




-
土壤物理性质反映土壤结构的稳定性,在一定程度上可以表征土壤的质量与退化状况[1]。除地形地貌、土壤特征、降雨差异和植被类型等外[2],人类活动也是影响土壤性质和土壤形成过程的重要因素[3−4]。适度的干扰会对物种多样性和生物量以及种群、群落、生态系统甚至整体景观格局有正向促进作用,而长期的人为干扰会造成土壤压实,降低土壤团聚体稳定性、持水能力和微生物活性等[5]。郭宝宝[6]通过研究不同人为干扰强度对森林土壤水分物理性质的影响发现:土壤容重随人为干扰强度的增加呈明显增加的趋势。庞学勇等[7]通过研究人为干扰对川西亚高山针叶林土壤物理性质的影响发现:随人为干扰强度的增加,土壤粉粒、黏粒含量和大团聚体数量减少,土壤表层孔隙度减小,土壤有效水降低。张希彪等[8]通过研究人为干扰对黄土高原子午岭油松Pinus tabuliformis人工林土壤物理性质的影响发现:随人为干扰强度的增加,0~50 cm土层土壤砂粒含量均增加,粉粒含量与黏粒含量均下降,土壤容重增加了0.11~0.41 g·cm−3,土壤总孔隙度、毛管孔隙度、非毛管孔隙度、毛管持水量、自然含水量、饱和持水量、田间持水量和土壤有效水含量均下降。人为干扰通过影响土壤物理性质会改变地表径流和土壤侵蚀程度[9],进而影响植物群落分布[10]、物种多样性和生物量[11]、土壤养分含量[12]以及生态系统稳定性[13]。目前,不同程度人为干扰对土壤物理性质的影响多集中在城市绿化用地[14]、山地和森林[15−16]、湖滨湿地[17]等方面,而针对河岸带湿地的研究相对较少。
河岸带是一种生态交错带类型,是水陆交互影响下形成的特殊生境和生态系统,具有防止河岸侵蚀与水土流失、调节洪水、净化水质、维持生物多样性等生态功能[18−19]。河岸带生态系统是一个脆弱的、易受水文、人类活动等干扰而退化的生态区域,具有明显的边缘效应和独特的生态过程[20],且其退化和破坏速度显著快于其他陆地生态系统[21]。农业开垦、城市开发、水利工程、道路和桥梁的修建等人为干扰影响植被类型演替[22−23],造成河岸带退化,加剧水土流失,危害河岸带景观的连续性,破坏了河岸带生态服务功能,严重威胁河岸带生态系统安全与健康[24−26]。
本研究以黄河中下游河岸带为例,根据实地调研结果,对不同干扰强度(轻度干扰、中度干扰、重度干扰)河岸带的土壤物理性质进行调查,分析河岸带土壤物理性质随土层深度的变化规律,对比不同干扰强度河岸带土壤物理性质的差异,探索不同干扰强度河岸带土壤物理性质的相互关系,为河岸带的生态修复和管理提供理论依据。
-
以黄河流域中下游洛阳—郑州段河岸带湿地作为研究对象,地理位置为34.823°~34.980°N,112.777°~113.665°E。研究区春季干旱少雨,夏季炎热多雨,秋季凉爽,冬季气候干燥雨雪稀少。年平均气温为15.0 ℃,最冷月平均气温为0 ℃;最热月平均气温为27.0 ℃,年均降水量为610.0 mm。主要植被类型有芦苇Phragmites australis、白茅Imperata cylindrica、狗牙根Cynodon dactylon、雀麦Bromus japonicus、柽柳Tamarix chinensis等。
-
在黄河洛阳—郑州段选择地形地貌和地表植被覆盖相对一致的河岸带区域,依据人为踩踏程度的不同,选择3种不同干扰强度的样地类型,分别为洛阳市偃师市邙岭乡轻度干扰河岸带、郑州市巩义市S237省道大桥中度干扰河岸带、郑州市黄河湿地公园重度干扰河岸带。样地具体情况如表1所示。其中,洛阳市偃师市邙岭乡河岸带位于黄河保护区内,周围采用栅栏圈围,受人为干扰程度较小,人流量20人·周−1;郑州市巩义市S237省道大桥下河岸带人流量50人·周−1;郑州市黄河湿地公园河岸带人类活动较多,人流量120人·周−1。
表 1 研究区样地概况
Table 1. Basic condition of study piots
干扰强度 地理位置 北纬(N) 东经(E) 海拔/m 优势植被 轻度 洛阳市偃师市邙岭乡 34.823° 112.785° 91 芦苇、白茅、雀麦、狗牙根、柽柳 中度 郑州市巩义市S237省道大桥 34.839° 113.060° 87 芦苇、白茅、雀麦、狗牙根 重度 郑州市黄河湿地公园 34.841° 113.140° 88 白茅、雀麦、狗牙根 2021年10月,在3种不同干扰强度的区域内选择近河岸带200 m×200 m样地,进行5×3网格化划分,共设置15个样点,记录每个样点的经纬度。以每个网格划分点为中心采样点设置5 m×5 m大样方,在每个大样方内随机设置3个1 m×1 m的小样方,用环刀法与土钻法分层采集0~10、10~20、20~30 cm土层样品,分别装入带编号的密封袋内。
-
土壤容重采用环刀法测定,土壤含水率采用烘干法测定,土壤毛管持水量、土壤最大持水量、土壤最小持水量、土壤总孔隙度、土壤毛管孔隙度和土壤非毛管孔隙度采用浸水法测定[1]。
采用Excel 2016和SPSS 25.0软件进行数据处理和分析。采用单因方差分析(one-way ANVOA)检验不同干扰程度土壤物理性质的差异显著性;采用 Pearson 相关分析法分析土壤物理性质之间的相关性;采用Origin 2019绘图。
-
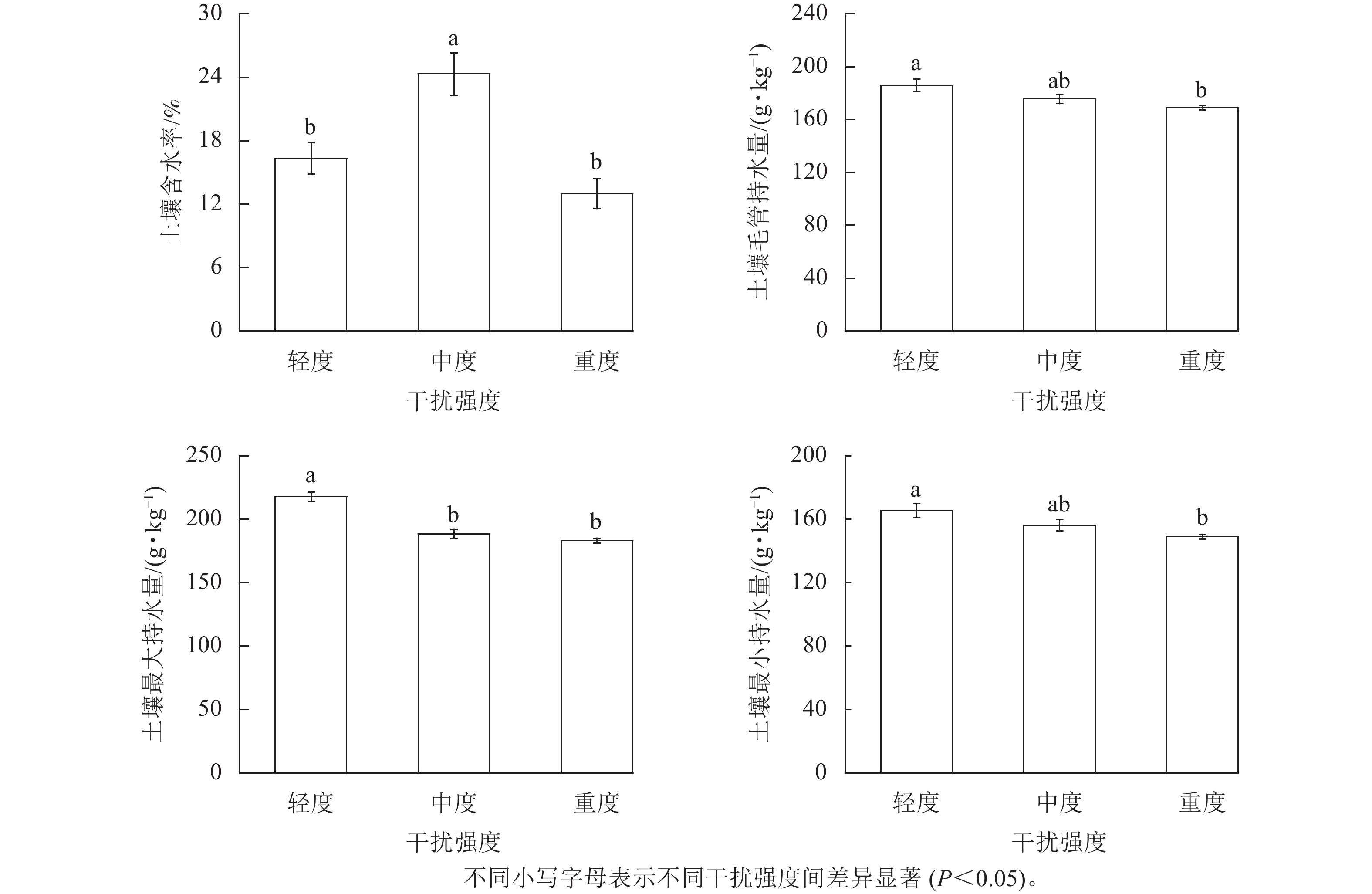
由图1可见:河岸带土壤总孔隙度、土壤毛管孔隙度随人为干扰程度的增加而减小;土壤容重、土壤非毛管孔隙度随人为干扰强度的增加而增加。河岸带0~30 cm土层土壤容重为1.28~1.41 g·cm−3,均值为1.37 g·cm−3,中度、重度干扰土壤容重显著(P<0.05)高于轻度干扰。河岸带0~30 cm土层土壤总孔隙度为44.5%~49.8%,均值为46.6%,中度和重度干扰土壤总孔隙度显著(P<0.05)低于轻度干扰;河岸带0~30 cm土层土壤毛管孔隙度为41.0%~43.1%,均值为42.0%,轻度干扰土壤毛管孔隙度显著(P<0.05)高于重度干扰;河岸带0~30 cm土层土壤非毛管孔隙度为7.1%~14.5%,均值为11.4%,中度、重度干扰土壤非毛管孔隙度显著(P<0.05)高于轻度干扰。
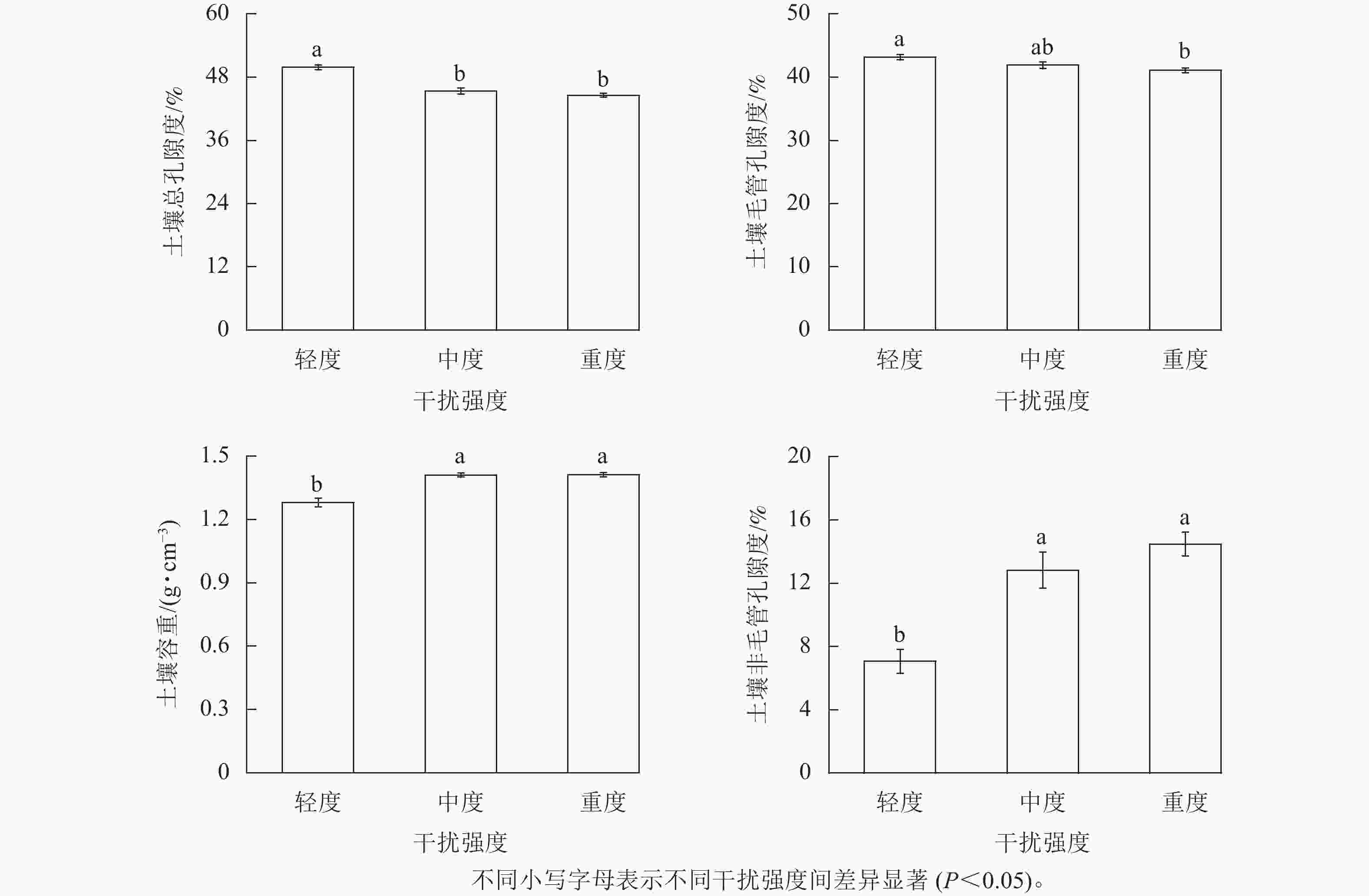
图 1 不同人为干扰强度对黄河河岸带0~30 cm土层土壤容重和土壤孔隙度的影响
Figure 1. Effects of different human disturbance intensity on soil bulk density and porosity of 0−30 cm riparian zone of Yellow River
由表2可见:3种不同人为干扰强度河岸带土壤容重均随土层深度增加而增加,但均无显著差异。轻度、重度干扰的土壤总孔隙度、土壤毛管孔隙度和土壤非毛管孔隙度在3个土层间均无显著差异。中度干扰下,0~10 cm土层土壤总孔隙度、土壤毛管孔隙度显著高于20~30 cm土层,而中度干扰下20~30 cm土层土壤非毛管孔隙度显著(P<0.05)高于0~10 cm土层。
表 2 不同人为干扰程度下土壤容重与土壤孔隙度分布特征
Table 2. Distribution characteristics of soil density and porosity under different degrees of human disturbance
干扰强度 土层深度/cm 土壤容重/(g·cm−3) 土壤总孔隙度/% 土壤毛管孔隙度/% 土壤非毛管孔隙度/% 轻度 0~10 1.24±0.03 a 50.73±0.79 a 43.85±0.82 a 5.42±1.38 a 10~20 1.26±0.02 a 49.38±0.88 a 42.57±0.59 a 8.05±1.18 a 20~30 1.32±0.03 a 49.45±0.82 a 42.86±0.79 a 7.69±1.34 a 中度 0~10 1.40±0.02 a 47.46±1.02 a 43.70±0.73 a 8.83±1.71 b 10~20 1.41±0.02 a 44.92±0.88 ab 41.45±0.87 ab 13.63±1.70 ab 20~30 1.43±0.02 a 43.65±1.13 b 40.39±1.00 b 15.96±2.09 a 重度 0~10 1.41±0.02 a 44.72±0.68 a 41.35±0.61 a 13.93±1.27 a 10~20 1.41±0.01 a 43.81±0.56 a 40.41±1.60 a 15.78±1.14 a 20~30 1.42±0.01 a 45.02±0.79 a 41.32±0.74 a 13.66±1.51 a 说明:不同小写字母表示不同土层间差异显著(P<0.05)。 -
由图2可见:河岸带0~30 cm土层土壤含水率为13.00%~24.29%,均值为17.86%,中度干扰土壤含水率显著(P<0.05)高于轻度、重度干扰。与轻度干扰相比,中度干扰土壤含水率显著增加了32.9%,重度干扰土壤含水率减少了25.43%。河岸带土壤毛管持水量、土壤最大持水量与土壤最小持水量随人为干扰强度的增加而减小。河岸带0~30 cm土层土壤毛管持水量为168.80~185.96 g·kg−1,均值为176.79 g·kg−1,轻度干扰土壤毛管持水量显著(P<0.05)高于重度干扰。河岸带0~30 cm土层土壤最大持水量为182.23~217.87 g·kg−1,均值为196.50 g·kg−1,轻度干扰土壤最大持水量显著(P<0.05)高于中度和重度干扰。河岸带0~30 cm土层土壤最小持水量为148.99~165.50 g·kg−1,均值为156.91 g·kg−1,轻度干扰土壤最小持水量显著(P<0.05)高于重度干扰。

图 2 不同人为干扰强度对黄河河岸带0~30 cm土层土壤水分特征的影响
Figure 2. Effects of different human disturbance intensity on river soil moisture characteristics of 0−30 cm riparian zone of Yellow River
由表3可见:轻度干扰河岸带0~10 cm土层土壤最小持水量显著(P<0.05)高于20~30 cm土层。中度干扰河岸带0~10 cm土层土壤最大持水量显著(P<0.05)高于20~30 cm土层。其他不同土层间土壤含水率、土壤毛管持水量、土壤最大持水量与土壤最小值持水量均无显著差异。
表 3 不同人为干扰强度下土壤水分分布特征
Table 3. Characteristics of soil moisture distribution under different degrees of human disturbance
干扰强度 土层深度/cm 土壤含水率/% 土壤毛管持水量/(g·kg−1) 土壤最大持水量/(g·kg−1) 土壤最小持水量/(g·kg−1) 轻度 0~10 18.58±3.19 a 195.71±3.48 a 227.13±5.87 a 175.99±3.87 a 10~20 16.15±2.37 a 189.83±2.90 a 219.42±5.55 a 169.02±2.02 ab 20~30 14.17±2.13 a 172.33±2.44 a 207.06±6.83 a 151.49±1.49 b 中度 0~10 25.71±3.64 a 186.28±6.18 a 199.37±6.50 a 166.16±6.18 a 10~20 23.79±3.61 a 173.14±5.71 a 186.63±5.49 ab 153.29±5.93 a 20~30 23.36±3.28 a 167.45±5.40 a 179.19±5.16 b 149.27±5.46 a 重度 0~10 13.07±2.67 a 170.32±3.03 a 184.25±3.26 a 150.91±2.59 a 10~20 12.83±2.54 a 166.74±2.27 a 180.90±2.71 a 146.81±2.31 a 20~30 13.09±2.37 a 169.35±3.48 a 184.55±3.84 a 149.26±3.25 a 说明:不同小写字母表示不同土层间差异显著(P<0.05)。 -
相关性分析表明(表4):土壤含水率与土壤毛管孔隙度、土壤最小持水量、土壤毛管孔隙度呈极显著(P<0.01)正相关,与土壤最大持水量呈显著(P<0.05)正相关,与土壤非毛管孔隙度呈显著(P<0.05)负相关;土壤容重与土壤毛管孔隙度、土壤最大持水量、土壤最小持水量、土壤总孔隙度呈极显著(P<0.01)负相关,与土壤非毛管孔隙度呈极显著(P<0.01)正相关。
表 4 黄河河岸带土壤物理性质之间的相关性
Table 4. Correlation between soil physical properties in riparian zone
相关性 含水率 容重 毛管持水量 最大持水量 最小持水量 总孔隙度 毛管孔隙度 容重 −0.067 1 毛管持水量 0.304** −0.508** 1 最大持水量 0.170* −0.788** 0.802** 1 最小持水量 0.366** −0.522** 0.980** 0.775** 1 总孔隙度 0.119 −0.516** 0.613** 0.879** 0.574** 1 毛管孔隙度 0.244** −0.058 0.626** 0.545** 0.608** 0.755** 1 非毛管孔隙度 −0.185* 0.339** −0.659** −0.783** −0.628** −0.953** −0.918** 说明:*表示P<0.05,**表示P<0.01。 -
土壤水分是植被生长的物质基础,影响着植被群落的演替和土壤养分的变化[10]。研究区黄河河岸带的土壤容重(1.28~1.41 g·cm−3)高于纳帕海湖滨湿地(1.00~1.30 g·cm−3)[17]和包头黄河湿地(1.01~1.49 g·cm−3)[27],低于敦煌阳关湿地(1.37~1.50 g·cm−3)[28]。研究区土壤含水率(13.00%~24.29%)低于金马河温江段护岸林地(21.46%~28.02%)[29]、包头黄河湿地(23.63%~57.32%)[27]、敦煌阳关湿地(8.82%~32.54%)[28],表明研究区黄河河岸带湿地的土壤含水量相对较低。
本研究发现:在0~30 cm各土层土壤含水率从高到低均为中度干扰、轻度干扰、重度干扰。张潇月等[29]研究发现:土壤含水量随着干扰强度的增加而减小,这与本研究结果不一致。中度干扰河岸带土壤含水率比轻度干扰显著增加了32.88%,可能是因为中度干扰对河岸带属于增益性干扰,可见适度人为干扰可以提高土壤蓄水保水能力,在土壤肥力和固碳增汇等方面起到更好的作用[30−31]。尤誉杰等[4]研究不同人为干扰措施对天然次生灌丛土壤肥力及蓄水能力的影响发现:适度人为干扰可以提高土壤蓄水保水能力,但人为干扰越强对河岸带土壤水分的负向影响就越大[32]。人为干扰强度增加超过河岸带生态系统承受范围,会导致地表的裸露程度增大,植物对土壤的保水能力下降,地表蒸发随之增加,土壤水分不易保持[9],从而导致重度干扰河岸带土壤含水率比轻度干扰减少了25.4%。土壤含水率对土壤有机质的分解和氮的转化及有效性有积极影响[14],重度人为干扰导致土壤含水率下降,影响植物生长发育,进而可能导致群落逐渐向逆向演替的方向发展,湿生的植被逐渐被中生和旱生的植被所替代。GUITET等[33]通过研究雨林发现:干扰是自然生态系统功能中多样性维持的重要驱动力,零星的强烈干扰及持续性的中等干扰会显著增加景观植物多样性;但当干扰更加频繁或更强烈时,景观植物多样性会略微减少。
土壤水分的蓄持能力和供给状况是限制植物生存的主要条件。土壤容重反映了土壤的紧实度,并直接影响土壤孔隙状况。土壤最大持水量、土壤毛管孔隙度分别反映了土壤的蓄水和供水能力[34],土壤非毛管孔隙直接影响着土壤透气与渗水能力。本研究发现:与轻度干扰相比,中度、重度干扰河岸带土壤容重分别显著增加了9.63%和9.75%;土壤总孔隙度、土壤毛管孔隙度分别显著减少了9.93%和11.98%、2.98%和5.05%,这与李茜[35]、杨大新等[36]的研究结果一致。土壤非毛管孔隙度是指孔隙直径大于0.1 mm的土壤孔隙所占土壤体积的百分比。本研究表明:土壤非毛管孔隙度随人为干扰强度增加而增加,这与张希彪等[8]的研究结果一致。中度和重度干扰河岸带土壤毛管持水量比轻度干扰分别减少了15.64%和18.90%,中度和重度干扰河岸带土壤最大持水量比轻度干扰分别减少了5.88%和10.16%,中度和重度干扰河岸带土壤最小持水量比轻度干扰分别显著减少了5.93%和11.08%,也证明人为干扰强度增加显著降低了河岸带土壤的蓄水和供水能力。这可能是因为随着人为踩踏等干扰强度的增加,河岸带土壤紧实度增加,直接或间接影响了土壤的颗粒组成以及团聚体数量和大小,从而导致土壤保水和持水能力下降,并导致土壤入渗能力降低,土壤的抗蚀能力下降,最终导致土壤退化[8]。
-
相关性分析表明:土壤容重与土壤毛管持水量、土壤最大持水量、土壤最小持水量、土壤总孔隙度呈极显著负相关,与土壤非毛管孔隙度呈极显著正相关。郭宝宝[6]在研究格氏栲Castanopsis kawakamii自然保护区时发现:土壤容重分别与土壤总孔隙度、土壤毛管孔隙度、土壤毛管持水量、土壤最大持水量、土壤最小持水量呈极显著负相关。丛晓峰等[14]在陕西省植物园的研究发现:土壤容重与土壤总孔隙度、土壤通气孔隙度、土壤田间持水量、土壤自然含水量、土壤饱和导水率之间呈极显著负相关。这表明土壤容重与土壤持水量呈负相关。土壤容重随土壤总孔隙度、土壤毛管孔隙度、土壤持水量的减小而显著增加,土壤中储存的有效水就越少,河岸带土壤持水能力就越差[37]。这说明土壤容重可以作为衡量土壤物理性质变化的重要指标之一。
长期和过度的人为干扰会显著影响河岸带土壤的含水量和容重等物理性质,进而对黄河河岸带滩涂湿地生态系统造成不可逆的胁迫和退化。研究表明:植被自然恢复过程能改善土壤的通气透水能力等物理性质[38],从而在一定程度上可以有效改善脆弱和退化生态系统环境[39]。李裕元等[40]在黄土高原北部的交错带研究表明:植被恢复可以降低土壤容重,增加土壤孔隙度,但是短期内(<30 a)植被恢复对土壤颗粒组成并无显著影响,需要长期的植被恢复过程才能有所改善[39]。因此,需要继续加强对不同人为干扰强度下河岸带滩涂湿地土壤特征的监测和研究,并进行合理的生态管理以维持河岸带生态系统的健康[41]。
-
本研究表明:黄河下游河岸带3种不同干扰强度的土壤含水率为13.0%~24.3%,均值为17.9%,中度干扰可以提高土壤含水率。土壤容重为1.28~1.41 g·cm−3,中度、重度干扰土壤容重显著高于轻度干扰。土壤的总孔隙度、毛管孔隙度、毛管持水量、最大持水量和最小持水量均随人为干扰强度的增加而减小。土壤容重、非毛管孔隙度随人为干扰强度的增加而增加。人为干扰对土壤物理性质的影响主要集中在0~10 cm表层土壤。
土壤含水率与土壤毛管孔隙度、土壤最小持水量、土壤毛管孔隙度呈极显著正相关,与土壤最大持水量呈显著正相关,与土壤非毛管孔隙度呈显著负相关。河岸带区域过度的人为干扰会显著影响河岸带土壤的水分含量和容重等物理性质变化,进而造成河岸带滩涂湿地土壤质量的退化。
Comparison of soil physical properties under different human disturbances in the Yellow River riparian zone
-
摘要:
目的 研究人为干扰强度对黄河河岸带土壤物理性质的影响,为黄河河岸带生态系统生态恢复与调控提供科学依据。 方法 2021年10月,在黄河洛阳—郑州段选择地形地貌和地表植被覆盖相对一致的河岸带区域,依据人为踩踏程度的不同,选择3种不同干扰强度的样地类型。按照5×3网格化划分近河岸带200 m×200 m样地,共设置15个样点,系统比较分析不同人为干扰强度(轻度、中度和重度)黄河河岸带土壤容重、土壤含水率、土壤毛管持水量、土壤最大持水量、土壤最小持水量、土壤总孔隙度、土壤毛管孔隙度和土壤非毛管孔隙度等土壤物理性质的差异。 结果 ①随人为干扰强度的增加,土壤容重、土壤非毛管孔隙度逐渐增加;土壤含水率先增加后减小;土壤毛管持水量、土壤最大持水量、土壤最小持水量、土壤总孔隙度与土壤毛管孔隙度逐渐减小。②土壤容重与土壤含水率、土壤毛管孔隙度呈负相关,但未达到显著水平;土壤容重与土壤毛管持水量、土壤最大持水量、土壤最小持水量、土壤总孔隙度呈极显著负相关(P<0.01),与土壤非毛管孔隙度呈极显著正相关(P<0.01)。 结论 过度的人为踩踏等干扰增加了河岸带土壤紧实度,导致土壤物理性质退化以及土壤保水和持水能力降低。图2表4参41 Abstract:Objective This study aims to explore the impact of human disturbance on soil physical properties in the riparian zone, in order to provide a scientific basis for ecological restoration and regulation of the Yellow River riparian ecosystem. Method In October 2021, three types of sample sites with different degrees of disturbance were selected in the Luoyang-Zhengzhou section of the Yellow River based on the degree of human trampling, where the topography and surface vegetation coverage were relatively consistent. The near riparian zone was divided into 200 m×200 m sample plots according to a 5×3 grid, and a total of 15 sample points were set up. Differences in soil physical properties such as bulk density, moisture content, capillary water capacity, maximum water capacity, minimum water capacity, total porosity, capillary porosity, and non-capillary porosity in the Yellow River riparian zone with different levels of human disturbance intensity (mild, moderate and severe) were systematically compared and analyzed. Result (1) With the increase of human disturbance intensity, soil bulk density and non-capillary porosity increased gradually. Soil moisture content first increased and then decreased. Soil capillary water capacity, maximum water capacity, minimum water capacity, total porosity and capillary porosity decreased gradually. (2) Soil bulk density was negatively correlated with soil moisture content and capillary porosity, but it did not reach a significant level. There was a significant negative correlation between soil bulk density and soil capillary water capacity, maximum water capacity, minimum water capacity and total porosity (P<0.01), and a significant positive correlation with non-capillary porosity (P<0.01). Conclusion Excessive human disturbance such as trampling increases the soil compaction in the riparian zone, leading to the degradation of soil physical properties, and a decrease in soil water retention and holding capacity. [Ch, 2 fig. 4 tab. 41 ref.] -
Key words:
- the Yellow River /
- riparian zone /
- human disturbance /
- physical property /
- soil moisture content /
- soil porosity
-
图 1 不同人为干扰强度对黄河河岸带0~30 cm土层土壤容重和土壤孔隙度的影响
Figure 1 Effects of different human disturbance intensity on soil bulk density and porosity of 0−30 cm riparian zone of Yellow River
图 2 不同人为干扰强度对黄河河岸带0~30 cm土层土壤水分特征的影响
Figure 2 Effects of different human disturbance intensity on river soil moisture characteristics of 0−30 cm riparian zone of Yellow River
表 1 研究区样地概况
Table 1. Basic condition of study piots
干扰强度 地理位置 北纬(N) 东经(E) 海拔/m 优势植被 轻度 洛阳市偃师市邙岭乡 34.823° 112.785° 91 芦苇、白茅、雀麦、狗牙根、柽柳 中度 郑州市巩义市S237省道大桥 34.839° 113.060° 87 芦苇、白茅、雀麦、狗牙根 重度 郑州市黄河湿地公园 34.841° 113.140° 88 白茅、雀麦、狗牙根  下载: 导出CSV
下载: 导出CSV
表 2 不同人为干扰程度下土壤容重与土壤孔隙度分布特征
Table 2. Distribution characteristics of soil density and porosity under different degrees of human disturbance
干扰强度 土层深度/cm 土壤容重/(g·cm−3) 土壤总孔隙度/% 土壤毛管孔隙度/% 土壤非毛管孔隙度/% 轻度 0~10 1.24±0.03 a 50.73±0.79 a 43.85±0.82 a 5.42±1.38 a 10~20 1.26±0.02 a 49.38±0.88 a 42.57±0.59 a 8.05±1.18 a 20~30 1.32±0.03 a 49.45±0.82 a 42.86±0.79 a 7.69±1.34 a 中度 0~10 1.40±0.02 a 47.46±1.02 a 43.70±0.73 a 8.83±1.71 b 10~20 1.41±0.02 a 44.92±0.88 ab 41.45±0.87 ab 13.63±1.70 ab 20~30 1.43±0.02 a 43.65±1.13 b 40.39±1.00 b 15.96±2.09 a 重度 0~10 1.41±0.02 a 44.72±0.68 a 41.35±0.61 a 13.93±1.27 a 10~20 1.41±0.01 a 43.81±0.56 a 40.41±1.60 a 15.78±1.14 a 20~30 1.42±0.01 a 45.02±0.79 a 41.32±0.74 a 13.66±1.51 a 说明:不同小写字母表示不同土层间差异显著(P<0.05)。
下载: 导出CSV
表 3 不同人为干扰强度下土壤水分分布特征
Table 3. Characteristics of soil moisture distribution under different degrees of human disturbance
干扰强度 土层深度/cm 土壤含水率/% 土壤毛管持水量/(g·kg−1) 土壤最大持水量/(g·kg−1) 土壤最小持水量/(g·kg−1) 轻度 0~10 18.58±3.19 a 195.71±3.48 a 227.13±5.87 a 175.99±3.87 a 10~20 16.15±2.37 a 189.83±2.90 a 219.42±5.55 a 169.02±2.02 ab 20~30 14.17±2.13 a 172.33±2.44 a 207.06±6.83 a 151.49±1.49 b 中度 0~10 25.71±3.64 a 186.28±6.18 a 199.37±6.50 a 166.16±6.18 a 10~20 23.79±3.61 a 173.14±5.71 a 186.63±5.49 ab 153.29±5.93 a 20~30 23.36±3.28 a 167.45±5.40 a 179.19±5.16 b 149.27±5.46 a 重度 0~10 13.07±2.67 a 170.32±3.03 a 184.25±3.26 a 150.91±2.59 a 10~20 12.83±2.54 a 166.74±2.27 a 180.90±2.71 a 146.81±2.31 a 20~30 13.09±2.37 a 169.35±3.48 a 184.55±3.84 a 149.26±3.25 a 说明:不同小写字母表示不同土层间差异显著(P<0.05)。
下载: 导出CSV
表 4 黄河河岸带土壤物理性质之间的相关性
Table 4. Correlation between soil physical properties in riparian zone
相关性 含水率 容重 毛管持水量 最大持水量 最小持水量 总孔隙度 毛管孔隙度 容重 −0.067 1 毛管持水量 0.304** −0.508** 1 最大持水量 0.170* −0.788** 0.802** 1 最小持水量 0.366** −0.522** 0.980** 0.775** 1 总孔隙度 0.119 −0.516** 0.613** 0.879** 0.574** 1 毛管孔隙度 0.244** −0.058 0.626** 0.545** 0.608** 0.755** 1 非毛管孔隙度 −0.185* 0.339** −0.659** −0.783** −0.628** −0.953** −0.918** 说明:*表示P<0.05,**表示P<0.01。
下载: 导出CSV
-
[1] 赵晓雪, 饶良懿, 申震洲. 砒砂岩区不同地形位置土壤物理性质分异特征[J]. 应用与环境生物学报, 2020, 26(6): 1359 − 1368. ZHAO Xiaoxue, RAO Liangyi, SHEN Zhenzhou. Heterogeneous characteristics of soil physical properties of different terrain locations in the Pisha sandstone area [J]. Chinese Journal of Applied Environmental Biology, 2020, 26(6): 1359 − 1368. [2] 杜姣姣, 周运超, 白云星, 等. 阔叶树种引入后马尾松人工林土壤水文物理性质研究[J]. 水土保持研究, 2021, 28(4): 105 − 112. DU Jiaojiao, ZHOU Yunchao, BAI Yunxing, et al. Research of soil hydrophysical properties of Pinus Massoniana plantation after the introduction of broad-leaved tree species [J]. Research of Soil and Water Conservation, 2021, 28(4): 105 − 112. [3] LI Jiangwen, DU Jing, ZHONG Qin, et al. Changes in the profile properties and chemical weathering characteristics of cultivated soils affected by anthropic activities[J/OL]. Scientific Reports, 2021, 11(1): 20822[2022-10-25]. doi: 10.22541/au.162012734.42874138/v1. [4] 尤誉杰, 王懿祥, 张华锋, 等. 不同人为干扰措施对天然次生灌丛土壤肥力及蓄水能力的影响[J]. 生态学报, 2018, 38(3): 1097 − 1105. YOU Yujie, WANG Yixiang, ZHANG Huafeng, et al. Effects of different human disturbances on soil water conversation and fertility of natural secondary shrub [J]. Acta Ecologica Sinica, 2018, 38(3): 1097 − 1105. [5] JIAO Shuying, LI Junran, LI Yongqiang, et al. Variation of soil organic carbon and physical properties in relation to land uses in the Yellow River Delta, China[J/OL]. Scientific Reports, 2020, 10(1): 20317[2022-10-25]. doi: 10.1038/s41598-020-77303-8. [6] 郭宝宝. 不同人为干扰程度对森林土壤水分物理性质的影响[J]. 亚热带资源与环境学报, 2014, 9(2): 70 − 74. GUO Baobao. Effects of different human interference on forest soil water-physical properties [J]. Journal of Subtropical Resources and Environment, 2014, 9(2): 70 − 74. [7] 庞学勇, 刘庆, 刘世全, 等. 人为干扰对川西亚高山针叶林土壤物理性质的影响[J]. 应用与环境生物学报, 2002, 8(6): 583 − 587. PANG Xueyong, LIU Qing, LIU Shiquan, et al. Effect of human-induced disturbance on soil physical properties of subalpine coniferous forests in western Sichuan [J]. Chinese Journal of Applied and Environmental Biology, 2002, 8(6): 583 − 587. [8] 张希彪, 上官周平. 人为干扰对黄土高原子午岭油松人工林土壤物理性质的影响[J]. 生态学报, 2006, 26(11): 3685 − 3695. ZHANG Xibiao, SHANGGUAN Zhouping. Effect of human-induced disturbance on physical properties of soil in artificial Pinus tabulaeformis Carr. forests of the Loess Plateau [J]. Acta Ecologica Sinica, 2006, 26(11): 3685 − 3695. [9] WANG Ting, XU Qing, GAO Deqiang, et al. Effects of thinning and understory removal on the soil water-holding capacity in Pinus massoniana plantations[J/OL]. Scientific Reports, 2021, 11(1): 13029[2022-10-25]. doi: 10.1038/s41598-021-92423-5. [10] 宋红丽, 牟晓杰, 刘兴土. 人为干扰活动对黄河三角洲滨海湿地典型植被生长的影响[J]. 生态环境学报, 2019, 28(12): 2307 − 2314. SONG Hongli, MOU Xiaojie, LIU Xingtu. Anthropogenic effect on wetland vegetation growth in the Yellow River Delta [J]. Ecology and Environmental Sciences, 2019, 28(12): 2307 − 2314. [11] 曾洪, 陈聪琳, 喻静, 等. 人为干扰对雅安苍坪山公园桉树人工林物种多样性和生物量的影响[J]. 浙江农林大学学报, 2021, 38(2): 253 − 261. ZENG Hong, CHEN Conglin, YU Jing, et al. Effects of human interference on species diversity and biomass of Eucalyptus grandis plantation in Cangping Mountain Park in Ya’an [J]. Journal of Zhejiang A&F University, 2021, 38(2): 253 − 261. [12] 李胜平, 王克林. 人为干扰对桂西北喀斯特山地植被多样性及土壤养分分布的影响[J]. 水土保持研究, 2016, 23(5): 20 − 27. LI Shengping, WANG Kelin. Effect of human disturbance on soil nutrient and plant diversity of grassland in karst mountain [J]. Research of Soil and Water Conservation, 2016, 23(5): 20 − 27. [13] 魏斌, 张霞, 吴热风. 生态学中的干扰理论与应用实例[J]. 生态学杂志, 1996, 15(6): 50 − 54. WEI Bin, ZHANG Xia, WU Refeng. Theoretical analysis of disturbance in ecology and its application [J]. Chinese Journal of Ecology, 1996, 15(6): 50 − 54. [14] 丛晓峰, 刘立成, 王宇超, 等. 不同干扰对城市绿化用地土壤物理性质的影响[J]. 中国农学通报, 2015, 31(28): 166 − 172. CONG Xiaofeng, LIU Licheng, WANG Yuchao, et al. Influence of different interference on physical properties of urban green space soil [J]. Chinese Agricultural Science Bulletin, 2015, 31(28): 166 − 172. [15] 王玲, 冯茂松, 洪志刚, 等. 兴文县香椿人工林土壤物理性质研究[J]. 土壤, 2014, 46(3): 512 − 517. WANG Ling, FENG Maosong, HONG Zhigang, et al. Soil physical properties of Toona sinensis plantation in Xingwen County [J]. Soils, 2014, 46(3): 512 − 517. [16] 严积有, 徐凯然, 申新山. 放牧干扰对人工林土壤物理性状的影响[J]. 水土保持通报, 2008, 28(6): 138 − 141. YAN Jiyou, XU Kairan, SHEN Xinshan. Influences of grazing disturbance on soil physical properties in planted forest [J]. Bulletin of Soil and Water Conservation, 2008, 28(6): 138 − 141. [17] 唐明艳, 杨永兴. 不同人为干扰下纳帕海湖滨湿地植被及土壤退化特征[J]. 生态学报, 2013, 33(20): 6681 − 6693. TANG Mingyan, YANG Yongxing. Analysis of vegetation and soil degradation characteristics under different human disturbance in lakeside wetland, Napahai [J]. Acta Ecologica Sinica, 2013, 33(20): 6681 − 6693. [18] 孙志高, 刘景双, 李彬. 中国湿地资源的现状、问题与可持续利用对策[J]. 干旱区资源与环境, 2006, 20(2): 83 − 88. SUN Zhigao, LIU Jingshuang, LI Bin. The actuality, problems and sustainable utilization countermeasures of wetland resources in China [J]. Journal of Arid Land Resources and Environment, 2006, 20(2): 83 − 88. [19] 郭二辉, 杨喜田, 陈利顶. 河岸带生态功能认知及河流景观偏好的调查研究[J]. 中国园林, 2017, 33(1): 95 − 99. GUO Erhui, YANG Xitian, CHEN Liding. Research of resident’s perceptions to ecological functions of riparian buffers and their preferences for river landscape [J]. Chinese Landscape Architecture, 2017, 33(1): 95 − 99. [20] 郭二辉, 方晓, 马丽, 等. 河岸带农田不同恢复年限对土壤碳氮磷生态化学计量特征的影响——以温榆河为例[J]. 生态学报, 2020, 40(11): 3785 − 3794. GUO Erhui, FANG Xiao, MA Li, et al. Effects of different recovery years on the ecological stoichiometry characteristics of soil carbon, nitrogen and phosphorus in riparian farmland: a case study of Wenyu River [J]. Acta Ecologica Sinica, 2020, 40(11): 3785 − 3794. [21] LAURANCE S G W, BAIDER C, VINCENT FLORENS F B, et al. Drivers of wetland disturbance and biodiversity impacts on a tropical oceanic island [J]. Biological Conservation, 2012, 149(1): 136 − 142. [22] ZHU Xianbin, WANG Lei, ZHANG Xun, et al. Effects of different types of anthropogenic disturbances and natural wetlands on water quality and microbial communities in a typical black-odor river[J/OL]. Ecological Indicators, 2022, 136: 108613[2022-08-25]. doi: 10.1016/j.ecolind.2022.108613. [23] BAI Junhong, ZALEWSKI M, ZHOU Demin, et al. Editorial: estuarine wetland ecohydrology and hydrobiology in Yellow River Delta of China, from prospective of intensive human disturbance [J]. Ecohydrology &Hydrobiology, 2020, 20(4): 473 − 474. [24] CHI Yuan, SHI Honghua, ZHENG Wei, et al. Spatiotemporal characteristics and ecological effects of the human interference index of the Yellow River Delta in the last 30 years [J]. Ecological Indicators, 2018, 89: 880 − 892. [25] 桑凯新, 胡淦林, 黄超, 等. 黄河河岸带5种植物类型根系结构特征对土壤渗透性的影响[J]. 中国水土保持科学, 2020, 18(5): 1 − 8. SANG Kaixin, HU Ganlin, HUANG Chao, et al. Effects of root structure characteristics of 5 plant types on soil infiltration in the Yellow River riparian [J]. Science of Soil and Water Conservation, 2020, 18(5): 1 − 8. [26] MEITZEN K M, PHILLIPS J N, PERKINS T, et al. Catastrophic flood disturbance and a community’s response to plant resilience in the heart of the Texas Hill Country [J]. Geomorphology, 2018, 305: 20 − 32. [27] 杨荣, 塞那, 苏亮, 等. 内蒙古包头黄河湿地土壤碳氮磷含量及其生态化学计量学特征[J]. 生态学报, 2020, 40(7): 2205 − 2214. YANG Rong, SAI Na, SU Liang, et al. Ecological stoichiometry characteristics of soil carbon, nitrogen and phosphorus of the Yellow River wetland in Baotou, Inner Mongolia [J]. Acta Ecologica Sinica, 2020, 40(7): 2205 − 2214. [28] 刘冬, 张剑, 包雅兰, 等. 水分对敦煌阳关湿地芦苇叶片与土壤C、N、P生态化学计量特征的影响[J]. 生态学报, 2020, 40(11): 3804 − 3812. LIU Dong, ZHANG Jian, BAO Yalan, et al. Effects of soil moisture on Phragmites australis leaves and soil C, N and P ecological stoichiometric characteristics in Yangguan wetland, Dunhuang [J]. Acta Ecologica Sinica, 2020, 40(11): 3804 − 3812. [29] 张潇月, 齐锦秋, 张柳桦, 等. 人为干扰对金马河温江段护岸林物种多样性和土壤理化性质的影响[J]. 植物研究, 2019, 39(1): 78 − 86. ZHANG Xiaoyue, QI Jinqiu ZHANG Liuhua, et al. Effects of human disturbance on species diversity and soil physical and chemical properties of revetment forest in Wenjiang Section of the Jinma River [J]. Bulletin of Botanical Research, 2019, 39(1): 78 − 86. [30] 何其华, 何永华, 包维楷. 岷江上游干旱河谷典型阳坡海拔梯度上土壤水分动态[J]. 应用与环境生物学报, 2004, 10(1): 68 − 74. HE Qihua, HE Yonghua, BAO Weikai. Dynamics of soil water contents on south-facing slope of dry valley area in the upper reaches of the Minjiang River [J]. Chinese Journal of Applied and Environmental Biology, 2004, 10(1): 68 − 74. [31] 姚俊宇, 齐锦秋, 张柳桦, 等. 人为干扰对碧峰峡山矾次生林群落物种多样性和土壤理化性质的影响[J]. 生态学杂志, 2018, 37(10): 2942 − 2950. YAO Junyu, QI Jinqiu, ZHANG Liuhua, et al. Effects of anthropogenic disturbance on species diversity and soil physicochemical properties of Symplocos sumuntia secondary forest in Bifengxia [J]. Chinese Journal of Ecology, 2018, 37(10): 2942 − 2950. [32] 张荣, 李婷婷, 金锁, 等. 人为干扰对蒙顶山木荷次生林物种多样性及土壤理化性质的影响[J]. 浙江农林大学学报, 2020, 37(5): 867 − 875. ZHANG Rong, LI Tingting, JIN Suo, et al. Effects of human disturbance on species diversity and soil physiochemical properties of Schima superba community in Mengding Mountain [J]. Journal of Zhejiang A&F University, 2020, 37(5): 867 − 875. [33] GUITET S, SABATIER D, BRUNAUX O, et al. Disturbance regimes drive the diversity of regional floristic pools across Guianan Rainforest Landscapes[J/OL]. Scientific Reports, 2018, 8(1): 3872[2022-10-25]. doi: 10.1038/s41598-018-22209-9. [34] 吴昊, 马昕昕, 肖楠楠, 等. 土壤物理性质对秦岭松栎林建群种形态及物种多样性的影响[J]. 土壤, 2020, 52(5): 1068 − 1075. WU Hao, MA Xinxin, XIAO Nannan, et al. Effects of soil physical properties on morphological traits of constructive trees and species diversity of pine-oak mixed forest in Qinling Mountains [J]. Soils, 2020, 52(5): 1068 − 1075. [35] 李茜. 旅游干扰对长白山景区土壤性质及植被群落的影响[J]. 山东农业大学学报(自然科学版), 2015, 46(1): 74 − 77. LI Qian. Impacts of tourist disturbance on soil properties and plant communities in the natural reservation area of Changbai Mountain [J]. Journal of Shandong Agricultural University (Natural Science Edition), 2015, 46(1): 74 − 77. [36] 杨大新, 巩合德, 潘洋, 等. 人为干扰对轿子山自然保护区典型植物群落及土壤理化性质的影响[J]. 科学技术与工程, 2016, 16(32): 8 − 14. YANG Daxin, GONG Hede, PAN Yang, et al. Effects of human disturbance on plant communities and physicochemical soil properties in the natural reservation area of Jiaozi mountain [J]. Science Technology and Engineering, 2016, 16(32): 8 − 14. [37] 杨益帆, 胡宗达, 李亚非, 等. 川西亚高山川滇高山栎灌丛地被物与土壤持水性能[J]. 应用与环境生物学报, 2020, 26(4): 951 − 960. YANG Yifan, HU Zongda, LI Yafei, et al. Water conservation capacity of ground cover and soils in the subalpine Quercus aquifolioides shrubs in of western Sichuan, China [J]. Chinese Journal of Applied and Environmental Biology, 2020, 26(4): 951 − 960. [38] 李松阳, 余杭, 罗清虎, 等. 灾害干扰受损区自然恢复初期土壤物理性质的变化特征[J]. 水土保持学报, 2020, 34(5): 162 − 168. LI Songyang, YU Hang, LUO Qinghu, et al. Change characteristics of soil physical properties at the initial natural recovery stage in disaster disturbed and damaged areas [J]. Journal of Soil and Water Conservation, 2020, 34(5): 162 − 168. [39] 李静鹏, 徐明锋, 苏志尧, 等. 不同植被恢复类型的土壤肥力质量评价[J]. 生态学报, 2014, 34(9): 2297 − 2307. LI Jingpeng, XU Mingfeng, SU Zhiyao, et al. Soil fertility quality assessment under different vegetation restoration patterns [J]. Acta Ecologica Sinica, 2014, 34(9): 2297 − 2307. [40] 李裕元, 邵明安, 陈洪松, 等. 水蚀风蚀交错带植被恢复对土壤物理性质的影响[J]. 生态学报, 2010, 30(16): 4306 − 4316. LI Yuyuan, SHAO Min’an, CHEN Hongsong, et al. Impacts of vegetation recovery on soil physical properties in the cross area of wind-water erosion [J]. Acta Ecologica Sinica, 2010, 30(16): 4306 − 4316. [41] 雷斯越, 郭晋伟, 何亮, 等. 黄土丘陵区退耕坡面草地恢复与土壤物理性质空间分异特征[J]. 水土保持学报, 2021, 35(1): 251 − 258. LEI Siyue, GUO Jinwei, HE Liang, et al. Spatial differentiation characteristics of grassland restoration and soil physical properties of slope in hilly-gully region of Loss Plateau [J]. Journal of Soil and Water Conservation, 2021, 35(1): 251 − 258. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220727
 点击查看大图
点击查看大图
计量
- 文章访问数: 1492
- HTML全文浏览量: 517
- PDF下载量: 38
- 被引次数: 0
