






-
花发育是植物完成生命周期的关键过程。植物从营养生长到生殖生长的转变是对环境因素和遗传因素的双重响应,为了确保子代的正常繁殖,高等植物必须在最适的环境条件下开花[1]。花发育分为成花诱导、成花启动及花器官发育共3个阶段[1]。植物通过外界环境和内源激素变化感受到开花信号,刺激茎顶端分生组织(shoot apical meristem,SAM)转变为花序分生组织 (inflorescence meristem,IM),随后花序分生组织在花分生组织特征基因(floral meristem identity gene)和花器官特征基因 (floral organ identity gene)的作用下形成一朵完整的花[2]。F-box蛋白是植物中最大的蛋白质超家族之一,N端存在F-box结构域,该结构域由40~60个氨基酸残基组成,能和Skp1、Cullinl (CUL1)/Cdc53和Rbxl/Rocl/Hrtl结合形成Skp1-Cullin1-F-box (SCF)复合体参与泛素化过程;C端为底物结合区域,存在不同的结构域,包括Kelch、LRR、FBD结构域等,F-box蛋白的C端决定了底物识别的特异性,根据结合底物的不同,F-box蛋白发挥不同的作用[3-5]。可见,F-box蛋白形成的SCF复合体能参与植物生命周期的各个方面,如种子萌发、花发育、自交不亲和性、生物胁迫和非生物胁迫以及光形态建成等[3, 6]。
UNUSUAL FLORAL ORGANS (UFO)基因是与花发育相关的F-box基因,是重要的花分生组织特征基因和花器官特征基因,依赖于LEAFY (LFY)基因发挥作用,并与LFY基因共同促进ABC模型中B类基因的表达,从而调控花瓣和雄蕊的发育[7-12]。除了LFY基因,UFO还能与ASK1结合形成 SCFUFO复合体对花瓣和雄蕊发育造成影响,ufo和ask1突变体都表现为花瓣和雄蕊发育不良[11]。在拟南芥Arabidopsis thaliana中UFO基因C末端区域是花瓣发育所必需的,而在百脉根Lotus japonicus和豌豆Pisum sativum中,UFO同源基因PROLIFERATING FLORAL ORGANS (PFO)和STAMINA PISTILLOIDA(STP)基因C端序列的缺失不仅对花瓣发育有影响,还对花分生组织确定性造成影响,形成次生花序[13-15]。
龙葵Solanum nigrum是茄科Solanaceae茄属Solanum的1年生草本植物,花序结构为聚伞花序[16]。目前对茄科的研究集中在番茄S. lycopersicum等经济作物上,在其他茄科植物中关于花发育的研究较为欠缺。龙葵生长周期短、植株矮小、遗传转化效率高且便于实验室栽培,是理想的研究材料。通过对龙葵花发育的研究可以丰富茄科植物花发育的研究内容,探讨UFO基因在不同物种的调控机制,为UFO基因在茄科花发育的调控方面提供理论支持。
-
以野生型植株及其转基因植株为材料,在室温25 ℃及光周期16 h/8 h的条件下栽培,备用。
-
根据龙葵转录组数据筛选出SnUFO2基因序列,设计上下游引物并利用在线网站NEBcutter (
http://tools.neb.com/NEBcutter/index.php3 )添加酶切位点BamHⅠ和SacⅠ以及保护碱基(表1)。引物序列送杭州有康生物科技有限公司合成。表 1 基因克隆及RT-qPCR引物列表
Table 1. Gene cloning and RT-qPCR primer list
引物名称 上游引物(5′→3′) 下游引物(5′→3′) SnUFO2 AAGGATCCATGGAAGCTTTTCATCATCCC AAGAGCTCTCAGTTGAAAGACTGAAAGGG qSnUFO2 GCTGTGGCTGGTGATAACTTG CGGCATACGGGCAATTTCTT SnAPRT GAGATGCATGTAGGTGCTGTGCAA GGCCCTTCAATTCTGGCAACTCAA 说明:下划线处为酶切位点,分别为BamHⅠ和SacⅠ 利用RNA提取试剂盒(普洛麦格生物产品有限公司,上海)提取盛花期野生型龙葵植株相同生长时期花苞的RNA,使用EasyScript®First-Strand cDNA Synthesis SuperMix反转录试剂盒(全式金生物技术有限公司,北京)获得cDNA,以其为模板克隆SnUFO2基因。PCR反应程序:97 ℃预变性3 min;95 ℃变性40 s,60 ℃退火40 s,72 ℃延伸1 min,35个循环;72 ℃总延伸10 min。PCR产物进行琼脂糖凝胶电泳检测片段大小,并纯化回收目的片段。目的片段连接到pEASY Blunt simple (全式金生物技术有限公司,北京)载体后,转入大肠埃希菌Escherichia coli DH5α 感受态中(唯地生物技术有限公司,上海)。通过菌落PCR筛选阳性单菌落,摇菌送杭州擎科生物有限公司测序。测序返回数据拼接后与转录组测序序列进行比对,获得2条目的基因序列。
-
通过在线网站Pfam (
http://pfam.xfam.org/ )蛋白质家族数据库进行结构域分析。将克隆得到的2条SnUFO2序列在美国国家生物信息中心(NCBI)上通过Blastx进行同源基因检索。利用MEGA7[17]进行氨基酸序列比对和系统进化树构建。进化树构建采用邻接法,Bootstrap检验1 000次。将筛选到的不同物种中UFO基因和龙葵中2条SnUFO2序列输入在线网站MEME Suite (https://meme-suite.org/meme/ ),选择合适的Motif数量后导出结果,并用TBtools[18]软件美化。 -
于野生型龙葵植株盛花期取样,分别采集植株不同部位,包括根、茎、叶和花苞并提取RNA,以反转录后的cDNA为模板,进行实时荧光定量PCR反应。内参基因为龙葵的APRT基因,SnAPRT和SnUFO2*定量引物如表1所示。根据RT-qPCR试剂盒TransStart® Tip Green qPCR SuperMix(全式金生物技术有限公司,北京)说明书设置反应体系:cDNA 1 μL ,上下游引物(10 μmol·L−1)各0.2 μL,2×TransStart® Tip Green qPCR SuperMix 5 μL,双蒸水补足至10 μL。反应条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火15 s,72 ℃延伸10 s,共反应45个循环。数据计算方法采用
$2^{ - \Delta \Delta C_t}$ 法计算相对表达量。 -
利用BamHⅠ和SacⅠ(赛默飞世尔科技公司,上海)将目的片段和pBI121载体进行双酶切,纯化回收后在16 ℃下过夜连接,并转入大肠埃希菌感受态。菌落经PCR验证后,提取阳性重组质粒,通过单双酶切验证超表达载体。
-
SDS法提取转化苗基因组DNA,以野生型龙葵为对照,通过PCR检测阳性转基因植株。通过尼康相机(COOLPIX P7100)拍照记录转基因植株和花序表型变化,Leica体视显微镜(M165FC)拍照记录转基因植株花表型变化。
-
采用海氏铁矾-苏木精染色法[19],在野生型与转基因植株盛花期取2.5 mm左右的花苞置于FAA固定液中固定,依次经50%、60%、70%、80%、90%、95%、100%体积分数的乙醇脱水,再依次经体积梯度比1∶2、1∶1、2∶1的二甲苯无水乙醇混合液和纯二甲苯透明后,浸蜡3 d,使石蜡缓慢进入材料中,将材料包埋至蜡块中。使用Leica转轮式切片机(RM2235)切片,用苏木精染色后封片。
-
以野生型龙葵盛花期花苞的cDNA为模板,克隆得到2条SnUFO2序列,都含有F-box结构域。一条序列与转录组测序结果一致,为龙葵中SnUFO2基因,编码456个氨基酸;另一条序列为短截版的SnUFO2基因,该基因在1 294~1 295 bp处有1个碱基G的插入,导致翻译提前终止,编码433个氨基酸,记为SnUFO2*。将SnUFO2*基因、全长SnUFO2基因和不同物种中UFO的同源基因构建系统进化树。SnUFO2*、SnUFO2基因和番茄、辣椒Capsicum annuum和矮牵牛Petunia × hybrida中的UFO基因在同一进化分支,同源关系较近,而与水稻Oryza sativa,拟南芥中的UFO基因同源关系较远(图1A)。

图 1 SnUFO2*系统发育树分析及Motif分析
Figure 1. SnUFO2* phylogenetic tree analysis and Motif analysis
利用MEME软件对SnUFO2*和SnUFO2蛋白进行Motif预测发现,这些蛋白相似性极高,预测到高度相似的Motif可能行使UFO蛋白最保守的功能(图1B)。SnUFO2*与其他UFO蛋白相比,Motif 8的缺失可能导致该基因的功能与其他物种中UFO基因的功能存在差异。通过氨基酸序列比对发现:SnUFO2*C末端比SnUFO2基因少了23个氨基酸,这段序列在茄科物种中高度保守,可能影响龙葵正常的花发育进程(图2)。

图 2 龙葵SnUFO2*氨基酸序列比对
Figure 2. Amino acid sequence alignment of S. nigrum SnUFO2*
-
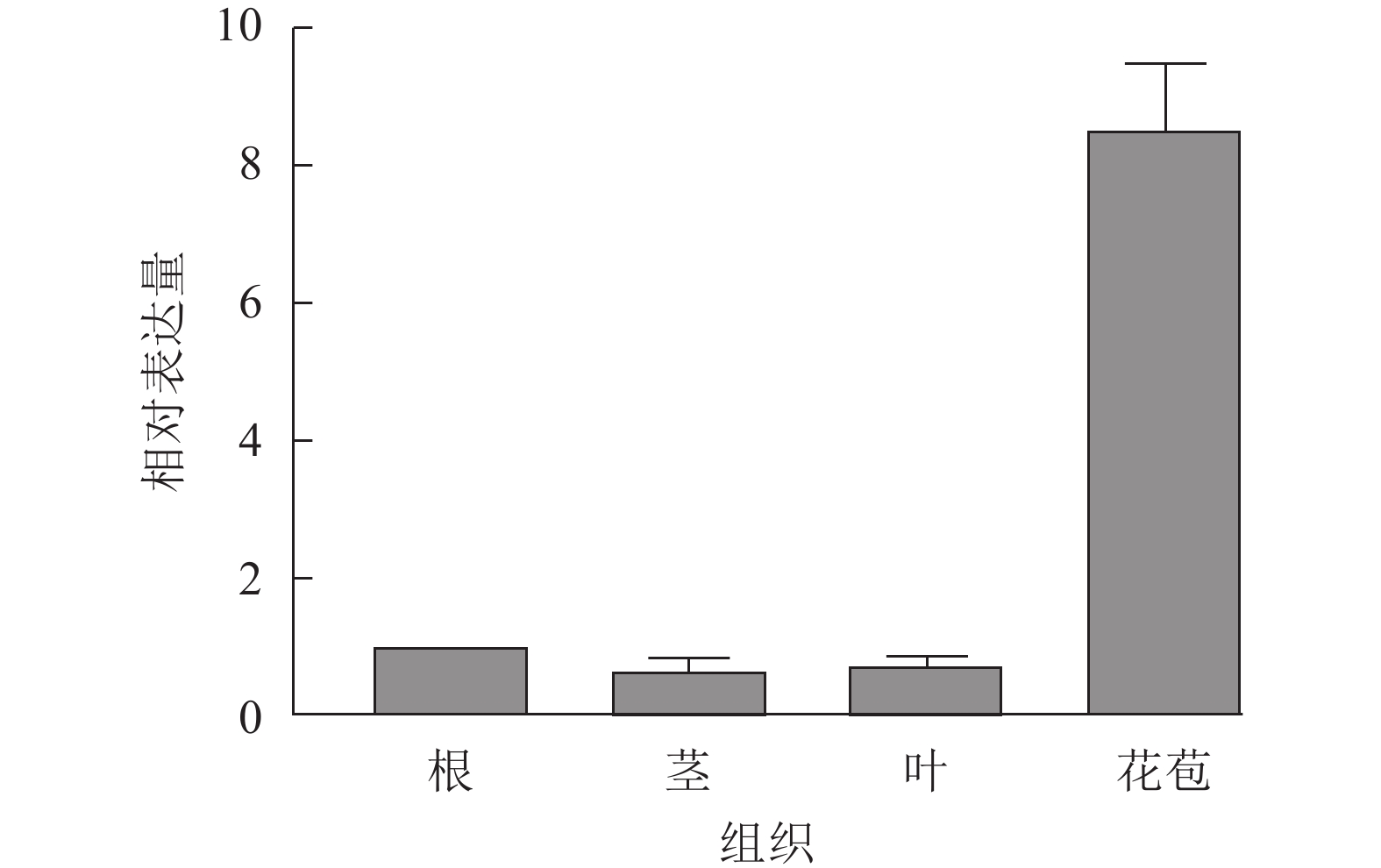
UFO基因是重要的花器官特征基因,在花发育初期发挥作用。为了探究SnUFO2*基因在龙葵营养器官及生殖器官中的表达水平,提取盛花期的根、茎、叶和花苞进行RT-qPCR分析。以龙葵APRT为内参基因,RT-qPCR结果表明:SnUFO2*在根、茎、叶中的表达量低,在花苞中相对表达量较高,推测SnUFO2*基因可能主要参与花器官发育(图3)。
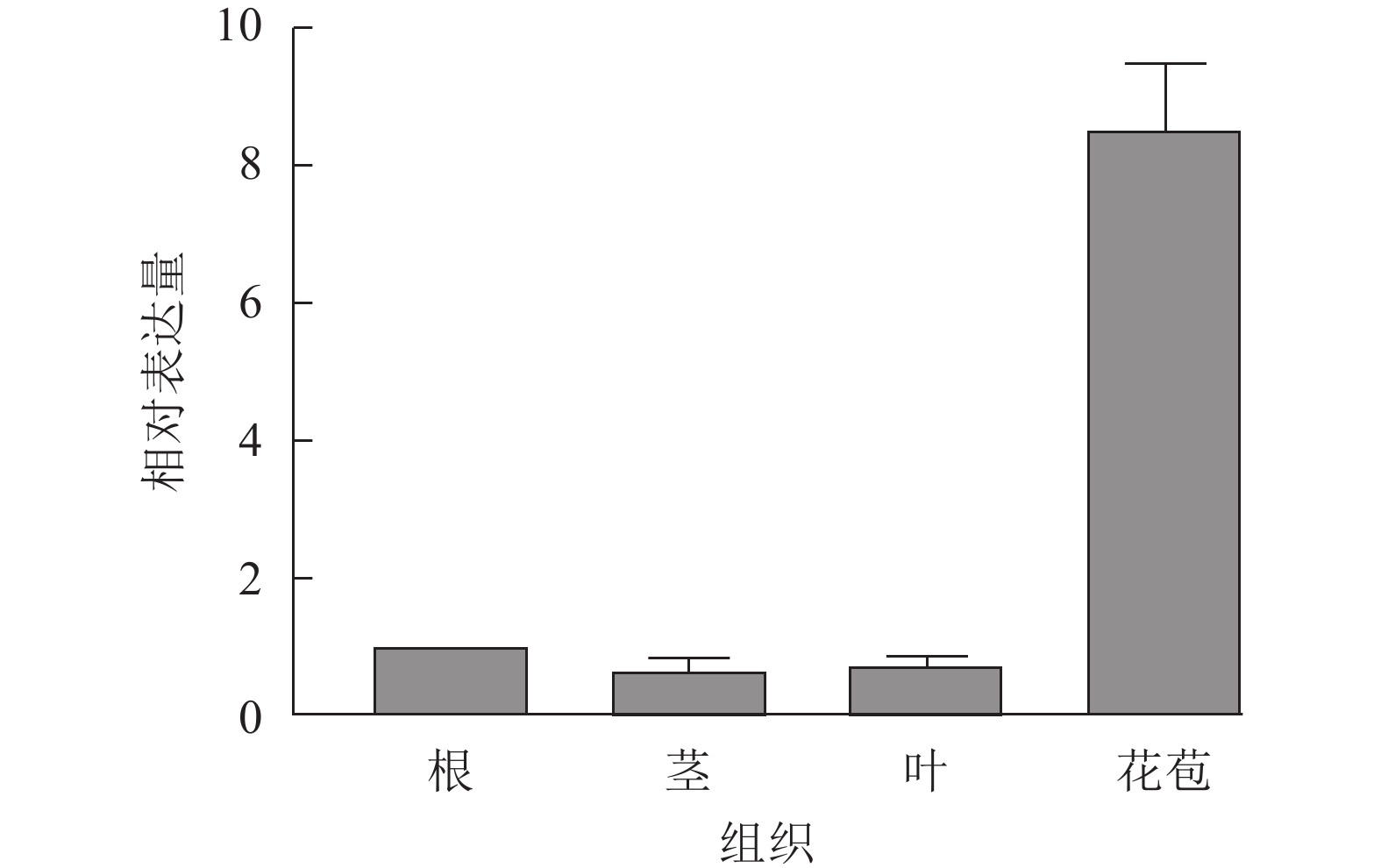
图 3 野生型龙葵中SnUFO2*不同组织部位表达量分析
Figure 3. Expression analysis of SnUFO2* in different tissues of wild-type S. nigrum
-
为探究龙葵C末端缺失的SnUFO2*基因对龙葵花发育的影响,构建了SnUFO2*的表达载体,将其转入龙葵植株中,经含有卡那霉素抗性的培养基筛选后,获得40个T0代独立抗性株系,经PCR鉴定共获得31个转基因阳性株系。选择表型明显的不同株系转基因植株进行分析,转基因植株的根、茎和叶等未观察到明显变化(图4A),而花器官发育异常:野生型龙葵花盛开后,花瓣呈白色,花瓣基部相连,雄蕊紧靠雌蕊生长于花中心(图4C);弱表型转基因SnUFO2-30株系的花部分花瓣中部形成绿色条纹状组织,偶尔形成萼片状花瓣,花瓣基部裂口变大,雄蕊花药未紧靠雌蕊,杂乱分布于四周(图4B和C);强表型SnUFO2-10株系的花瓣完全萼片化,这类花最终不能形成正常的果实和种子(图4A~C)。

图 4 35S::SnUFO2* 转基因植株表型图
Figure 4. Phenotypic map of 35S::SnUFO2* transgenic plants
-
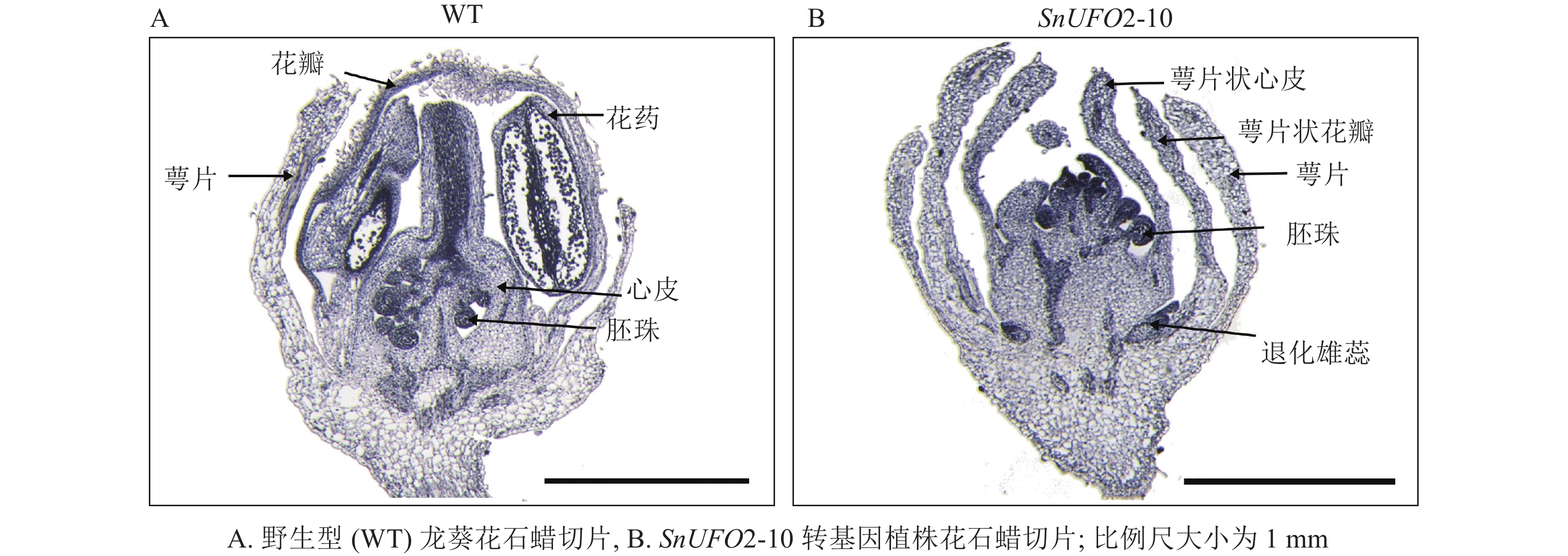
为了进一步确定转基因植株花内部结构及其细胞变化,通过对野生型和SnUFO2-10转基因植株的花苞进行石蜡切片表明:野生型株系的花苞由外到内依次存在萼片、花瓣、雄蕊和雌蕊;SnUFO2-10株系花苞生成萼片状花瓣,雄蕊缺失,心皮发育异常,没有花柱和柱头产生,偶尔在发育中的心皮两侧观察到胚珠的产生(图5)。
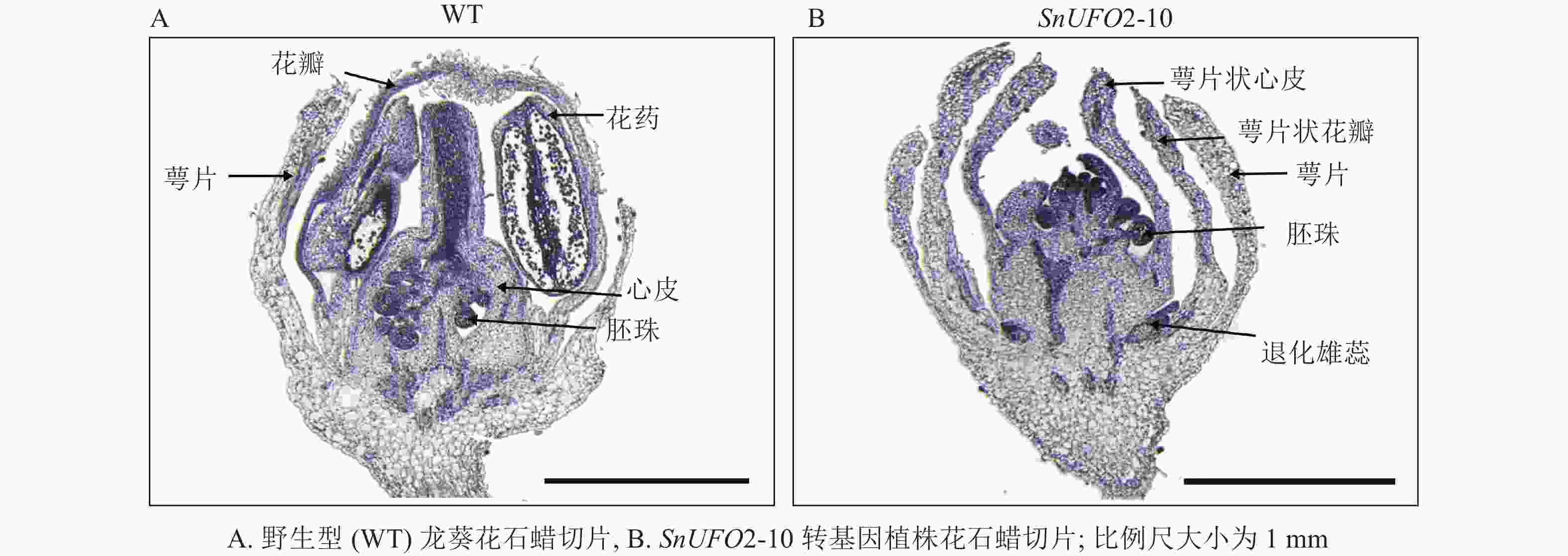
图 5 野生型及35S::SnUFO2*转基因株系花苞石蜡切片
Figure 5. Paraffin sections of flower buds of wild-type and 35S::SnUFO2 * transgenic lines
-
本研究克隆了1条短截版的SnUFO2基因,探究C端序列的完整性对龙葵花发育的影响。生物信息学分析发现:SnUFO2*基因属于F-box基因家族,且C末端缺失的序列可能具有保守的功能。F-box蛋白家族通常以SCF复合体的形式参与植物各项生命活动。过往研究认为F-box蛋白的N端与SKP1类基因结合,形成SCF复合体,而C端与靶蛋白结合,通过泛素链引导至26S蛋白酶体从而降解结合蛋白[3]。基于C端序列在F-box基因中的重要作用推测,SnUFO2* C末端23个保守的氨基酸缺失可能会影响泛素化过程。
UFO及其同源基因已被证明在植物花发育过程中发挥作用[7-15]。通过RT-qPCR分析发现:SnUFO2*基因在龙葵的根、茎和叶中表达水平较低,而在花苞中表达量较高,推测该基因和其他UFO基因一样在花发育过程中发挥作用。于是构建SnUFO2*超表达载体并将其转入龙葵中。在矮牵牛中过表达DOUBLE TOP(DOT)基因会导致植株矮化,形成一朵单花[20]。然而形态学观察发现:35S::SnUFO2*转基因植株未出现矮化表型,根、茎和叶也无明显变化,但是花器官发生明显变化,萼片内侧的花瓣、雄蕊和雌蕊都被萼片状的器官代替。这与C末端缺失的UFO基因突变体表型相似,在拟南芥ufo-2突变体中,UFO基因翻译提前终止,编码262个氨基酸,产生了强烈的表型变化:生成萼片状花瓣、花丝及心皮状结构[21]。在豌豆突变体,stp-4中,STP基因只编码了252个氨基酸,导致豌豆花缺少花瓣和雄蕊,且有次生花产生[15]。黄瓜Cucumis sativus ufo突变体中,其花瓣的位置产生了叶状器官,雄蕊发育不正常[22]。以上研究表明:UFO基因的C末端对该基因的功能具有重要作用,虽然不同C末端缺失突变体表型不完全相同,但这可能与该基因短截的位点不同有关,也可能与物种特性有关。
花发育是一个受多基因调控的复杂生理过程,需要相关花分生组织特征基因和花器官特征基因的共同作用。SnUFO2-10花苞的石蜡切片结果显示:在4轮花器官形成过程中,除最外轮萼片部分形成正常外,花瓣萼片化,花分生组织不确定导致内轮不断产生增殖的萼片状器官。缺少C末端的 ufo突变体的共同特征为花瓣和雄蕊的缺失或是畸形发育,例如拟南芥ufo-2花瓣萼片化及雄蕊数量减少,豌豆stp突变体表现为花瓣向萼片转化[15, 21]。其次是心皮发育异常或花分生组织的不确定性,例如金鱼草fimbriata620 (fim620)突变体产生的侧生花及花的萼片数目不确定和百脉根 pfo突变体产生的不断增殖的萼片状器官[14, 23]。这些表型的出现与花器官特征基因中的B类和C类基因有关,因此推测UFO基因C端序列的缺失对该基因的功能造成影响,从而直接或间接影响B类和C类基因的正常表达。过往研究表明:SCFUFO可能促进LFY基因的转录活性,且UFO基因能与LFY共同促进B、C类基因的表达[11-12, 20-21]。我们认为:SnUFO2*可能通过SCF复合体的形式,影响LFY基因的表达进而促进相关花同源异型基因的表达,然而SnUFO2*基因C末端序列的缺失影响了SCF复合体的功能,从而影响4轮花器官的正常发育。转基因植株形成的萼片状花与lfy突变体表型相似,间接证明了这种猜想[11]。
综上所述,不仅是UFO基因的F-box结构域对其功能具有重要作用,C端序列的完整性对基因功能也极其重要。之后的研究中,观察全长SnUFO2转基因植株表型的变化,尤其是花器官发育变化,对深入探究UFO基因的功能是必要的。
Effect of conservation of C-terminal sequence of Solanum nigrum UNUSUAL FLORAL ORGANS family SnUFO2 on flower development
-
摘要:
目的 UNUSUAL FLORAL ORGANS (UFO)基因属于F-box基因家族,是重要的花器官特征基因。UFO基因N端能与Skp1类基因结合形成Skp1-Cullin1-F-box (SCF)复合体,参与泛素化过程并降解C端结合的靶蛋白。为了探究C端序列对龙葵Solanum nigrum花发育的影响,本研究克隆了一个C末端缺失的SnUFO2*基因并构建其表达载体转入龙葵植株中,观察转基因龙葵植株花器官变化,从而深入探讨UFO基因完整的C末端序列在龙葵花发育中的重要作用。 方法 利用生物信息学分析软件对SnUFO2*和全长的SnUFO2比较分析,采用实时荧光定量PCR(RT-qPCR)对SnUFO2*基因在野生型龙葵植株根、茎、叶、花苞中进行表达分析;通过超表达载体的构建、转基因植株表型的观察及石蜡切片技术验证SnUFO2基因的功能。 结果 SnUFO2*基因ORF长度为1302 bp,编码433个氨基酸,与龙葵中完整的SnUFO2基因相比,其C末端缺失了23个氨基酸。RT-qPCR结果显示:SnUFO2*基因在野生型植株的花苞中特异性表达。对转基因植株的表型观察发现:35S:: SnUFO2*转基因龙葵植株的花瓣向萼片转化。石蜡切片分析发现:转基因龙葵植株雄蕊缺失,雌蕊处有不确定的分生组织产生。 结论 35S:: SnUFO2*转基因龙葵植株花瓣、雄蕊和心皮发育异常。C端结构缺失可能降低了SnUFO2蛋白特异性识别靶蛋白的能力,说明该基因完整的C末端对龙葵花器官发育至关重要。图5表1参23 Abstract:Objective UNUSUAL FLORAL ORGANS (UFO) gene belongs to the F-box gene family and is one of the most important floral organ identity genes. The N-terminal of UFO gene can combine with Skp1 genes to form Skp1-Cullin1-F-box (SCF) complex, which participates in the ubiquitination process and degrades the target protein binding with the C-terminal. In order to explore the effect of C-terminal sequence on the flower development of Solanum nigrum, a C-terminal deleted SnUFO2* gene was cloned and its expression vector was constructed transferred into S. nigrum plants to observe the changes of floral organs of transgenic S. nigrum plants, so as to further explore the important role of the complete C-terminal sequence of UFO gene in the flower development of S. nigrum. Method Comparative analysis of SnUFO2* and full-length SnUFO2 was performed using bioinformatics analysis software. Moreover, the expression levels of SnUFO2* in roots, stems, leaves, and buds of wild-type S. nigrum were analyzed by real-time quantitative PCR (RT-qPCR); the function of the SnUFO2* gene was verified by the construction of over expression vector, observation of phenotype of transgenic plants, and the employment of paraffin section technique. Result The ORF length of SnUFO2* was 1 302 bp, encoding 433 amino acids, when compared with the complete SnUFO2 in S. nigrum, the C-terminal of which was missing 23 amino acids. RT-qPCR results showed that SnUFO2* was specifically expressed in the buds of wild-type plants. The phenotypic observation of transgenic plants demonstrated that the petals of 35S::SnUFO2* transgenic S. nigrum plants transformed into sepals. Paraffin section analysis presented that the stamens of transgenic S. nigrum plants were missing and uncertain meristem was produced in the pistil. Conclusion 35S::SnUFO2* transgenic plants lead to abnormal development of petals, stamens, and carpels of S. nigrum. The deletion of the C-terminal structure may reduce the ability of SnUFO2 protein to specifically recognize target proteins, indicating that the complete C-terminal of the gene is important for organ development of S. nigrum. [Ch, 5 fig. 1 tab. 23 ref. ] -
Key words:
- transgenic Solanum nigrum /
- SnUFO2 /
- floral meristem /
- floral organ development /
- flower development
-
图 1 SnUFO2*系统发育树分析及Motif分析
Figure 1 SnUFO2* phylogenetic tree analysis and Motif analysis
图 3 野生型龙葵中SnUFO2*不同组织部位表达量分析
Figure 3 Expression analysis of SnUFO2* in different tissues of wild-type S. nigrum
图 5 野生型及35S::SnUFO2*转基因株系花苞石蜡切片
Figure 5 Paraffin sections of flower buds of wild-type and 35S::SnUFO2 * transgenic lines
表 1 基因克隆及RT-qPCR引物列表
Table 1. Gene cloning and RT-qPCR primer list
引物名称 上游引物(5′→3′) 下游引物(5′→3′) SnUFO2 AAGGATCCATGGAAGCTTTTCATCATCCC AAGAGCTCTCAGTTGAAAGACTGAAAGGG qSnUFO2 GCTGTGGCTGGTGATAACTTG CGGCATACGGGCAATTTCTT SnAPRT GAGATGCATGTAGGTGCTGTGCAA GGCCCTTCAATTCTGGCAACTCAA 说明:下划线处为酶切位点,分别为BamHⅠ和SacⅠ  下载: 导出CSV
下载: 导出CSV
-
[1] CHO L H, YOON J, AN G. The control of flowering time by environmental factors [J]. The Plant Journal, 2017, 90(4): 708 − 719. [2] 杨传平, 刘桂丰, 魏志刚. 高等植物成花基因的研究[J]. 遗传, 2002, 24(3): 379 − 384. YANG Chuanping, LIU Guifeng, WEI Zhigang. Studies of flowering genes of plants [J]. Hereditas, 2002, 24(3): 379 − 384. [3] ABD-HAMID N A, AHMAD-FAUZI M I, ZAINAL Z, et al. Diverse and dynamic roles of F-box proteins in plant biology[J/OL]. Planta, 2020, 251(3): 68[2022-01-20]. doi: 10.1007/s00425-020-03356-8. [4] XU Guixia, MA Hong, NEI M, et al. Evolution of F-box genes in plants: different modes of sequence divergence and their relationships with functional diversification [J]. Proceedings of the National Academy of Sciences, 2009, 106(3): 835 − 840. [5] MO Fulei, ZHANG Nian, QIU Youwen, et al. Molecular characterization, gene evolution and expression analysis of the F-box gene family in tomato (Solanum lycopersicum)[J/OL]. Genes, 2021, 12(3): 417[2022-01-20]. doi: 10.3390/genes12030417. [6] ZHANG Shulin, TIAN Zailong, LI Haipeng, et al. Genome-wide analysis and characterization of F-box gene family in Gossypium hirsutum L. [J/OL]. BMC Genomics, 2019, 20(1): 993[2022-01-20]. doi: 10.1186/s12864-019-6280-2. [7] LEVIN J Z, MEYEROWITZ E M. UFO: an Arabidopsis gene involved in both floral meristem and floral organ development [J]. Plant Cell, 1995, 7(5): 529 − 548. [8] LEE I, WOLFE D S, NILSSON O, et al. A LEAFY co-regulator encoded by UNUSUAL FLORAL ORGANS [J]. Current Biology, 1997, 7(2): 95 − 104. [9] SAMACH A, KLENZ J E, KOHALMI S E, et al. The UNUSUAL FLORAL ORGANS gene of Arabidopsis thaliana is an F-box protein required for normal patterning and growth in the floral meristem[J]. The Plant Journal, 1999, 20(4): 433 − 445. [10] RISSEEUW E, VENGLAT P, XIANG Daoquan, et al. An activated form of UFO alters leaf development and produces ectopic floral and inflorescence meristems[J/OL]. PLoS One, 2013, 8(12): e83807[2022-09-23]. doi: 10.1371/journal.pone.0083807. [11] ZHAO Dazhong, YU Qilu, CHEN Min, et al. The ASK1 gene regulates B function gene expression in cooperation with UFO and LEAFY in Arabidopsis[J]. Development, 2001, 128(14): 2735 − 2746. [12] CHAE E, TAN Q K, HILL T A, et al. An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development [J]. Development, 2008, 135(7): 1235 − 1245. [13] DURFEE T, ROE J L, SESSIONS R A, et al. The F-box-containing protein UFO and AGAMOUS participate in antagonistic pathways governing early petal development in Arabidopsis[J]. Proceedings of the National Academy of Sciences, 2003, 100(14): 8571 − 8576. [14] ZHANG Shulu, SANDAL N, POLOWICK P L, et al. PROLIFERATING FLORAL ORGANS (PFO), a Lotus japonicus gene required for specifying floral meristem determinacy and organ identity, encodes an F-box protein[J]. The Plant Journal, 2003, 33(4): 607 − 619. [15] TAYLOR S, HOFER J, MURFET I. STAMINA PISTILLOIDA, the pea ortholog of FIM and UFO, is required for normal development of flowers, inflorescences, and leaves[J]. Plant Cell, 2001, 13(1): 31 − 46. [16] 孙皎, 倪彦博, 冯舒, 等. 不同栽培条件对龙葵生物碱积累的影响[J]. 北方园艺, 2017(13): 157 − 160. SUN Jiao, NI Yanbo, FENG Shu, et al. Effect of different cultivation conditions on accumulation of alkaloids of Solalum nigrum [J]. Northern Horticutlure, 2017(13): 157 − 160. [17] KUMAR S, STECHER G, TAMURA K. MEGA7: Molecular evolutionary genetics analysis Version 7.0 for bigger datasets [J]. Molecular Biology and Evolution, 2016, 33(7): 1870 − 1874. [18] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194 − 1202. [19] 祁宏英, 姚美玲, 徐洪国. 龙葵花芽分化形态解剖学研究[J]. 北方园艺, 2017(9): 135 − 138. QI Hongying, YAO Meiling, XU Hongguo. Anatomical and morphological characteristics of development of flower bud differentiation in Solanum nigrum L. [J]. Northern Horticulture, 2017(9): 135 − 138. [20] SOUER E, REBOCHO A B, BLIEK M, et al. Patterning of inflorescences and flowers by the F-box protein DOUBLE TOP and the LEAFY homolog ABERRANT LEAF AND FLOWER of Petunia [J]. Plant Cell, 2008, 20(8): 2033 − 2048. [21] NI Weimin, XIE Daoxin, HOBBIE L, et al. Regulation of flower development in Arabidopsis by SCF complexes [J]. Plant Physiology, 2004, 134(4): 1574 − 1585. [22] CHEN Yue, WEN Haifan, PAN Jian, et al. CsUFO is involved in the formation of flowers and tendrils in cucumber [J]. Theoretical and Applied Genetics, 2021, 134(7): 2141 − 2150. [23] SIMON R, CARPENTER R, DOYLE S, et al. Fimbriata controls flower development by mediating between meristem and organ identity genes [J]. Cell, 1994, 78(1): 99 − 107. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220217
 点击查看大图
点击查看大图

计量
- 文章访问数: 2179
- HTML全文浏览量: 622
- PDF下载量: 93
- 被引次数: 0

