





-
种间关系是植物群落中不同种群间相互作用的依赖或竞争关系,是群落结构形成的关键驱动力[1−3],分析种间关系对揭示群落演替与更新机制至关重要[4−6]。种间联结用以描述不同物种在空间分布上的相互关联性,作为定性指标常用来反映物种同时出现的相似尺度[7]。生态位可以体现种群在生态系统中的时空位置与功能关系,揭示种群地位、作用及生境需求[8]。研究树种生态位及种间联结性可掌握种群生存现状与群落演替特征,为种群恢复和物种保护提供理论基础[9]。
楠木是一类珍贵木材资源的总称,主要包括润楠属Machilus、楠属Phoebe在内的多种植物及其近缘类群。由于长期人为破坏及楠木自身更新不良等因素,中国楠木种群数量逐渐减少,在各类天然林中呈零星分布状态,少有成片分布[10]。任晴[11]研究发现:红楠Machilus thunbergii与枫香Liquidambar formosana、榆树Ulmus pumila、白栎Quercus fabri等树种具有高生态位重叠值,它们对资源竞争激烈。减少生态位重叠程度高的树种,可以达到保护相关物种的目的。李冰[12]研究指出:毛竹Phyllostachys edulis对浙江楠Phoebe chekiangensis存在种间竞争,伐去部分毛竹可促进浙江楠群落可持续生长。李铁华等[13]通过分析闽楠P. bournei与木荷Schima superba 林分空间结构与种间关系,发现木荷是闽楠较好的伴生树种。在浙江省建德市寿昌林场的绿荷塘林区,存在着1片被誉为亚洲最大的天然古楠木林,面积超过30 hm2,楠木种类有刨花润楠M. pauhoi、华东楠M. leptophylla、红楠、紫楠P. sheareri。目前对该楠木林研究报道较少。何贵平等[14]发现:在该群落中刨花润楠占主导地位;吴初平等[15]认为:楠木具有较好的更新能力;吴丹婷等[16]指出:该群落中乔木层、灌木层和草本层均处于不稳定阶段。本研究通过1 hm2大样地的调查,分析建德楠木林中各种群的群落结构与种间联结性特征,试图从种间关系的角度揭示楠木天然林的物种共存机制,旨在为建德楠木林的生态恢复和生物多样性保护提供参考。
-
建德楠木林位于浙江省建德市绿荷塘国家森林公园,地理位置为29°25′45″~29°27′01″N,119°08′45″~119°11′00″E,属千里岗山脉。该地区属亚热带季风气候,四季分明。年平均气温为17.6 ℃,极端最高气温为42.2 ℃,极端最低气温为−13.9 ℃。年平均降水量为1 700.0 mm,平均相对湿度为82%,年日照数为1 762.0 h,全年无霜期为265.0 d。土壤以酸性红壤为主,土层厚度为30~80 cm。
-
2023年7月,在对绿荷塘国家森林公园内的楠木林进行全面踏查的基础上,设置1 hm2大样地1个,将其分为25个20 m×20 m的调查样方。对样方内所有胸径≥1 cm的林木个体进行每木检尺,详细记录树种名称、胸径、树高、枝下高及空间坐标等信息。
-
通过物种重要值(VI)确定群落中木本植物的优势树种。物种重要值参考马克平等[17]的方法计算。
-
参考PIANKA[18]的方法,计算木本植物优势树种的Levins生态位宽度指数(BL)和Shannon生态位宽度指数(BS),并使用Pianka指数(Oik)计算生态位重叠指数。
-
采用方差比率(RV),利用统计量(W)来检验物种的整体关联程度[19]。
$$ {R}_{\mathrm{V}}=\frac{1}{r}\displaystyle\sum _{j=1}^{r}{({T}_{j}-t)}^{2}/\displaystyle\sum _{i=1}^{S}\left(1-\frac{{r}_{i}}{r}\right)\frac{{r}_{i}}{r} \text{;} $$ $$ W={R}_{\mathrm{V}}\times r 。 $$ 其中:r为样方总数;$ {T}_{j} $为样方j内出现的物种数;t为所有样方内物种平均数;S为总物种数;$ \dfrac{{r}_{i}}{r} $为物种i出现的频度;ri为物种i出现的样方数。
-
参照江常春等[20]的方法,采用物种联结性χ2 统计量测定种间关联性,以确定实测值与期望值之间偏差的显著程度。
$$ \chi^2=\frac{r\left(\right|ad-bc|-0.5r{)}^{2}}{(a+b)(c+d)(a+c)(b+d)}。 $$ 其中:r为样方总数。当ad>bc时,表示种对呈正联结;当ad<bc时,表示种对呈负联结。χ2<3.841时,表示种对间联结不显著(P>0.05);χ2>6.635时,表示种对间联结极显著(P<0.01);3.841<χ2<6.635时,表示种对间联结显著(0.01<P<0.05)。
按照郭志华等[21]的方法计算种间联结系数(CA)和共同出现百分率(CP)。
-
建德楠木林群落共有75种木本植物,隶属于26科47属,其中重要值≥1%的植物种类有19种(表1),重要值总和为84.41%。从表1可知:刨花润楠和甜槠Castanopsis eyrei的重要值分别为23.44%和13.92%,是该群落的建群树种,其他楠木种类还有华东楠、红楠、紫楠,重要值分别为1.73%、1.68%、1.20%,排名分别为第12、13、19位。19个优势种的Shannon生态位宽度和Levins生态位宽度平均值分别为2.12和7.34。该群落中生态位宽度最大的树种为矩形叶鼠刺Itea oblonga,其Shannon生态位宽度和Levins生态位宽度分别为2.72和12.06。刨花润楠在群落中重要值虽然排在第1位,为23.44,但其Shannon生态位宽度和Levins生态位宽度为2.62和9.44,排名为第3位和第6位。华东楠、红楠、紫楠的Shannon生态位宽度分别为1.41、1.97、1.52,排名分别为第18、13、17位,Levins生态位宽度分别为2.98、5.65、3.76,排名分别为第19、14、16位。群落中刨花润楠有较高的重要值且占据较大的生态位宽度,华东楠、红楠、紫楠重要值和生态位宽度均较小。
表 1 主要优势树种重要值及生态位宽度
Table 1. Importance values and niche width of dominant tree species
排名 植物 平均胸径/cm 平均树高/m 个体数/株 重要值/% 生态位宽度指数 Shannon Levins 1 刨花润楠 Machilus pauhoi 12.2 9.6 347 23.44 2.62 9.44 2 甜槠 Castanopsis eyrei 13.9 9.5 159 13.92 2.34 8.10 3 乌楣栲Castanopsis jucunda 10.6 8.1 34 5.30 2.56 10.80 4 栲 Castanopsis fargesii 9.0 7.5 76 4.98 2.63 10.11 5 矩形叶鼠刺 Itea oblonga 10.5 6.8 123 4.84 2.72 12.06 6 青冈 Cyclobalanopsis glauca 15.6 10.5 58 4.53 2.48 9.02 7 毛花连蕊茶 Camellia fraternal 10.6 7.6 124 4.51 2.52 9.82 8 微毛柃 Eurya hebeclados 9.9 7.3 126 4.31 2.35 9.31 9 细叶香桂 Cinnamomum subavenium 9.9 7.9 72 2.98 2.25 8.15 10 枫香 Liquidambar formosana 10.0 7.7 12 2.02 1.98 6.55 11 拟赤杨 Alniphyllum fortunei 6.8 7.2 36 1.82 1.52 3.66 12 华东楠 Machilus leptophylla 9.5 8.6 28 1.73 1.41 2.98 13 红楠 Machilus thunbergii 10.9 8.3 19 1.68 1.97 5.65 14 毛八角枫 Alangium kurzii 10.8 7.4 31 1.66 1.76 4.06 15 木荷 Schima superba 19.5 11.3 16 1.47 1.37 3.12 16 老鼠矢 Symplocos stellaris 11.1 6.9 12 1.40 2.34 9.80 17 檵木 Loropetalum chinense 6.3 6.7 9 1.31 1.89 6.37 18 厚皮香 Ternstroemia gymnanthera 7.8 7.0 12 1.30 2.14 6.76 19 紫楠 Phoebe sheareri 23.5 16.3 10 1.20 1.52 3.76 -
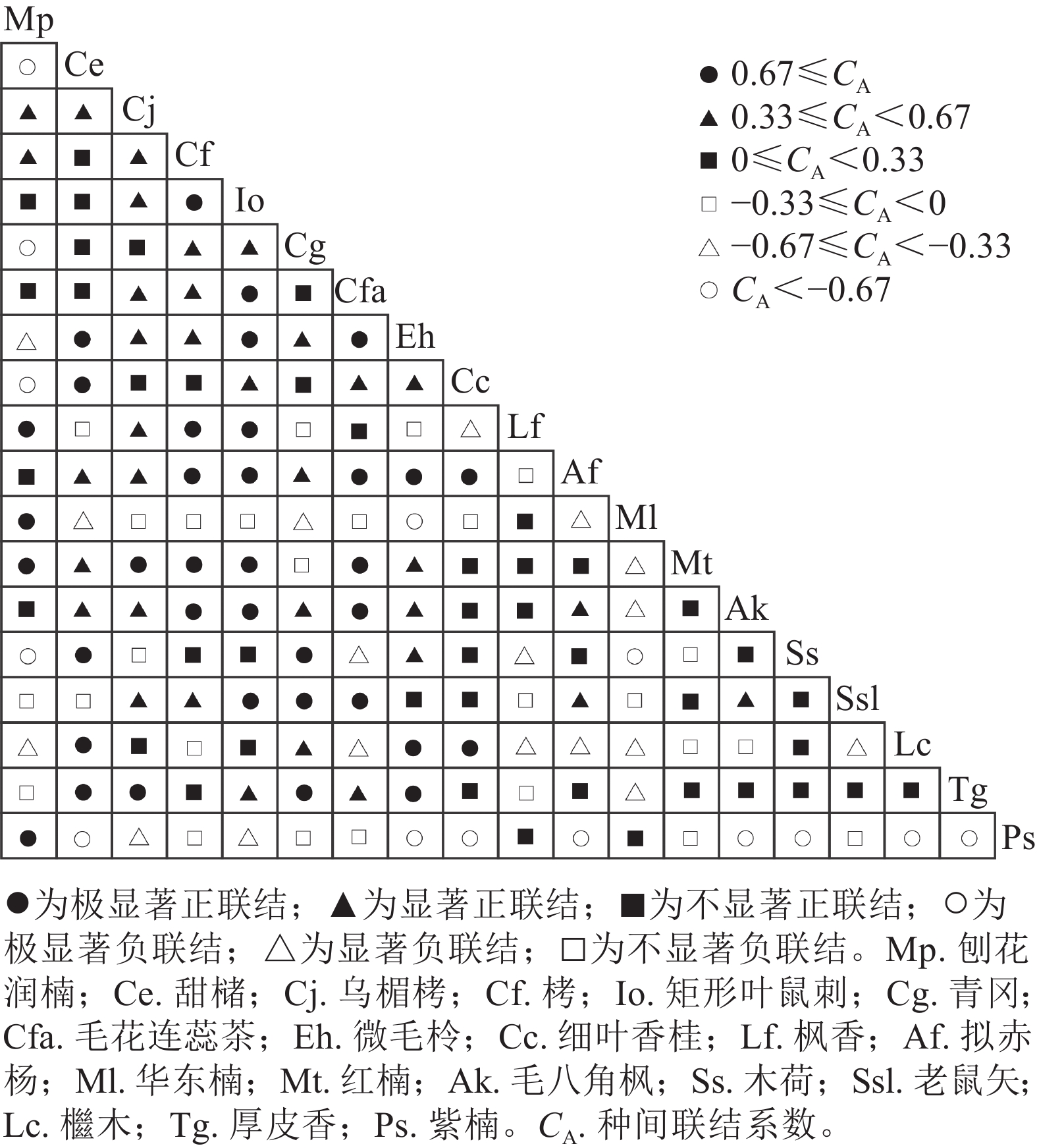
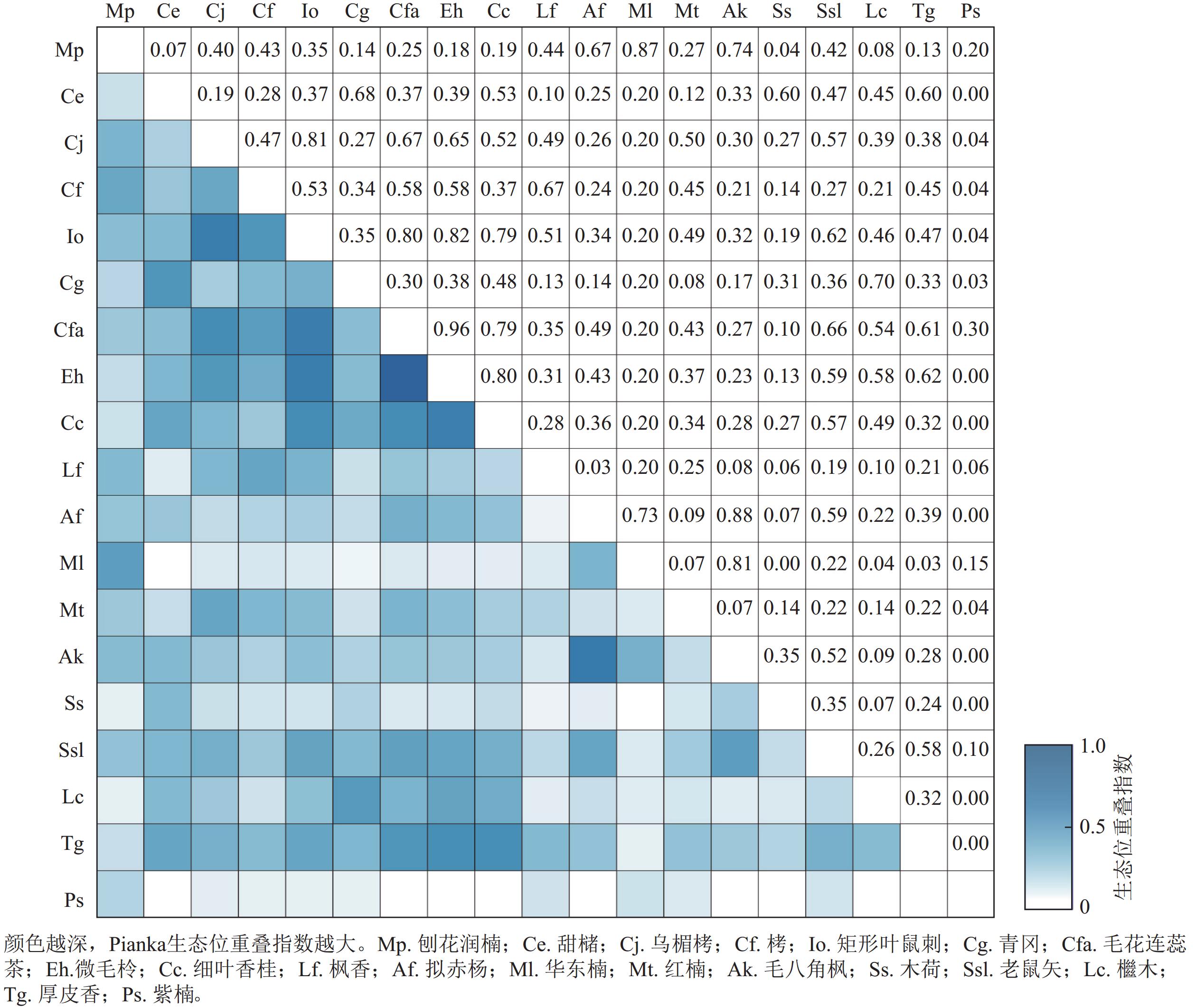
建德楠木林群落优势树种的Pianka生态位重叠指数如图1所示。重要值≥1%的优势树种组成的171组种对中,有164组种对间存在生态位重叠,占所有种对数的95.91%,所有种对的生态位重叠指数平均值为0.28。微毛柃Eurya hebeclados和毛花连蕊茶Camellia fraternal的Pianka生态位重叠指数最高,为0.96。Pianka生态位重叠指数≥0.50的种对有37对,占21.64%,小于0.50的种对数有134对,占78.36%。刨花润楠与群落中其他18个优势树种都存在较明显的生态位重叠,Pianka生态位重叠指数平均值为0.32,与华东楠的重叠指数达0.87,为所有种对中的最大值。
图 1 主要优势树种 Pianka 生态位重叠指数
Figure 1. Pianka niche overlap index of dominant tree species
华东楠的Pianka生态位重叠指数平均值为0.24,与毛八角枫、拟赤杨生态位重叠指数分别为0.81和0.73,排名分别为第2位和第3位。与木荷不存在生态位重叠。红楠的Pianka生态位重叠指数平均值为0.23,与乌楣栲生态位重叠指数最高,达0.50,与矩形叶鼠刺、栲的生态位重叠指数分别为0.49、0.45,排名第2位和第3位。紫楠的Pianka生态位重叠指数均值为0.06,与毛花连蕊茶生态位重叠指数最高,为0.30,与乌楣栲、栲、矩形叶鼠刺、红楠生态位重叠指数均为0.04,与甜槠、微毛柃、细叶香桂、拟赤杨等8个树种间没有生态位重叠。
-
建德楠木林群落总体关联性的方差比率(RV)=3.21,大于1,表明群落中优势物种的总体关联性呈现为正联结。χ2临界值[ χ2 (0.95,25)和χ2(0.05,25)]分别为14.611和37.652,检验统计量(W)为80.29。通过检验统计量(W)检验方差比率(RV)与1的偏差程度,W并未落在χ2(0.95)<W<χ2(0.05)范围内,表明优势树种之间联结显著(P<0.05),总体联结性为显著正联结(P<0.05)。
-
建德楠木林重要值≥1%的优势树种 χ2统计量检验结果如表2所示。有9对种间联结性达极显著水平(P<0.01),占比为5.27%;有11对种间联结性显著(P<0.05),占比为6.43%;有151对种间联结性不显著,占比为88.30%。紫楠-甜槠(10.62)、紫楠-微毛柃(7.28)种间联结性达极显著(P<0.01),刨花润楠-木荷(3.87)、红楠-乌楣栲(4.52)、紫楠-细叶香桂(4.98)等种间联结性达显著水平(P<0.05),华东楠与所有优势树种联结性均不显著。该群落优势树种间独立性较强。
表 2 优势树种χ2统计量检验
Table 2. χ2 correlation test of dominant tree species
树种 Mp Ce Cj Cf Io Cg Cfa Eh Cc Lf Af Ml Mt Ak Ss Ssl Lc Tg Ce 1.14 Cj 2.04 2.09 Cf 0.91 0.53 4.15* Io 0.17 3.14 9.66** 9.77** Cg 0.83 0.31 0.00 0.93 0.93 Cfa 0.21 0.83 9.69** 1.50 5.47* 0.06 Eh 0.08 14.52** 2.92 1.71 5.37* 0.72 4.72* Cc 2.98 5.50* 0.09 0.01 0.81 0.09 0.59 5.03* Lf 0.83 0.12 0.95 1.39 1.39 0.00 0.06 0.00 1.32 Af 0.21 0.90 0.50 1.00 1.00 0.50 2.10 2.01 3.64 0.06 Ml 0.35 1.71 0.34 0.12 0.12 2.52 0.73 3.08 0.13 0.18 0.04 Mt 1.14 0.41 4.52* 1.83 1.83 0.31 3.51 1.50 0.02 0.12 0.00 0.41 Ak 0.00 0.41 1.52 1.83 1.83 1.52 3.51 1.50 0.02 0.12 7.65** 0.41 0.05 Ss 3.87* 2.62 0.34 0.12 0.12 2.03 0.04 1.16 0.13 0.18 0.04 1.06 0.11 0.11 Ssl 0.08 0.15 0.78 0.50 2.93 3.04 2.01 0.08 0.03 0.00 4.72* 0.02 0.21 8.83** 0.02 Lc 2.81 3.51 0.06 0.01 0.01 0.50 0.21 2.01 3.64 0.50 0.21 0.04 0.00 0.00 0.04 0.27 Tg 0.01 6.95** 2.21 0.26 0.26 2.20 1.40 5.69* 0.33 0.07 0.41 0.74 0.59 0.59 1.11 0.01 2.39 Ps 0.35 10.62** 2.52 0.12 2.23 0.34 0.73 7.28** 4.98* 0.18 1.51 0.00 0.11 2.62 1.06 1.06 1.51 3.30 说明:**为联结性极显著(P<0.01);*为联结性显著(P<0.05)。Mp. 刨花润楠;Ce. 甜槠;Cj. 乌楣栲;Cf. 栲;Io. 矩形叶鼠刺;Cg. 青冈;Cfa. 毛花连蕊茶;Eh. 微毛柃;Cc. 细叶香桂;Lf. 枫香;Af. 拟赤杨;Ml. 华东楠;Mt. 红楠;Ak. 毛八角枫;Ss. 木荷;Ssl. 老鼠矢;Lc. 檵木;Tg. 厚皮香;Ps. 紫楠。 -
由图2可知:分析的171个种对的联结情况中,呈正联结种对数有113对,占比为66.1%,负联结种对数有58对,占比为33.9%,正负关联比为2∶1。极显著正联结(P<0.01)的种对数有36对,占总种对数的21.1%。显著正联结(P<0.05)的种对数有34对,占比为19.8%。不显著正联结有43对,占比为25.1%。极显著负联结(P<0.01)有14对,占比为8.2%,显著负联结(P<0.05)有16对,占比为9.4%。不显著负联结有28对,占比为16.4%。刨花润楠与枫香、紫楠、红楠、华东楠等4个种对呈极显著正联结(P<0.01),与木荷、细叶香桂、青冈、甜槠等4个种对呈极显著负联结(P<0.01);华东楠与微毛柃呈极显著负联结(P<0.01);红楠与矩形叶鼠刺、栲、毛花连蕊茶、乌楣栲呈极显著正联结(P<0.01);紫楠与甜槠、微毛柃、细叶香桂、拟赤杨、毛八角枫、木荷、檵木、厚皮香呈极显著负联结(P<0.01)。
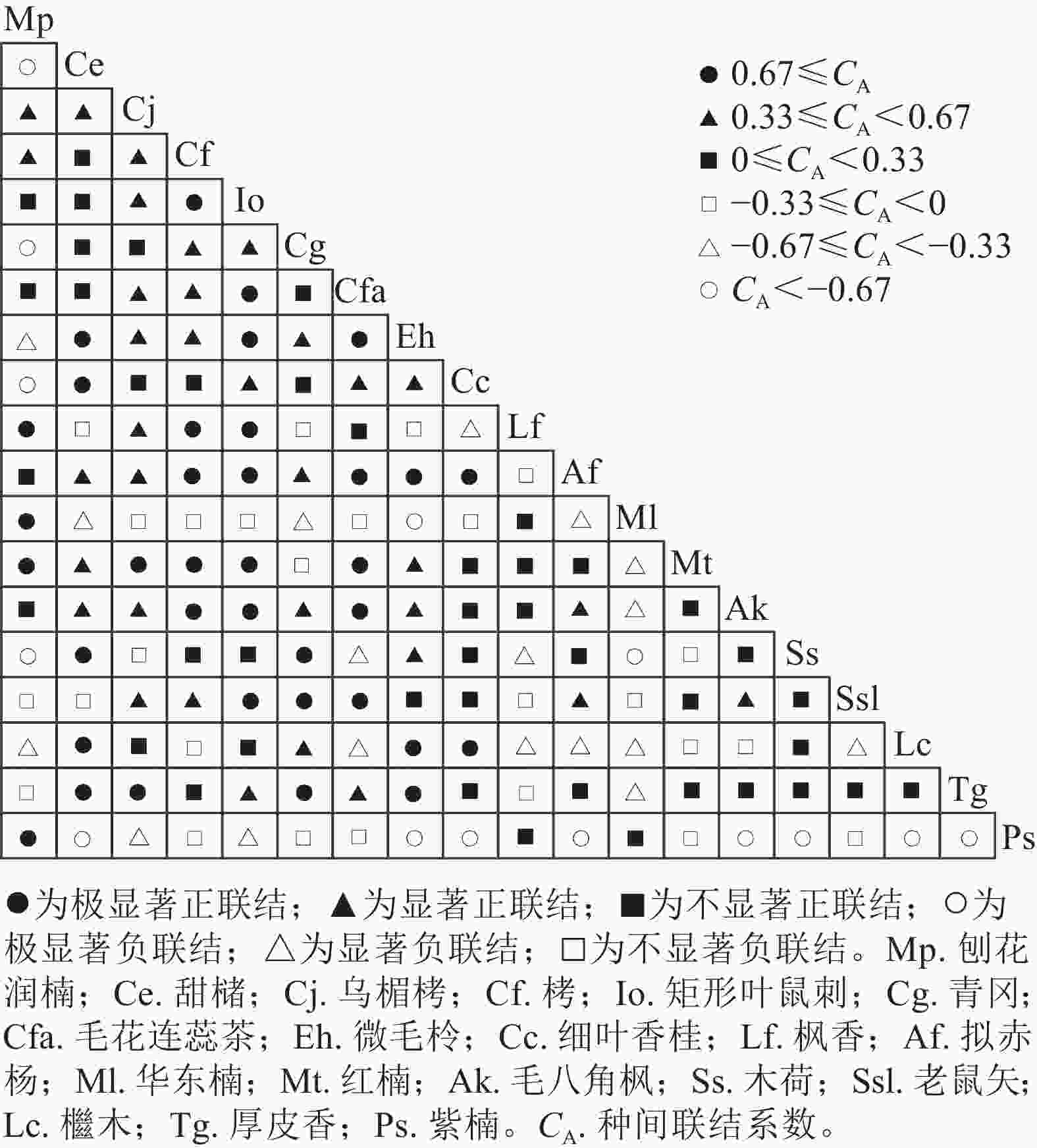
图 2 优势植物种间联结性半矩阵图
Figure 2. Semi-matrix diagram of association coefficients of interspecific associations of dominant plants
-
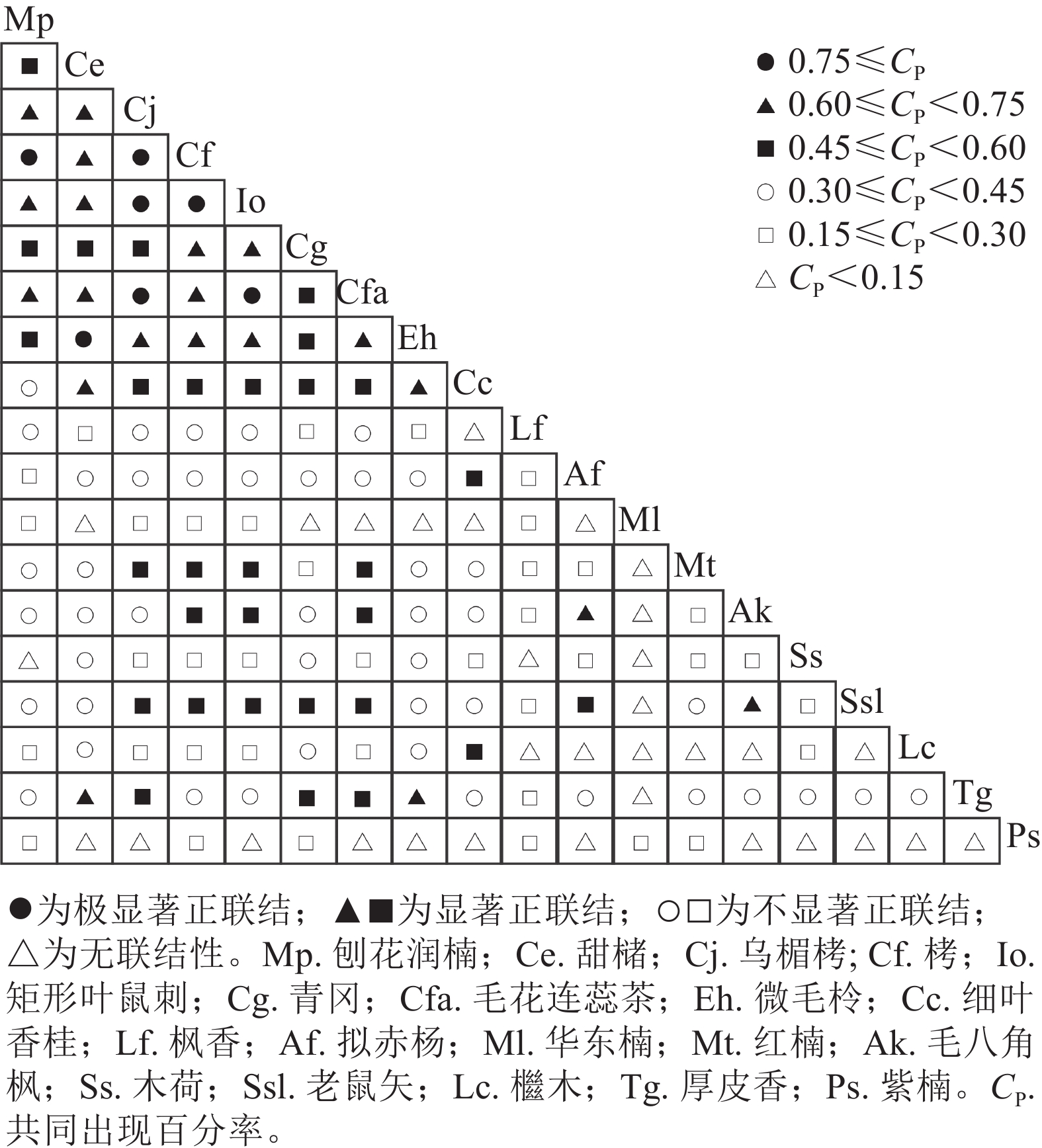
共同出现百分率检验结果由图3可知:极显著正联结(P<0.01)的种对有7对,占比为4.1%;显著正联结(P<0.05)有50对,占比为29.2%;不显著正联结(P>0.05)有82对,占48.0%;无联结性(P>0.05)为32对,占总种对的18.7%。刨花润楠与栲呈极显著正联结(P<0.01),与木荷无联结性;华东楠与厚皮香、檵木、老鼠矢等10个树种无联结性;红楠与毛花连蕊茶、矩形叶鼠刺、栲、乌楣栲呈显著正联结(P<0.05);紫楠与木荷、细叶香桂、毛花连蕊茶等12个树种无联结性,与栲、青冈等6个树种呈不显著正联结(P>0.05)。

图 3 优势植物共同出现百分率半矩阵图
Figure 3. Semi-matrix diagram of co-occurrence percentage for dominant plant species
-
建德楠木林优势树种的Pianka生态位重叠值与种间联结系数、共同出现百分率的回归分析如图4所示。从图4可知:建德楠木林优势树种生态位重叠值与种间联结系数、共同出现百分率之间存在极显著的正相关关系(P<0.01)。

图 4 生态位重叠值与种间联结系数、共同出现百分率的线性回归分析
Figure 4. Linear regression analysis between niche overlap and interspecific association coefficient, and percentage of co-occurrence
-
生态位宽度是评估物种对资源环境利用程度的关键量化指标,可以反映物种在群落中的生态位置。物种与其他物种的生态位重叠度也反映了物种对资源和环境的利用差异,以及与其他物种的竞争关系。这些指标共同构成了理解群落结构和物种相互作用的重要基础[22]。研究区内楠木有刨花润楠、华东楠、红楠、紫楠,其中刨花润楠的重要值最高,且Shannon生态位宽度和Levins生态位宽度均较大,表明刨花润楠对环境资源具有较高的利用率。野外调查发现,群落内刨花润楠大多是大胸径的高大个体,竞争能力强,分布于各样方,对环境影响大,成年个体对环境适应性强。从建德楠木林种群结构的分布情况来看[23],华东楠仅在少数几个样方中有分布,其重要值排在第12位,而红楠、紫楠、毛八角枫、木荷等物种重要值虽然比华东楠低,但其生态位宽度均比华东楠高,这可能与这些物种在样方内分布均匀有关。这一结果表明:华东楠种群在环境资源利用方面表现出相对较弱的能力,且其地理分布区狭窄,表明该物种的生态位宽度受到分布频率、植物生态学特征等多方面影响[24]。紫楠重要值和生态位宽度均较小,导致紫楠与大多数树种没有生态位重叠,这可能是紫楠数量太少的缘故。在生态位重叠程度方面,刨花润楠与其他优势种重叠程度显著,生态位重叠指数均值为0.32,与华东楠重叠指数最大。华东楠与毛八角枫、拟赤杨有较高的生态位重叠,说明它们对共同资源的竞争程度较高。红楠、紫楠与其他优势树种重叠不明显。综上所述,刨花润楠种群具备较强的生存和竞争能力,而华东楠、红楠、紫楠各自占据的生态位空间相对独立,与其他物种联系较少,彼此之间在资源利用上的直接竞争压力较小。
-
总体联结性揭示群落演替阶段、群落结构和物种组成的稳定性[25]。当群落趋向于顶级演替时,群落越趋向正联结,群落结构也越完整,且物种间充分利用环境资源,相互促进生长[26−27]。研究区域内优势树种的总体联结性呈显著正相关,正联结种对数量明显多于负联结种对,极显著关联种对数量较少,这在一定程度上揭示了该群落物种间生态习性的相似性,以及对生境资源需求的互补性。这也表明:群落内优势树种之间在资源共享和生态位分化方面存在积极的相互作用,有助于维持群落的稳定性和多样性[28],王如等[29]也认为:群落呈显著正相关且正联结数量较多,群落稳定性更强,物种能够共存。本研究发现:刨花润楠与其他优势树种大多有联结性,极显著联结和显著联结比为2∶1,极显著和显著联结的树种数占总优势树种数的66.7%。华东楠、红楠、紫楠与其他树种大多联结性不显著,无联结的种对数也有很大一部分,这与前述的生态位重叠程度分析结果一致。分析联结系数发现:刨花润楠与其他18个物种组成的种对中,显著正联结的数量多于χ²检验和共同出现百分率分析的结果,这一现象可能归因于刨花润楠在25个局域小生境中的集中分布和高频出现,这种分布模式增加了刨花润楠与其他物种在同一样地中的共现概率,进而提升了它们之间的联结显著度。这与叶权平等[30]得出的联结系数会扩大种对间的联结显著度的结论一致[31]。刨花润楠与栲、矩叶鼠刺、青冈、微毛柃等物种χ2检验与联结系数均不显著,但在样地调查中发现刨花润楠与其他物种共同出现的频次较高,说明尽管这些物种间未显示出明显的关联性,但它们凭借各自的独特生态习性或林分垂直分层特性,得以占据不同的生态位,从而实现物种间的共存状态。本研究发现:刨花润楠与华东楠、红楠、紫楠呈极显著正联结,原因可能是它们对特定环境条件有共同适应性,这与张滋芳等[32]认为物种间有相似的环境适应性时,更容易表现为正向联系的结果一致。4种楠木同属樟科 Lauraceae,有着相似的生活习性,彼此间联系较强,通过相互协作和依赖,共同适应并占据了群落中特定的生态位。这也是建德楠木林中4种楠木共存的理论基础[33]。
-
建德楠木林群落中的刨花润楠具有最大的重要值和较高的生态位宽度,与其他树种的生态位重叠程度均较高,种间联结性较强。华东楠、红楠、紫楠的重要值、生态位宽度和生态位重叠程度均较小,种间联结性较弱。群落总体呈现显著正联结,种间正联结占优势,树种联结性较弱,树种之间独立性较强。群落种间联结比较松散,群落演替尚未达到成熟阶段。
-
为了有效保护建德楠木林,适当间伐与楠木存在负联结的树种,如与刨花润楠呈极显著负联结的木荷、细叶香桂、青冈、甜槠;与华东楠呈极显著负联结的微毛柃;与紫楠呈负联结的拟赤杨、毛八角枫、檵木,从而有效缓解树种间的竞争矛盾,促进楠木的健康生长与生态平衡;在较大林窗处,可适当补植与楠木有显著正联结的树种,如与红楠呈显著正联结的枫香、乌楣栲、矩形叶鼠刺、毛花连蕊茶、栲等。同时需开展长期定位监测,以掌握该群落的演替过程与动态,为更好保护建德楠木林提供数据支撑。
Ecological niche and interspecific association of dominant tree species in a Machilus and Phoebe forest in Jiande, Zhejiang Province
-
摘要:
目的 建德楠木森林公园分布着亚洲现存最大的天然楠木林。分析该群落中楠木的生态位和种间关系,以期为建德楠木林的保护、种群恢复及近自然经营提供理论依据。 方法 基于2023年7月对1 hm2样地的调查数据,通过物种组成、生态位宽度、生态位重叠指数、χ2检验、方差比例法、种间联结系数和共同出现百分率等,研究包括刨花润楠Machilus pauhoi、华东楠M. leptophylla、红楠M. thunbergii、紫楠Phoebe sheareri等19种重要值≥1%的优势树种的生态位和种间联结性。 结果 ①样地中,刨花润楠重要值最大,达23.44%,为群落的建群种;华东楠、红楠、紫楠重要值为1.20%~1.80%,排在第12位以后。②样地中刨花润楠与其他优势树种有较高的生态位重叠值,其中与华东楠生态位重叠值高达0.87;华东楠、红楠、紫楠与其他树种间生态位重叠值较小,为0.10~0.50。③建德楠木林优势树种种间总体呈现显著正相关(P<0.05)。χ2检验、种间联结系数、共同出现百分率均表明:刨花润楠与18个优势树种种间关联性较强,华东楠、红楠、紫楠与优势树种种间关联性较弱。④回归分析显示:主要优势树种的种间联结系数和共同出现百分率与生态位重叠值呈极显著正相关(P<0.01)。 结论 浙江建德楠木林群落中各物种相对独立分布,种间关系较为松散,群落正处于演替中期。可采取适当择伐、林下补植等措施,实现楠木的长期稳定生长。图4表2参33 Abstract:Objective The objective is to analyze the ecological niche and interspecific relationship of the Machilus and Phoebe forest in Jiande Forest Park, which is the largest existing natural Machilus and Phoebe forest in Asia, so as to provide theoretical basis for its protection, population restoration and near-nature management. Method Based on the survey data of 1 hm2 sample plot in July 2023, the ecological niche and interspecific association of 19 dominant tree species with important values≥1% were examined, including M. pauhoi, M. leptophylla, M. thunbergii, and P. sheareri, through species composition, niche breadth, niche overlap index, χ2 test, variance ratio method, interspecific association coefficient, and co-occurrence percentage. Result (1) M. pauhoi had the highest important value (23.44%), and was the constructive species of the community. The important values of M. leptophylla, M. thunbergii, and P. sheareri were 1.20%−1.80%, ranking after the 12th place. (2) In the sample plots, there was a high niche overlap between M. pauhoi and other dominant tree species, among which the ecological niche overlap value with M. leptophylla was as high as 0.87. The niche overlap values between M. leptophylla, M. thunbergii, P. sheareri, and other tree species were relatively small, ranging from 0.10 to 0.50. (3) There was a significant positive correlation (P<0.05) among the dominant trees in Machilus and Phoebe forest in Jiande. The χ2 test, interspecific association coefficient, and percentage of co-occurrence revealed that there was a strong correlation between M. pauhoi and 18 dominant tree species, while the correlation between M. leptophylla, M. thunbergii, and P. sheareri and the dominant tree species was weak. (4) Regression analysis showed that interspecific association coefficient, and percentage of co-occurrence of the main dominant tree species were significantly positively correlated with the ecological niche overlap value (P<0.01). Conclusion The species of Machilus and Phoebe forest in Jiande are relatively independently distributed and in the middle stage of succession, with loose interspecific relationship. Appropriate measures such as selective logging and understory replanting can be taken to achieve long-term stable growth of Machilus and Phoebe trees. [Ch, 4 fig. 2 tab. 33 ref.] -
图 1 主要优势树种 Pianka 生态位重叠指数
Figure 1 Pianka niche overlap index of dominant tree species
图 2 优势植物种间联结性半矩阵图
Figure 2 Semi-matrix diagram of association coefficients of interspecific associations of dominant plants
图 3 优势植物共同出现百分率半矩阵图
Figure 3 Semi-matrix diagram of co-occurrence percentage for dominant plant species
图 4 生态位重叠值与种间联结系数、共同出现百分率的线性回归分析
Figure 4 Linear regression analysis between niche overlap and interspecific association coefficient, and percentage of co-occurrence
表 1 主要优势树种重要值及生态位宽度
Table 1. Importance values and niche width of dominant tree species
排名 植物 平均胸径/cm 平均树高/m 个体数/株 重要值/% 生态位宽度指数 Shannon Levins 1 刨花润楠 Machilus pauhoi 12.2 9.6 347 23.44 2.62 9.44 2 甜槠 Castanopsis eyrei 13.9 9.5 159 13.92 2.34 8.10 3 乌楣栲Castanopsis jucunda 10.6 8.1 34 5.30 2.56 10.80 4 栲 Castanopsis fargesii 9.0 7.5 76 4.98 2.63 10.11 5 矩形叶鼠刺 Itea oblonga 10.5 6.8 123 4.84 2.72 12.06 6 青冈 Cyclobalanopsis glauca 15.6 10.5 58 4.53 2.48 9.02 7 毛花连蕊茶 Camellia fraternal 10.6 7.6 124 4.51 2.52 9.82 8 微毛柃 Eurya hebeclados 9.9 7.3 126 4.31 2.35 9.31 9 细叶香桂 Cinnamomum subavenium 9.9 7.9 72 2.98 2.25 8.15 10 枫香 Liquidambar formosana 10.0 7.7 12 2.02 1.98 6.55 11 拟赤杨 Alniphyllum fortunei 6.8 7.2 36 1.82 1.52 3.66 12 华东楠 Machilus leptophylla 9.5 8.6 28 1.73 1.41 2.98 13 红楠 Machilus thunbergii 10.9 8.3 19 1.68 1.97 5.65 14 毛八角枫 Alangium kurzii 10.8 7.4 31 1.66 1.76 4.06 15 木荷 Schima superba 19.5 11.3 16 1.47 1.37 3.12 16 老鼠矢 Symplocos stellaris 11.1 6.9 12 1.40 2.34 9.80 17 檵木 Loropetalum chinense 6.3 6.7 9 1.31 1.89 6.37 18 厚皮香 Ternstroemia gymnanthera 7.8 7.0 12 1.30 2.14 6.76 19 紫楠 Phoebe sheareri 23.5 16.3 10 1.20 1.52 3.76  下载: 导出CSV
下载: 导出CSV
表 2 优势树种χ2统计量检验
Table 2. χ2 correlation test of dominant tree species
树种 Mp Ce Cj Cf Io Cg Cfa Eh Cc Lf Af Ml Mt Ak Ss Ssl Lc Tg Ce 1.14 Cj 2.04 2.09 Cf 0.91 0.53 4.15* Io 0.17 3.14 9.66** 9.77** Cg 0.83 0.31 0.00 0.93 0.93 Cfa 0.21 0.83 9.69** 1.50 5.47* 0.06 Eh 0.08 14.52** 2.92 1.71 5.37* 0.72 4.72* Cc 2.98 5.50* 0.09 0.01 0.81 0.09 0.59 5.03* Lf 0.83 0.12 0.95 1.39 1.39 0.00 0.06 0.00 1.32 Af 0.21 0.90 0.50 1.00 1.00 0.50 2.10 2.01 3.64 0.06 Ml 0.35 1.71 0.34 0.12 0.12 2.52 0.73 3.08 0.13 0.18 0.04 Mt 1.14 0.41 4.52* 1.83 1.83 0.31 3.51 1.50 0.02 0.12 0.00 0.41 Ak 0.00 0.41 1.52 1.83 1.83 1.52 3.51 1.50 0.02 0.12 7.65** 0.41 0.05 Ss 3.87* 2.62 0.34 0.12 0.12 2.03 0.04 1.16 0.13 0.18 0.04 1.06 0.11 0.11 Ssl 0.08 0.15 0.78 0.50 2.93 3.04 2.01 0.08 0.03 0.00 4.72* 0.02 0.21 8.83** 0.02 Lc 2.81 3.51 0.06 0.01 0.01 0.50 0.21 2.01 3.64 0.50 0.21 0.04 0.00 0.00 0.04 0.27 Tg 0.01 6.95** 2.21 0.26 0.26 2.20 1.40 5.69* 0.33 0.07 0.41 0.74 0.59 0.59 1.11 0.01 2.39 Ps 0.35 10.62** 2.52 0.12 2.23 0.34 0.73 7.28** 4.98* 0.18 1.51 0.00 0.11 2.62 1.06 1.06 1.51 3.30 说明:**为联结性极显著(P<0.01);*为联结性显著(P<0.05)。Mp. 刨花润楠;Ce. 甜槠;Cj. 乌楣栲;Cf. 栲;Io. 矩形叶鼠刺;Cg. 青冈;Cfa. 毛花连蕊茶;Eh. 微毛柃;Cc. 细叶香桂;Lf. 枫香;Af. 拟赤杨;Ml. 华东楠;Mt. 红楠;Ak. 毛八角枫;Ss. 木荷;Ssl. 老鼠矢;Lc. 檵木;Tg. 厚皮香;Ps. 紫楠。
下载: 导出CSV
-
[1] AKATOV V V, AKATOVA T V, CHEFRANOV S G, et al. Degree of dominance and species richness in plant communities with high and low intensity of interspecies competition[J]. Biology Bulletin Reviews, 2018, 8(5): 389−400. [2] VAJARI K A. Influence of interspecies competition on beech (Fagus orientalis Lipsky) trees and some features of stand in mixed broad-leaved forest[J/OL]. Environmental Monitoring & Assessment, 2018, 190 [2024-04-30]. DOI: 10.1007/s10661-018-6754-8. [3] 张璐, 苏志尧, 陈北光. 中国特有植物——华南五针松群落的种间关系[J]. 生态学报, 2006, 26(4): 1063−1072. ZHANG Lu, SU Zhiyao, CHEN Beiguang, et al. Interspecific relationships in the forest community dominated by Pinus kwangtungensis, a Chinese native species[J]. Acta Ecologica Sinica, 2006, 26(4): 1063−1072. [4] 薛卫星, 李春辉, 艾训儒, 等. 鹅掌楸天然林优势树种生态位与种间联结性[J]. 森林与环境学报, 2023, 43(1): 26−34. XUE Weixing, LI Chunhui, AI Xunru, et al. Niche and interspecific association of dominant tree species in Liriodendron chinense natural forest[J]. Journal of Forest and Environment, 2023, 43(1): 26−34. [5] 刘雨婷, 侯满福, 贺露炎, 等. 滇东菌子山喀斯特森林群落乔木优势树种生态位和种间联结[J]. 应用生态学报, 2023, 34(7): 1771−1778. LIU Yuting, HOU Manfu, HE Luyan, et al. Niche and interspecific association of dominant tree species in karst forest of Junzi Mountain, eastern Yunnan, China[J]. Chinese Journal of Applied Ecology, 2023, 34(7): 1771−1778. [6] 商侃侃. 浙江天目山孑遗植物群落主要种群的种间关系[J]. 浙江农林大学学报, 2013, 30(2): 206−214. SHANG Kankan. Interspecific relationship of dominant populations in relict deciduous forests on Mount Tianmu, eastern China[J]. Journal of Zhejiang A&F University, 2013, 30(2): 206−214. [7] 刘润红, 陈乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J]. 生态学报, 2020, 40(6): 2057−2071 LIU Runhong, CHEN Le, TU Hongrun, et al. Ecological study of Cyclobalanopsis glauca communityon the karst hills of Guilin[J]. Acta Ecologica Sinica, 2020, 40(6): 2057−2071 [8] LEVINE J M, HILLERISLAMBERS J. The importance of niches for the maintenance of species diversity[J]. Nature, 2009, 461(7261): 254−257. [9] 方庆, 谭菊荣, 许惠春, 等. 珍稀濒危植物细果秤锤树群落物种组成与生态位分析[J]. 浙江农林大学学报, 2022, 39(5): 931−939. FANG Qing, TAN Jurong, XU Huichun et al. Species composition and niche of Sinojackia microcarpa, a rare and endangered plant[J]. Journal of Zhejiang A&F University, 2022, 39(5): 931−939. [10] 孙杰杰, 江波, 吴初平, 等. 浙江檫木群落生境特征及区划研究[J]. 生态学报, 2019, 39(3): 884−894. SUN Jiejie, JIANG Bo, WU Chuping, et al. Study on the habitat and niche of Sassafras tzumu (Hemsl. ) Hemsl[J]. Acta Ecologica Sinica, 2019, 39(3): 884−894. [11] 任晴. 珍贵树种红楠适生区预测及生境地群落特征研究[D]. 杭州: 浙江农林大学, 2020. REN Qing. Research on Suitable Habitat and Community Characteristics of Precious Tree Machilus thunbergii[D]. Hangzhou: Zhejiang A&F University, 2020. [12] 李冰. 氮添加对毛竹和浙江楠生理生长特征及种间关系的影响[D]. 杭州: 浙江农林大学, 2022. LI Bing. Effects of Nitrogen Addition on Physiological Growth Characteristics and Interspecific Relationship of Phyllostachys edulis and Phoebe chekiangensis[D]. Hangzhou: Zhejiang A&F University, 2022. [13] 李铁华, 李阳宁, 闫旭, 等. 闽楠-木荷混交林空间结构及其养分利用[J]. 中南林业科技大学学报, 2024, 44(2): 1−9. LI Tiehua, LI Yangning, YAN Xu, et al. Spatial structure and nutrient utilization of mixed forest of Phoebe bournei and Schima superba[J]. Journal of Central South University of Forestry and Technology, 2024, 44(2): 1−9. [14] 何贵平, 骆文坚, 冯建民, 等. 不同立地条件刨花楠天然群落物种多样性比较研究[J]. 浙江林业科技, 2003, 23(5): 2−3, 13. HE Guiping, LUO Wenjian, FENG Jianmin, et al. Comparison study on diversity of natural Machilus pauhoi community under different site conditions[J]. Journal of Zhejiang Forestry Science and Technology, 2003, 23(5): 2−3, 13. [15] 吴初平, 袁位高, 盛卫星, 等. 浙江省典型天然次生林主要树种空间分布格局及其关联性[J]. 生态学报, 2018, 38(2): 537−539. WU Chuping, YUAN Weigao, SHENG Weixing, et al. Spatial distribution patterns and associations of tree species in typical natural secondary forest communities in Zhejiang Province[J]. Acta Ecologica Sinica, 2018, 38(2): 537−539. [16] 吴丹婷, 吴初平, 盛卫星, 等. 浙江建德楠木天然林群落种间联结动态[J]. 浙江农林大学学报, 2021, 38(4): 671−681. WU Danting, WU Chuping, SHENG Weixing, et al. Interspecific association dynamics of Nanmu natural forest in Jiande, Zhejiang Province[J]. Journal of Zhejiang A&F University, 2021, 38(4): 671−681. [17] 马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究 (Ⅱ) 丰富度、均匀度和物种多样性指数[J]. 生态学报, 1995, 15(3): 286−277. MA Keping, HUANG Jianhui, YU Shunli, et al. Plant community diversity in Dongling Mountain, Beijing, China (Ⅱ) species richness, evenness, and species diversities[J]. Acta Ecologica Sinica, 1995, 15(3): 286−277. [18] PIANKA R E. The structure of lizard communities[J]. Annual Review of Ecology, Evolution and Systematics, 1973, 4(1): 53−74. [19] SCHLUTER D. A variance test for detecting species associations, with some example applications[J]. Ecology, 1984, 65(3): 998−1005. [20] 江常春, 梁希妮, 田磊, 等. 浙江省常绿阔叶林优势树种种间联结分析[J]. 浙江师范大学学报(自然科学版), 2022, 45(4): 437−445. JIANG Changchun, LIANG Xini, TIAN Lei, et al. Analysis on interspecific association of dominant species in evergreen broad-leaved forests in Zhejiang Province[J]. Journal of Zhejiang Normal University (Natural Sciences), 2022, 45(4): 437−445. [21] 郭志华, 卓正大, 陈洁, 等. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J]. 植物生态学报, 1997, 21(5): 424−432. GUO Zhihua, ZHUO Zhengda, CHEN Jie, et al. Interspecific association of trees in mixed evergreen and deciduous broadleaved forest in Lushan Mountain[J]. Acta Phytoecologica Sinica, 1997, 21(5): 424−432. [22] 陈林, 辛佳宁, 苏莹, 等. 异质生境对荒漠草原植物群落组成和种群生态位的影响[J]. 生态学报, 2019, 39(17): 6187−6205. CHEN Lin, XIN Jianing, SU Ying, et al. Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China[J]. Acta Ecologica Sinica, 2019, 39(17): 6187−6205. [23] 李德志, 石强, 臧润国, 等. 物种或种群生态位宽度与生态位重叠的计测模型[J]. 林业科学, 2006, 42(7): 95−103. LI Dezhi, SHI Qiang, ZANG Runguo, et al. Models for niche breadth and niche overlap of species or populations[J]. Scientia Silvae Sinicae, 2006, 42(7): 95−103. [24] 叶兴状, 王妙青, 程诺, 等. 福建天台山半枫荷天然群落的物种组成生态位和种间关系[J]. 植物资源与环境学报, 2021, 30(6): 19−28. YE Xingzhuang, WANG Miaoqing, CHENG Nuo, et al. Species composition, niche, and interspecific relationships of Semiliquidambar cathayensis natural community in Tiantai Mountain of Fujian Province[J]. Journal of Plant Resources and Environment, 2021, 30(6): 19−28. [25] 胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报, 2009, 29(7): 3670−3677. HU Zhenghua, QIAN Haiyuan, YU Mingjian. The niche of dominant species populations in Castanopsis eyrei forest in Gutian Mountain National Nature Reserve[J]. Acta Ecologica Sinica, 2009, 29(7): 3670−3677. [26] 马丰丰, 潘高, 李锡泉, 等. 桂西南喀斯特山地木本植物群落种间关系及CCA排序[J]. 北京林业大学学报, 2017, 39 (6): 32−44. MA Fengfeng, PAN Gao, LI Xiquan, et al. Interspecific relationship and canonical correspondence analysis within woody plant communities in the karst mountains of southwest Guangxi, southern China[J]. Journal of Beijing Forestry University, 2017, 39 (6): 32−44. [27] 高浩杰, 袁佳, 高平仕. 舟山群岛次生林优势种群的生态位与种间联结[J]. 热带亚热带植物学报, 2017, 25(1): 73−80 GAO Haojie, YUAN Jia, GAO Pingshi. Niche and interspecific association of dominant species in secondary forest of Zhoushan Islands[J]. Journal of Tropical and Subtropical Botany, 2017, 25(1): 73−80. [28] 何江, 徐来仙, 艾训儒. 星斗山天然次生林优势种的生态位和种间联结动态[J]. 林业科学研究, 2024, 37(1): 171−182. HE Jiang, XU Laixian, AI Xunru. Dynamics in niche and interspecific association of dominant species in natural secondary forest of Xingdou Mountain[J]. Forest Research, 2024, 37(1): 171−182. [29] 王如, 彭文成, 杨佳, 等. 尖峰岭杉木人工林群落主要木本植物的生态位和种间联结[J]. 应用生态学报, 2024, 35(6): 325−333. WANG Ru, PENG Wencheng, YANG Jia, et al. Niche and interspecific association of main woody plants in Cunninghamia lanceolata plantation community in Jianfengling, China[J]. Chinese Journal of Applied Ecology, 2024, 35(6): 325−333. [30] 叶权平, 张文辉, 于世川, 等. 桥山林区麻栎群落主要乔木种群的种间联结性[J]. 生态学报, 2018, 38(9): 3165−3174. YE Quanping, ZHANG Wenhui, YU Shichuan, et al. Interspecific association of the main tree populations of the Quercus acutissima community in the Qiaoshan forest area[J]. Acta Ecologica Sinica, 2018, 38(9): 3165−3174. [31] 李锦婷, 穆君, 申开平, 等. 小黄花茶群落优势木本植物生态位及种间联结性[J]. 生态学报, 2024, 44(1): 283−294. LI Jinting, MU Jun, SHEN Kaiping, et al. Niche and interspecific association of dominant woody plants in Camellia luteoflora community[J]. Acta Ecologica Sinica, 2024, 44(1): 283−294. [32] 张滋芳, 毕润成, 张钦弟, 等. 珍稀濒危植物矮牡丹生存群落优势种种间联结性及群落稳定性[J]. 应用与环境生物学报, 2019, 25(2): 291−299. ZHANG Zifang, BI Runcheng, ZHANG Qindi, et al. Community stability and inter-specific associations between the rare plant Paeonia suffruticosa Andr. var. spontanea Rehd and dominant species[J]. Chinese Journal of Applied and Environmental Biology, 2019, 25(2): 291−299. [33] 张珂, 廖绍波, 孙冰, 等. 广东3地典型彩叶树种岭南槭所在天然群落特征比较[J]. 浙江农林大学学报, 2018, 35(1): 10−19. ZHANG Ke, LIAO Shaobo, SUN Bing, et al. Comparison of natural community characteristics of a typical colored tree species, Acer tutcheri at three sites in Guangdong[J]. Journal of Zhejiang A&F University, 2018, 35(1): 10−19. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240378
 点击查看大图
点击查看大图
计量
- 文章访问数: 1461
- HTML全文浏览量: 452
- PDF下载量: 32
- 被引次数: 0
