




-
全球气候持续变暖会导致森林产生不同程度的响应。基于2050年气候变化情景,THOMAS等[1]研究发现:中等气候变化背景下,覆盖地球表面20%样本区域将有15%~37%的物种面临灭绝。物种分布模型(SDM)常用来模拟物种潜在分布及适应生境变化[2]。目前,主要的物种分布模型有生物气候模型(Bioclim)、药用植物全球产地生态适宜性区划信息系统(GMPGIS)、气候动态分析模型(Climex)、规则集预测的遗传算法模型(GARP)和最大熵模型(MaxEnt)[3−5],其中最大熵模型较其他同类模型具有预测结果准确性高、灵活性强等优点[6]。因此,最大熵模型在一些植物的潜在分布区研究中有广泛应用,如银杉Cathaya argyrophylla[7]、祁连圆柏Juniperus przewalskii[8]、沙冬青Ammopiptanthus mongolicus[9]等。
樟子松Pinus sylvestris var. mongolica作为欧洲赤松P. sylvestris的一个地理变种,属松科Pinaceae松属Pinus,因其较强的耐寒、耐旱、耐贫瘠土壤和较速生等优良特性被广泛用于三北工程等人工林体系的建设[10−11]。早在1955年,樟子松作为防风固沙林首次成功引种到章古台。迄今为止,樟子松已先后在中国三北工程区的13个省(市、自治区)590余县引种成功。然而,年平均气温达约7 ℃时,樟子松人工林无法自然更新;因章古台、毛乌素沙地和塞罕坝等地受降水限制[12−17],樟子松出现生产力水平降低和枯死等现象。根据《三北工程六期规划(2021—2035年)》,樟子松仍是三北工程中最重要的常绿针叶造林树种。为防止樟子松引种后出现严重衰退或更新困难等情况,本研究利用优化后MaxEnt模型预测不同气候环境下樟子松潜在分布,以期开展未来樟子松引种和优良种质资源培育工作。
-
2018—2021年内蒙古自治区林业和草原监测规划院通过实地调查,在内蒙古自治区范围内樟子松分布区域共获取130个分布数据;结合中国数字植物标本馆(CVH)、中国国家标本植物平台(NSII)、全球生物多样性信息网(GBIF)及其他相关文献共获取樟子松分布点数据115个,经检索汇总后共保留245条记录。将文献记载及实地调研数据整合,配合ArcGIS软件对样点经纬度进行筛选剔除。为避免樟子松群体分布得不均匀,以经纬度2.5′×2.5′为1个网格单元,每个单元筛选1条信息记录,截至2021年最终保留200个样点进行后续拟合分析。
-
本研究选取空间分辨率均为30″的19个生物气候因子和1个地形因子作为环境因子(表1)。气候数据来源于世界气候数据库(
https://www.worldclim.org ),地图数据来源于国家基础地理信息中心的中国标准地图。表 1 全球气候数据库的环境因子
Table 1. Environmental factors in the Global Climate Database
代号 内容 单位 代号 内容 单位 Bio1 年平均气温 ℃ Bio11 最冷季度平均气温 ℃ Bio2 月平均昼夜温差 ℃ Bio12 年降水量 mm Bio3 等温性 Bio13 最湿月降水量 mm Bio4 气温季节性变化 Bio14 最干月降水量 mm Bio5 最热月最高气温 ℃ Bio15 降水量季节性变化 Bio6 最冷月最低气温 ℃ Bio16 最湿季度降水量 mm Bio7 气温年较差 ℃ Bio17 最干季度降水量 mm Bio8 最湿季度平均气温 ℃ Bio18 最暖季度降水量 mm Bio9 最干季度平均气温 ℃ Bio19 最冷季度降水量 mm Bio10 最暖季度平均气温 ℃ Elev 高程 m 未来气候数据选取第6次国际耦合模式计划(CMIP6)的国家气候中心中等分辨率气候系统模式(BCC-CSM2-MR)。此模式为中国未来的气候模式。为更均衡合理模拟未来时期(2050s和2100s)樟子松潜在分布,同时考虑社会经济共享路径(SSP)和代表性浓度路径(RCP),选择SSP-RCP2.6 (SSP126)、SSP-RCP4.5 (SSP245)和SSP-RCP8.5 (SSP585)等3个情景对樟子松未来分布区进行预测。SSP126情景使用可持续发展路径,代表温室气体排放量处于较低水平;SSP245情景使用中度发展路径,代表温室气体排放量处于中等水平;SSP585情景使用以化石燃料为主的发展路径,代表温室气体排放处于高水平[18]。
为避免20个生物气候变量间的相似性过高导致模型过度拟合,通过方差膨胀因子(VIF)和Pearson相关性检验,结合MaxEnt模型预实验中各个因子贡献率,选择相关系数小于0.8、VIF小于10且对樟子松分布具有较高贡献率和生态学意义的因子。最终保留8个气候因子和1个地形因子进行建模,包括年平均气温、最热月最高气温、最冷月最低气温、年降水量、最冷季度平均气温、气温季节变动系数、最干季度平均气温、降水量季节性变化和海拔。
-
本研究利用MaxEnt模型预测当前和未来时期樟子松在中国的潜在分布。将樟子松200个分布点和9个环境因子导入MaxEnt 3.4.1软件,java环境下运行,随机选取25%的样本作为测试数据集,75%的样本作为训练数据集[19],以Bootstrap模式,重复迭代10次,建模结果以Logistic形式输出。
MaxEnt模型准确度可通过受试者工作特征曲线(ROC)进行检验。用该曲线下的面积(AUC)表示所建模型的精度,AUC通常为0.5~1.0,其值越大,表明模型预测越精准。其中AUC在0.5~0.6时表明模型预测失败,0.6~0.7时表明模型预测效果较差,0.7~0.8时表明模型预测效果一般,0.8~0.9时表明模型预测效果良好,0.9~1.0时表明预测效果极好[20],可以较准确地反映物种潜在分布。
-
本模型采用优化方法为Checkerboard 2法,调整调控倍率(RM)和特征组合(FC) 2个参数改变模型正规化水平。通过改变特征数量、组合方式以及RM数值,将MaxEnt模型中的5种特征:线性特征(L)、二次型特征(Q)、片段化特征(H)、乘积型特征(P)和阈值型特征(T),与16个RM值(0.5~8.0,以0.5为间隔梯度)排列组合出148种组合方式。利用R语言中的ENMeval程序包对其进行优化测试,结果中各组合的ΔAICc值和10%测试遗漏率越低,表示模型预测精准度越高[21−22]。
-
运用ArcGIS将模型运行后的数据进行适宜区划分和可视化处理,根据软件中提供的多种分类方法预测樟子松适宜性阈值。根据冉巧等[7]对孑遗植物银杉进行适宜区划分的方法,本研究使用自然断点法划分樟子松适宜指数。参照张伟萍等[8]对祁连圆柏适宜区划分的等级,将樟子松生境划分为不适宜区、较不适宜区、一般适宜区和高度适宜区(适宜指数分别为0~0.1、>0.1~0.2、>0.2~0.6、>0.6~1.0)。对比樟子松不同时期的适宜区分布差异,获得樟子松不同适宜区空间分布图;利用ArcGIS 10.4.1软件中SDMtoolbox 2.0工具包,计算樟子松在不同时期的适生区质心位置得到樟子松分布区的迁移方向,并计算其质心迁移距离。
-
根据200个樟子松分布点和9个环境因子利用MaxEnt模型对樟子松的潜在分布区域进行模拟预测。在MaxEnt为默认参数设置时,RM为1.0,FC为LQHPT,∆AICc为29.77。当RM为1.5, FC为LQHPT时,∆AICc为0,此时模型最优,10%的训练遗漏率降低4.63% (表2)。因此选取RM为1.5, FC为LQHPT为模型最终参数。该参数下10次模拟训练AUC均值为0.964 (图1),说明预测结果精确。
表 2 不同参数MaxEnt数模型优化结果
Table 2. Environmental results of MaxEnt model under different parameter settings
模型评价 特征组合 调控倍频 ∆AICc 10%训练遗漏率 默认 LQHPT 1.0 29.77 0.169 75 最优 LQHPT 1.5 0 0.161 89 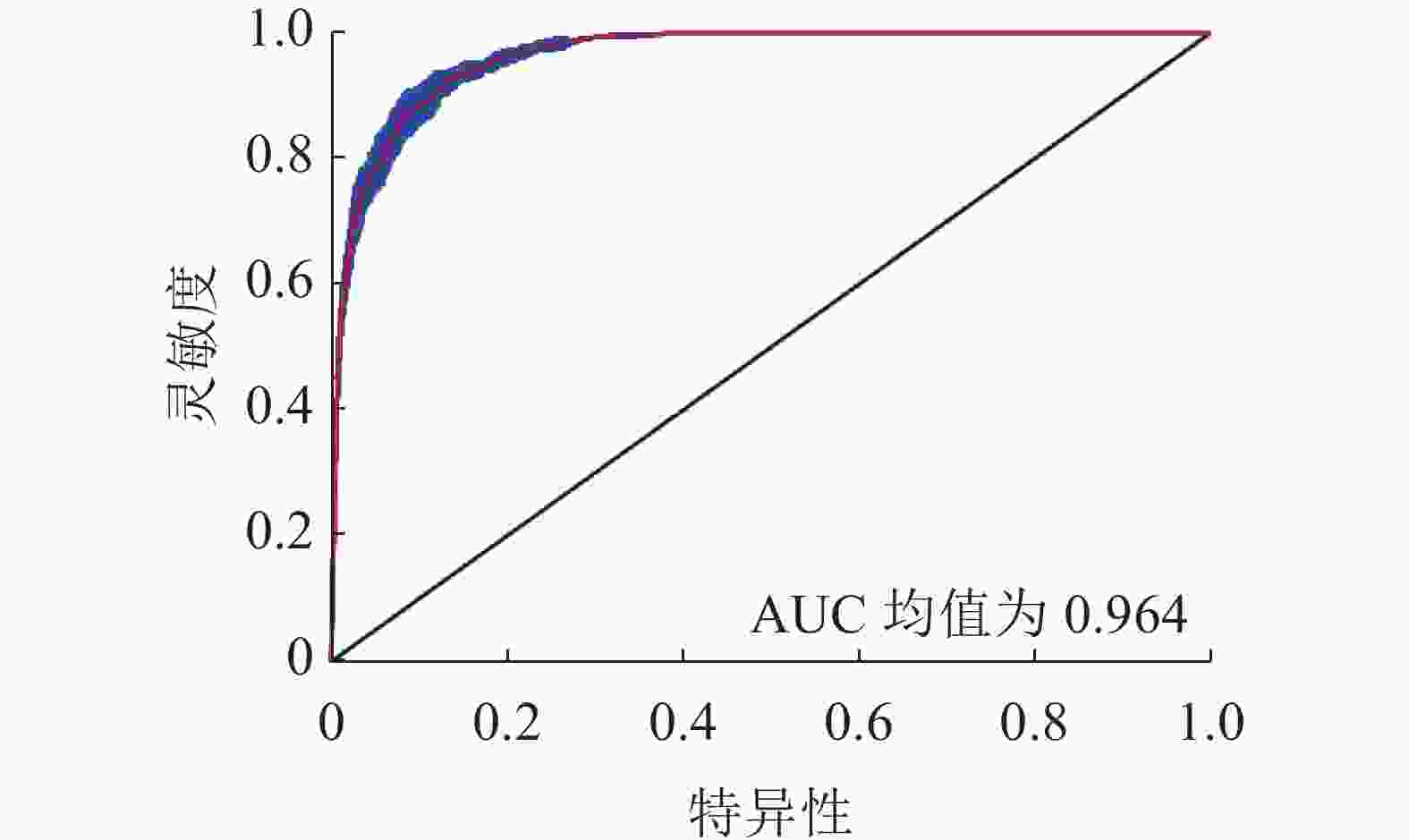
图 1 MaxEnt模型的ROC响应曲线
Figure 1. ROC response curve under MaxEnt model
-
樟子松当前气候条件下的潜在适生区集中在53.55°N以南,135.09°E以西,总适生区面积(一般适生区面积与高度适生区面积之和)为645 104.2 km2,主要位于大兴安岭地区、内蒙古中部、黑龙江西北部,在山西、河北、吉林、辽宁也有少量分布[23],其中,高度适生区面积为147 638.9 km2,一般适生区面积为497 465.3 km2。樟子松高度适应区绝大部分在内蒙古东北部,黑龙江西北部呈零星分布。
-
未来时期不同气候情景下,樟子松适生区面积较当前时期有不同程度减少。2050s,樟子松总适生区在SSP126、SSP245和SSP585气候情景下降35.7%、52.5%和82.6%;2100s与2050s相比,总适生区面积在SSP126、SSP245和SSP585情景下呈下降趋势,降幅分别为62.1%、37.4%和100.0%。SSP126-2050s情景下,樟子松高度适生区面积相较当前大幅下降,面积约为当前的6.6%,仅存在大兴安岭较高海拔地区;SSP245-2100s气候情景下,樟子松高度适生区将完全消失。
在3种气候条件下樟子松适生区面积均下降,SSP245和SSP585气候情景下樟子松适生区对气候的响应最为明显 (表3和表4)。SSP126气候情景下,2050s樟子松适生区面积变化幅度最小,适生区面积下降230 225.7 km2,降幅为35.7%,主要变化区域位于呼伦贝尔市西南部;2100s适生区面积下降487 968.8 km2,降幅为75.6%,主要在兴安盟和锡林郭勒盟的东北部出现较大面积消失。SSP245气候情景下,2050s樟子松适生区面积下降338 449.4 km2,降幅为52.5%,其变化区域主要位于内蒙古自治区呼伦贝尔市西东两侧地区和赤峰市;2100s其适生区面积下降453 211.8 km2,降幅为70.3%,内蒙古自治区东部和黑龙江省的大部分地区樟子松适生区消失。SSP585气候情景下,樟子松适生区面积变化幅度最大,2050s樟子松适生区面积下降532 882.0 km2,比当前减少82.6%,仅在呼伦贝尔市中部有樟子松分布;2100s樟子松适生区将在中国完全消失。
表 3 中国不同气候情境下樟子松适生区面积
Table 3. Suitable growing area of P. sylvestris var. mongolica under different climates scenarios in China
气候变化情景 高度适生区/km2 一般适生区/km2 较不适生区/km2 不适生区/km2 总适生区/km2 当前 147 638.9 497 465.3 896 614.6 8 058 281.8 645 104.2 SSP126-2050s 9 809.0 405 069.4 618 090.3 8 567 031.4 414 878.5 SSP126-2100s 4 219.8 152 916.7 571 493.1 8 871 371.6 157 135.4 SSP245-2050s 4 433.2 302 221.6 848 215.5 8 445 129.9 306 654.8 SSP245-2100s 0 191 892.4 802 395.8 8 605 711.9 191 892.4 SSP585-2050s 0 112 222.2 437 517.4 9 050 260.5 112 222.2 SSP585-2100s 0 0 108 142.4 9 491 857.8 0 表 4 不同时期樟子松适生区空间格局变化
Table 4. Dynamic changes in the suitable area for P. sylvestris var. mongolica under different combination of climates cenarios
气候变化情景 面积/km2 面积变化率/% 丧失区 增加区 保留区 丧失率 增加率 保留率 SSP126-2050s 300 586.5 782 87.1 337 725.7 46.59 12.13 52.35 SSP126-2100s 277 630.1 173 36.2 138 584.8 41.12 2.56 20.53 SSP245-2050s 378 636.3 446 69.6 258 933.5 57.79 6.81 39.53 SSP245-2100s 185 735.3 706 42.3 118 506.6 27.11 10.31 17.30 SSP585-2050s 549 974.5 200 00.8 889 97.1 82.69 3.00 13.38 SSP585-2100s 111 059.9 0 0 15.97 0 0 -
在空间分布格局方面,未来时期不同气候情景下樟子松适生区的迁移位置存在差异,但总体上呈现出向西北和西南方向迁移(表5)。当前时期樟子松适生区的质心在内蒙古兴安盟阿尔山市(47.09°N,120.25°E)。气候情景为SSP126-2100s时,樟子松适生区向西北方向迁移,此时樟子松适生区质心位于内蒙古自治区呼伦贝尔市鄂温克族自治旗(48.41°N,120.17°E),迁移距离为161.47 km;气候情景为SSP585-2050s时,樟子松向西北方向迁移,此时其适生区质心位于内蒙古自治区呼伦贝尔市新巴尔虎左旗(47.88°N,119.06°E),迁移距离为126.36 km;气候情景为SSP245-2100s时,樟子松向西南方向迁移,此时其适生区质心位于内蒙古自治区锡林郭勒盟东乌珠穆沁旗(45.66°N,117.39°E),迁移距离为270.80 km。未来时期不同气候变化情景下,全球增温增湿使得樟子松在中国适生区的质心整体向西北和西南迁移,质心随着适生区分布范围消减可能消失。
表 5 不同气候情景下樟子松适生区质心变化
Table 5. Core distributional shifts under different climate scenario/periods for P. sylvestris var. mongolica
时期 气候情景 地理坐标 迁移距离/km 北纬(N) 东经(E) 当前 47.09° 120.25° 2050s SSP126 47.11° 120.04° 16.04 SSP245 46.65° 119.29° 87.86 SSP585 47.88° 119.06° 126.36 2100s SSP126 48.41° 120.17° 161.47 SSP245 45.66° 117.39° 270.80 说明:SSP585气候情景下在2100s时期无樟子松适生区。 -
根据Jackknife法分析结果(表6),樟子松潜在地理分布贡献率超过5%的环境因子分别为最冷季度平均气温(22.1%)、降水量季节性变化(19.8%)、最冷月最低气温(18.1%)、气温季节变动系数(17.8%)、最干季度平均气温(9.0%)、最热月最高气温(6.1%)。这6个环境因子总贡献率达92.9%。
表 6 参与建模的环境因子贡献率及置换重要值
Table 6. Percentage contribution and permutation importance of environment variables for P. sylvestris var. mongolica in the MaxEnt model
因子 描述 贡献率/% 置换重要值 因子 描述 贡献率/% 置换重要值 Bio11 最冷季度平均气温 22.1 2.7 Bio5 最热月最高气温 6.1 6.0 Bio15 降水量季节性变化 19.8 12.4 Bio12 年降水量 4.9 9.0 Bio6 最冷月最低气温 18.1 4.3 Elev 海拔 1.7 48.2 Bio4 气温季节变动系数 17.8 3.5 Bio1 年平均气温 0.5 9.4 Bio9 最干季度平均气温 9.0 4.3 -
樟子松分布主要环境因子与潜在分布区的关系如表7所示。SSP126气候情景下,2050s和2100s樟子松生境适宜度比当前降低了0.38和0.54;SSP245气候情景下,2050s和2100s樟子松适宜度比当前降低了0.46和0.55;SSP585气候情景下,樟子松生境适宜度急剧下降,2050s下降了0.55,到了2100s下降0.67,降幅为91%。
表 7 樟子松适生区环境因子结果分析
Table 7. Results analysis of major environmental variables in P. sylvestris var. mongolica suitable area
时期 气候
情景年平均
气温/℃气温季节
变动系数最热月最高
气温/℃最冷月最低
气温/℃最干季度平均
气温/℃最冷季度平均
气温/℃年降水
量/mm降水量季节性
变化海拔/
m物种生境
适宜度当前 1.60 1 524.06 24.35 −29.18 −18.79 −20.64 425.20 112.41 828.22 0.74 2050s SSP126 1.90 1 483.54 26.94 −25.97 −16.44 −17.75 430.87 113.05 828.22 0.36 SSP245 2.49 1 511.33 27.97 −25.37 −16.05 −17.45 445.52 110.16 828.22 0.28 SSP585 3.20 1 517.97 28.53 −25.13 −15.46 −17.14 469.78 114.43 828.22 0.19 2100s SSP126 2.20 1 479.34 26.75 −26.44 −16.12 −17.77 460.45 116.35 828.22 0.20 SSP245 3.68 1 519.68 28.91 −23.84 −14.88 −16.47 463.79 115.97 828.22 0.19 SSP585 6.11 1 494.04 30.98 −21.86 −12.59 −13.42 515.40 121.54 828.22 0.07 200个樟子松分布点的年平均气温与其生境适宜度变化正好相反,2050s,SSP126、SSP245和SSP585情景下年平均气温分别增加19.5%、56.6%和101.3%;2100s与2050s相比,SSP126、SSP245和SSP585情景下年平均气温分别增加15.9%、48.1%和90.7%。
200个樟子松分布点的年降水量在SSP126、SSP245和SSP585情景下均为递增趋势,且年降水量与年平均气温变化趋势相同,都与樟子松物种适宜度变化相反。2050s,SSP126、SSP245和SSP585情景下年降水量分别增加5.67、20.32和44.58 mm;2100s与2050s相比,SSP126、SSP245和SSP585情景下年降水量分别增加29.58、18.27和45.62 mm。
-
本研究基于气候和地形因子的研究,应用ENMeval数据包中的Checkerboard 2法来优化模型[23]。作为北方主要造林树种,樟子松集中分布在中国北方地区,具有喜光、耐寒、耐旱、耐贫瘠和抗病能力强的生长特性。在中国的大兴安岭北部,西起莫尔道戈、金河、根河,东到新林、呼玛以北有连续的分布,在南伊图里河的银河、免渡河、阿尔山等地呈带状或块状分布着樟子松纯林以及其与兴安落叶松Larix gmelinii构成的混交林[24]。自1955年樟子松被成功引种到辽宁省阜新市彰武县章古台以来,已推广到科尔沁沙地、浑善达克沙地、毛乌素沙地等地。本研究基于优化的最大熵模型预测结果显示:当前樟子松高度适生区也位于上述地区,预测结果与实际高度一致,表明经过优化后的MaxEnt模型用于樟子松适生区的预测结果精准可靠。
-
Jackknife法分析表明:年平均气温、最热月最高气温、最冷月最低气温、年降水量、最冷季度平均气温、气温季节变动系数、最干季度平均气温、降水量季节性变化和海拔是影响樟子松分布的主要环境因子。当樟子松存在概率大于0.6时,对应的环境因子最适合植物生长。主导环境因子分析表明:气温和降水相关因子对樟子松分布贡献率达到98.3%,对其分布起决定性作用。这与赵晓彬[25]研究中樟子松适生于年平均气温−3.7~−1.5 ℃、年降水量375 mm的结论相符。本研究预测樟子松的最适宜海拔为800 m左右,与戴继先等[26]研究发现樟子松天然分布在300~900 m的阳坡或向阳坡的结论一致。水热条件一定程度上决定了樟子松潜在地理分布,地形在一定程度上也会对水热资源进行二次分配,因此,气温、降水和地形对樟子松分布格局的相互作用都不能忽视。
-
2050s和2100s不同二氧化碳排放情景下,高温高湿使樟子松适生区不断减少。2100s樟子松适生区显著减少,其降幅明显大于2050s。未来不同的气候情景下,樟子松适生区的空间分布变化随着气候变暖加剧,整体向西北高纬度和西南季节性降水量增加的地区收缩。SSP126情景下,2050s适生区质心迁移距离最短,说明气候小幅变暖对适生区分布范围影响不大。SSP245情景下樟子松适生区质心点受季节性降水量影响整体向西南方向降水量充沛地区迁移,2100s与2050s相比其质心点迁移距离较大。SSP585情景下温室气体排放增加,2050s樟子松适生区质心点明显向西北高纬度地区迁移,2100s适生区在中国消失。张日升等[27]研究表明:随着温室气体排放等环境问题加剧,年平均气温达7 ℃左右时樟子松将出现生长不良,导致大面积退化。吴祥云等[28]也曾提出:樟子松人工林引种到年平均气温为7.1 ℃的章古台会发生“早衰”现象,导致面积减退。未来气候情景下,温室气体排放,气温升高导致樟子松适生区面积逐步北移,在中国分布减少甚至至消失。综上所述,樟子松迁移趋势与CHEN[29]研究中国寒温带森林树种地理分布在未来全球变暖下往高纬度和降水量充沛地区迁移的趋势相符。
-
本研究通过对MaxEnt模型的优化,发现樟子松潜在分布最优模型的特征组合为片段化特征(LQHPT)、乘积型特征(P)和阈值性特征(H),调控倍率为1.5。此模型复杂度低,预测结果较准确,预测樟子松适生区范围与实际分布情况基本一致。中国樟子松当前高度适生区集中在47.2°~53.3°N, 118.8°~121.9°E的大兴安岭北部高纬度地区。不同气候情景下,樟子松对水热变化最为敏感,特别是在年平均气温升高5.0 ℃左右时将导致其在中国的分布区发生显著变化。
通过樟子松不同适生区的划分,根据各自区划气候特点提出以下保护对策:①在不同地区选取生长良好的樟子松单株进行种质资源监测、保护、采集和培育,培育最适合当地气候条件的优良品种。②未来在不适合樟子松生长地区引种时,应避免引种樟子松。③提前进行系统的种源区划研究,培育出不同特性的樟子松并在本地区推广。应该在年平均气温为−3.7~1.6 ℃,降水量为375~425 mm的东北等高纬度地区引种樟子松,以发挥其最大生态效益。
Prediction of the potential distribution pattern of Pinus sylvestris var. mongolica in China under climate change
-
摘要:
目的 预测不同气候条件下樟子松Pinus sylvestris var. mongolica在中国的潜在分布及迁移,确定影响樟子松分布的主要环境变量,为樟子松合理引种与保护提供理论依据。 方法 根据200个樟子松分布点和20个环境变量,利用R语言中ENMeval数据包优化最大熵模型(MaxEnt)并利用ArcGIS空间分析技术对当前气候条件下樟子松在中国潜在分布进行模拟,通过Pearson相关分析和方差膨胀因子分析结合预建模结果对环境因子的筛选,综合Jackknife检验和相关系数,分析樟子松主导限制因子,预测樟子松从当前到未来时期(2050s和2100s)的3种气候情景(SSP126、SSP245和SSP585)下适生区变化趋势。 结果 MaxEnt模型受训者工作特征曲线都大于0.94,说明模型精度较高,能较好预测樟子松潜在分布;影响樟子松分布的主导因子为最冷季度平均气温、降水量季节性变化、最冷月最低气温、温度季节变动系数、最干季度平均气温和最热月最高气温,累计贡献率为92.9%;当前气候条件下,樟子松的适宜分布区主要位于中国大兴安岭地区,总适宜区面积占中国总面积的6.72%;未来时期不同气候条件下樟子松潜在分布区面积减少,质心向西北高纬度和西南降水量充沛地区迁移。 结论 以年为单位的温度和降水是樟子松分布的主要影响因子,当前樟子松的适生区主要在中国大兴安岭地区,未来樟子松分布区有向现有分布区的西北和西南地区迁移的趋势。图1表7参29 Abstract:Objective This study aims to predict the potential distribution and migration of Pinus sylvestris var. mongolica under different climate conditions in China and to determine the main environmental variables affecting its distribution, so as to provide theoretical basis for rational introduction and protection of P. sylvestris var. mongolica. Method Based on 200 distribution points and 20 environmental variables, the potential distribution of P. sylvestris var. mongolica under current climate conditions was simulated by using ENMeval packet optimization maximum entropy model (MaxEnt) in R language and ArcGIS spatial analysis technology. Through Pearson correlation analysis and variance inflation factor analysis combined with the screening of environmental factors based on pre-modeling results, Jackknife test and correlation coefficient were integrated to analyze the dominant limiting factors of P. sylvestris var. mongolica, and predict the change trend of suitable habitat under three climate scenarios (SSP126, SSP245 and SSP585) from the current to the future (2050s and 2100s). Result The area under ROC curve of the MaxEnt model was greater than 0.94, indicating that the model had high accuracy and could better predict the potential distribution of P. sylvestris var. mongolica. The main factors affecting the distribution were the average temperature in the coldest quarter, seasonal variation of precipitation, minimum temperature in the coldest month, seasonal variation coefficient of temperature, average temperature in the driest quarter and maximum temperature in the hottest month, with a cumulative contribution rate of 92.9%. Under the current climate conditions, the suitable distribution area of P. sylvestris var. mongolica was mainly located in the Greater Hinggan Mountains of China, and the total suitable area accounted for 6.72% of the total area of China. In the future, the potential distribution area of P. sylvestris var. mongolica would decrease under different climatic conditions, and the centroid would migrate to the northwest area at high latitude and southwest area with abundant precipitation. Conclusion The annual temperature and precipitation are the main factors affecting the distribution of P. sylvestris var. mongolica. At present, the suitable growing areas are mainly concentrated in the Greater Hinggan Mountains of China, and its distribution will migrate to the northwest and southwest of the existing distribution area in the future. [Ch, 1 fig. 7 tab. 29 ref.] -
表 1 全球气候数据库的环境因子
Table 1. Environmental factors in the Global Climate Database
代号 内容 单位 代号 内容 单位 Bio1 年平均气温 ℃ Bio11 最冷季度平均气温 ℃ Bio2 月平均昼夜温差 ℃ Bio12 年降水量 mm Bio3 等温性 Bio13 最湿月降水量 mm Bio4 气温季节性变化 Bio14 最干月降水量 mm Bio5 最热月最高气温 ℃ Bio15 降水量季节性变化 Bio6 最冷月最低气温 ℃ Bio16 最湿季度降水量 mm Bio7 气温年较差 ℃ Bio17 最干季度降水量 mm Bio8 最湿季度平均气温 ℃ Bio18 最暖季度降水量 mm Bio9 最干季度平均气温 ℃ Bio19 最冷季度降水量 mm Bio10 最暖季度平均气温 ℃ Elev 高程 m  下载: 导出CSV
下载: 导出CSV
表 2 不同参数MaxEnt数模型优化结果
Table 2. Environmental results of MaxEnt model under different parameter settings
模型评价 特征组合 调控倍频 ∆AICc 10%训练遗漏率 默认 LQHPT 1.0 29.77 0.169 75 最优 LQHPT 1.5 0 0.161 89
下载: 导出CSV
表 3 中国不同气候情境下樟子松适生区面积
Table 3. Suitable growing area of P. sylvestris var. mongolica under different climates scenarios in China
气候变化情景 高度适生区/km2 一般适生区/km2 较不适生区/km2 不适生区/km2 总适生区/km2 当前 147 638.9 497 465.3 896 614.6 8 058 281.8 645 104.2 SSP126-2050s 9 809.0 405 069.4 618 090.3 8 567 031.4 414 878.5 SSP126-2100s 4 219.8 152 916.7 571 493.1 8 871 371.6 157 135.4 SSP245-2050s 4 433.2 302 221.6 848 215.5 8 445 129.9 306 654.8 SSP245-2100s 0 191 892.4 802 395.8 8 605 711.9 191 892.4 SSP585-2050s 0 112 222.2 437 517.4 9 050 260.5 112 222.2 SSP585-2100s 0 0 108 142.4 9 491 857.8 0
下载: 导出CSV
表 4 不同时期樟子松适生区空间格局变化
Table 4. Dynamic changes in the suitable area for P. sylvestris var. mongolica under different combination of climates cenarios
气候变化情景 面积/km2 面积变化率/% 丧失区 增加区 保留区 丧失率 增加率 保留率 SSP126-2050s 300 586.5 782 87.1 337 725.7 46.59 12.13 52.35 SSP126-2100s 277 630.1 173 36.2 138 584.8 41.12 2.56 20.53 SSP245-2050s 378 636.3 446 69.6 258 933.5 57.79 6.81 39.53 SSP245-2100s 185 735.3 706 42.3 118 506.6 27.11 10.31 17.30 SSP585-2050s 549 974.5 200 00.8 889 97.1 82.69 3.00 13.38 SSP585-2100s 111 059.9 0 0 15.97 0 0
下载: 导出CSV
表 5 不同气候情景下樟子松适生区质心变化
Table 5. Core distributional shifts under different climate scenario/periods for P. sylvestris var. mongolica
时期 气候情景 地理坐标 迁移距离/km 北纬(N) 东经(E) 当前 47.09° 120.25° 2050s SSP126 47.11° 120.04° 16.04 SSP245 46.65° 119.29° 87.86 SSP585 47.88° 119.06° 126.36 2100s SSP126 48.41° 120.17° 161.47 SSP245 45.66° 117.39° 270.80 说明:SSP585气候情景下在2100s时期无樟子松适生区。
下载: 导出CSV
表 6 参与建模的环境因子贡献率及置换重要值
Table 6. Percentage contribution and permutation importance of environment variables for P. sylvestris var. mongolica in the MaxEnt model
因子 描述 贡献率/% 置换重要值 因子 描述 贡献率/% 置换重要值 Bio11 最冷季度平均气温 22.1 2.7 Bio5 最热月最高气温 6.1 6.0 Bio15 降水量季节性变化 19.8 12.4 Bio12 年降水量 4.9 9.0 Bio6 最冷月最低气温 18.1 4.3 Elev 海拔 1.7 48.2 Bio4 气温季节变动系数 17.8 3.5 Bio1 年平均气温 0.5 9.4 Bio9 最干季度平均气温 9.0 4.3
下载: 导出CSV
表 7 樟子松适生区环境因子结果分析
Table 7. Results analysis of major environmental variables in P. sylvestris var. mongolica suitable area
时期 气候
情景年平均
气温/℃气温季节
变动系数最热月最高
气温/℃最冷月最低
气温/℃最干季度平均
气温/℃最冷季度平均
气温/℃年降水
量/mm降水量季节性
变化海拔/
m物种生境
适宜度当前 1.60 1 524.06 24.35 −29.18 −18.79 −20.64 425.20 112.41 828.22 0.74 2050s SSP126 1.90 1 483.54 26.94 −25.97 −16.44 −17.75 430.87 113.05 828.22 0.36 SSP245 2.49 1 511.33 27.97 −25.37 −16.05 −17.45 445.52 110.16 828.22 0.28 SSP585 3.20 1 517.97 28.53 −25.13 −15.46 −17.14 469.78 114.43 828.22 0.19 2100s SSP126 2.20 1 479.34 26.75 −26.44 −16.12 −17.77 460.45 116.35 828.22 0.20 SSP245 3.68 1 519.68 28.91 −23.84 −14.88 −16.47 463.79 115.97 828.22 0.19 SSP585 6.11 1 494.04 30.98 −21.86 −12.59 −13.42 515.40 121.54 828.22 0.07
下载: 导出CSV
-
[1] THOMAS C D, CAMERON A, GREEN R E, et al. Extinction risk from climate change [J]. Nature, 2004, 427(6970): 145 − 148. [2] BEAUMONT L J, HUGHES L, POULSEN M. Predicting species distributions: use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions [J]. Ecological Modelling, 2005, 186(2): 251 − 270. [3] PATIISON R R, MACK R N. Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: evaluating CLIMEX predictions with field trials [J]. Global Change Biology, 2008, 14(4): 813 − 826. [4] 邓阳川, 向丽, 汤欢, 等. 基于GMPGIS的杜仲全球产地生态适宜性分析[J]. 世界科学技术—中医药现代化, 2019, 21(4): 755 − 763. DENG Yangchuan, XIANG Li, TANG Huan, et al. Suitability analysis of Eucommia ulmoides global ecological adaptability area based on GMPGIS [J]. Modernization of Traditional Chinese Medicine and Materia Medica-World Science and Technology, 2019, 21(4): 755 − 763. [5] 叶芸, 孔德英, 王振华, 等. 基于CLIMEX的西方散白蚁在中国潜在适生区分析[J]. 湖北农业科学, 2016, 55(15): 3894 − 3896. YE Yun, KONG Deying, WANG Zhenhua, et al. The potential geographical distribution of Reticulitermes hesperus in China based on CLIMEX [J]. Hubei Agricultural Science, 2016, 55(15): 3894 − 3896. [6] 杨芙蓉, 张琴, 孙成忠, 等. 蒙古黄芪潜在分布区预测的多模型比较[J]. 植物科学学报, 2019, 37(2): 136 − 143. YANG Furong, ZHANG Qin, SUN Chengzhong, et al. Comparative evaluation of multiple models for predicting the potential distribution areas of Astragalus membranaceus var. mongholicus [J]. Journal of Plant Science, 2019, 37(2): 136 − 143. [7] 冉巧, 卫海燕, 赵泽芳, 等. 气候变化对孑遗植物银杉的潜在分布及生境破碎度的影响[J]. 生态学报, 2019, 39(7): 2481 − 2493. RAN Qiao, WEI Haiyan, ZHAO Zefang, et al. Impact of climate change on the potential distribution and habitat fragmentation of the relict plant Cathaya argyrophylla Chun et Kuang [J]. Acta Ecologica Sinica, 2019, 39(7): 2481 − 2493. [8] 张伟萍, 胡云云, 李智华, 等. 气候变化情景下祁连圆柏在青海省的适宜分布区预测[J]. 应用生态学报, 2021, 32(7): 2514 − 2524. ZHANG Weiping, HU Yunyun, LI Zhihua, et al. Predicting suitable distribution areas of Juniperus przewalskii in Qinghai Province under climate change scenarios [J]. Chinese Journal of Applied Ecology, 2021, 32(7): 2514 − 2524. [9] 牛若恺, 高润红, 侯艳青, 等. 气候变化下沙冬青适宜分布区预测[J]. 西北林学院学报, 2021, 36(1): 102 − 107. NIU Ruokai, GAO Runhong, HOU Yanqing, et al. Prediction of the geographic distribution of Ammopiptanthus mongolicus under climate change [J]. Journal of Northwest College of Forestry, 2021, 36(1): 102 − 107. [10] 焦树仁. 辽宁省章古台樟子松固沙林提早衰弱的原因与防治措施[J]. 林业科学, 2001, 37(2): 131 − 138. JIAO Shuren. Reasons and prevention measures for early weakness of forest in Zhanggutai, Liaoning Province [J]. Scientia Silvae Sinicae, 2001, 37(2): 131 − 138. [11] ZHU Jiaojun, FAN Zhiping, ZENG Dehui, et al. Comparison of stand structure and growth between artificial and natural forests of Pinus sylvestiris var. mongolica on sandy land [J]. Journal of Forestry Research, 2003, 14(2): 103 − 111. [12] 刘亚玲, 信忠保, 李宗善, 等. 河北坝上樟子松人工林径向生长及其对气候因素的响应[J]. 生态学报, 2022, 42(5): 1830 − 1840. LIU Yaling, XIN Zhongbao, LI Zongshan, et al. Response of radial growth of Pinus sylvestris var. mongolica to climate factors in Bashang area of Hebei Province [J]. Acta Ecologica Sinica, 2022, 42(5): 1830 − 1840. [13] 王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系[J]. 植物生态学报, 2011, 35(3): 294 − 302. WANG Xiaochun, SONG Laiping, ZHANG Yuandong. Climate-tree growth relationships of Pinus sylvestris var. mongolica in the northern Daxing’ an Mountains, China [J]. Chinese Journal of Plant Ecology, 2011, 35(3): 294 − 302. [14] 徐静, 郭滨德, 孙洪志. 帽儿山地区不同种源樟子松树轮对气候因子的响应[J]. 林业科学研究, 2016, 29(4): 581 − 586. XU Jing, GUO Bingde, SUN Hongzhi. Tree ring response of scots pine provenances to climate factors at Maoershan, northeastern China [J]. Forestry Research, 2016, 29(4): 581 − 586. [15] 李俊霞, 白学平, 张先亮, 等. 大兴安岭林区南、北部天然樟子松生长对气候变化的响应差异[J]. 生态学报, 2017, 37(21): 7232 − 7241. LI Junxia, BAI Xueping, ZHANG Xianliang, et al. Different responses of natural Pinus sylvestris var. mongolica growth to climate change in southern and northern forested areas in the Great Xing’ an Mountains [J]. Acta Ecologica Sinica, 2017, 37(21): 7232 − 7241. [16] 尚建勋, 时忠杰, 高吉喜, 等. 呼伦贝尔沙地樟子松年轮生长对气候变化的响应[J]. 生态学报, 2012, 32(4): 1177 − 1184. SHANG Jianxun, SHI Zhongjie, GAO Jixi, et al. Response of tree-ring width of Pinus sylvestris var. mongolica to climate change in Hulunbuir Sand Land, China [J]. Acta Ecologica Sinica, 2012, 32(4): 1177 − 1184. [17] 雷帅, 张劲松, 孟平, 等. 中国北部不同地点樟子松人工林径向生长对气候响应的差异[J]. 生态学报, 2020, 40(13): 4479 − 4492. LEI Shuai, ZHANG Jinsong, MENG Ping, et al. Differences in tree-ring growth response of Pinus sylvestris var. mongolica to climatic variation at different locations in northern China [J]. Acta Ecologica Sinica, 2020, 40(13): 4479 − 4492. [18] LI Suyuan, MIAO Lijuan, JIANG Zihong, et al. Projected drought conditions in northwest China with CMIP6 models under combined SSPs and RCPs for 2015−2099 [J]. Advances in Climate Change Research, 2020, 11: 210 − 217. [19] YI Yujun, CHENG Xi, YANG Zhifeng, et al. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China [J]. Ecological Engineering, 2016, 92: 260 − 269. [20] RONG Zhanlei, ZHAO Chuanyan, LIU Junjie, et al. Modeling the effect of climate change on the potential distribution of Qinghai spruce (Picea crassifolia Kom.) in Qilian Mountains[J/OL]. Forests, 2019, 10(1): 62[2022-06-03]. doi: 10.3390/f10010062. [21] 赵光华, 樊保国. 末次间冰期以来濒危植物藤枣适生区空间迁移预测[J]. 西南农业学报, 2021, 34(1): 174 − 182. ZHAO Guanghua, FAN Baoguo. Prediction on spatial migration of suitable distribution of Eleutharrhena macrocarpa (Diels) forman since last inter glacial [J]. Southwest China Journal of Agricultural Sciences, 2021, 34(1): 174 − 182. [22] 李安, 李良涛, 高萌萌, 等. 基于MaxEnt模型和气候变化情景入侵种黄顶菊在中国的分布区预测[J]. 农学学报, 2020, 10(1): 60 − 67. LI An, LI Liangtao, GAO Mengmeng, et al. Distribution prediction of invasive species Flaveria bidentis in China: based on MaxEnt Model and climate change scenario [J]. Journal of Agriculture, 2020, 10(1): 60 − 67. [23] 刘佳琪, 魏广阔, 史常青, 等. 基于MaxEnt模型的北方抗旱造林树种适宜区分布[J]. 北京林业大学学报, 2022, 44(7): 63 − 77. LIU Jiaqi, WEI Guangkuo, SHI Changqing, et al. Suitable distribution area of drought-resistant afforestation tree species in north China based on MaxEnt model [J]. Journal of Beijing Forestry University, 2022, 44(7): 63 − 77. [24] 赵光华, 崔馨月, 王智, 等. 气候变化背景下我国酸枣潜在适生区预测[J]. 林业科学, 2021, 57(6): 158 − 168. ZHAO Guanghua, CUI Xinyue, WANG Zhi, et al. Prediction of potential distribution of Ziziphus jujuba var. spinosa in China under context of climate change [J]. Scientia Silvae Sinicae, 2021, 57(6): 158 − 168. [25] 赵晓彬. 榆林沙区樟子松造林技术研究[D]. 杨凌: 西北农林科技大学, 2007. ZHAO Xiaobin. Study on Afforestation Techniques of Pinus Sylvestris var. mongolica in Yunlin Sandy Land[D]. Yangling: Northwest A&F University, 2007. [26] 戴继先, 杨国林, 杨战阳. 治沙造林先锋树种——樟子松造林技术研究[J]. 林业实用技术, 2003(10): 5 − 7. DAI Jixian, YANG Guolin, YANG Zhanyang. Sand control forestation pioneer tree species: study on afforestation technology of Pinus sylvestris var.mongolica [J]. Practical Forestry of Technology, 2003(10): 5 − 7. [27] 张日升, 贾树海, 张国剑, 等. 基于GIS的樟子松种植适宜性评价研究[J]. 土壤通报, 2019, 50(3): 555 − 561. ZHANG Risheng, JIA Shuhai, ZHANG Guojian, et al. Suitability evaluation for Pinus sylvestris var. mongolica planting based on GIS [J]. Chinese Journal of Soil Science, 2019, 50(3): 555 − 561. [28] 吴祥云, 姜凤岐, 李晓丹, 等. 樟子松人工固沙林衰退的规律和原因[J]. 应用生态学报, 2004, 15(12): 2225 − 2228. WU Xiangyun, JIANG Fengqi, LI Xiaodan, et al. Decline regularity and causes of Pinus sylvestris var. mongolica plantation on sandy land [J]. Chinese Journal of Applied Ecology, 2004, 15(12): 2225 − 2228. [29] CHEN Xiongwen. Modeling the effects of global climatic change at the ecotone of boreal larch forest and temperate forest in northeast China [J]. Climatic Change, 2012, 55(1/2): 77 − 97. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220451
 点击查看大图
点击查看大图

计量
- 文章访问数: 1799
- HTML全文浏览量: 536
- PDF下载量: 69
- 被引次数: 0
