐射传感器观测,风速(SW,m·s−1)采用010C-1风速传感器观测,林内20 cm土壤温度(Ts,℃) 采用109土壤温度传感器观测。设置数据采集时间间隔为30 min,并采用CR1000数据采集与记录器收集数据。选取水汽压亏缺(DVP)指标综合反映空气相对湿度与空气温度的协同效应。
$$ {D}_{\mathrm{V}\mathrm{P}}\text=\text{0.611}{\mathrm{e}}^{\frac{17.502T}{T+240.97}}{(}{\text{1}}-{H}_{{\rm{R}}}{)} 。 $$ (2) -
采用baseliner软件将温差数据转化为液流数据,使用Excel整理和计算树干液流、气象因子数据,使用SPSS软件与R语言进行相关分析,使用Origin绘图。图表中所用数据均为平均值±标准差。采用配对样本t检验分析各月整日树干液流量与昼间树干液流量的差异性;采用Pearson相关分析法分析气象因子与树干液流的相关关系;采用随机森林回归模型中相对重要性得分指标[IncMSE,IncMSE指去除自变量后随机森林回归模型预测误差的百分数,自变量关于IncMSE得分越高,说明自变量越重要]与逐步回归方程中因子进入方程时拟合度(R2)的变化值分析气象因子对树干液流的影响程度(随机森林是基于分类树的算法[13],该算法预测精度较高,被多个领域用于评估指标重要性);采用随机森林回归法与逐步回归法构建气象因子与树干液流速率相关模型。
-
根据观测数据可知:研究期间太阳辐射、气温、土壤温度、空气相对湿度整体呈下降趋势,日太阳辐射为14.36~451.83 W·m−2,平均辐射为218.42 W·m−2;日气温为7.5~28.6 ℃,平均气温为20.0 ℃;日地温为12.0~28.6 ℃,平均地温为22.0 ℃;日相对湿度为23.22%~98.83%,平均相对湿度为66.58%。降水主要在8—10月,最大降水发生在2021年9月19日,降水量为61.7 mm。
-
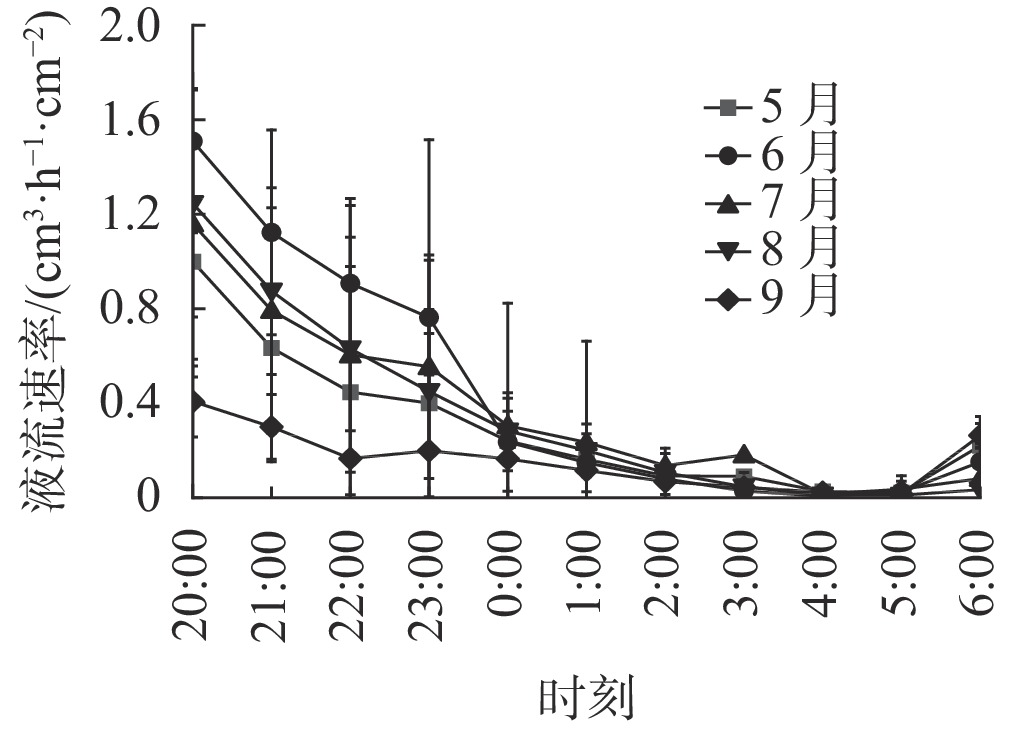
由图1可知:各月刺槐树干液流日变化规律相对一致,呈先增后减变化趋势。晴天树干液流启动时间在6:00左右,启动后树干液流速率迅速上升,到12:00左右达到峰值,随后液流开始下降;夜间(20:00—6:00)有明显的树干液流,变化不稳定,但是呈现一定规律性,因此有必要进一步研究夜间液流的变化特征。
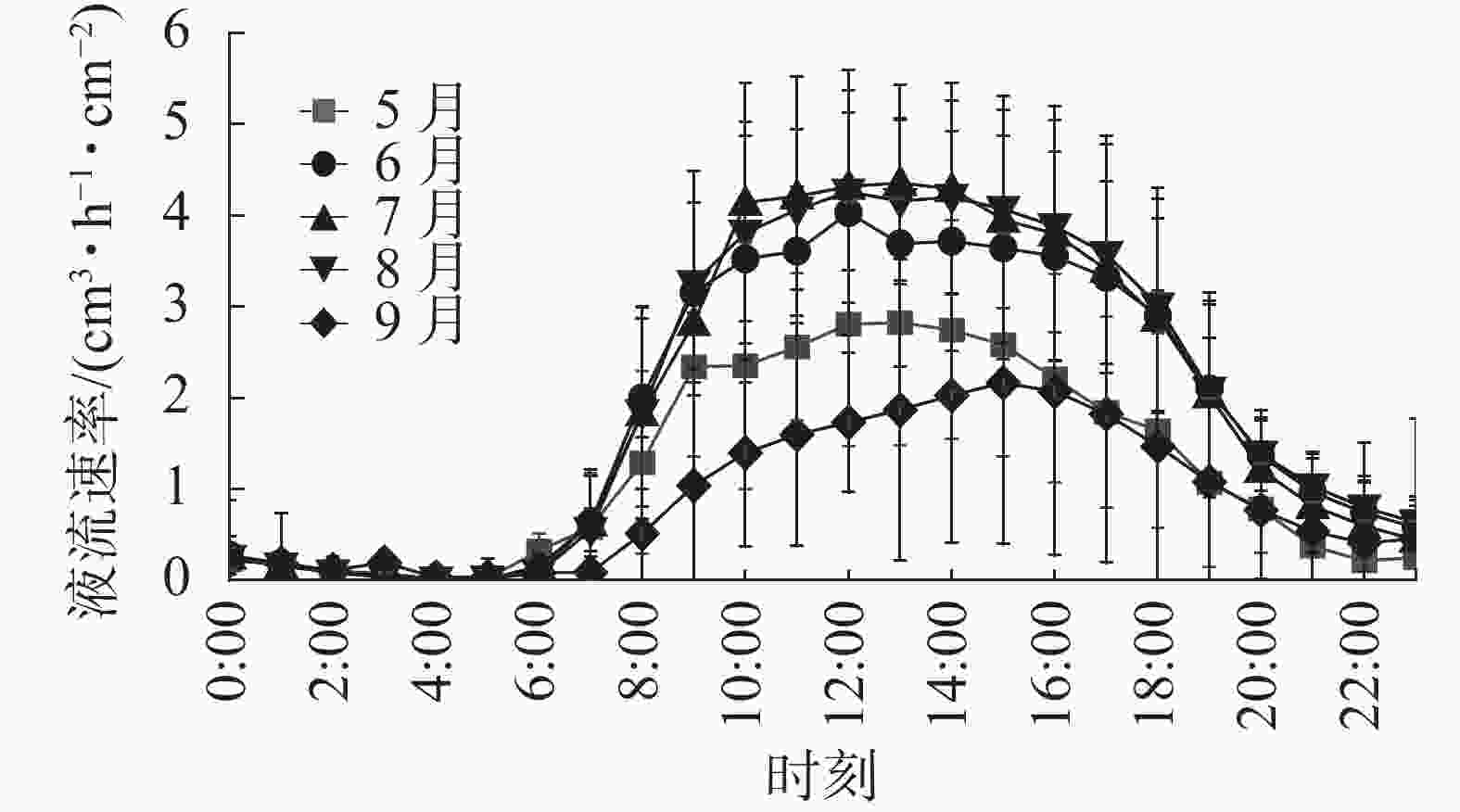
图 1 刺槐树干液流速率日变化
Figure 1. Daily variation of sap flow rate in R. pseudoacacia
-
由图2可知:各月夜间刺槐树干液流速率均表现出递减趋势,前半夜(20:00—24:00)树干液流速率大于后半夜(24:00—6:00)树干液流速率,整个夜晚树干液流速率处于持续下降状态,凌晨5:00左右液流速率降到最低。
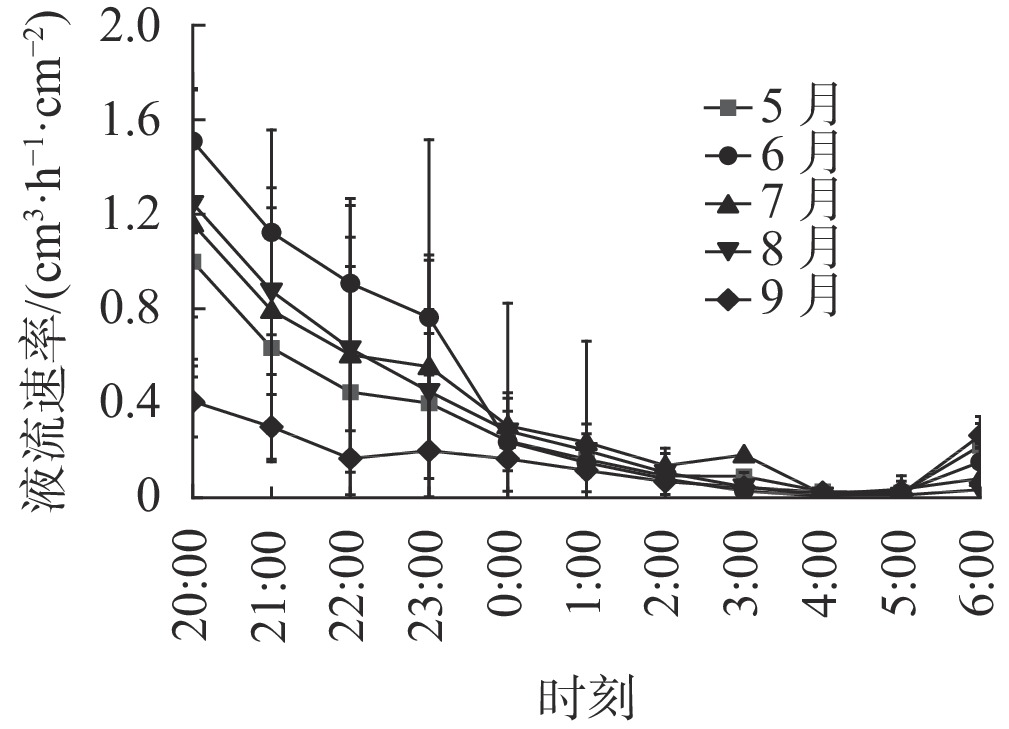
图 2 刺槐树干液流速率夜间变化
Figure 2. Nocturnal variation of sap flow rate in R. pseudoacacia
-
计算刺槐生长季各月昼间、夜间树干液流平均速率与它们对整日液流的贡献率(表2)。结果表明:各月昼夜刺槐树干液流速率及其贡献率差异明显。在5—9月,昼夜平均液流速率呈先增后减变化,昼间平均树干液流速率在7月最大,9月最小;夜间平均液流速率在6月最大,9月最小;各月昼间液流速率对整日液流速率贡献率为88%~93%,贡献率在9月最小,8月最大;各月夜间树干液流速率对整日液流速率贡献率为7%~12%,9月贡献率最大。
表 2 生长季树干液流的昼夜特征
Table 2. Diurnal and nocturnal characteristics of sap flow rate in growing season
月份 昼间平均液流速率/
(cm3·h−1·cm−2)贡献
率/%夜间平均液流速率/
(cm3·h−1·cm−2)贡献
率/%5 2.71±1.07 91.26 0.29±0.23 8.74 6 2.94±1.05 91.16 0.43±0.21 8.84 7 3.24±0.85 92.25 0.35±0.17 7.75 8 3.18±1.61 92.52 0.33±0.18 7.48 9 1.74±1.26 88.39 0.27±0.12 11.61 对整日刺槐树干液流量与昼间树干液流量进行相关性分析和配对样本T检验发现(表3):各月整日刺槐树干液流量与昼间树干液流量呈极显著相关(P<0.01), 整日液流量与昼间液流量呈极显著差异(P<0.01),说明各月夜间树干液流对整日树干液流的影响不可忽视。
表 3 整日与昼间刺槐树干液流量t检验
Table 3. Paired samples t test between daily and diurnal sap flow of R. pseudoacacia
月份 相关分析 配对样本t检验 R2 P t P 5 0.991 0.000 12.582 0.000 6 0.993 0.000 9.261 0.000 7 0.998 0.000 9.054 0.000 8 0.976 0.000 8.234 0.000 9 0.993 0.000 7.982 0.000 -
Pearson相关分析(表4)结果表明:各月昼间刺槐树干液流速率与太阳辐射、土壤温度、气温、水汽压亏缺呈极显著正相关(P<0.01),与空气相对湿度呈极显著负相关(P<0.01),5、6月风速与树干液流速率不存在相关关系(P>0.05)。各月夜间刺槐树干液流速率与土壤温度、气温、水汽压亏缺度呈极显著正相关(P<0.01),与空气相对湿度呈极显著负相关(P<0.01)。
表 4 昼夜树干液流速率与气象因子Pearson相关系数
Table 4. Correlation analysis between diurnal and nocturnal sap flow rate of R. pseudoacacia and meteorological factors
昼夜 月份 太阳辐射 气温 土壤温度 水汽压亏缺 相对湿度 风速 昼间 5 0.575** 0.483** 0.429** 0.290** −0.278** 0.067 6 0.670** 0.539** 0.331** 0.389** −0.305** −0.031 7 0.762** 0.767** 0.497** 0.691** −0.722** 0.192** 8 0.689** 0.747** 0.546** 0.641** −0.638** 0.295** 9 0.674** 0.737** 0.608** 0.696** −0.692** 0.331* 夜间 5 0.582** 0.585** 0.556** −0.266** 0.351** 6 0.213** 0.224** 0.043** −0.049** 0.123** 7 0.512** 0.475** 0.483** −0.394** 0.338** 8 0.747** 0.546** 0.641** −0.638** 0.295** 9 0.141** 0.160** 0.107** −0.058** 0.215** 说明:**表示极显著相关(P<0.01); * 表示显著相关(P<0.05) 基于随机森林模型中IncMSE指标(IncMSE越大自变量越重要)与逐步线性回归R2变化值来判断各月气象因子对刺槐树干液流速率的影响程度。昼间,刺槐在不同月份产生树干液流速率的驱动因子差异不大,随机森林IncMSE指标与逐步线性回归均显示各月对树干液流速率影响较大的气象因子为太阳辐射和气温。夜间,刺槐在不同月份产生树干液流速率的驱动因子存在较大差异,随机森林IncMSE(图3)显示:5月气象因子对树干液流影响程度从大到小排序为水汽压亏缺、土壤温度、风速、相对湿度、气温;6月为土壤温度、水汽压亏缺、相对湿度、气温、风速;7月为气温、相对湿度、风速、土壤温度、水汽压亏缺;8月为水汽压亏缺、相对湿度、气温、土壤温度、风速;9月为风速、水汽压亏缺、土壤温度、相对湿度、气温。夜间,逐步线性回归表明(表5):5、6月土壤温度对刺槐树干液流速率影响最大;8月水汽压亏缺对树干液流速率影响最大;7月气温对树干液流速率影响最大;9月风速对树干液流速率影响最大。气象因子对昼夜树干液流速率的影响程度在2种分析方法上存在不同,但2种方法都表明昼间气象因子对刺槐树干液流速率的影响在各月差异较小,太阳辐射与气温是影响各月昼间树干液流速率的主导因子;夜间气象因子对树干液流速率的影响在不同月份差异较大。除7月外,各月均表现出水汽压亏缺对夜间树干液流速率影响的重要性。

图 3 气象因子对刺槐树干液流速率重要性排序
Figure 3. Meteorological factors of importance on the rate of flow
表 5 刺槐昼夜液流速率与气象因子逐步回归R2变化量
Table 5. Changes of stepwise regression R2 of R. pseudoacacia sap flow rate and meteorological factors
因子 逐步回归R2变化量 昼间 夜间 5月 6月 7月 8月 9月 5月 6月 7月 8月 9月 太阳辐射 − 0.449 0.095 0.015 0.040 − − − − − 气温 0.334 0.026 0.589 0.559 0.543 0.127 − 0.362 − 0.008 水汽压亏缺 0.152 0.150 0.016 0.029 − 0.057 0.148 0.005 0.360 0.232 相对湿度 0.087 − 0.032 0.053 − 0.014 0.084 0.050 0.117 0.008 土壤温度 − 0.007 0.007 0.074 0.014 0.342 0.350 − − − 风速 − − 0.002 0.029 0.018 0.042 − 0.158 0.030 0.246 说明:−表示气象因子未进入到逐步回归方程,以上数据均具有显著性 -
采用随机森林回归法与逐步回归法构建各月昼夜树干液流与气象因子的相关模型。随机森林回归模型无解析表达式,逐步回归模型表达式如表6。根据拟合度来评估2个模型准确度,在6、7、8月2个模型拟合较好,5、9月2个模型拟合较差。总体而言,基于随机森林回归构建的模型拟合度优于逐步回归模型。
表 6 基于逐步回归法和随机森林回归法的刺槐昼夜树干液流与气象因子模型
Table 6. Models of day and night sap flow and meteorological factors based on stepwise regression method and random forest algorithm
昼夜 月份 逐步回归模型 随机森林回归法拟合度/% 模型 拟合度/% 昼间 5 y=−1.29+0.579x2−3.427x4−0.063x5 57.3 67.3 6 y=−0.767+0.003x1+0.301x2−0.149x3−0.869x4 63.2 73.2 7 y=1.509+0.002x1+0.444x2−0.194x3−1.862x4−0.053x5+0.071x6 74.1 86.3 8 y=1.864+0.001x1+0.506x2−0.208x3−2.468x4−0.065x5+0.144x6 75.9 90.6 9 y=1.104+0.001x1+0.300x2−0.204x3+0.297x6 61.5 81.1 夜间 5 y=2.852+0.002x1+0.182x2−0.279x3+0.154x6 48.2 59.4 6 y=4.943+0.003x1+0.346x2−0.363x3−1.252x4−0.025x5−0.110x6 61.6 67.7 7 y=3.966+0.003x1+0.415x2−0.209x3−2.094x4−0.069x5 57.5 61.6 8 y=4.869+0.002x1+0.486x2−0.311x3−2.296x4−0.074x5+0.091x6 50.7 63.9 9 y=−5.886+0.502x2−3.033x4 49.4 54.6 说明:y表示液流速率,x1表示太阳辐射,x2表示气温,x3表示土壤温度,x4表示水汽压亏缺,x5表示相对湿度,x6表示风速 -
根据随机森林回归模型推算刺槐林分各月蒸腾量可知:7月林分蒸腾量最大,为37.5 mm,其他月份依次为8月33.5 mm,6月30.8 mm,5月30.6 mm,9月林分蒸腾量最小,为24.3 mm。推算结果符合刺槐生长规律。生长旺期的蒸腾最大,生长末期的蒸腾最小。
-
刺槐在不同月份的单日树干液流速率均呈先增后减的变化趋势,在日出前后液流启动,峰值时间在12:00左右,并且在10:00—16:00保持较高水平。这与赵楠[14]、凡超等[15]、吴旭等[16]的研究结果一致。各月昼间树干液流速率从小到大依次为9月、5月、6月、8月、7月,说明刺槐昼间蒸腾速率满足生长盛期(6—8月)最大,生长初期(5月)次之,生长末期(9月)最小的规律,且生长季各月昼间树干液流速率对整日液流速率贡献率均在88%以上,说明生长季刺槐白天的蒸腾活动较为强烈。
刺槐在各月夜间树干液流速率从小到依次为9月、5月、8月、7月、6月,生长盛期的夜间树干液流速率显著大于生长初期与生长末期夜间树干液流速率。目前,有关夜间树干液流贡献率的研究较多,如王志超等[17]、HAYAT等[18]研究表明:尾叶桉Larix kaempferi与沙柳Salix psammophila夜间树干液流分别占整日树干液流的6.62%和11.00%。本研究发现:各月夜间液流对整日液流的贡献率范围为7.00%~12.00%,夜间树干液流对整日树干液流的贡献率从大到小依次为生长末期、生长初期、生长盛期,而徐志彬等[19]、张婕等[8]研究表明:生长初期夜间树干液流速率与贡献率较大,可能是研究树种与研究区气象要素不同导致研究结果出现差异。此外,研究不同树种树干液流在不同时间尺度上动态变化的目的是为了能够精准估算各树种耗水变化。本研究发现:生长季各月昼夜树干液流差异较大,夜间树干液流的占比不可忽视,因此今后在月尺度上估算刺槐蒸腾耗水量时需要考虑到月尺度的差异性以及夜间树干液流差异性。
-
植物树干液流速率的变化受到周围环境的影响。本研究发现:刺槐各月昼间树干液流速率与太阳辐射、气温、水汽压亏缺、土壤温度呈正相关,与相对湿度呈负相关。这与刘崴等[20]、YIN等[21]研究结果基本一致。主导昼间树干液流速率的气象因子在各月差异不显著,主要为气温与太阳辐射。气温升高有助于加快叶片表面的水分散发,促进植物根系吸水从而增强植物蒸腾作用。但孙旭等[22]研究认为:土壤温度是影响油松树干液流的主导因子之一,与本研究的结果不一致。可能是研究的时间尺度、植被类型、植被覆盖度差异导致,因此关于树种耗水的研究需要综合考虑时间尺度与地下植被的影响。
目前,关于气象因子对夜间树干液流速率影响统一的结论是太阳辐射与夜间液流没有相关性。本研究结果显示:刺槐各月夜间液流速率与气温、水汽压亏缺、土壤温度、风速呈正相关,与相对湿度呈负相关。各月主导刺槐夜间树干液流速率的气象因子存在差异,5—6月土壤温度与水汽压亏缺是影响夜间树干液流速率的主导因子;7月气温是影响夜间树干液流速率的主要因子;8月水汽压亏缺与相对湿度是影响夜间树干液流速率的主导因子;9月风速与水汽压亏缺是影响夜间树干液流速率的主导因子。周翠鸣等[23]从季节尺度上分析了广州地区木荷Schima superba夜间树干液流和环境因子的关系,认为夜间液流产生的主要原因是树形特征和生物量而非气象因子;李洁等[24]对晋西北山地油松和落叶松Larix principis-rupprechtii研究发现:影响植物夜间液流速率的环境因子有气温、水汽压亏缺、土壤温度,其中土壤温度是主要驱动因子。可见,树种、地点都会影响树干液流速率对环境因子的响应。
-
随着研究的深入,普遍认为夜间液流主要用途为补充树体水分与植物蒸腾[8-9]。区分两者的方法主要有2种:一种为通过仪器测量比较树干上下枝液流量差异来判断,另一种为根据液流速率与环境因子的关系来判断。本研究采用第2种方法来区分各月刺槐夜间树干液流的主要作用。刺槐生长季各月夜间树干液流速率与水汽压亏缺均有相关性,表明各月夜间树干液流均有蒸腾作用。除7月外,水汽压亏缺是各月主导环境因子之一,说明5、6、8、9月夜间树干液流用于蒸腾比例较大,7月夜间树干液流用于树体补水比例较大,因为水汽压亏缺通过改变叶片的气孔导度来影响植物蒸腾,水汽压亏缺能很好地解释夜间液流变化则说明夜间液流主要为蒸腾。由于夜间蒸腾与补水是同步发生的,本研究根据气象因子只能分辨出夜间液流的主要作用,因此在今后的研究中应该探索植物生理特性,以便更好地了解植物的夜间液流机制。
-
在生长季,刺槐昼间与夜间树干液流速率特征具有规律性。刺槐夜间液流量对整日液流量的贡献率远远小于昼间液流的贡献率,但夜间树干液流弥补了昼间蒸腾造成的水分亏缺,因此夜间树干液流的重要性不可忽视。生长季不同月份影响刺槐昼夜蒸腾速率的主导气象因子存在差异,且构建昼夜液流速率与气象因子关系模型时,不同月份模型参数与因子不同。综上,生长季各月在根据气象因子构建蒸腾模型时需考虑昼夜差异。
Diurnal and nocturnal changes in stem sap flow of Robinia pseudoacacia during growing season and its response to meteorological factors
-
摘要:
目的 研究刺槐Robinia pseudoacacia生长季树干液流昼夜变化规律,探究气象因子对刺槐蒸腾的影响,为估算林木耗水和林分水资源管理提供理论依据。 方法 在山西省吉县蔡家川流域刺槐样地选择8株样树,于2021年5—9月采用热扩散探针(TDP)对样树树干液流进行连续观测,并同步监测太阳辐射、气温、土壤温度、风速、相对湿度等气象因子,并采用随机森林与逐步线性回归法分析气象因子对树干液流的影响。 结果 ①生长季各月昼间树干液流速率从小到大依次为9月、5月、6月、8月、7月,昼间树干液流速率对整日树干液流速率的贡献率为88%~93%;夜间树干液流速率从小到大依为9月、5月、8月、7月、6月,夜间树干液流速率对整日液流速率的贡献率为7%~12%。②影响昼间树干液流速率的主导气象因子在各月基本一致,主要为太阳辐射和气温;影响夜间液流速率的主导气象因子在各月存在差异,5、6月主要为土壤温度、水汽压亏缺,7月主要为气温,8月主要为水汽压亏缺、相对湿度,9月主要为风速与水汽压亏缺。③采用随机森林回归法构建的各月昼夜树干液流速率模型拟合度优于逐步回归法。 结论 昼夜树干液流速率在各月存在明显差异,夜间树干液流对刺槐耗水的影响不可忽视,气象因子对昼夜树干液流速率的影响程度不同,在根据气象因子模拟树木蒸腾耗水时应该区分昼间和夜间。图3表6参24 Abstract:Objective The objective is to analyze the diurnal and nocturnal changes in stem sap flow of Robinia pseudoacacia in the growing season and explore the impact of meteorological factors on transpiration of R. pseudoacacia , so as to provide theoretical basis for estimating forest water consumption and forest water resource management. Method Eight sample trees were selected from R. pseudoacacia sample plots in Caijiachuan watershed, Ji County of Shanxi Province. From May to September 2021, thermal diffusion probe (TDP) was used to continuously observe stem sap flow of sample trees, and meteorological factors such as solar radiation, air temperature, soil temperature, wind speed and relative humidity were monitored simultaneously. Random forest and stepwise linear regression were used to analyze the influence of meteorological factors on stem sap flow. Result (1) The order of diurnal sap flow rate from small to large in the growing season was September, May, June, August and July, and the contribution rate of diurnal sap flow to full-day sap flow was 88%−93%. The nocturnal sap flow rate ranging from small to large was September, May, August, July and June, and the contribution rate of nocturnal sap flow to full-day sap flow was 7%−12%. (2) The main meteorological factors affecting the diurnal sap flow rate were basically the same, mainly solar radiation and air temperature. The dominant meteorological factors affecting nocturnal sap flow were different in each month. The main factors were soil temperature and water vapor pressure deficit in May and June, air temperature in July, water vapor pressure deficit and relative humidity in August, and wind speed and water vapor pressure deficit in September. (3) The fitting degree of the monthly day-night flow rate model constructed by random forest regression method was better than that by stepwise regression method. Conclusion There are obvious differences in diurnal and nocturnal sap flow rate in each month. The effect of nocturnal sap flow on water consumption of R. pseudoacacia should not be ignored, and the effect of meteorological factors on diurnal and nocturnal sap flow rate is different. Daytime and nighttime should be distinguished when simulating water consumption of tree transpiration according to meteorological factors. [Ch, 3 fig. 6 tab. 24 ref.] -
图 3 气象因子对刺槐树干液流速率重要性排序
Figure 3 Meteorological factors of importance on the rate of flow
表 1 测定样树主要参数
Table 1. Main parameters of sample trees
样树编号 胸径/cm 树高/m 边材面积/cm2 样树编号 胸径/cm 树高/m 边材面积/cm2 1 22.5 10.2 142.2 5 16.8 9.5 85.2 2 21.9 11.2 141.5 6 18.0 11.0 93.4 3 20.6 9.8 138.7 7 17.4 10.3 90.2 4 21.3 11.2 140.2 8 18.2 10.8 92.6  下载: 导出CSV
下载: 导出CSV
表 2 生长季树干液流的昼夜特征
Table 2. Diurnal and nocturnal characteristics of sap flow rate in growing season
月份 昼间平均液流速率/
(cm3·h−1·cm−2)贡献
率/%夜间平均液流速率/
(cm3·h−1·cm−2)贡献
率/%5 2.71±1.07 91.26 0.29±0.23 8.74 6 2.94±1.05 91.16 0.43±0.21 8.84 7 3.24±0.85 92.25 0.35±0.17 7.75 8 3.18±1.61 92.52 0.33±0.18 7.48 9 1.74±1.26 88.39 0.27±0.12 11.61
下载: 导出CSV
表 3 整日与昼间刺槐树干液流量t检验
Table 3. Paired samples t test between daily and diurnal sap flow of R. pseudoacacia
月份 相关分析 配对样本t检验 R2 P t P 5 0.991 0.000 12.582 0.000 6 0.993 0.000 9.261 0.000 7 0.998 0.000 9.054 0.000 8 0.976 0.000 8.234 0.000 9 0.993 0.000 7.982 0.000
下载: 导出CSV
表 4 昼夜树干液流速率与气象因子Pearson相关系数
Table 4. Correlation analysis between diurnal and nocturnal sap flow rate of R. pseudoacacia and meteorological factors
昼夜 月份 太阳辐射 气温 土壤温度 水汽压亏缺 相对湿度 风速 昼间 5 0.575** 0.483** 0.429** 0.290** −0.278** 0.067 6 0.670** 0.539** 0.331** 0.389** −0.305** −0.031 7 0.762** 0.767** 0.497** 0.691** −0.722** 0.192** 8 0.689** 0.747** 0.546** 0.641** −0.638** 0.295** 9 0.674** 0.737** 0.608** 0.696** −0.692** 0.331* 夜间 5 0.582** 0.585** 0.556** −0.266** 0.351** 6 0.213** 0.224** 0.043** −0.049** 0.123** 7 0.512** 0.475** 0.483** −0.394** 0.338** 8 0.747** 0.546** 0.641** −0.638** 0.295** 9 0.141** 0.160** 0.107** −0.058** 0.215** 说明:**表示极显著相关(P<0.01); * 表示显著相关(P<0.05)
下载: 导出CSV
表 5 刺槐昼夜液流速率与气象因子逐步回归R2变化量
Table 5. Changes of stepwise regression R2 of R. pseudoacacia sap flow rate and meteorological factors
因子 逐步回归R2变化量 昼间 夜间 5月 6月 7月 8月 9月 5月 6月 7月 8月 9月 太阳辐射 − 0.449 0.095 0.015 0.040 − − − − − 气温 0.334 0.026 0.589 0.559 0.543 0.127 − 0.362 − 0.008 水汽压亏缺 0.152 0.150 0.016 0.029 − 0.057 0.148 0.005 0.360 0.232 相对湿度 0.087 − 0.032 0.053 − 0.014 0.084 0.050 0.117 0.008 土壤温度 − 0.007 0.007 0.074 0.014 0.342 0.350 − − − 风速 − − 0.002 0.029 0.018 0.042 − 0.158 0.030 0.246 说明:−表示气象因子未进入到逐步回归方程,以上数据均具有显著性
下载: 导出CSV
表 6 基于逐步回归法和随机森林回归法的刺槐昼夜树干液流与气象因子模型
Table 6. Models of day and night sap flow and meteorological factors based on stepwise regression method and random forest algorithm
昼夜 月份 逐步回归模型 随机森林回归法拟合度/% 模型 拟合度/% 昼间 5 y=−1.29+0.579x2−3.427x4−0.063x5 57.3 67.3 6 y=−0.767+0.003x1+0.301x2−0.149x3−0.869x4 63.2 73.2 7 y=1.509+0.002x1+0.444x2−0.194x3−1.862x4−0.053x5+0.071x6 74.1 86.3 8 y=1.864+0.001x1+0.506x2−0.208x3−2.468x4−0.065x5+0.144x6 75.9 90.6 9 y=1.104+0.001x1+0.300x2−0.204x3+0.297x6 61.5 81.1 夜间 5 y=2.852+0.002x1+0.182x2−0.279x3+0.154x6 48.2 59.4 6 y=4.943+0.003x1+0.346x2−0.363x3−1.252x4−0.025x5−0.110x6 61.6 67.7 7 y=3.966+0.003x1+0.415x2−0.209x3−2.094x4−0.069x5 57.5 61.6 8 y=4.869+0.002x1+0.486x2−0.311x3−2.296x4−0.074x5+0.091x6 50.7 63.9 9 y=−5.886+0.502x2−3.033x4 49.4 54.6 说明:y表示液流速率,x1表示太阳辐射,x2表示气温,x3表示土壤温度,x4表示水汽压亏缺,x5表示相对湿度,x6表示风速
下载: 导出CSV
-
[1] 代亚丽, 蔡江碧, 王宏丽. 植被建设在黄土高原生态环境建设中的地位和作用[J]. 西北农业大学学报, 2000, 28(6): 130 − 134. DAI Yali, CAI Jiangbi, WANG Hongli. The position and role of vegetation construction in ecological environment construction in Loess Plateau [J]. J Northwest Agric Univ, 2000, 28(6): 130 − 134. [2] 邵明安, 贾小旭, 王云强, 等. 黄土高原土壤干层研究进展与展望[J]. 地球科学进展, 2016, 31(1): 14 − 22. SHAO Ming’an, JIA Xiaoxu, WANG Yunqiang, et al. A review of studies on dried soil layers in the Loess Plateau [J]. Adv Earth Sci, 2016, 31(1): 14 − 22. [3] 孙长忠, 黄宝龙, 陈海滨, 等. 黄土高原人工植被与其水分环境相互作用关系研究[J]. 北京林业大学学报, 1998, 20(3): 10 − 17. SUN Changzhong, HUANG Baolong, CHEN Haibin, et al. Interaction between soil water conditions and different kinds of artificial plant cover in the Loess Plateau [J]. J Beijing For Univ, 1998, 20(3): 10 − 17. [4] LIU Hong, GUO Jianbin, LIU Zebin, et al. Time-lag effect between sap flow and environmental factors of Larix principis-rupprechtii Mayr [J]. Forests, 2019, 10(11): 971 − 971. [5] DALEY M J, PHILLIPS N. Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest [J]. Tree Physiol, 2006, 26(4): 411 − 419. [6] ZEPPEL M J B, TISSUE D T, TAYLOR D, et al. Rates of nocturnal transpiration in two evergreen temperate woodland species with differing water-use strategies [J]. Tree Physiol, 2010, 30(8): 988 − 1000. [7] 孔喆, 陈胜楠, 律江, 等. 欧美杨单株液流昼夜组成及其影响因素分析[J]. 林业科学, 2020, 56(3): 8 − 20. KONG Zhe, CHEN Shengnan, LÜ Jiang, et al. Characteristics of Populus euramericana sap flow over day and night and its influencing factors [J]. Sci Silv Sin, 2020, 56(3): 8 − 20. [8] 张婕, 蔡永茂, 陈立欣, 等. 北京山区元宝枫夜间液流活动特征及影响因素[J]. 生态学报, 2019, 39(9): 3210 − 3223. ZHANG Jie, CAI Yongmao, CHEN Lixin, et al. Influencing factors and characteristics of nighttime sap flow of Acer truncatum in Beijing mountainous area [J]. Acta Ecol Sin, 2019, 39(9): 3210 − 3223. [9] 方伟伟, 吕楠, 傅伯杰. 植物夜间液流的发生、生理意义及影响因素研究进展[J]. 生态学报, 2018, 38(21): 7521 − 7529. FANG Weiwei, LÜ Nan, FU Bojie. Research advances in nighttime sap flow density, its physiological implications, and influencing factors in plants [J]. Acta Ecol Sin, 2018, 38(21): 7521 − 7529. [10] 任启文, 毕君, 李联地, 等. 冀北山地油松树干液流特征及其对环境因子的响应[J]. 水土保持学报, 2017, 31(5): 183 − 188. REN Qiwen, BI Jun, LI Liandi, et al. Stem sap flow characteristics of Pinus tabulaeformis and its response to environmental factors at northern mountain areas of Hebei Province [J]. J Soil Water Conserv, 2017, 31(5): 183 − 188. [11] 贾国栋, 陈立欣, 李瀚之, 等. 北方土石山区典型树种耗水特征及环境影响因子[J]. 生态学报, 2018, 38(10): 3441 − 3452. JIA Guodong, CHEN Lixin, LI Hanzhi, et al. The effect of environmental factors on plant water consumption characteristics in a northern rocky mountainous area [J]. Acta Ecol Sin, 2018, 38(10): 3441 − 3452. [12] 徐利岗, 苗正伟, 杜历, 等. 干旱区枸杞树干液流变化特征及其影响因素[J]. 生态学报, 2016, 36(17): 5519 − 5527. XU Ligang, MIAO Zhengwei, DU Li, et al. Analysis of variation in and factors influencing sap flow in stems of Lycium barbarum in an arid area [J]. Acta Ecol Sin, 2016, 36(17): 5519 − 5527. [13] 方匡南, 吴见彬, 朱建平, 等. 随机森林方法研究综述[J]. 统计与信息论坛, 2011, 26(3): 32 − 38. FANG Kuangnan, WU Jianbin, ZHU Jianping, et al. A review of random forest methods [J]. Stat Inf Forum, 2011, 26(3): 32 − 38. [14] 赵楠. 干旱绿洲区葡萄和白杨树干液流动态及对环境因子的响应[D]. 兰州: 兰州大学, 2021. ZHAO Nan. Sap Flow Dynamics and Environmental Responses of Grape and Poplar in Arid Oasis[D]. Lanzhou: Lanzhou University, 2021. [15] 凡超, 邱燕萍, 李志强, 等. 荔枝树干液流速率与气象因子的关系[J]. 生态学报, 2014, 34(9): 2401 − 2410. FAN Chao, QIU Yanping, LI Zhiqiang, et al. Relationships between stem sap flow rate of litchi trees and meteorological parameters [J]. Acta Ecol Sin, 2014, 34(9): 2401 − 2410. [16] 吴旭, 陈云明, 唐亚坤. 黄土丘陵区刺槐和侧柏人工林树干液流特征及其对降水的响应[J]. 植物生态学报, 2015, 39(12): 1176 − 1187. WU Xu, CHEN Yunming, TANG Yakun. Sap flow characteristics and its responses to precipitation in Robinia pseudoacacia and Platycladus orientalis plantations [J]. Chin J Plant Ecol, 2015, 39(12): 1176 − 1187. [17] 王志超, 许宇星, 竹万宽, 等. 雷州半岛尾叶桉人工林夜间耗水特征及驱动因素[J]. 浙江农林大学学报, 2020, 37(4): 646 − 653. WANG Zhichao, XU Yuxing, ZHU Wankuan, et al. Characteristics and driving factors of nocturnal water consumption of Eucalyptus urophylla plantations in Leizhou Peninsula [J]. J Zhejiang A&F Univ, 2020, 37(4): 646 − 653. [18] HAYAT M, IQBAL S, ZHA T, et al. Biophysical control on nighttime sap flow in Salix psammophila in a semiarid shrubland ecosystem[J/OL]. Agric For Meteorol, 2021, 300(4): 108329[2022-01-10]. doi. org/10.1016/j. agrformet. 2021.108329. [19] 徐志彬, 陈胜楠, 陈立欣, 等. 半干旱区油松林分夜间液流变化特征及其影响因子[J]. 中国水土保持科学, 2021, 19(5): 37 − 44. XU Zhibin, CHEN Shengnan, CHEN Lixin, et al. Nocturnal sap flow variations and its influencing factors of Pinus tabulaeformis stand in a semi-arid environment [J]. Sci Soil Water Conserv, 2021, 19(5): 37 − 44. [20] 刘崴, 魏天兴, 朱清科. 半干旱黄土丘陵区河北杨和油松生长季树干液流特征[J]. 浙江农林大学学报, 2018, 35(6): 1045 − 1053. LIU Wei, WEI Tianxing, ZHU Qingke. Growing season sap flow of Populus hopeiensis and Pinus tabulaeformis in the semi-arid Loess Plateau, China [J]. J Zhejiang A&F Univ, 2018, 35(6): 1045 − 1053. [21] YIN Lihe, ZHOU Yangxiao, HUANG Jinting, et al. Dynamics of willow tree (Salix matsudana ) water use and its response to environmental factors in the semi-arid Hailiutu River catchment, Northwest China [J]. Environ Earth Sci, 2014, 71(12): 4997 − 5006. [22] 孙旭, 杨文慧, 焦磊, 等. 不同时间尺度北京蟒山油松树干液流对环境因子的响应研究[J]. 生态学报, 2022, 42(10): 1 − 11. SUN Xu, YANG Wenhui, JIAO Lei, et al. Relationship between sap flux density of Pinus tabulaeformis trees and environmental factors at different temporal scales in Mangshan National Forest Park of Beijing [J]. Acta Ecol Sin, 2022, 42(10): 1 − 11. [23] 周翠鸣, 赵平, 倪广艳, 等. 广州地区荷木夜间树干液流补水的影响因子及其对蒸腾的贡献[J]. 应用生态学报, 2012, 23(7): 1751 − 1757. ZHOU Cuiming, ZHAO Ping, NI Guangyan, et al. Water recharge through nighttime stem sap flow of Schima superba in Guangzhou region of Guangdong Province, South China: affecting factors and contribution to transpiration [J]. Chin J Appl Ecol, 2012, 23(7): 1751 − 1757. [24] 李洁, 任启文, 马香玲, 等. 冀西北山地油松和落叶松树干液流昼夜变化特征及其与环境因子的关系[J]. 东北林业大学学报, 2018, 46(7): 18 − 22. LI Jie, REN Qiwen, MA Xiangling, et al. Daytime and nighttime variation of sap flow velocity for Pinus tabulaeformis and Larix principis-rupprechtii and its relationship with environmental factors in northwest mountain of Hebei Province [J]. J Northeast For Univ, 2018, 46(7): 18 − 22. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220159
 点击查看大图
点击查看大图

计量
- 文章访问数: 1699
- HTML全文浏览量: 269
- PDF下载量: 73
- 被引次数: 0


