




-
自1979年以来,中国乡村空间结构经历着剧烈的重构与分离,导致了生活空间、生态空间、生产空间的分散化、无序化、污损化等问题[1]。在面临大量内外刺激的变动中[2],乡村振兴战略[3]是当前乡村空间活力提升与重构的政策导向[4]。然而乡村振兴战略实施以来,是否推动了乡村可持续发展还犹未可知[5]。探索乡村空间活力的内涵和评价方法,并因地制宜提出活化策略,是中国解决乡村空间发展不平衡不充分问题的迫切需要。
在此背景下,针对乡村空间活力,国内外学者主要从乡村空间理论研究、乡村空间活力维系与提升、乡村公共空间活力评价、国际经验借鉴[6−8]等方面开展了系统研究。随着对乡村空间特征与所面临问题的深入探索,学界把关注重心逐步转向了定量研究。从研究内涵上看,将重点从乡村振兴内涵目标[9]、城乡融合[10]、乡村性[11]及乡村发展潜力[12]角度展开;从研究方法上看,采用“定性+定量”“主观+客观”的方法,一般涉及专家打分法、层次分析法、熵值法、因子分析法[13−15];从研究尺度上看,学者大多以全国、省(市)域等宏观层面为主[11],近年来有向县域、村域等微观尺度深入的趋势[9, 12]。乡村发展评价一直是政府和学术界关注的焦点,国外较早开展系统性的乡村发展评价研究[16−18],其中,乡村空间活力评价作为诊断乡村振兴状态、制定乡村政策、指导乡村科学发展的重要技术手段,在英国、日本、德国已得到广泛应用[6]。然而,中国有关乡村空间活力内涵及其评价研究目前还较为欠缺,且缺乏对发达省份的欠发展区域的研究。
为了深入解析山区县域乡村空间活力,本研究以浙江省典型山区县泰顺县为例,根据各乡(镇)调研数据,构建乡村空间活力评价指标体系,评价分析泰顺县各乡(镇)的乡村空间活力特征与差异,并因地制宜地提出差异化提升策略,旨在为推进县域乡村振兴战略实施、山区跨越式高质量发展提供参考,以期早日实现共同富裕目标。此外,本研究对浙江省相对欠发达地区实现乡村振兴发展有一定的指导作用,在一定程度上也完善了乡村空间活力理论与评价研究。
-
泰顺县地处浙江省最南端,温州市西南部,地理区位特殊,自然条件复杂,生态环境脆弱,经济发展和城镇化水平较低,区域发展差异大。在浙江省提出高质量发展,建设共同富裕示范区背景下,泰顺县实施乡村空间活力提升,促进乡村振兴发展在整个浙江省相对欠发达地区具有重要意义。
本研究范围为泰顺县全域12镇7乡,包含罗阳镇、司前畲族镇、百丈镇、筱村镇、泗溪镇、彭溪镇、雅阳镇、仕阳镇、三魁镇、南浦溪镇、龟湖镇、西旸镇12镇和包垟乡、东溪乡、凤垟乡、柳峰乡、雪溪乡、大安乡、竹里畲族乡7乡,总面积为1 768 km2。
考虑研究区数据的可获得性、科学性和代表性,选取2020年作为时间断面,涉及数据主要包括空间矢量数据、遥感影像数据、统计数据和实地调查资料,空间矢量数据来源于中国科学院资源环境科学数据中心(
https://www.resdc.cn/ );遥感影像数据来源于地理空间数据云2020年10月Landsat 8卫星遥感影像(http://www.gscloud.cn/ );统计数据及相关指标数据来源于高德开放平台、浙江省人民政府网(http://www.zj.gov.cn/ )、泰顺县人民政府网(http://www.ts.gov.cn/ )、2020年温州市、泰顺县统计年鉴以及通过遥感影像数据运算后得到的结果。 -
乡村空间活力指数评价体系由多种指标构成,为了提高数据统计结果的可信度,先对所有评估指数进行极大值标准化处理,以消除各数据集的量纲影响,其计算过程参考相关研究成果[19−20]。正向指标:
$$ {R}_{ij}'=\frac{{R}_{ij}-\mathrm{m}\mathrm{i}\mathrm{n}\left({R}_{ij}\right)}{\mathrm{m}\mathrm{a}\mathrm{x}\left({R}_{ij}\right)-\mathrm{m}\mathrm{i}\mathrm{n}\left({R}_{ij}\right)}。$$ (1) 逆向指标:
$$ {R}_{ij}'=\frac{\mathrm{m}\mathrm{a}\mathrm{x}\left({R}_{ij}\right)-{R}_{ij}}{\mathrm{m}\mathrm{a}\mathrm{x}\left({R}_{ij}\right)-\mathrm{m}\mathrm{i}\mathrm{n}\left({R}_{ij}\right)}\mathrm{。} $$ (2) 式(1)~(2)中:$ {R}_{ij}' $为i镇j项指标数据标准化值;$ {R}_{ij} $为实际值;$ \mathrm{m}\mathrm{a}\mathrm{x}\left({R}_{ij}\right) $和$ \mathrm{m}\mathrm{i}\mathrm{n}\left({R}_{ij}\right) $分别为i镇j项指标的最大值和最小值。
采用熵值法确定各个指标权重$ {w}_{j} $,其计算过程参考相关研究成果[12, 19],并计算各乡(镇)乡村空间活力指数(Vi):
$$ {V}_{i}=\sum _{j=1}^{n}{R}_{ij}\times {w}_{j}\mathrm{。} $$ (3) 式(3)中:$ {w}_{j} $为j项指标权重;$ {V}_{i} $为乡村空间活力指数,数值越大表明乡村空间活力越高。
最后利用ArcGIS 10.6将数据按自然间断点分级法把乡村空间活力指数水平等级划分为匮乏区(0.11<Vi≤0.18)、一般区(0.18<Vi≤0.28)和活力区(0.28<Vi≤0.53) 3类。数值越高,乡村空间活力越高。
-
乡村空间活力提升路径探索必须首先明确乡村地域特征与差异,在科学划分类型的前提下,分类施策、精准发力。基于乡村空间活力内涵解读和乡村空间活力评价结果,并参考龙花楼等[11]、张利国等[21]乡村类型识别方法,深入分析乡村空间活力类型特征,进而能够更加有效推动不同类型山区乡村空间活力的提升,促进山区人民走向共同富裕。具体判断过程如下:
$$ \left\{\begin{array}{*{20}{c}}\dfrac{{R}_{p}}{{A}_{p}+{S} _{p}}\geqslant 0.75 & 第p项活力为该乡(镇)主导活力\\ 0.5\leqslant \dfrac{{R}_{p}}{{A}_{p}+{S} _{p}}<0.75 & 第p项活力为该乡(镇)非主导活力\\ \dfrac{{R}_{p}}{{A}_{p}+{S} _{p}}<0.50 & 第p项活力为该乡(镇)短板活力\end{array}\right.。$$ (4) 式(4)中:Rp为该乡(镇)第p项活力值;Ap 为所有乡(镇)第p项活力的均值;Sp 为所有乡(镇)第p项活力的标准差。识别主导活力、非主导活力及短板活力之后,按表1的划分原则对各乡(镇)乡村空间活力类型进行划分。当3种空间活力同时为某类活力时,划分为综合发展型。在此条件下,若3种空间活力同时为主导活力则划分为强综合型,若同时为短板活力则划分为弱综合型,若同时为非主导活力则划分为协调综合型。当存在且少于3种主导活力时,按是否存在短板活力划分为极化发展型和主导发展型,并以较强的主导活力或更弱的短板活力划分亚类。当不存在主导活力且存在多个短板活力时,划分为欠缺发展型,以更弱的短板活力划分亚类。
表 1 泰顺县乡村空间活力发展类型划分原则
Table 1. Classification of rural spatial vitality levels in Taishun County
是否存在
主导活力是否存在
短板活力主导活力项数/项 活力发展类型 活力发展亚类 是 否 0<N<3 主导发展型 按主导活力类型划分 是 0<N<3 极化发展型 主导活力-短板活力 否 是 N=0 欠缺发展型 按短板活力类型划分 − − N=0或N=3或n=3 综合发展型 若N=3,则为强综合型;若N=0,则为弱综合型;若n=3,则为协调综合型 说明:N为主导活力项数,n为非主导活力项数;−表示不确定,需根据综合发展类型判断。 -
生物的活力是由自身的生存能力和外部环境的相互作用所决定的。现如今,“活力”概念已被广泛应用,如城市活力、人才活力等[22−23]。而生活空间、生态空间、生产空间(“三生”空间)是中国建立陆地空间规划体系所要涉及的首要概念,是衔接、整合与协调各层级规划的核心[24]。
“乡村空间活力”是以乡村“三生”空间为载体,以乡村地域空间发展状况以及各个空间系统发挥的效能为载体,且相互作用所表征出来的能量形式,是一个复杂的综合性概念。本研究将乡村空间活力划分为乡村生活空间活力、生态空间活力、生产空间活力等3个子系统。其中,乡村生活空间活力所对应的载体——生活空间,是乡村空间活力存续的核心,是以培育乡村发展社会行为主体(人)、完善公共服务和社会保障体系、传承乡土文化、缩小城乡差距为主要内涵[1];乡村生态空间活力所对应的载体——生态空间,是乡村空间活力存续的先决条件,始终是各类型空间及其相互作用发展的根本立足点,提供生活、生产活动的空间载体,既养育了人,也塑造了相应产业;乡村生产空间活力所对应的载体——生产空间,是乡村空间活力的驱动因素,以产业培育为核心,并随着乡村经济基础能力的提高,为生态空间活力、生活空间活力提升创造物质基础。
激活乡村空间活力是实现乡村振兴战略的重要保障,亦是乡村振兴的关键路径,其核心是构建生态空间、生活空间、生产空间的耦合格局,促进其协调共振发展。在当今乡村转型发展的关键时期,应当首先认识和探索乡村空间活力的内涵和评价方法,构建一套科学合理的评价体系,评价并探析乡村空间活力的特征与差异,进而因地制宜地实施活化策略。通过主体的干预与调控,重组乡村要素、优化空间结构、提升地域功能,以实现乡村生态空间、生活空间和生产空间的协调重构与活力提升,最终使乡村振兴乃至共同富裕目标得以实现。
-
为全面体现研究区域内乡村空间活力水平的实际状况,按照科学性和代表性等指标构建原则,本研究在参考冯旭等[6]、何焱洲等[20]的研究以及《美丽乡村建设指南》《国家乡村振兴战略规划(2018—2022)》《国家质量兴农战略规划(2018—2022)》《浙江省山区26县跨越式高质量发展实施方案》等综合设置的基础上,以城乡规划学科的研究视角,基于“三生”空间理论,以乡村振兴战略总要求为指导,从生活空间活力、生态空间活力、生产空间活力等3个维度构建乡村空间活力指数评价体系,包括17个指标(表2)。①乡村生活空间主要是乡村居民日常生活的场所,乡村空间的宜居性以及居民幸福感是生活空间活力的核心需求。乡村空间人居环境、设施建设的完善程度、人文活动的开展情况等是乡村生活空间活力的根本体现。选取美丽乡村精品村占比、文化旅游体育与传媒占总支出比例、人均一般公共预算收入、金融服务水平、医疗服务水平、生活服务水平、教育服务水平等7项指标综合体现乡村生活空间的活力水平和宜居程度。其中,美丽乡村精品村占比表明乡村生活整体水平;文化旅游体育与传媒支出占总支出比例、人均一般公共预算收入代表政府的经济实力与扶持力度,数值越大说明乡村生活水平具有更大的提升潜力;金融、医疗、生活、教育服务水平从各个方面展现县域对乡村设施的支持能力,同时也能说明村民的生活便利程度。②乡村生态空间活力主要体现为乡村本底自然生态资源条件以及生态资源的保护利用程度,因此,植被覆盖率、3A景区村庄数占比、地均生态服务价值、土地开发强度、休闲农业基础设施保障等5项指标表现乡村生态空间的活力。指数值越高,生态空间的稳定性与价值转换的可持续性越高。其中,3A景区村庄占比表明整体乡村生态水平;植被覆盖率、地均生态服务价值表明乡村本底自然资源条件;土地开发强度、休闲农业基础设施保障代表乡村自然资源利用程度与能力。③乡村生产空间的核心任务在于牢牢守住保障国家粮食安全和不发生规模性返贫2条底线。农业的稳产增产能力和一二三产业的融合发展水平是乡村生产空间活力的重要表征内容,因此选取人均耕地面积、村均经营性集体经济、农林水事务支持强度、休闲旅游功能、工业企业个数等5项指标表征乡村生产空间活力,其值越高,说明越有利于维持乡村生产空间系统的稳定。其中,人均耕地面积体现农业的发展水平与潜力;工业企业个数表明乡村第二产业的发展情况;村均经营性集体经济、农林水事务支持强度、休闲旅游功能则用来综合体现乡村一二三产业的融合发展水平。
表 2 乡村空间活力指数评价体系
Table 2. Evaluation system of rural spatial vitality index
目标层 准则层 指标层 性质 指标意义 权重系数 乡村空间活力 生活空间活力
(0.385)美丽乡村精品村占比/% + 精品村数量/行政村总数[来源泰顺县人民政
府网]0.043 文化旅游体育与传媒支出占
总支出比重/%+ 文化旅游体育与传媒支出/政府财政总支出[来
源人民政府部门决算公开]0.065 人均一般公共预算收入/(元·人−1) + 一般公共预算收入/2020年户籍人口 0.073 金融服务水平/家 + 万人拥有金融机构网点数 0.082 医疗服务水平/家 + 万人拥有医疗机构数 0.050 生活服务水平/个 + 万人拥有商店、超市、农贸市场数 0.045 教育服务水平/个 + 万人拥有学校数 0.027 生态空间活力
(0.206)植被覆盖率/% + (林地面积+草地面积)/县域总面积 0.031 3A景区村庄数占比/% + 3A景区村庄数/行政村总数 0.037 地均生态服务价值/(元·hm−2) + 区域生态服务价值总和/镇域面积 0.056 土地开发强度 − 建设用地面积/镇域面积 0.016 休闲农业基础设施保障/万元 + 区域农林水务投入 0.066 生产空间活力
(0.409)人均耕地面积/(hm2·人−1) + 耕地面积/2020年户籍人口 0.048 村均经营性集体经济/(万元·村−1) + 村经营性集体经济总量/行政村总数 0.125 农林水事务支持强度/(万元·hm−2) + 农林水事务支出/耕地面积 0.054 休闲旅游功能/个 + 万人拥有农家乐个数、观光园个数 0.055 工业企业个数/个 + 工业企业的户数 0.126 说明:指标性质相对于目标层而确定,其中+表示正向指标,−表示负向指标。地均生态服务价值通过谢高地等[25]方法计算,2020年户籍人口根据2019年户籍人口预测。 -
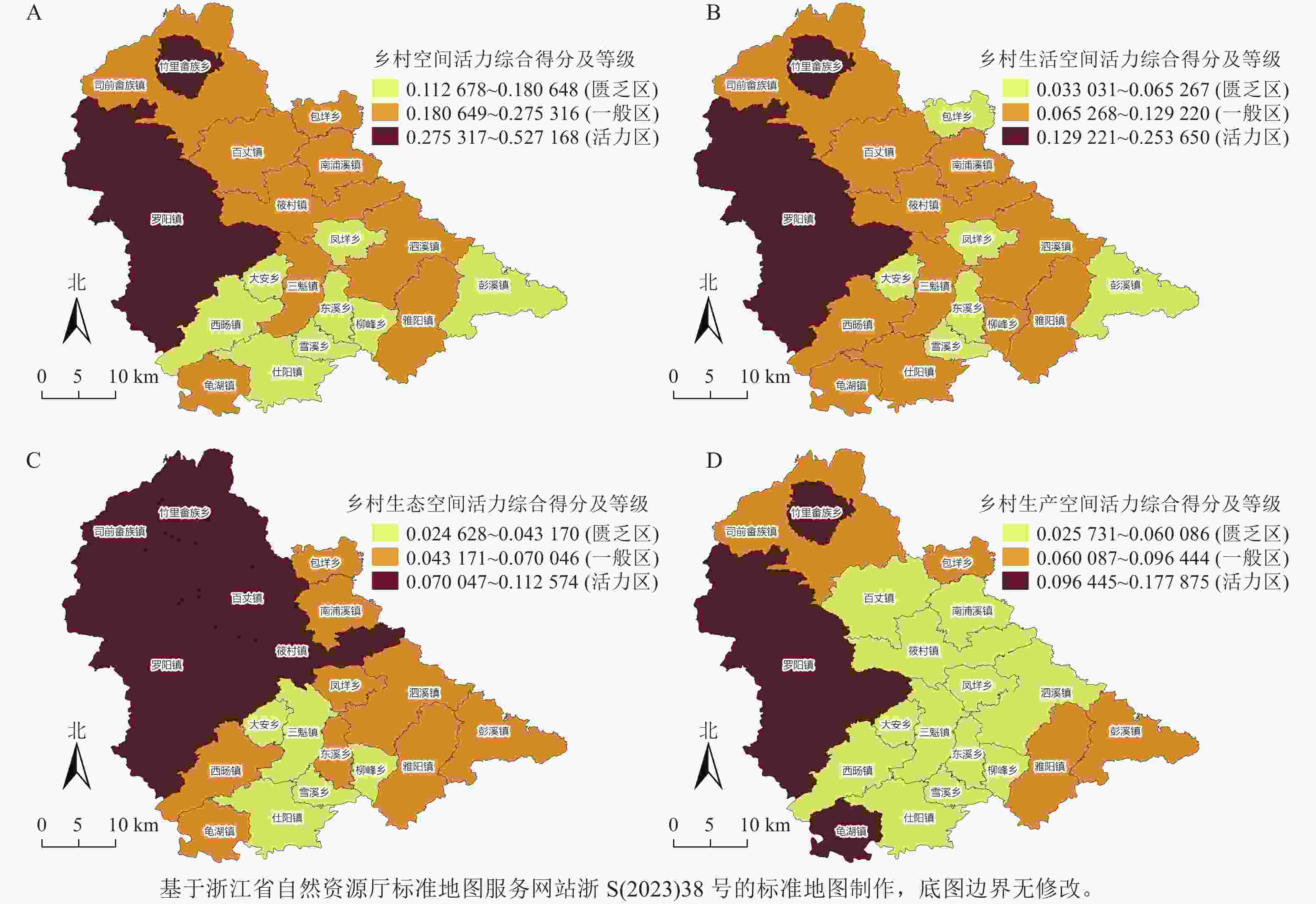
图 1 乡村空间活力和各子系统活力的空间分布
Figure 1. The rural spatial vitality and the spatial distribution of the vitality of each subsystem
-
匮乏区的乡(镇)占泰顺县的42.11%,集中分布于泰顺县南部,乡村活力差异相对较小;一般区的乡(镇)占泰顺县的47.37%,包括位于东北部的百丈镇、包垟乡等8个乡(镇)和西南部的龟湖镇;活力区的乡(镇)仅占泰顺县的10.53%,包括活力最高的竹里畲族乡和县政府驻地罗阳镇。从整体上看,呈“局部高值,北高南低”的空间分布特征。生产空间活力指数总体均值为0.073 (13个地区未达到均值),生态空间活力指数总体均值为0.061 (12个地区未达到均值),生活空间活力指数总体平均值为0.063 (12个地区未达到均值),综合活力水平为0.229 (12个地区未达到均值),大部分乡(镇)都未达到平均水平,说明泰顺县乡村空间活力总体比较低,生态空间活力、生产空间活力和生活空间活力均较低(表3,图1A)。
表 3 泰顺县各乡镇乡村空间活力发展类型统计
Table 3. Statistics of spatial vitality development types of villages and towns in Taishun County
活力发展类型 活力发展亚类 乡(镇) 现状特征 乡镇数量/个 综合发展型 强综合型 竹里畲族乡、罗阳镇 各类活力水平较为均衡,乡村空间活力水平高 2 弱综合型 大安乡、雪溪乡 各类活力水平较为均衡,乡村空间活力水平低 2 主导发展型 生活主导型 泗溪镇 各类公共服务和配套设施完善,民生福祉不断增进 1 生态主导型 司前畲族乡 境内竹木、水力资源丰富,生态环境较好 1 生产主导型 龟湖镇、雅阳镇 区位优势突出,乡村集体经济和工业发展较好 2 极化发展型 生态主导-生活滞后型 筱村镇、包垟乡 具有丰富的自然、人文资源,主要以农业为主,但公共服务和基础设施建设水平较低,对居民文化生活的关注略显不足 2 生态主导-生产滞后型 百丈镇 生态环境优越,以农业生产为主,但农村收入较低,产业发展存在一定问题 1 欠缺发展型 生活滞后型 彭溪镇 生活配套设施建设较为落后 1 生态滞后性 柳峰乡、仕阳镇 生态环境较差,整体乡村生态空间活力也居于末位 2 生产滞后型 南浦溪镇、三魁镇、西旸
镇、凤垟乡、东溪乡工业和休闲旅游业务发展均较弱 5 -
从生活空间活力水平分析来看(图1B),总体呈“局部低值,西高东低”的空间分布特征。共有13个乡(镇)的乡村生活空间活力为活力区和一般区,占全县的68.42%,其中活力区的乡(镇)只有竹里畲族乡和罗阳镇,而包垟乡、大安乡等6个乡(镇)的乡村生活空间活力均较为匮乏,且差距较大。罗阳镇和竹里畲族乡的分值高达0.221和0.254,第3位的泗溪镇仅为0.129。可以发现泰顺县整体的基础设施和公共服务设施建设分布不均,例如罗阳镇、泗溪镇万人有医院数为17.09、10.59家,而竹里畲族乡、东溪乡、雪峰乡均为0,万人拥有商店、超市、农贸市场数亦是如此,罗阳镇高达20.21家,雪溪乡低至0.99家。
-
从生态空间活力水平来看(图1C),总体呈“北高南低”的空间分布特征。活力区集中连片,包括罗阳镇、百丈镇等5个乡(镇),这些乡(镇)地均生态服务价值均高于平均值3.11万元·hm−2,百丈镇优势最为显著,其值高达4.86万元·hm−2。此外,活力区的休闲农业基础设施保障也较为完善,区域农林水事务支持强度为221.28万元·hm−2,远高于研究区均值161.22万元·hm−2。在投入过程中积极融合当地的畲族文化和生态资源,并与全域旅游相互衔接,为激发当地乡村空间活力提供了坚实的保障。一般区与匮乏区大都位于泰顺县南部,生态条件较北部略显欠缺,且生态保护与开发投入不足,区域农林水务投入和地均生态服务价值均值仅为139.78、2.96万元·hm−2。
-
乡村生产空间活力水平总体呈“局部突出,四周高中间低”的空间分布特征(图1D)。活力区位于罗阳镇、竹里畲族乡和龟湖镇,各自都有产业发展优势。乡村生产空间活力匮乏区占泰顺县的一半以上,高达63.16%,包括东溪乡、凤垟乡、大安乡等12个乡(镇),这些乡(镇)的工业与休闲旅游业发展能力十分薄弱,工业企业个数和农家乐观光园个数平均为12.75、2.18个,与活力区(77.00、3.75个)相差悬殊,缺少支柱性特色产业与产业发展引导,且正处于旅游开发的初期,未能形成完善的产业体系,导致村集体经济发展落后(活力区和匮乏区村均经营性集体经济均值分别为0.58和0.17万元·村−1),村民收入低,人口流失严重,使得这些乡(镇)的乡村生产空间活力在全县处于弱势。
-
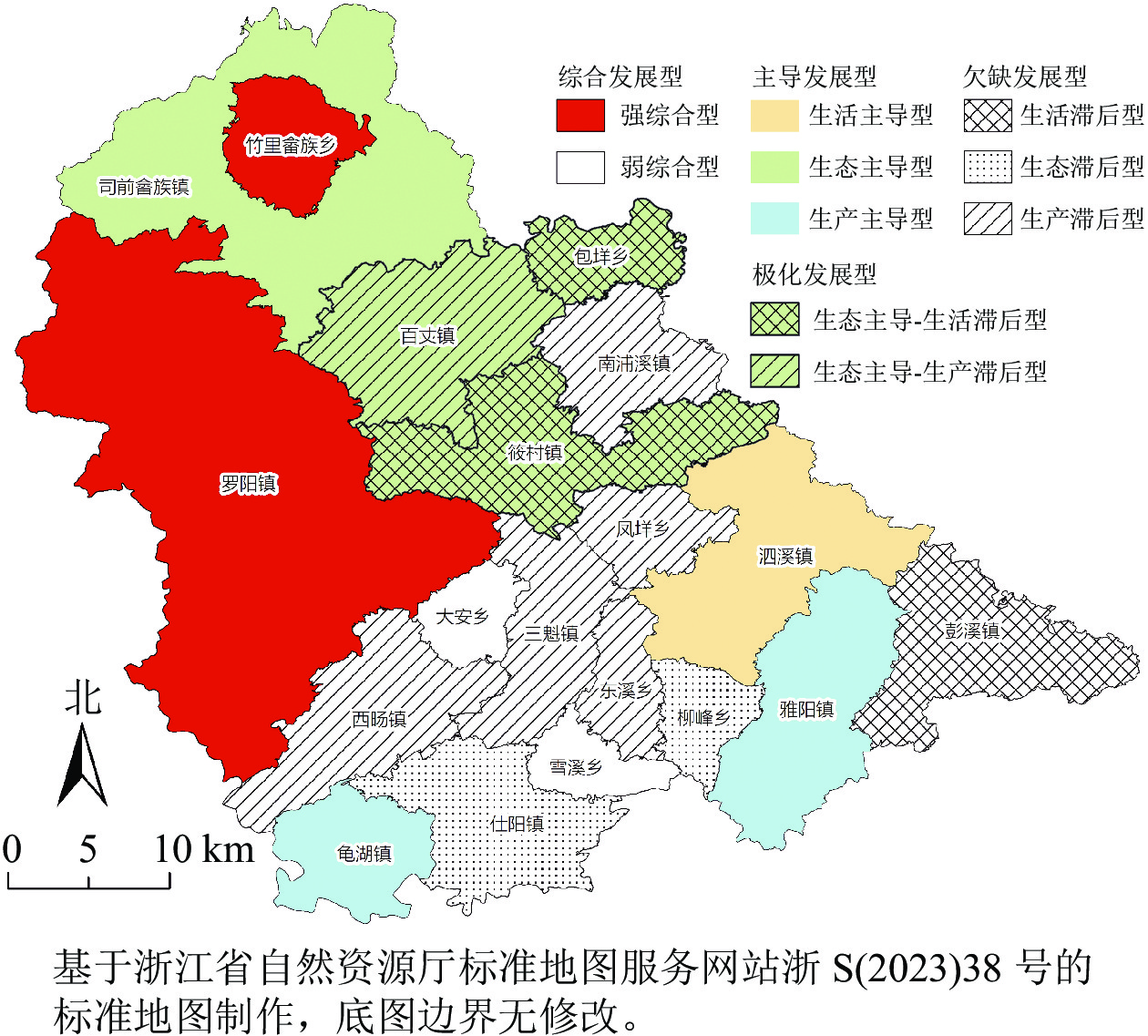
通过上述划分原则,最终确认活力发展类型共4类,包括活力发展亚类10类,各类活力发展类型乡(镇)统计和现状特征如表3所示,其空间分异特征见图2。
图 2 泰顺县乡村空间活力发展类型空间分异
Figure 2. 2 Spatial differentiation of rural spatial vitality development types in Taishun County
-
综合发展型乡(镇)是指生活空间、生态空间、生产空间活力水平较为均衡的乡(镇)。研究区的综合发展型乡(镇)共4个,分布较为分散,主要位于泰顺县北部和南部的边界处,与其他县域、省域相邻。因活力水平层次存在较大差异性,又将综合发展型划分为强综合型(2个)和弱综合型(2个)。强综合型乡(镇)各类活力均为主导活力,而弱综合型乡(镇)各类活力均为短板活力。对于这类乡(镇),应高效利用各自的优势资源,形成强对弱的对点帮扶,联合活化,推进泰顺县乡村空间活力的跨越式提升。①强综合型。该类乡(镇)有2个,包括竹里畲族乡和罗阳镇,都位于泰顺县北部,紧邻G235国道,交通便利且地势较为平坦。这2个乡(镇)的乡村空间活力水平均较高,医疗、教育、金融等公共服务与基础设施建设完善,农业发展水平较高,生态资源以及人文资源均十分优越,提前依靠自身优势进行全域旅游开发,推进一二三产业融合发展,形成了具有特色的产业发展带。依托其区位交通优势,立足共同富裕的长远目标,制定合理的活力提升路径既有助于扩大自身活力优势,也能发挥辐射带动作用,助推周边地区乡村空间活力提升。未来应以全域旅游快速发展为基础,依托现有优势生态资源,推动当地发展医疗、康养等健康产业;高效利用各自的优势资源,积极与周边区域融合发展、联合活化;利用好现有产业园、特色小镇等平台,并建设具备项目孵化、人才引进等平台载体,推动返乡创业、就近就业、创业带动就业。②弱综合型。该类乡(镇)有2个,包括大安乡和雪溪乡,总体活力水平(0.130)远低于平均值(0.229),与强综合型乡(镇)形成鲜明两级对比。这类乡(镇)的乡村空间活力提升难度较大,需要大量的资本、人力推动发展,同时也需要周边乡(镇)的协同融合发展。因此,可以重点从以下几方面展开:鼓励支持强综合型乡(镇)与弱综合型乡(镇)的结对帮扶,尽快完善基础设施、公共服务设施、医疗卫生体系、社会保障体系建设;发展山区特色生态农业,植入农业观光、采摘等低成本体验型业态,带动当地村民就地就业;因地制宜推广“大搬快聚富民安居”工程,推进撤村并居,走出以中心村为发展核心,以一般村为补充的具有山区特色的乡村振兴之路。
-
主导发展型乡(镇)是指有一类活力具有较强的发展优势,其他各类活力相对协调且不存在短板活力的乡(镇)。研究区该类乡镇共有4个,沿泰顺县边界呈点状分布,整体活力较高,其乡村空间活力均值(0.262)仅次于强综合型乡(镇) (0.352),根据乡(镇)的主导活力又划分为生活主导型(1个)、生态主导型(1个)和生产主导型(2个)。对于这类乡镇,应发挥自身优势活力整合协调其他活力,带动乡村全面协调可持续发展。①生活主导型。该类乡(镇)仅有泗溪镇。泗溪镇位于泰顺县东南部,是浙南闽北的重要交通枢纽,各类乡村空间活力,尤其是乡村生活空间活力水平均较高。随着泗溪镇15 min品质文化生活圈、残疾人之家工程、泰顺县人民医院泗溪分院迁建工程等社会事业的推进建设和各类配套设施的完善,民生福祉不断增进。未来,面对苍泰高速带来的发展机遇,应积极拓展发展空间,搭建重大产业平台,加快镇域高质量发展;在此基础上以“美好生活”为目标,进一步推进人居环境政治和文明创建工作,让居民的生活品质更进一步。②生态主导型。该类乡(镇)仅有司前畲族镇,位于泰顺县北部,境内竹木、水力资源丰富,生态环境较好,植被覆盖率达89.06% (研究区平均为80.66%),地均生态服务价值为3.39万元·hm−2(研究区平均为3.10万元·hm−2)。司前畲族镇利用自身生态优势,以乌岩岭景区和白鹤山庄为依托,融合畲族文化、森林、乌饭等元素,形成了乡村特色民宿、竹林康养、畲族风情体验等业态,带动了乡村空间活力协调发展。未来要进一步保障生态空间规模,提升生态资源质量,把生态产业作为增强乡村空间活力的核心,建设特色的产业体系,打造绿色富民“金钥匙”。③生产主导型。该类乡(镇)有2个,包括龟湖镇和雅阳镇,位于泰顺县南部,是重要省际边贸地区,区域优势突出,人均耕地面积为0.06 hm2·人−1(研究区平均为0.05 hm2·人−1),村均集体经济和工业企业数均较高,生产空间活力远远超过其他乡(镇)。近年来,依靠叶蜡石、有机茶等特色资源与旅游产业的融合,积极推动文化产业园、全域景区化等提升项目,快速带动了该区域的乡村产业空间活力。未来应坚持把特色资源作为核心,谋划布局“一镇一品牌、一村一产业” ,建成现代农业园区,通过改造低效旱地,重点发展特色经济作物种植,并发挥龙头企业带头作用,持续打响特色农业品牌,适当拉长产业链,进一步激发乡村产业空间活力。
-
极化发展型乡(镇)是指存在主导活力的同时也存在短板活力的乡(镇)。研究区这类乡(镇)共有3个,分布集中连片,位于泰顺县北部,与生态主导型乡(镇)司前畲族镇和强综合性乡(镇)紧密相连,其生态空间活力均较高(均值为0.086,在四大类中分值最高),但因受其他短板活力的影响,整体乡村空间活力较低。因此根据这类乡(镇)的短板活力将其划分为生态主导-生活滞后型和生态主导-生产滞后型。对于这类乡(镇)应利用自身优势资源,进一步提升主导活力。在此基础上应积极探索特色发展道路,补齐短板活力,实现各类乡村空间活力协同发展。①生态主导-生活滞后型。这类乡(镇)共2个,为筱村镇和包垟乡,具有丰富的自然、人文资源,周边旅游资源丰富,拥有丰富的历史古村古民居和名木古树。目前,这类乡(镇)主要发展生态果林无公害蔬菜、规模养殖和绿化苗。但公共服务和基础设施建设水平较低,对居民文化生活的关注略显不足。未来,应重点发展社会事业,加快改善基础设施,持续提升公共服务水平,促进各类乡村活力协同发展。此外,应利用生态优势挖掘古民居文化、非遗文化等,丰富和创新相关旅游业态,发展乡村文化旅游;以“一户一策一干部”帮促低收入农户,对产业、就业、安居、医疗、就学等多方面实施帮扶;大力发展来料加工业,实现家门口就业,全面促进农民增收,提升生活品质。②生态主导-生产滞后型。这类乡(镇)仅有百丈镇,位于泰顺县北部飞云湖南岸,拥有7 667 hm2生态公益林,生态环境优越,农业主要以种植杨梅Myrica rubra、茶Camellia sinensis为主,但农民收入较低,产业发展存在一定的薄弱环节。未来应进一步强化生态空间涵养能力,提高生态系统服务效益,并积极创新生态补偿及财政转移支付等政策措施,在不损害生态环境的基础上,积极发展生态旅游业;不断探索农体旅融合发展,引导农企合作,深入挖掘培育飞云湖茶、杨梅等产业品牌,开展标准化生产、品牌化营销,有效提高农产品品牌附加值。
-
欠缺发展型乡(镇)是指没有主导活力且存在短板活力的乡(镇)。这些乡(镇)虽然具备一定的资源,但因其缺乏正确的发展定位与产业布局,乡村空间活力难以提升。研究区这类乡(镇)共8个,是整个泰顺县占比最高的活力发展类型,集中分布于泰顺县南部。根据短板活力类型将其划分为生活滞后型、生态滞后型和生产滞后型。对于这类乡(镇)应首先立足于全县长远目标,深入挖掘该类乡(镇)潜在的文化、生态等核心资源,明确发展定位,精准打造一批“金名片”,助力乡村振兴发展。①生活滞后型:这类乡(镇)只有彭溪镇,彭溪镇是泰顺县东部中心镇,作为县域东大门入口,工业企业个数仅次于罗阳镇,工业较为发达,但生活配套设施建设较为落后。镇区应以提升完善公共服务配套和基础设施配套为主,并通过文化节庆和有机特色农产品丰富旅游休闲度假,提升乡村空间活力的整体协调;还需加强农村思想道德和公共文化建设,改善农民精神风貌,提高乡村社会文明程度,焕发乡村文明新气象。②生态滞后型。这类乡(镇)共有2个,为柳峰乡和仕阳镇,位于泰顺县南部,与福建省接壤,植被覆盖率仅为71.41% (研究区平均为80.66%),生态环境较差,乡村生态空间活力整体居于末位。未来应立足绿色发展,推进生物多样性保护和可持续利用,积极完善森林生态效益补偿机制,大力推进生产方式绿色低碳循环化改造,健全生态综合保护利用机制;在此基础上,加快推进旅游保护开发,依托仕阳梯田、土楼等特色资源,构建全域旅游新格局。③生产滞后型。这类乡(镇)共5个,包括南浦溪镇、三魁镇、西旸镇、凤垟乡和东溪乡,境内森林覆盖率高,污染少,生态环境较好,旅游资源丰富,但工业和休闲旅游业发展均较弱,人均耕地面积也低于平均水平。未来应特色发展山区生态农业,加大粮食功能区和高标准农田建设支持力度,培育壮大南浦溪果酒、东溪茶文化等特色产业,并植入农业观光、采摘等低成本体验型旅游业态,带动当地村民就地就业,提高收入水平,推动乡村生产空间活力提升。
-
本研究基于乡村空间活力内涵及理论框架的解析,从乡村生活空间活力、生态空间活力、生产空间活力3个方面构建乡村空间活力指数评价体系,测度了浙江省山区县泰顺县19个乡(镇)的乡村空间活力水平,开展了空间特征分析,并通过识别各乡(镇)主导活力和短板活力,探索了各类型区差异化提升路径,以主导活力和短板活力为原则划分活力发展类型,分别提出相应的提升策略。首先,从整体上看,泰顺县乡村空间活力总体呈“局部高值,北高南低”的空间分布特征,大部分乡(镇)的乡村生态空间活力、生产空间活力和生活空间活力均较低。其次,各项子系统活力不尽相同,乡村生活空间活力总体呈现“局部低值,西高东低”的空间分布特征;乡村生态空间活力总体呈“北高南低”的空间分布特征;乡村生产空间活力总体呈“局部突出,四周高中间低”的空间分布特征。最后,将泰顺县各乡(镇)划分为综合发展型、主导发展型、极化发展型和欠缺发展型四大类,并根据不同地域类型提出活力提升策略,旨在为加快推进山区县域乡村振兴与共同富裕提供决策参考,在一定程度上也完善了乡村空间活力理论与评价研究。
泰顺县是浙江省共同富裕示范区的加快发展县——山区26县的重点区域。与国内相关研究对比,泰顺县具有较强的典型性,对其进行实证研究与时空特征分析,既能很好地明确共同富裕示范区的乡村空间活力特征与差异,在比较优势的基础上理性揭示乡村发展的区域异质性,研判乡村发展阶段与问题,进而开展有针对性的政策设计。同时以县域为研究尺度,有利于科学划分乡村活力发展类型,从而对各个乡(镇)精准施策。
本研究仍存在不足:①乡村空间活力自提出以来便是以政策为导向,发达国家的实践探索也证明了其有效性。近年来,在中国的区域背景下,围绕乡村发展与乡村活力相继提出了美丽乡村建设、乡村振兴、共同富裕等概念,应进一步完善乡村空间活力理论体系,将乡村活力与中国乡村发展政策内涵有效衔接,确保乡村活力理论体系的系统性、包容性与发展性,积极为政策落实与指导做出贡献。②由于数据获取等问题,只选取了泰顺县进行分析,在其他区域的实用性可能会受到一定的限制,难以涵盖广大山区县域特征的多样性。下一步可从全国范围选取不同地域类型的山区县域进行横向对比分析,提出更具有针对性的乡村发展策略建议。此外,所采用的研究方法、研究思路及所建立的评价指标体系也有待进一步完善。③乡村发展有自身的规律,不同的社会经济发展阶段会有不同的活力发展状态,呈现发展、维持或衰退的状态,而在相同的环境下,也会呈现不同的状态,造成这种现象的原因有待进一步研究。通过对不同社会经济形态不同地域空间格局的乡村发展核心影响因素进行量化研究,找到影响乡村发展的关键要素,才能准确研判乡村活力状况,进一步预测或决定乡村未来的发展。
Construction and measurement analysis of evaluation index system for rural spatial vitality in mountainous counties
-
摘要:
目的 在乡村振兴和共同富裕背景下,探索提升不同地域类型乡村空间活力的具体路径,为乡村发展提供科学依据。 方法 以浙江省泰顺县为研究对象,从乡村生活、生态、生产活力3个维度系统构建乡村空间活力指数评价体系,应用综合评价法研究乡村空间活力发展的空间差异,利用纵横对比法识别各乡(镇)主导活力和短板活力,探索各类型区差异化提升路径。 结果 ①泰顺县乡村空间活力空间分异特征明显,总体呈“局部高值,北高南低”,大部分乡(镇)的活力水平比较低;各项子系统活力不尽相同,乡村生活、生态、生产空间活力分别呈现“局部低值,西高东低”“北高南低” “局部突出,四周高中间低”的空间分布特征。②以主导活力和短板活力为划分原则,将泰顺县划分为综合发展型、主导发展型、极化发展型和欠缺发展型四大类,并根据不同地域类型提出活力提升策略:综合发展型应高效利用各自的优势资源,形成强对弱的对点帮扶,联合活化,推进泰顺县乡村空间活力的跨越式提升;主导发展型应发挥自身优势活力整合协调其他活力;极化发展型应在提升主导活力的基础上重点考虑积极探索特色发展道路,补齐短板活力;欠缺发展型应深入挖掘文化、生态等核心资源,明确发展定位。 结论 泰顺县乡村活力水平整体较低,各乡(镇)也有不同的优劣势,体现了活力水平发展的不平衡不充分特征。提出差异化提升路径以期对泰顺县实现乡村振兴发展有一定的指导作用。图2表3参25 Abstract:Objective This study aims to explore specific paths to improve the vitality of different regional types of rural areas in the context of rural revitalization and common prosperity, so as to provide scientific basis for rural development. Method Taking Taishun County in Zhejiang Province as the research object, a rural space vitality index evaluation system was constructed from the dimensions of rural life, ecology, and production vitality. The comprehensive evaluation method was applied to study the spatial differences in the development of rural spatial vitality, and the vertical and horizontal comparison method was used to identify leading and weak vitality of each township (town), and explore differentiated upgrading paths for various types of districts. Result (1) The spatial differentiation characteristics of rural spatial vitality in Taishun County were obvious, displaying an overall value of high in local areas, and high in the north and low in the south. The vitality level of most townships (towns) was relatively low. The vitality of each subsystem was different. The vitality of rural life, ecology and production space respectively showed the spatial distribution characteristics of locally low, high in the west and low in the east, high in the north and low in the south, locally prominent and high in the surroundings and low in the middle. (2) Based on the dividing principles of leading vitality and weak vitality, Taishun County was divided into four categories: comprehensive development, leading development, polarized development and underdevelopment, and the following vitality promotion strategies were proposed according to different regional types. The comprehensive development type should make full use of their respective advantageous resources to offer targeted assistance and form joint activation between the strong and the weak to promote a leap-forward improvement in rural spatial vitality. The leading development type should give full play to its own advantages and vitality to integrate and coordinate other vitalities. The polarized development should focus on exploring distinctive development path and filling gaps in vitality while enhancing the leading vitality. The underdevelopment type should deeply tap into core resources such as culture and ecology, and clarify the development orientation. Conclusion The overall level of rural vitality in Taishun County is low, and each township (town) also has different advantages and disadvantages, reflecting the unbalanced and inadequate development of vitality level. The proposed differentiated upgrading path is expected to have a certain guiding role in achieving rural revitalization and development in Taishun County. [Ch, 2 fig. 3 tab. 26 ref.] -
Key words:
- rural spatial vitality /
- index system /
- common prosperity /
- development strategy
-
图 1 乡村空间活力和各子系统活力的空间分布
Figure 1 The rural spatial vitality and the spatial distribution of the vitality of each subsystem
图 2 泰顺县乡村空间活力发展类型空间分异
Figure 2 2 Spatial differentiation of rural spatial vitality development types in Taishun County
表 1 泰顺县乡村空间活力发展类型划分原则
Table 1. Classification of rural spatial vitality levels in Taishun County
是否存在
主导活力是否存在
短板活力主导活力项数/项 活力发展类型 活力发展亚类 是 否 0<N<3 主导发展型 按主导活力类型划分 是 0<N<3 极化发展型 主导活力-短板活力 否 是 N=0 欠缺发展型 按短板活力类型划分 − − N=0或N=3或n=3 综合发展型 若N=3,则为强综合型;若N=0,则为弱综合型;若n=3,则为协调综合型 说明:N为主导活力项数,n为非主导活力项数;−表示不确定,需根据综合发展类型判断。  下载: 导出CSV
下载: 导出CSV
表 2 乡村空间活力指数评价体系
Table 2. Evaluation system of rural spatial vitality index
目标层 准则层 指标层 性质 指标意义 权重系数 乡村空间活力 生活空间活力
(0.385)美丽乡村精品村占比/% + 精品村数量/行政村总数[来源泰顺县人民政
府网]0.043 文化旅游体育与传媒支出占
总支出比重/%+ 文化旅游体育与传媒支出/政府财政总支出[来
源人民政府部门决算公开]0.065 人均一般公共预算收入/(元·人−1) + 一般公共预算收入/2020年户籍人口 0.073 金融服务水平/家 + 万人拥有金融机构网点数 0.082 医疗服务水平/家 + 万人拥有医疗机构数 0.050 生活服务水平/个 + 万人拥有商店、超市、农贸市场数 0.045 教育服务水平/个 + 万人拥有学校数 0.027 生态空间活力
(0.206)植被覆盖率/% + (林地面积+草地面积)/县域总面积 0.031 3A景区村庄数占比/% + 3A景区村庄数/行政村总数 0.037 地均生态服务价值/(元·hm−2) + 区域生态服务价值总和/镇域面积 0.056 土地开发强度 − 建设用地面积/镇域面积 0.016 休闲农业基础设施保障/万元 + 区域农林水务投入 0.066 生产空间活力
(0.409)人均耕地面积/(hm2·人−1) + 耕地面积/2020年户籍人口 0.048 村均经营性集体经济/(万元·村−1) + 村经营性集体经济总量/行政村总数 0.125 农林水事务支持强度/(万元·hm−2) + 农林水事务支出/耕地面积 0.054 休闲旅游功能/个 + 万人拥有农家乐个数、观光园个数 0.055 工业企业个数/个 + 工业企业的户数 0.126 说明:指标性质相对于目标层而确定,其中+表示正向指标,−表示负向指标。地均生态服务价值通过谢高地等[25]方法计算,2020年户籍人口根据2019年户籍人口预测。
下载: 导出CSV
表 3 泰顺县各乡镇乡村空间活力发展类型统计
Table 3. Statistics of spatial vitality development types of villages and towns in Taishun County
活力发展类型 活力发展亚类 乡(镇) 现状特征 乡镇数量/个 综合发展型 强综合型 竹里畲族乡、罗阳镇 各类活力水平较为均衡,乡村空间活力水平高 2 弱综合型 大安乡、雪溪乡 各类活力水平较为均衡,乡村空间活力水平低 2 主导发展型 生活主导型 泗溪镇 各类公共服务和配套设施完善,民生福祉不断增进 1 生态主导型 司前畲族乡 境内竹木、水力资源丰富,生态环境较好 1 生产主导型 龟湖镇、雅阳镇 区位优势突出,乡村集体经济和工业发展较好 2 极化发展型 生态主导-生活滞后型 筱村镇、包垟乡 具有丰富的自然、人文资源,主要以农业为主,但公共服务和基础设施建设水平较低,对居民文化生活的关注略显不足 2 生态主导-生产滞后型 百丈镇 生态环境优越,以农业生产为主,但农村收入较低,产业发展存在一定问题 1 欠缺发展型 生活滞后型 彭溪镇 生活配套设施建设较为落后 1 生态滞后性 柳峰乡、仕阳镇 生态环境较差,整体乡村生态空间活力也居于末位 2 生产滞后型 南浦溪镇、三魁镇、西旸
镇、凤垟乡、东溪乡工业和休闲旅游业务发展均较弱 5
下载: 导出CSV
-
[1] 龙花楼, 屠爽爽. 论乡村重构[J]. 地理学报, 2017, 72(4): 563 − 576. LONG Hualou, TU Shuangshuang. Rural restructuring: theory, approach and research prospect [J]. Acta Geographica Sinica, 2017, 72(4): 563 − 576. [2] 王勇, 周雪, 李广斌. 苏南不同类型传统村落乡村性评价及特征研究——基于苏州12个传统村落的调查[J]. 地理研究, 2019, 38(6): 1311 − 1321. WANG Yong, ZHOU Xue, LI Guangbin. The rurality appraisal and characteristics of different types of traditional villages in southern Jiangsu Province: based on surveys of 12 traditional villages in Suzhou [J]. Geographical Research, 2019, 38(6): 1311 − 1321. [3] 刘彦随. 中国新时代城乡融合与乡村振兴[J]. 地理学报, 2018, 73(4): 637 − 650. LIU Yansui. Research on the urban-rural integration and rural revitalization in the new era in China [J]. Acta Geographica Sinica, 2018, 73(4): 637 − 650. [4] 李红波, 胡晓亮, 张小林, 等. 乡村空间辨析[J]. 地理科学进展, 2018, 37(5): 591 − 600. LI Hongbo, HU Xiaoliang, ZHANG Xiaolin, et al. On the analysis of rural space [J]. Progress in Geography, 2018, 37(5): 591 − 600. [5] 陈文盛, 范水生, 邱生荣, 等. 福建省乡村发展水平及主导类型划定[J]. 地域研究与开发, 2016, 35(5): 143 − 148. CHEN Wensheng, FAN Shuisheng, QIU Shengrong, et al. Development level and dominant types partition of rural in Fujian Province [J]. Areal Research and Development, 2016, 35(5): 143 − 148. [6] 冯旭, 张湛新, 潘传杨, 等. 人口收缩情境下的乡村活力分析与实践——基于美国、德国、日本、英国的比较研究[J]. 国际城市规划, 2022, 37(3): 42 − 49, 88. FENG Xu, ZHANG Zhanxin, PAN Chuanyang, et al. Analysis and practice of rural vitality under the circumstances of population shrinkage: comparative study based on the United States, Germany, Japan and the United Kingdom [J]. Urban Planning International, 2022, 37(3): 42 − 49, 88. [7] 干靓, 钱玲燕, 杨秀. 乡村内生型发展活力测评——德国巴伐利亚州的实践与启示[J]. 国际城市规划, 2020, 35(5): 23 − 34. GAN Jing, QIAN Lingyan, YANG Xiu. “Vitalitäts-check” for endogenous development of rural communities: practice in Bavaria, Germany and its enlightenments [J]. Urban Planning International, 2020, 35(5): 23 − 34. [8] WOODS M. Rural geography: blurring boundaries and making connections [J]. Progress in Human Geography, 2009, 33(6): 849 − 858. [9] 沈剑波, 王应宽, 朱明, 等. 乡村振兴水平评价指标体系构建及实证[J]. 农业工程学报, 2020, 36(3): 236 − 243. SHEN Jianbo, WANG Yingkuan, ZHU Ming, et al. Evaluation index system and empirical analysis of rural revitalization level [J]. Transactions of the Chinese Society of Agricultural Engineering, 2020, 36(3): 236 − 243. [10] 何杰, 金晓斌, 梁鑫源, 等. 城乡融合背景下淮海经济区乡村发展潜力——以苏北地区为例[J]. 自然资源学报, 2020, 35(8): 1940 − 1957. HE Jie, JIN Xiaobin, LIANG Xinyuan, et al. Research of rural system development potential in Huaihai Economic Zone: a case study of Northern Jiangsu Province under the background of urban-rural integration [J]. Journal of Natural Resources, 2020, 35(8): 1940 − 1957. [11] 龙花楼, 刘彦随, 邹健. 中国东部沿海地区乡村发展类型及其乡村性评价[J]. 地理学报, 2009, 64(4): 426 − 434. LONG Hualou, LIU Yansui, ZOU Jian. Assessment of rural development types and their rurality in eastern coastal China [J]. Acta Geographica Sinica, 2009, 64(4): 426 − 434. [12] 吕兆群, 王方民, 杨馗, 等. 基于村级尺度的重庆荣昌区村庄发展潜能评价与类型划分[J]. 中国农业资源与区划, 2022, 43(4): 248 − 258. LÜ Zhaoqun, WANG Fangmin, YANG Kui, et al. Village development potential assessment and classification from the village level in Rongchang District, Chongqing Municipality [J]. Chinese Journal of Agricultural Resources and Regional Planning, 2022, 43(4): 248 − 258. [13] 孙玉, 程叶青, 张平宇. 东北地区乡村性评价及时空分异[J]. 地理研究, 2015, 34(10): 1864 − 1874. SUN Yu, CHENG Yeqing, ZHANG Pingyu. Spatio-temporal dynamics of rurality in northeast China [J]. Geographical Research, 2015, 34(10): 1864 − 1874. [14] 李智, 张小林, 李红波, 等. 基于村域尺度的乡村性评价及乡村发展模式研究——以江苏省金坛市为例[J]. 地理科学, 2017, 37(8): 1194 − 1202. LI Zhi, ZHANG Xiaolin, LI Hongbo, et al. Research on rurality at village scale and rural development model: a case of Jintan City, Jiangsu Province [J]. Scientia Geographica Sinica, 2017, 37(8): 1194 − 1202. [15] 熊鹰, 黄利华, 邹芳, 等. 基于县域尺度乡村地域多功能空间分异特征及类型划分——以湖南省为例[J]. 经济地理, 2021, 41(6): 162 − 170. XIONG Ying, HUANG Lihua, ZOU Fang, et al. Multifunctional spatial characteristics of rural areas and their type identification based on county scale: a case of Hunan Province [J]. Economic Geography, 2021, 41(6): 162 − 170. [16] PHILLIPS M. Baroque rurality in an English village [J]. Journal of Rural Studies, 2014, 33: 56 − 70. [17] HALFACREE K H. Talking about rurality: social representations of the rural as expressed by residents of six English parishes [J]. Journal of Rural Studies, 1995, 11(1): 1 − 20. [18] DUENCKMANN F. The village in the mind: applying Q-methodology to re-constructing constructions of rurality [J]. Journal of Rural Studies, 2010, 26(3): 284 − 295. [19] 田超, 程琳琳, 殷婷婷. 多功能视角下县域乡村发展水平评价与类型划分研究[J]. 江西农业大学学报, 2020, 42(4): 829 − 838. TIAN Chao, CHENG Linlin, YIN Tingting. Evaluation of development level and type division of country rural areas from multi-functional perspective [J]. Acta Agriculturae Universitatis Jiangxiensis, 2020, 42(4): 829 − 838. [20] 何焱洲, 王成. 乡村生产空间系统功能评价与格局优化——以重庆市巴南区为例[J]. 经济地理, 2019, 39(3): 162 − 171. HE Yanzhou, WANG Cheng. Evaluation and pattern optimization of rural production space system function: a case study of Ba’nan District, Chongqing Municipality [J]. Economic Geography, 2019, 39(3): 162 − 171. [21] 张利国, 王占岐, 魏超, 等. 基于村域多功能视角的乡村振兴策略——以鄂西郧阳山区为例[J]. 资源科学, 2019, 41(9): 1703 − 1713. ZHANG Liguo, WANG Zhanqi, WEI Chao, et al. Rural revitalization strategy based on the perspective of rural multifunctions: a case of the mountainous areas in Yunyang District of Shiyan City, western Hubei Province [J]. Resources Science, 2019, 41(9): 1703 − 1713. [22] 刘军. 城市人才活力评价与实证研究——以深圳为例[J]. 中国人力资源开发, 2006(1): 4 − 9. LIU Jun. Evaluation and empirical research on the vitality of urban talents: taking Shenzhen as an example [J]. Human Resources Development of China, 2006(1): 4 − 9. [23] 童明. 城市肌理如何激发城市活力[J]. 城市规划学刊, 2014(3): 85 − 96. TONG Ming. How urban fabric can help sustain the vitality of cities [J]. Urban Planning Forum, 2014(3): 85 − 96. [24] 孙璐, 王江萍. 空间规划视域下武汉市乡村“三生空间”分类与功能评价研究[J]. 现代城市研究, 2020(7): 9 − 16. SUN Lu, WANG Jiangping. Research on classification and functional evaluation of rural production-living-ecological space in Wuhan from a spatial planning perspective [J]. Modern Urban Research, 2020(7): 9 − 16. [25] 谢高地, 张彩霞, 张雷明, 等. 基于单位面积价值当量因子的生态系统服务价值化方法改进[J]. 自然资源学报, 2015, 30(8): 1243 − 1254. XIE Gaodi, ZHANG Caixia, ZHANG Leiming, et al. Improvement of the evaluation method for ecosystem service value based on per unit area [J]. Journal of Natural Resources, 2015, 30(8): 1243 − 1254. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230478
 点击查看大图
点击查看大图
计量
- 文章访问数: 1379
- HTML全文浏览量: 368
- PDF下载量: 29
- 被引次数: 0
