




-
香榧Torreya grandis ‘Merrillii’是红豆杉科Taxaceae榧属Torreya榧树Torreya grandis中的优良栽培类型,是中国南方特色干果树种,属于国家二级保护植物[1]。其坚果风味香酥、营养丰富,富含油脂、蛋白质、各种微量元素等,尤其富含多不饱和脂肪酸和生育酚、角鲨烯等多种生物活性成分,具有很高的保健、药用和综合开发利用价值[2−7]。近年来,香榧产业不断发展扩大,目前,香榧种植区域覆盖浙江、安徽、江西、贵州等全国11个省,面积达到10余万hm2[8−9]。然而,在长期栽培管理过程中发现,香榧幼龄期容易出现树体徒长、生长过旺的问题,导致树冠光照强度降低,抑制了叶片光合能力,这不仅影响了树体的结实情况,同时严重影响种实品质,极大地限制了香榧产业的发展[9]。
树形调整是影响果实产量品质的关键整形技术,主要指对果树骨干枝数目和开张角度进行调整,并辅以适当的修剪、环剥等手段,从而影响树体骨架结构、枝条空间分布等,最终影响树冠光照强度、树体通风透光能力、叶片光合能力、营养运输等[10]。研究发现:在诸多影响果实品质的因素中,果树树形结构直接影响果实色泽、口感与营养价值[11]。荔枝Litchi chinensis树形缺乏科学管理,枝条分布密集,使得叶片之间相互遮挡,导致树冠内膛郁闭,降低了叶片对光能的利用,影响了光合同化物的积累和分配,导致果实品质降低[12]。苹果Malus pumila果实品质受树冠光照影响,无效光区越大,果实品质越差,对树体结构进行优化后,果实内光合产物分配和积累更多,产量和品质明显提高[13]。梨Pyrus spp.骨干枝数目调整后,平均单果质量、可溶性固形物、可溶性总糖等含量均明显上升,其中棚架形树冠可溶性固形物含量最高,果实品质一致性最好[14]。在早实核桃Juglans regia树形调整中也发现:与自然圆头形相比,骨干分层形和开心形树冠内通风透光性好,种实粗脂肪,蛋白质等含量较高,坚果品质最好[15]。澳洲坚果Macadamia spp.多干开心形和疏散分层形光合性能最佳、坚果品质最好,尤其是多干开心形对脂肪酸含量的提高效果最显著[16]。此外,研究表明拉枝可以打破树体原有的养分平衡关系,调节树体营养物质运输,使光合产物更多供给果实发育,在果实品质形成方面具有决定性作用。研究发现:开张角度对板栗Castanea mollissima树冠内膛光照强度和果实营养物质积累有显著影响,当开张角度大于60°时,果实内的可溶性糖和淀粉含量达到最高[17]。同样,开张角度60°处理后,枣Ziziphus jujuba果中的可溶性糖和维生素C含量升高,果实品质明显提高[18]。随着开张角度增大,泡核桃Juglans sigillata坚果品质明显提升,尤其是显著增加了种实蛋白质和粗脂肪含量[19]。综上所述,科学合理的树形调整不仅有利于果树树形培养,同时可显著提高树体光照强度和光合能力,促进果实营养物质积累,提高果实品质。近年来,有关树形调整技术在核桃等坚果类树种中虽有少量报道,但并未得到广泛的推广应用。目前有关香榧的优质丰产栽培理论与配套树形调控技术仍不完善,关于树形调整如何影响其树冠光照及果实品质的研究仍未见报道。因此,本研究针对香榧树形调整展开研究,分别设置不同的骨干枝数目和骨干枝开张角度处理,分析其对树体光照强度、光合能力、种实光合碳同化物质量分数、油脂品质等的影响,以揭示树形调整对香榧树冠光照和果实品质的影响,探索香榧树体适宜的骨干枝数目和开张角度,为香榧优质丰产、提高品质提供科学依据。
-
研究区位于浙江省杭州市临安区浙江农林大学潘母岗试验基地(30°14′N,119°42′E)。该区属于亚热带季风气候,全年平均气温为17.8 ℃,平均年降水量为1 454.0 mm,平均相对湿度为70.3%,年日照时数为1 765.0 h。土壤全氮质量分数为1.2 g·kg−1,全磷质量分数为1.1 g·kg−1,全钾质量分数为15.7 g·kg−1。基地采用统一的常规施肥管理,每年施用2 ~ 3次复合肥,每年每株共计750 g,每2年施用1次有机肥。
-
材料为2007年种植的香榧树,树龄为20 a,平均地径为12.2 cm,平均树高为1.8 m。2021年11月选择长势一致的香榧树进行不同树形处理,包括骨干枝数目和骨干枝开张角度的单因素和交互试验。在前期预试验的基础上,骨干枝数目设置单骨干枝自然圆头形(对照,ck)、3条骨干枝开心形(N3)和4条骨干枝开心形(N4)处理;骨干枝开张角度设置30°(对照,A30°)、60°(A60°)和80°(A80°)处理,处理时先将骨干枝简单揉压,用拉枝绳缓慢拉动枝条基部,调节至合适角度。采用随机区组设计,每组9株香榧树,每个处理重复5次,共计45株。
-
沿树冠垂直方向由上至下分为上层、中层、下层,每层间隔50 cm,使用照度计(TES-1339R) 测定不同处理下的各层光照强度。采用便携式光合仪(Li-6400)测定叶片净光合速率,测量时选择天气晴朗且无云的上午9:00—11:00。光合有效辐射(PAR)设定为1 200 μmol·m−2·s−1,空气流速为400 µmol·s−1,叶室温度为25 ℃,具体参照刘琏等[20]的方法。
-
可溶性糖和淀粉质量分数测定采用蒽酮比色法,可溶性蛋白质量分数测定采用考马斯亮蓝染色法,具体参照成豪等[21]的方法。
-
含油率的提取参照GB/T 14772—2008《食品中粗脂肪的测定》索氏抽提法[22]。脂肪酸组分分析采用气相色谱仪(Trace 1300),具体参照ZHOU等[23]的方法。
-
数据用3次重复的平均值±标准误表示。采用SPSS 25对数据进行单因素分析,采用Excel 2013绘制图表。
-
本研究调查了不同树形处理下,香榧树冠5—9月的光照强度变化(表1~2)。骨干枝开张角度单因素分析发现:60°处理显著增加了5—9月香榧树冠中层和下层的光照强度,但80°处理会降低树冠光照强度。骨干枝数目和开张角度的交互处理显著提高了5—7月香榧树冠光照强度,其中,N3-A60°和N4-A60°效果最好(P<0.05)。
表 1 树形调整对香榧中层光照强度变化的影响
Table 1. Effect of tree shape adjustment on the change of central layer light intensity
处理 光照强度/klx 5月 6月 7月 8月 9月 ck-A30° 16.92±0.26 c 18.11±0.21 d 32.26±0.57 b 38.48±0.34 a 22.10±0.42 bc ck-A60° 20.28±0.27 b 23.00±0.24 a 37.59±0.37 a 39.85±0.86 a 26.09±0.21 a ck-A80° 15.03±0.17 d 19.97±0.48 c 26.99±0.62 c 36.37±0.69 b 21.66±0.31 cd N3-A30° 20.08±0.35 b 21.11±1.53 bc 27.90±1.16 c 35.55±0.27 bc 22.86±0.44 b N3-A60° 26.85±0.50 a 22.15±0.35 ab 36.72±0.88 a 39.67±0.67 a 25.96±0.22 a N3-A80° 17.61±0.26 c 18.14±0.18 d 31.09±1.08 b 33.88±0.32 c 21.01±0.27 d N4-A30° 16.19±0.27 d 18.11±0.52 d 38.01±0.45 a 34.58±0.61 c 22.09±0.23 bc N4-A60° 25.25±0.72 a 23.33±0.30 a 36.95±0.66 a 39.15±0.36 a 25.83±0.21 a N4-A80° 18.01±0.59 c 14.96±0.27 e 36.10±0.81 ab 33.93±0.43 c 22.08±0.33 bc NE 0.000 0.000 0.000 0.000 0.032 AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.000 ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 表 2 树形调整对香榧下层光照强度变化的影响
Table 2. Effect of tree shape adjustment on the change of lower layer light intensity
处理 光照强度/klx 5月 6月 7月 8月 9月 ck-A30° 8.55±0.09 c 11.22±0.32 b 11.75±0.49 cd 14.25±0.43 b 10.15±0.33 cde ck-A60° 9.22±0.23 c 14.18±0.45 a 15.78±1.12 ab 16.55±0.22 a 11.73±0.14 a ck-A80° 7.38±0.46 d 9.83±0.68 c 9.93±1.17 d 12.53±0.35 c 9.64±0.16 de N3-A30° 7.41±0.09 d 9.50±0.43 c 10.64±0.69 d 12.60±0.46 c 10.31±0.19 c N3-A60° 13.93±0.45 a 13.82±0.37 a 13.70±0.45 bc 16.08±0.40 a 11.25±0.22 ab N3-A80° 9.15±0.27 c 8.41±0.12 c 12.20±0.79 cd 12.77±0.37 c 9.56±0.14 e N4-A30° 7.38±0.23 d 10.03±0.36 c 12.10±0.57 cd 13.22±0.36 c 10.34±0.10 c N4-A60° 11.54±0.23 b 13.54±0.33 ab 16.55±0.36 a 16.00±0.27 a 10.72±0.11 bc N4-A80° 7.32±0.17 d 6.65±0.13 d 10.21±0.37 d 12.67±0.62 c 10.22±0.22 cd NE 0.000 0.000 0.004 0.111 ns AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.031 ns 0.007 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 基于光照强度的结果,进一步以3条骨干枝为代表,于光照条件好的5、7和9月测定并分析骨干枝数目和开张角度交互处理对香榧叶片光合能力的影响。结果表明:骨干枝数目和开张角度的交互处理可有效提高5、7和9月香榧叶片净光合速率、气孔导度、蒸腾速率(图1)。其中,N3-A60°处理效果最好,分别使7和9月香榧叶片的净光合速率提高了46%和38% (P<0.05),气孔导度提高了35%和54%,蒸腾速率提高了34%和30%。
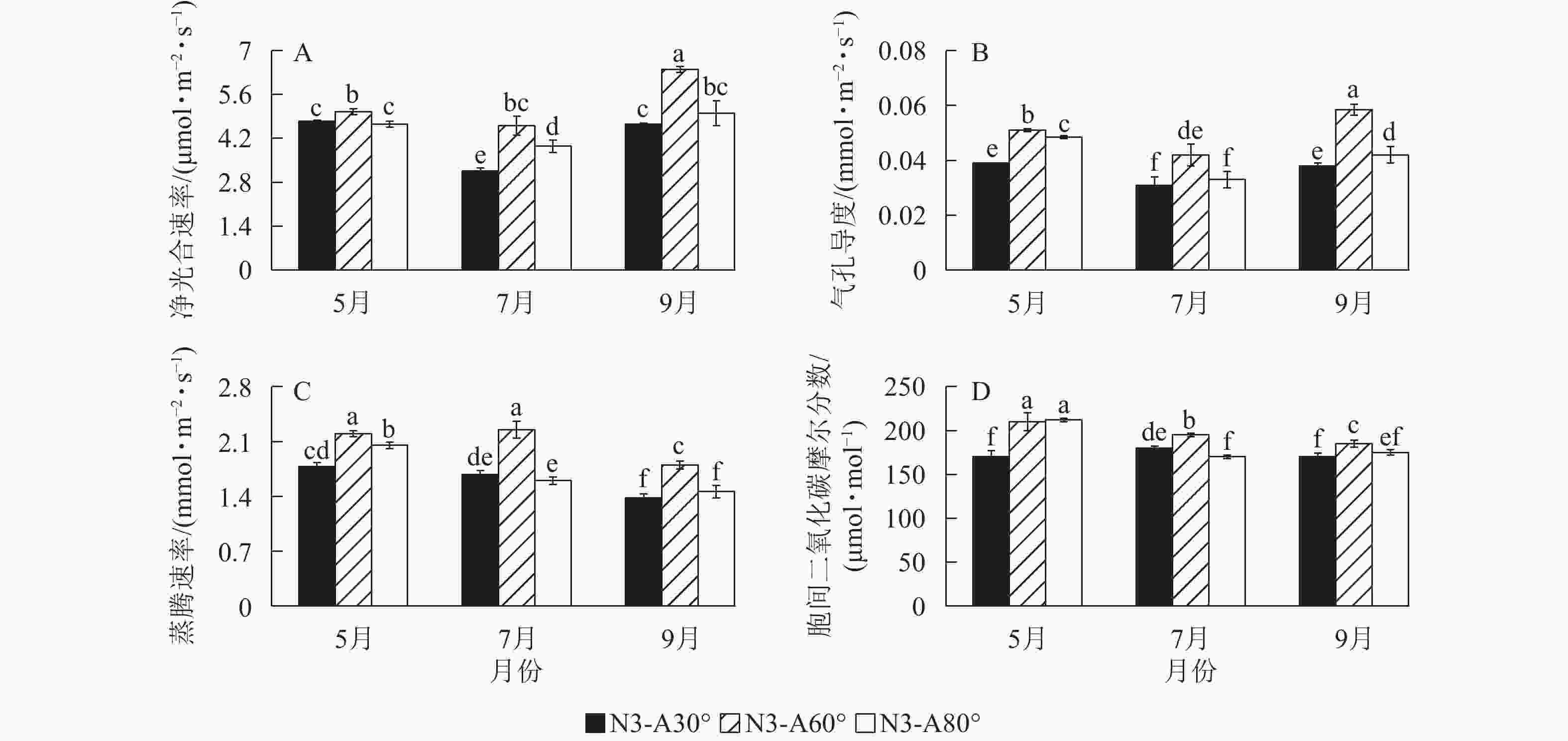
图 1 不同树形处理下香榧叶片光合变化
Figure 1. Effect of tree shape adjustment on the change of leaf photosynthesis
-
由表3~4可见:5—9月香榧种实逐渐成熟,种实内部积累的糖、淀粉等营养物质逐渐转化为蛋白质和油脂,种实中的淀粉和可溶性糖总体呈逐渐下降趋势。与A30°相比,A60°和A80°处理下5—9月香榧种实中可溶性糖质量分数均显著增加(P<0.05)。ck-A60°和ck-A80°处理下5月种实中可溶性糖质量分数分别达到565.11和577.53 mg·g−1,与ck-A30°相比分别高出34%和37%。与A30°相比,A60°处理也显著增加了香榧种实中淀粉质量分数(P<0.05)。相较于单因素试验,骨干枝数目和开张角度的交互处理对5—7月种实中可溶性糖质量分数增加幅度更大,N3-A60°和N4-A60°处理下,5月种实可溶性糖和淀粉质量分数分别达到573.41和576.63 mg·g−1。
表 3 树形调整对香榧种实可溶性糖质量分数的影响
Table 3. Effect of tree shape adjustment on the content of soluble sugar of the seeds
处理 可溶性糖质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 421.32±4.11 e 487.16±6.23 d 338.34±0.36 d 190.71±4.46 cd 158.85±0.67 c ck-A60° 565.11±8.55 c 541.84±10.31 bc 416.54±2.76 a 228.51±6.35 a 191.20±3.63 a ck-A80° 577.53±3.57 bc 512.94±10.18 cd 387.12±6.36 bc 215.29±2.45 ab 178.70±1.22 b N3-A30° 520.20±14.86 d 479.39±13.79 d 346.86±2.55 d 202.64±3.27 bcd 171.07±2.16 c N3-A60° 590.90±5.07 bc 566.07±15.20 ab 378.60±6.24 c 210.84±1.61 abc 190.53±1.40 a N3-A80° 641.06±8.71 a 585.95±2.65 a 367.76±6.82 cd 198.62±4.46 bcd 178.51±1.99 b N4-A30° 522.59±9.70 d 521.33±17.14 cd 354.34±2.69 d 183.24±1.08 d 166.30±2.92 c N4-A60° 609.05±8.22 b 567.31±2.17 ab 414.99±7.73 a 210.98±0.71 abc 185.28±0.99 ab N4-A80° 566.06±10.40 c 546.50±8.53 bc 408.28±6.78 ab 203.51±10.23 bc 180.70±2.30 ab NE 0.000 0.015 0.000 ns ns AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.038 0.017 ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 表 4 树形调整对香榧种实淀粉质量分数的影响
Table 4. Effect of tree shape adjustment on the content of starch of the seeds
处理 淀粉质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 521.82±5.06 bc 267.04±16.07 f 110.38±4.14 d 96.55±5.21 d 107.33±1.39 c ck-A60° 650.25±38.93 a 362.11±2.06 cd 141.85±7.26 bc 117.53±8.18 bc 123.31±2.66 a ck-A80° 519.67±7.61 bc 497.72±7.55 a 166.86±9.52 ab 118.15±1.39 bc 104.11±0.37 c N3-A30° 490.65±10.37 c 311.77±8.97 ef 131.72±7.93 cd 115.05±4.91 c 116.52±1.88 b N3-A60° 573.41±12.95 b 393.56±4.00 bc 164.49±3.53 ab 141.76±1.17 a 110.77±0.72 bc N3-A80° 494.41±9.80 c 369.79±17.69 cd 117.27±4.69 cd 113.43±1.68 c 107.72±2.23 c N4-A30° 483.12±1.16 c 323.66±17.75 de 142.93±10.24 bc 118.15±2.31 bc 81.75±1.17 e N4-A60° 576.63±11.28 b 481.64±20.17 a 180.87±4.97 a 130.92±0.99 ab 106.09±1.71 c N4-A80° 508.92±13.08 c 433.41±1.51 b 121.59±5.50 cd 120.08±0.58 bc 95.36±1.77 d NE 0.021 0.001 ns 0.005 0.000 AE 0.000 0.000 0.000 0.000 0.000 NE × AE ns 0.000 0.000 0.038 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 -
由表5可见:随着香榧种实成熟,种实可溶性蛋白逐渐积累,8月达到最高值,9月相比8月略有下降。骨干枝开张角度单因素处理对香榧种实可溶性蛋白积累有显著影响(P<0.05),其中A60°处理可显著提高种实可溶性蛋白质量分数,但是开张角度继续增大则不利于种实中可溶性蛋白的增加。ck-A80°处理下,7—9月种实可溶性蛋白质量分数较ck-A30°显著降低(P<0.05)。同时,骨干枝数目增加也显著提高了种实中可溶性蛋白的质量分数,N3-A30°和N4-A30°处理下,种实中可溶性蛋白质量分数分别达到484.14和460.3 mg·g−1。骨干枝数目和开张角度的交互处理也可显著提高种实中可溶性蛋白的质量分数,以60°开张角度与3和4条骨干枝的交互处理促进效果最佳,N3-A60°和N4-A60°处理下,8月种实可溶性蛋白质量分数分别达到647.12和752.61 mg·g−1,较ck-A60°分别提高了13%和31%。
表 5 树形调整对香榧种实可溶性蛋白质量分数的影响
Table 5. Effect of tree shape adjustment on the content of soluble protein of the seeds
处理 可溶性蛋白质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 135.89±6.72 d 146.02±5.22 e 172.30±3.13 fg 402.95±14.91 f 343.21±10.93 d ck-A60° 198.53±10.20 b 217.51±3.28 c 270.21±14.30 c 573.63±3.96 c 573.37±6.35 b ck-A80° 140.62±9.01 cd 151.01±3.82 e 161.66±2.30 g 235.22±5.10 h 235.33±14.75 f N3-A30° 205.62±3.35 b 215.97±10.83 c 240.41±5.43 d 484.14±5.23 d 478.95±5.14 c N3-A60° 286.88±0.64 a 329.74±6.35 b 407.50±11.09 b 647.12±12.21 b 585.41±11.49 b N3-A80° 197.94±8.84 b 200.59±7.38 cd 209.55±4.84 e 270.19±9.19 g 297.18±6.63 e N4-A30° 189.08±2.21 b 200.59±2.57 cd 223.38±2.30 e 456.88±8.05 de 460.3±7.14 c N4-A60° 283.63±8.86 a 363.95±9.76 a 444.75±11.75 a 752.61±7.16 a 680.07±15.65 a N4-A80° 163.67±2.30 c 181.76±2.74 d 195.71±1.51 ef 431.99±13.05 ef 296.71±6.88 e NE 0.000 0.000 0.000 0.000 0.000 AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.027 0.000 0.000 0.000 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 -
含油率是衡量香榧种实品质的重要指标,油脂一般在7月开始积累,因此主要检测各处理7—9月种实的含油率。随着7—9月种实逐渐成熟,含油率逐渐增加,9月达到最大值。与A30°相比,A60°处理下种实含油率显著增加(P<0.05),但开张角度过大(A80°)不利于种实含油率积累。单独的骨干枝数目增加对种实含油率无显著影响,交互处理对种实含油率也无显著影响,说明树形调整对种实含油率的影响主要受到骨干枝开张角度的影响(表6)。
表 6 树形调整对香榧种实含油率的影响
Table 6. Effect of tree shape adjustment on the oil content of the seeds
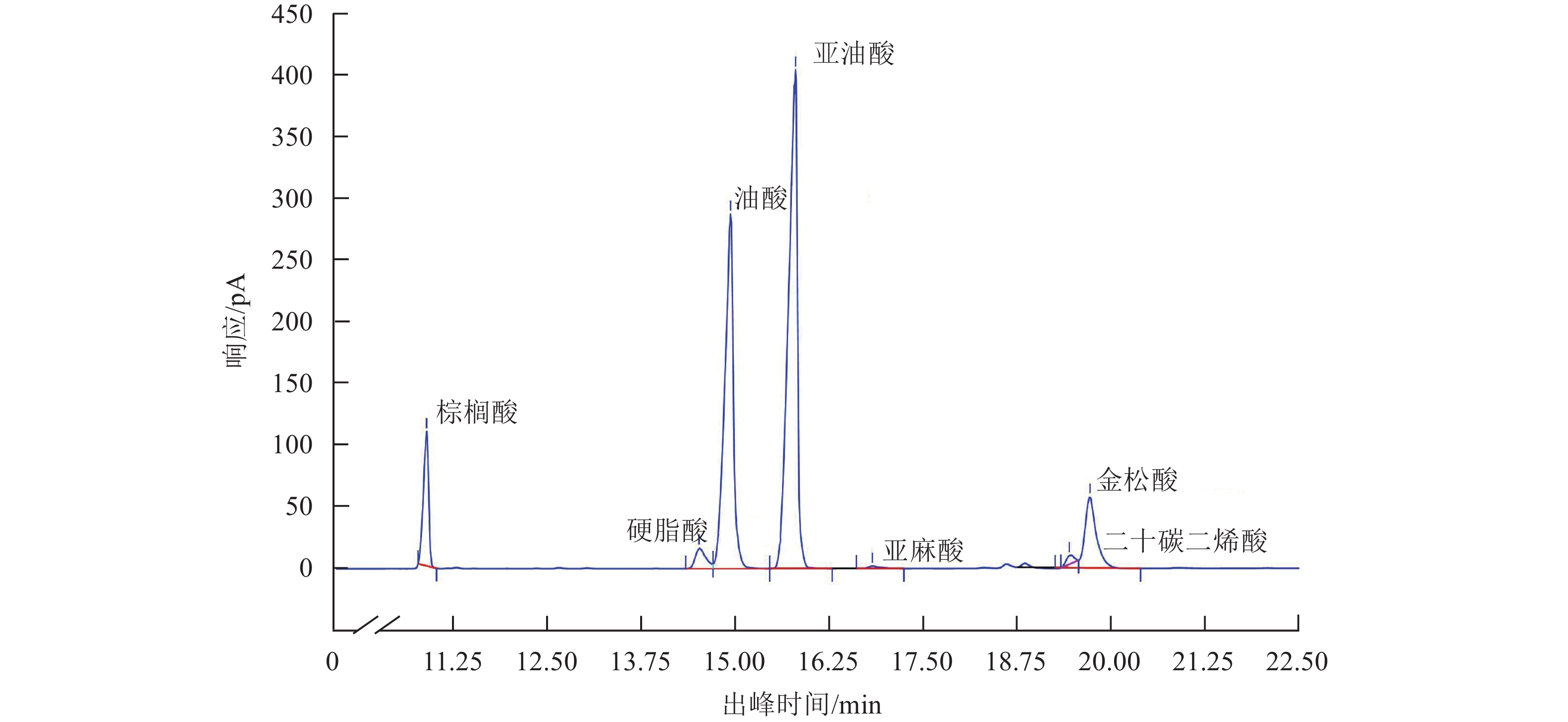
处理 含油率/% 7月 8月 9月 ck-A30° 28.34±0.16 c 45.96±0.01 abc 51.53±0.63 bcd ck-A60° 29.53±0.12 a 46.52±0.13 ab 53.59±0.53 a ck-A80° 28.09±0.32 c 45.11±0.12 c 50.19±0.44 bcd N3-A30° 28.56±0.21 c 46.17±0.23 abc 51.93±0.27 ab N3-A60° 29.65±0.19 a 47.08±0.24 a 53.89±0.69 a N3-A80° 28.12±0.26 c 45.61±0.11 bc 49.50±0.59 cd N4-A30° 28.65±0.17 bc 45.81±0.73 abc 51.26±0.28 bc N4-A60° 29.46±0.34 ab 47.03±0.38 a 53.90±0.48 a N4-A80° 28.34±0.19 c 45.14±0.20 c 49.04±0.30 d NE ns ns ns AE 0.000 0.001 0.000 NE × AE ns ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。 9月香榧成熟种实中脂肪酸组分分析发现:香榧油脂主要由棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸和金松酸等组分组成,其中,不饱和脂肪酸油酸和亚油酸占比最高,金松酸质量分数也较高(图2)。研究发现不同树形处理对脂肪酸组分质量分数变化有显著影响(P<0.05)。如表7所示:骨干枝开张角度增大,各脂肪酸组分变化显著(P<0.05),其中,油酸质量分数和占比逐渐降低,亚油酸和金松酸质量分数和占比均逐渐升高,但开张角度过大,不利于油酸、亚油酸、金松酸的积累。骨干枝数目和开张角度的交互处理同样可以显著提高种实中亚油酸和金松酸的质量分数(P<0.05),其中,以N3-A60°和N4-A60°处理下亚油酸和金松酸质量分数最高,分别达到288.81和65.21 mg·g−1。

图 2 香榧种实主要脂肪酸组分气相色谱图
Figure 2. GC-MS chromatogram of main fatty acids in the seeds
表 7 树形调整对香榧种实脂肪酸组分的影响
Table 7. Effect of tree shape adjustment on the main fatty acid compositions of the seeds
处理 脂肪酸组分/(mg·g−1) 棕榈酸 硬脂酸 油酸 亚油酸 亚麻酸 二十碳二烯酸 金松酸 ck-A30° 39.04±0.23 cd 11.47±0.07 b 164.59±0.95 b 240.73±1.39 f 1.60±0.01 fg 3.64±0.02 a 47.48±0.28 f ck-A60° 44.52±0.21 a 10.50±0.05 c 151.62±0.71 c 262.57±1.22 d 1.82±0.01 d 3.45±0.02 b 54.59±0.26 c ck-A80° 38.96±0.16 cd 9.92±0.04 d 149.08±0.62 d 242.67±1.00 f 1.66±0.01 e 2.83±0.01 e 50.35±0.21 e N3-A30° 40.75±0.10 b 12.36±0.03 a 176.29±0.43 a 231.78±0.57 g 1.61±0.00 f 3.01±0.01 c 46.51±0.11 g N3-A60° 41.01±0.25 b 6.84±0.04 h 132.17±0.80 e 288.81±1.74 a 1.94±0.01 b 1.98±0.01 i 59.40±0.36 b N3-A80° 38.76±0.22 d 7.38±0.04 f 117.32±0.66 f 265.62±1.49 cd 1.89±0.01 c 2.56±0.01 g 55.18±0.31 c N4-A30° 41.03±0.11 b 9.96±0.03 d 150.38±0.39 cd 246.87±0.64 e 1.57±0.00 g 2.92±0.01 d 53.37±0.14 d N4-A60° 44.31±0.19 a 7.98±0.03 e 132.52±0.56 e 277.39±1.16 b 1.87±0.01 c 2.78±0.01 f 65.21±0.38 a N4-A80° 39.36±0.11 c 7.24±0.02 g 104.93±0.30 g 268.38±0.77 c 2.11±0.01 a 2.26±0.01 h 59.88±0.17 b NE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 AE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著。 -
果实品质是衡量果实好坏的重要指标,提高果实品质是增加经济效益的关键,也是果树栽植和整形的出发点和落脚点[24]。研究表明:科学的树形有利于培养良好的树体结构,在提高树冠通风透光能力的同时,促进叶片光合作用[25−26]。研究发现:桃树Prunus persica ‘V’形树形处理下冠层内光照分布最为均匀,净光合速率最高,果实色泽最好,香气含量最高[27−28]。‘砀山酥梨’‘玉露香梨’‘黄冠’等不同品种梨树不同树形处理也发现:Y字形树冠整体光照情况、净光合速率、果实品质均优于其他树形[29]。此外,骨干枝开张角度也是树形调整的重要因素。研究发现:开张角度105°和120°处理提高了‘富士’苹果树冠内膛和中下部叶片的光合能力和果实中可溶性固形物含量[30]。本研究也发现:骨干枝60°开张角度显著增加了5—9月香榧树冠中层和下层的光照强度。同时,3条骨干枝开心形和开张角度60°交互处理显著提高了5、7和9月香榧叶片净光合速率、气孔导度、蒸腾速率。以上研究表明:树形调整对香榧树冠整体光照强度及叶片光合能力有显著影响。
可溶性糖、淀粉等碳水化合物是果实中重要的营养物质,其含量高低也是果实品质形成的重要因素[31−32]。研究表明:合理的树形调整不仅能够提高树冠光照强度和叶片光合能力,同时能够促进糖、淀粉等营养物质在果实中积累,提高果实品质[33]。例如,扇形树形有利于葡萄Vitis vinifera果实中总糖的积累[34]。多主枝平棚形鲜黄梨Pyrus pyrifolia树冠冠层开度和冠下光照显著高于V字形和疏散分层形树形,其透光性最好,光合能力最强,果实中可溶性总糖、葡萄糖、果糖、蔗糖含量最高[35]。不同开张角度处理的‘富士’苹果中可溶性糖质量分数也显著提高,其中,开张角度为110°~120°时达到最高[36−37]。本研究也发现:树形调整有利于香榧种实中营养物质的积累,骨干枝数目和开张角度增加均能显著提高5—7月香榧种实中可溶性糖、淀粉质量分数。二者交互处理效果更好,N4-A60°和N4-A60°处理下,5月种实可溶性糖和淀粉质量分数分别达到609.05和576.63 mg·g−1。以上研究表明:香榧树形调整可显著促进种实中碳水化合物的积累。
核桃、香榧等坚果种实中蛋白质以及油脂质量分数高低是评价其品质的重要指标。在核桃和澳洲坚果研究中发现:多主枝开心形和分层形树形可有效提高坚果品质,增加种实蛋白和脂肪酸质量分数,尤其是显著提高了不饱和脂肪酸油酸和亚油酸的占比[16, 38]。开张角度也显著增加了泡核桃种实蛋白质和粗脂肪质量分数[18]。本研究发现:随着香榧种实逐渐成熟,5—9月种实可溶性糖、淀粉质量分数逐渐降低,蛋白质和油脂质量分数逐渐积累。N3-A60°和N4-A60°处理下,蛋白质和油脂质量分数最高。脂肪酸组分分析也表明:N3-A60°和N4-A60°处理可显著提高香榧种实中不饱和脂肪酸亚油酸和金松酸的质量分数,9月种实成熟时,二者质量分数分别达到288.81和65.21 mg·g−1。以上研究表明:不同树形调整促进了糖、淀粉等碳水化合物在香榧种实中的积累,同时也显著促进了种实中碳水化合物向蛋白质和油脂的转化。因此,树形调整对香榧种实品质形成具有重要作用,生产上可通过构建和培养合理树形来实现香榧的提质增效。
-
树形调整对香榧种实品质有显著的促进作用,其中以3和4条骨干枝与60°开张角度处理最佳。树形调整显著提高了香榧树冠通风透光能力,增加了树冠中层和下层的光照强度,使叶片可以更好地进行光合作用,净叶片光合作用显著增强,显著促进了5—7月香榧种实中可溶性糖、淀粉的积累,显著促进了7—9月种实中可溶性糖、淀粉向蛋白质和油脂的转化,提高了种实中可溶性蛋白质量分数和含油率,同时促进了不饱和脂肪酸亚油酸和金松酸的积累。
Effects of tree shape adjustment on canopy light intensity and seed quality of Torreya grandis ‘Merrillii’
-
摘要:
目的 研究不同树形调整对香榧Torreya grandis ‘Merrillii’树冠光照强度和光合速率的影响,阐明不同骨干枝数目和开张角度处理下香榧种实中碳水化合物、油脂等变化规律,揭示树形调整对香榧种实品质的影响,为香榧树形培养、提质增效提供科学依据。 方法 设置不同骨干枝数目和骨干枝开张角度单因素和交互处理试验,测定树冠光照强度、叶片光合速率,分析可溶性糖、淀粉、可溶性蛋白质量分数、含油率及脂肪酸组分变化。 结果 骨干枝开张角度调整对香榧树冠光照强度有显著影响。与对照相比,60°处理下5—9月香榧树冠中层和下层的光照强度显著增加(P<0.05),7月树冠中层光照强度最高达到40 klx,但80°开张角度则会降低树冠光照强度。同时,骨干枝数目和开张角度的交互处理下,5、7和9月香榧叶片净光合速率、气孔导度、蒸腾速率均显著提高(P<0.05)。碳水化合物检测发现,3和4条骨干枝与60°开张角度(N3-A60°和N4-A60°)处理下香榧种实中可溶性糖和淀粉质量分数显著增加(P<0.05),5月香榧种实可溶性糖和淀粉质量分数分别达到609.05和576.63 mg·g−1。树形调整也显著促进了香榧种实中可溶性蛋白和油脂的积累(P<0.05)。 结论 树形调整对香榧树冠光照强度、叶片光合能力和种实品质有明显的促进作用,其中以3和4条骨干枝与60°开张角度处理最佳。图2表7参38 Abstract:Objective This study aimed to investigate the impact of adjusting the number and opening angle of skeleton branches of Torreya grandis ‘Merrillii’ on canopy light intensity and leaf photosynthesis. Additionally, it aimed to elucidate alterations in carbohydrate, oil content, and other components to reveal the effect of tree shape adjustment on seed quality and provide a scientific basis for the tree cultivation. Method Experiments with different numbers and opening angles of skeleton branches were conducted using single-factor and interactive treatments to examine canopy light intensity, leaf photosynthesis, and the contents of soluble sugar, starch, soluble protein, oil and fatty acid components. Result Adjusting the opening angle of skeleton branches significantly influenced canopy light intensity. Under the opening angle of 60°, the light intensity in the central and lower layers of the canopy notably increased (P<0.05) from May to September, peaking at 40 klx in the central layer in July. However, an excessively wide opening angle of 80° led to a decrease of canopy light intensity. Under the interactive treatment of different numbers and opening angles of skeleton branches, the net photosynthetic rate, stomatal conductance, and transpiration rate of the leaves significantly increased in May, July and September (P<0.05). Carbohydrate analysis revealed that the contents of soluble sugar and starch in seeds under 3 and 4 branches with the opening angle of 60° adjustment (N3-A60° and N4-A60°) were significantly increased (P<0.05). The content of soluble sugar and starch in the seeds reached 609.05 and 576.63 mg·g−1, respectively. Furthermore, tree shape adjustment significantly promoted the accumulation of soluble protein and oil in the seeds. Conclusion The tree shape adjustment of T. grandis ‘Merrillii’ can enhance the canopy light intensity, leaf photosynthesis and seed quality, and the 3 and 4 skeleton branches with the opening angle of 60° has the best promotion effect. [Ch, 2 fig. 7 tab. 38 ref.] -
Key words:
- Torreya grandis ‘Merrillii’ /
- tree shape adjustment /
- light intensity /
- seed quality
-
图 1 不同树形处理下香榧叶片光合变化
Figure 1 Effect of tree shape adjustment on the change of leaf photosynthesis
表 1 树形调整对香榧中层光照强度变化的影响
Table 1. Effect of tree shape adjustment on the change of central layer light intensity
处理 光照强度/klx 5月 6月 7月 8月 9月 ck-A30° 16.92±0.26 c 18.11±0.21 d 32.26±0.57 b 38.48±0.34 a 22.10±0.42 bc ck-A60° 20.28±0.27 b 23.00±0.24 a 37.59±0.37 a 39.85±0.86 a 26.09±0.21 a ck-A80° 15.03±0.17 d 19.97±0.48 c 26.99±0.62 c 36.37±0.69 b 21.66±0.31 cd N3-A30° 20.08±0.35 b 21.11±1.53 bc 27.90±1.16 c 35.55±0.27 bc 22.86±0.44 b N3-A60° 26.85±0.50 a 22.15±0.35 ab 36.72±0.88 a 39.67±0.67 a 25.96±0.22 a N3-A80° 17.61±0.26 c 18.14±0.18 d 31.09±1.08 b 33.88±0.32 c 21.01±0.27 d N4-A30° 16.19±0.27 d 18.11±0.52 d 38.01±0.45 a 34.58±0.61 c 22.09±0.23 bc N4-A60° 25.25±0.72 a 23.33±0.30 a 36.95±0.66 a 39.15±0.36 a 25.83±0.21 a N4-A80° 18.01±0.59 c 14.96±0.27 e 36.10±0.81 ab 33.93±0.43 c 22.08±0.33 bc NE 0.000 0.000 0.000 0.000 0.032 AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.000 ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。  下载: 导出CSV
下载: 导出CSV
表 2 树形调整对香榧下层光照强度变化的影响
Table 2. Effect of tree shape adjustment on the change of lower layer light intensity
处理 光照强度/klx 5月 6月 7月 8月 9月 ck-A30° 8.55±0.09 c 11.22±0.32 b 11.75±0.49 cd 14.25±0.43 b 10.15±0.33 cde ck-A60° 9.22±0.23 c 14.18±0.45 a 15.78±1.12 ab 16.55±0.22 a 11.73±0.14 a ck-A80° 7.38±0.46 d 9.83±0.68 c 9.93±1.17 d 12.53±0.35 c 9.64±0.16 de N3-A30° 7.41±0.09 d 9.50±0.43 c 10.64±0.69 d 12.60±0.46 c 10.31±0.19 c N3-A60° 13.93±0.45 a 13.82±0.37 a 13.70±0.45 bc 16.08±0.40 a 11.25±0.22 ab N3-A80° 9.15±0.27 c 8.41±0.12 c 12.20±0.79 cd 12.77±0.37 c 9.56±0.14 e N4-A30° 7.38±0.23 d 10.03±0.36 c 12.10±0.57 cd 13.22±0.36 c 10.34±0.10 c N4-A60° 11.54±0.23 b 13.54±0.33 ab 16.55±0.36 a 16.00±0.27 a 10.72±0.11 bc N4-A80° 7.32±0.17 d 6.65±0.13 d 10.21±0.37 d 12.67±0.62 c 10.22±0.22 cd NE 0.000 0.000 0.004 0.111 ns AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.031 ns 0.007 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。
下载: 导出CSV
表 3 树形调整对香榧种实可溶性糖质量分数的影响
Table 3. Effect of tree shape adjustment on the content of soluble sugar of the seeds
处理 可溶性糖质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 421.32±4.11 e 487.16±6.23 d 338.34±0.36 d 190.71±4.46 cd 158.85±0.67 c ck-A60° 565.11±8.55 c 541.84±10.31 bc 416.54±2.76 a 228.51±6.35 a 191.20±3.63 a ck-A80° 577.53±3.57 bc 512.94±10.18 cd 387.12±6.36 bc 215.29±2.45 ab 178.70±1.22 b N3-A30° 520.20±14.86 d 479.39±13.79 d 346.86±2.55 d 202.64±3.27 bcd 171.07±2.16 c N3-A60° 590.90±5.07 bc 566.07±15.20 ab 378.60±6.24 c 210.84±1.61 abc 190.53±1.40 a N3-A80° 641.06±8.71 a 585.95±2.65 a 367.76±6.82 cd 198.62±4.46 bcd 178.51±1.99 b N4-A30° 522.59±9.70 d 521.33±17.14 cd 354.34±2.69 d 183.24±1.08 d 166.30±2.92 c N4-A60° 609.05±8.22 b 567.31±2.17 ab 414.99±7.73 a 210.98±0.71 abc 185.28±0.99 ab N4-A80° 566.06±10.40 c 546.50±8.53 bc 408.28±6.78 ab 203.51±10.23 bc 180.70±2.30 ab NE 0.000 0.015 0.000 ns ns AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.038 0.017 ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。
下载: 导出CSV
表 4 树形调整对香榧种实淀粉质量分数的影响
Table 4. Effect of tree shape adjustment on the content of starch of the seeds
处理 淀粉质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 521.82±5.06 bc 267.04±16.07 f 110.38±4.14 d 96.55±5.21 d 107.33±1.39 c ck-A60° 650.25±38.93 a 362.11±2.06 cd 141.85±7.26 bc 117.53±8.18 bc 123.31±2.66 a ck-A80° 519.67±7.61 bc 497.72±7.55 a 166.86±9.52 ab 118.15±1.39 bc 104.11±0.37 c N3-A30° 490.65±10.37 c 311.77±8.97 ef 131.72±7.93 cd 115.05±4.91 c 116.52±1.88 b N3-A60° 573.41±12.95 b 393.56±4.00 bc 164.49±3.53 ab 141.76±1.17 a 110.77±0.72 bc N3-A80° 494.41±9.80 c 369.79±17.69 cd 117.27±4.69 cd 113.43±1.68 c 107.72±2.23 c N4-A30° 483.12±1.16 c 323.66±17.75 de 142.93±10.24 bc 118.15±2.31 bc 81.75±1.17 e N4-A60° 576.63±11.28 b 481.64±20.17 a 180.87±4.97 a 130.92±0.99 ab 106.09±1.71 c N4-A80° 508.92±13.08 c 433.41±1.51 b 121.59±5.50 cd 120.08±0.58 bc 95.36±1.77 d NE 0.021 0.001 ns 0.005 0.000 AE 0.000 0.000 0.000 0.000 0.000 NE × AE ns 0.000 0.000 0.038 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。
下载: 导出CSV
表 5 树形调整对香榧种实可溶性蛋白质量分数的影响
Table 5. Effect of tree shape adjustment on the content of soluble protein of the seeds
处理 可溶性蛋白质量分数/(mg·g−1) 5月 6月 7月 8月 9月 ck-A30° 135.89±6.72 d 146.02±5.22 e 172.30±3.13 fg 402.95±14.91 f 343.21±10.93 d ck-A60° 198.53±10.20 b 217.51±3.28 c 270.21±14.30 c 573.63±3.96 c 573.37±6.35 b ck-A80° 140.62±9.01 cd 151.01±3.82 e 161.66±2.30 g 235.22±5.10 h 235.33±14.75 f N3-A30° 205.62±3.35 b 215.97±10.83 c 240.41±5.43 d 484.14±5.23 d 478.95±5.14 c N3-A60° 286.88±0.64 a 329.74±6.35 b 407.50±11.09 b 647.12±12.21 b 585.41±11.49 b N3-A80° 197.94±8.84 b 200.59±7.38 cd 209.55±4.84 e 270.19±9.19 g 297.18±6.63 e N4-A30° 189.08±2.21 b 200.59±2.57 cd 223.38±2.30 e 456.88±8.05 de 460.3±7.14 c N4-A60° 283.63±8.86 a 363.95±9.76 a 444.75±11.75 a 752.61±7.16 a 680.07±15.65 a N4-A80° 163.67±2.30 c 181.76±2.74 d 195.71±1.51 ef 431.99±13.05 ef 296.71±6.88 e NE 0.000 0.000 0.000 0.000 0.000 AE 0.000 0.000 0.000 0.000 0.000 NE × AE 0.027 0.000 0.000 0.000 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。
下载: 导出CSV
表 6 树形调整对香榧种实含油率的影响
Table 6. Effect of tree shape adjustment on the oil content of the seeds
处理 含油率/% 7月 8月 9月 ck-A30° 28.34±0.16 c 45.96±0.01 abc 51.53±0.63 bcd ck-A60° 29.53±0.12 a 46.52±0.13 ab 53.59±0.53 a ck-A80° 28.09±0.32 c 45.11±0.12 c 50.19±0.44 bcd N3-A30° 28.56±0.21 c 46.17±0.23 abc 51.93±0.27 ab N3-A60° 29.65±0.19 a 47.08±0.24 a 53.89±0.69 a N3-A80° 28.12±0.26 c 45.61±0.11 bc 49.50±0.59 cd N4-A30° 28.65±0.17 bc 45.81±0.73 abc 51.26±0.28 bc N4-A60° 29.46±0.34 ab 47.03±0.38 a 53.90±0.48 a N4-A80° 28.34±0.19 c 45.14±0.20 c 49.04±0.30 d NE ns ns ns AE 0.000 0.001 0.000 NE × AE ns ns ns 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著;ns表示无显著差异。
下载: 导出CSV
表 7 树形调整对香榧种实脂肪酸组分的影响
Table 7. Effect of tree shape adjustment on the main fatty acid compositions of the seeds
处理 脂肪酸组分/(mg·g−1) 棕榈酸 硬脂酸 油酸 亚油酸 亚麻酸 二十碳二烯酸 金松酸 ck-A30° 39.04±0.23 cd 11.47±0.07 b 164.59±0.95 b 240.73±1.39 f 1.60±0.01 fg 3.64±0.02 a 47.48±0.28 f ck-A60° 44.52±0.21 a 10.50±0.05 c 151.62±0.71 c 262.57±1.22 d 1.82±0.01 d 3.45±0.02 b 54.59±0.26 c ck-A80° 38.96±0.16 cd 9.92±0.04 d 149.08±0.62 d 242.67±1.00 f 1.66±0.01 e 2.83±0.01 e 50.35±0.21 e N3-A30° 40.75±0.10 b 12.36±0.03 a 176.29±0.43 a 231.78±0.57 g 1.61±0.00 f 3.01±0.01 c 46.51±0.11 g N3-A60° 41.01±0.25 b 6.84±0.04 h 132.17±0.80 e 288.81±1.74 a 1.94±0.01 b 1.98±0.01 i 59.40±0.36 b N3-A80° 38.76±0.22 d 7.38±0.04 f 117.32±0.66 f 265.62±1.49 cd 1.89±0.01 c 2.56±0.01 g 55.18±0.31 c N4-A30° 41.03±0.11 b 9.96±0.03 d 150.38±0.39 cd 246.87±0.64 e 1.57±0.00 g 2.92±0.01 d 53.37±0.14 d N4-A60° 44.31±0.19 a 7.98±0.03 e 132.52±0.56 e 277.39±1.16 b 1.87±0.01 c 2.78±0.01 f 65.21±0.38 a N4-A80° 39.36±0.11 c 7.24±0.02 g 104.93±0.30 g 268.38±0.77 c 2.11±0.01 a 2.26±0.01 h 59.88±0.17 b NE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 AE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 NE × AE 0.000 0.000 0.000 0.000 0.000 0.000 0.000 说明:ck表示单骨干枝自然圆头形;N3~N4分别表示3、4条骨干枝开心形;A30°、A60°、A80°分别表示30°、60°、80°开张角度;同列不同小写字母表示不同处理间差异显著(P<0.05)。NE表示骨干枝效应;AE表示角度效应;NE×AE表示交互效应;P<0.05表示差异显著。
下载: 导出CSV
-
[1] WU Jiasheng, HUANG Jiandiao, HONG Yiwei, et al. De novo transcriptome sequencing of Torreya grandis reveals gene regulation in sciadonic acid biosynthesis pathway [J]. Industrial Crops &Products, 2018, 120: 47 − 60. [2] SUO Jinwei, TONG Ke, WU Jiasheng, et al. Comparative transcriptome analysis reveals key genes in the regulation of squalene and β-sitosterol biosynthesis in Torreya grandis [J]. Industrial Crops &Products, 2019, 131: 182 − 193. [3] SUO Jinwei, MA Zhenmin, ZHAO Bing, et al. Metabolomics reveal changes in flavor quality and bioactive components in post-ripening Torreya grandis nuts and the underlying mechanism [J/OL]. Food Chemistry, 2023, 406: 134987[2024-01-11]. doi: 10.1016/j.foodchem.2022.134987. [4] YAN Jingwei, ZENG Hao, CHEN Wenchao, et al. New insights into the carotenoid biosynthesis in Torreya grandis kernels [J]. Horticultural Plant Journal, 2023, 2(6): 1 − 11. [5] ZHANG Zuying, TAO Liu, GAO Lingling, et al. Transcription factors TgbHLH95 and TgbZIP44 cotarget terpene biosynthesis gene TgGPPS in Torreya grandis nuts [J]. Plant Physiology, 2023, 193(2): 1161 − 1176. [6] LOU Heqiang, ZHENG Shan, CHEN Wenchao, et al. Transcriptome-referenced association study provides insights into the regulation of oil and fatty acid biosynthesis in Torreya grandis kernel [J/OL]. Journal of Advanced Research, 2023[2024-01-11]. doi: 10.1016/j.jare.2023.01.007. [7] YAN Jingwei, ZENG Hao, CHEN Weijie, et al. Effects of tree age on flavonoids and antioxidant activity in Torreya grandis nuts via integrated metabolome and transcriptome analyses [J]. Food Frontiers, 2023, 4: 358 − 367. [8] HU Yuanyuan, ZHANG Yongling, YU Weiwu, et al. Novel insights into the influence of seed sarcotesta photosynthesis on accumulation of seed dry matter and oil content in Torreya grandis cv. “Merrillii” [J]. Frontiers in Plant Science, 2018, 8: 2179. [9] 何风杰, 洪小玲, 邱智敏, 等. 台州市香榧产业发展现状、存在问题及对策[J]. 果树资源学报, 2023, 4(2): 77 − 80. HE Fengjie, Hong Xiaoling, QIU Zhimin, et al. Development status, existing problem and countermeasure of Torreya grandis ‘Merrillii’ in Taizhou city [J]. Journal of Fruit Resources, 2023, 4(2): 77 − 80. [10] 叶淑媛, 曾燕如, 胡渊渊, 等. 香榧初结果母枝性状变化规律与结实能力的关系[J]. 浙江农林大学学报, 2022, 39(1): 41 − 49. YE Shuyuan, ZENG Yanru, HU Yuanyuan, et al. Relationship between character changing and seed-bearing capacity of initial seed-bearing mother shoots in Torreya grandis ‘Merrillii’ [J]. Journal of Zhejiang A&F University, 2022, 39(1): 41 − 49. [11] 郝婕, 索相敏, 李学营, 等. 不同整形修剪模式下富士苹果树体结构及相关因素分析[J]. 东北农业科学, 2017, 18(12): 2528 − 2535. HAO Jie, SUO Xiangmin, LI Xueying, et al. Analysis on Fuji apple tree structures and related factors under different pruning modes [J]. Journal of Northeast Agricultural Sciences, 2017, 18(12): 2528 − 2535. [12] 贺梅英. 荔枝龙眼示范园间伐技术经济效益评价[J]. 广东农业科学, 2015, 42(18): 37 − 42. HE Meiying. Economic benefit evaluation of thinning technology in litchi and longan demonstration orchards [J]. Guangdong Agricultural Sciences, 2015, 42(18): 37 − 42. [13] 苏渤海, 范崇辉, 李国栋, 等. 红富士苹果改形过程中不同树形光照分布及其对产量品质的影响[J]. 西北农林科技大学学报(自然科学版), 2008, 36(1): 158 − 162. SU Bohai, FAN Chonghui, LI Guodong, et al. Effects of modifying between light distribution, yield and quality of different shapes on ‘Red Fuji’ apple [J]. Journal of Northwest A&F University (Natural Science Edition), 2008, 36(1): 158 − 162. [14] 牟红梅, 于强, 李庆余, 等. ‘莱阳茌梨’高光效树形光合特性与果实品质的研究[J]. 北方园艺, 2018, 12: 58 − 62. MOU Hongmei, YU Qiang, LI Qingyu, et al. Research on photosynthetic characteristics and fruit quality of high light use efficiency tree shape on ‘Laiyang pear’ [J]. Northern Horticulture, 2018, 12: 58 − 62. [15] 刘丙花, 唐贵敏, 梁静. 不同树形对早实核桃‘鲁光’坚果产量和品质的影响[J]. 果树学报, 2021, 38(1): 73 − 81. LIU Binghua, TANG Guimin, LIANG Jing. Effects of different tree shapes on yield and quality of ‘Luguang’ walnut [J]. Journal of Fruit Science, 2021, 38(1): 73 − 81. [16] 何双凌, 岳海, 陈宇春, 等. 不同树形澳洲坚果‘O. C’坐果特征与产量的关系[J]. 西部林业科学, 2023, 52(5): 7 − 12. HE Shuangling, YUE Hai, CHEN Yuchun. Effect of setting fruit characteristics and yield of ‘O. C’ macadamia nut in different tree shapes [J]. Journal of West China Forestry Science, 2023, 52(5): 7 − 12. [17] 熊欢, 郭素娟, 彭晶晶, 等. 树体结构对板栗冠层光照分布和果实产量及品质的影响[J]. 南京林业大学学报(自然科学版), 2014, 38(2): 68 − 74. XIONG Huan, GUO Sujuan, PENG Jingjing. Effect of tree structure on light distribution and yield and quality of Castanea mollissima Bl. [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2014, 38(2): 68 − 74. [18] 李艳芳. 拉枝对枣树结果、叶片营养和光合特性的影响[D]. 保定: 河北农业大学, 2010. LI Yanfang. The Effects of Branch on Fruiting, Leaf Nutrition and Photosynthetic Characteristics in Chinese Jujube [D]. Baoding: Agricultural University of Hebei Province, 2010. [19] 蔡虎, 张文娥, 吴浪, 等. 拉枝对黔核7号泡核桃叶片营养、成花及坚果品质的影响[J]. 果树学报, 2022, 39(1): 60 − 67. CAI Hu, ZHANG Wene, WU Lang, et al. Effects of branch bending angle on leaf nutrition, flower formation and nut quality of Qianhe 7 walnut [J]. Journal of Fruit Science, 2022, 39(1): 60 − 67. [20] 刘琏, 陆吕佳, 胡渊渊, 等. 不同月份不同树龄香榧叶片光合特性的变化[J]. 浙江农林大学学报, 2022, 39(1): 32 − 40. LIU Lian, LU Lüjia, HU Yuanyuan, et al. Changes of photosynthesis in leaves of Torreya grandis ‘Merrillii’ in different months and different tree ages [J]. Journal of Zhejiang A&F University, 2022, 39(1): 32 − 40. [21] 成豪, 吴家胜, 马爽, 等. 树形调整对香榧成花和坐果的影响[J]. 林业科学, 2023, 59(11): 49 − 58. CHENG Hao, WU Jiasheng, MA Shuang, et al. Effects of tree shape adjustment on flower formation and fruit setting of Torreya grandis cv. Merrillii [J]. Scientia Silvae Sinicae, 2023, 59(11): 49 − 58. [22] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. 食品中粗脂肪的测定: GB/T 14772—2008[S]. 北京: 中国标准出版社, 2009. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China. Determination of Crude Fat in Foods: GB/T 14772−2008 [S]. Beijing: Standards Press of China, 2009. [23] ZHOU Minying, SONG Lili, YE Shan, et al. New sights into lipid metabolism regulation by low temperature in harvested Torreya grandis nuts [J]. Research Article, 2019, 99: 4226 − 4234. [24] HAMADZIRIPI E T, THERON K I, MULLER M, et al. Apple compositional and peel color differences resulting from canopy microclimate affect consumer preference for eating quality and appearance [J]. Nature, 2006, 439(7075): 457 − 461. [25] 魏钦平, 鲁韧强, 张显川, 等. 富士苹果高干开心形光照分布与产量品质的关系研究[J]. 园艺学报, 2004, 31(3): 291 − 296. WEI Qinping, LU Renqiang, ZHANG Xianchuan, et al. Relationships between distribution of relative light intensity and yield and quality in different tree canopy shapes for ‘Fuji’ apple [J]. Acta Horticulturae Sinica, 2004, 31(3): 291 − 296. [26] 史祥宾, 刘凤之, 程存刚, 等. 设施葡萄不同新梢间距处理对冠层光环境及果实品质的影响[J]. 园艺学报, 2018, 45(3): 436 − 446. SHI Xiangbin, LIU Fengzhi, CHENG Cungang, et al. Effects of different new shoots spacing on canopy light environment and fruit quality of grapevine under protected cultivation [J]. Acta Horticulturae Sinica, 2018, 45(3): 436 − 446. [27] 张海旺, 张文臣, 贾浩, 等. 不同树形桃树冠层光照分布对果实产量和品质的影响[J]. 北方园艺, 2024(4): 21 − 27. ZHANG Haiwang, ZHANG Wenchen, JIA Hao, et al. Effects of light distribution in canopy of peach trees with different tree shapes on fruit yield and quality [J]. Northern Horticulture, 2024(4): 21 − 27. [28] 张红欢, 杨兴旺, 冀晓昊, 等. 树形对促早栽培桃冠层结构、光合特性及果实品质的影响[J]. 果树学报, 2024, 41(3): 470 − 480. ZHANG Honghuan, YANG Xingwang, JI Xiaohao, et al. Effects of tree shape on canopy structure, photosynthetic characteristics and fruit quality of early cultivated peach [J]. Journal of Fruit Science, 2024, 41(3): 470 − 480. [29] 刘珊珊, 令狐田, 赵志华, 等. 不同树形对砀山酥梨冠层结构、光合特性及果实品质的影响[J]. 果树资源学报, 2021, 3(5): 41 − 49. LIU Shanshan, LING Hutian, ZHAO Zhihua, et al. Effects of different tree shapes on canopy structure, photosynthetic characteristics, fruit quality of ‘Dangshan’ pear [J]. Journal of Fruit Resources, 2021, 3(5): 41 − 49. [30] 赵国栋, 贾林光, 张新生, 等. 拉枝角度对‘宫崎短枝富士’树体生长、果实品质和质地的影响[J]. 西北农业学报, 2021, 30(1): 102 − 108. ZHAO Guodong, JIA Linguang, ZHANG Xinsheng, et al. Effect of branch bending angle on growth, fruit quality and texture properties of Malus domestica Borkh ‘Miyazakifuji’ [J]. Acta Agriculturae Boreali-occidentalis Sinica, 2021, 30(1): 102 − 108. [31] 黄丽, 王亮, 赵迎丽, 等. 3种梨贮藏期间果实品质、可溶性糖和有机酸含量变化[J]. 食品研究与开发, 2023, 44(10): 46 − 52. HUANG Li, WANG Liang, ZHAO Yingli, et al. Changes in fruit quality, soluble sugar and organic acid content of three pear species during storage [J]. Food Research and Development, 2023, 44(10): 46 − 52. [32] 沈乐意, 王立如, 徐悦, 等. 不同砧木对‘阳光玫瑰’葡萄果实品质及糖异生相关基因表达的影响[J]. 农业生物技术学报, 2023, 31(12): 2490 − 2505. SHEN Leyi, WANG Liru, XU Yue, et al. Effect of different rootstocks on fruit quality and expression of genes related to gluconeogenesis in ‘Shine Muscat’ grapes (Vitis vinfera) [J]. Journal of Agricultural Biotechnology, 2023, 31(12): 2490 − 2505. [33] 于文剑, 杨丽, 张俊环, 等. 杏果实风味形成及调控机制研究进展[J]. 果树学报, 2023, 40(12): 2624 − 2637. YU Wenjian, YANG Li, ZHANG Junhuan, et al. Research progress on the mechanism of flavor formation and regulation in apricot [J]. Journal of Fruit Science, 2023, 40(12): 2624 − 2637. [34] 赵旗峰, 郭文娇, 荀志丽, 等. 树形对酿酒葡萄果实糖含量及蔗糖代谢相关酶活性的影响[J]. 河南农业科学, 2021, 50(5): 122 − 128. ZHAO Qifeng, GUO Wenjiao, XUN Zhili, et al. Effect of tree shape on sugar content and enzymes activities of sucrose metabolism in wine grape fruit [J]. Journal of Henan Agricultural Sciences, 2021, 50(5): 122 − 128. [35] 汪志辉, 熊碧玲, 刘燕, 等. 树形对鲜黄梨果实糖积累及山梨醇转化相关酶活性的影响[J]. 林业科学, 2011, 47(4): 27 − 32. WANG Zhihui, XIONG Biling, LIU Yan, et al. Effects of tree shape on the sugar accumulation and activity of sorbitol-invertase of Pyrus pyrifolia ‘Sunhwang’ [J]. Scientia Silvae Sinicae, 2011, 47(4): 27 − 32. [36] 林敏娟, 王建军, 吴翠云. 不同拉枝角度富士苹果叶片营养物质含量与果实品质之间的关系[J]. 塔里木大学学报, 2012, 24(3): 36 − 39. LIN Minjuan, WANG Jianjun, WU Cuiyun. Leaf nitrogen, phosphorus and potassium of different branch angle impact on fruit quality in Fuji [J]. Journal of Tarim University, 2012, 24(3): 36 − 39. [37] 王振磊, 林敏娟, 吴翠云. 不同拉枝角度对富士苹果树体生理特性和果实品质的影响[J]. 新疆农业科学, 2013, 50(8): 1462 − 1467. WANG Zhenlei, LIN Minjuan, WU Cuiyun. Effect of different branch bending angles on physiological characteristics and fruit quality of Fuji apple [J]. Xinjiang Agricultural Sciences, 2013, 50(8): 1462 − 1467. [38] 刘雪芳, 刘慧芳, 常月梅. ‘中林1号’核桃不同树形坚果品质分析[J]. 天津农业科学, 2021, 27(9): 19 − 23. LIU Xuefang, LIU Huifang, CHANG Yuemei. analysis on quality of ‘Zhonglin No. 1’ walnut with different tree shapes [J]. Tianjin Agricultural Sciences, 2021, 27(9): 19 − 23. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240178
 点击查看大图
点击查看大图
计量
- 文章访问数: 1647
- HTML全文浏览量: 346
- PDF下载量: 52
- 被引次数: 0
