






-
海岛生态系统具有明显的独立性与资源短缺性,属于典型的脆弱型生态系统,在独特的自身环境与复杂的干扰下,表现出易受损性和难恢复性[1]。目前,海岛植物的研究集中在植物恢复方面。如WANG等[2]基于植物功能性状的筛选方法,对海岛植物恢复进行植物选择。石娇星[3]对舟山群岛物种进行统计和植物制图,为海岛植物恢复提供了资源信息。俞意等[4]对珠海市海岛林地植物恢复技术及效果进行研究,认为恢复海岛土壤基质是解决海岛植物恢复的关键。关于海岛植物群落结构和物种组成及其与土壤相关因子的研究较少,认识土壤因子变化与海岛植物群落结构和物种组成的内在联系,对于阐明海岛植物退化及修复机制具有重要的生态学意义。
群落结构是认识群落组成、变化及发展趋势的基础,能够反映群落对环境的适应,反映群落的动态和机能,并能影响生态系统的功能特征[5]。群落的垂直结构和径级结构是群落结构中的重要特征[6]。陈材等[7]在研究雷州半岛时发现:随着幼树的生长及分化,灌丛面积随之增长,粗糠柴Mallotus philippensis、九节Psychotria rubra、皂帽花Dasymaschalon trichophorum等物种的径级结构呈现倒“J”型分布。金乐薇等[8]对上海大金山岛研究时发现:随着演替的进行和植物的发育,森林结构逐渐完善,不同演替阶段径级分布模式不同。张川英等[9]研究发现:遂昌乌溪江流域山蜡梅Chimonanthus nitens种群和生境群落径级结构均呈倒“J”型,为增长型结构。因此,研究群落径级结构及其更新机制,对阐明种群生态特性、更新对策及群落的形成和稳定性等具有重要意义。
关于环境因子对海岛群落物种多样性的影响受到越来越多的关注,与台州地区陆地海岸线相比,海岛植物群落物种多样性还受岛屿面积[10]、陆地海岸线距离[11]、干旱[12]和台风等的影响。土壤因子对海岛植物物种组成变化的影响已有较多研究。已有研究表明:土壤含水率、pH、全磷和全钾是影响海岛植物群落变化的主要环境因子[13]。闫潍虹[14]研究发现:土壤有效钾、速效磷是影响海岛植物多样性的重要因素。与其他具有人类开发活动的海岛类似,大陈岛受到人为的影响显著,岛上种植的大量木麻黄Casuarina equisetifolia成为大陈岛重要的植物类型,但是人为影响与土壤因子和植物组成的变化是否存在紧密联系尚不清楚。鉴于此,本研究选择大陈岛典型植物群落为研究对象,对大陈岛植物群落结构、物种组成及其与土壤因子的关系进行研究,以期为大陈岛海岛植物的维护与管理提供科学依据。
-
大陈岛位于中国浙江省台州湾东南海域,是台州列岛106个岛中的主岛,总面积近12 km2。它由上大陈、下大陈、竹屿、洋岐、猪腰屿、屏风山等29个岛(礁)组成。地形呈东北西南走向,地势起伏不平,以丘陵地形为主。大陈岛属中亚热带季风气候,年平均气温为16.7 ℃,年平均降水量为1 378.5 mm,年平均风速为6.8 m·s−1。植被是以柃木Eurya japonica、天仙果Ficus erecta为优势物种的常绿阔叶林和针阔混交林群落,同时以木麻黄为优势物种的人工栽培经过长期自然演化的针叶林群落。
-
2023年9—12月在大陈岛上设置15个20 m×20 m的样方(分别为S01~S15)。按顺序将每个20 m×20 m样方划分为16个5 m×5 m的小样方。依据方精云等[15]的研究方法,标定样方中所有胸径(DBH)≥1 cm的木本植物个体,记录每株植物的名称、胸径和高度,同时记录每个样地环境因子,包括岩石裸露率、土壤深度、凋落物盖度、凋落物厚度、腐殖质层厚度和土壤电导率。在每个20 m×20 m的样方中随机选取3个监测点,用环刀取土壤,根据《森林土壤水分物理性质的测定》方法测定pH、有机碳、全碳、全氮、全磷、硝态氨、铵态氮和有效磷等土壤理化性质。
-
以数量、胸径和高度为指标,计算每个群落中物种的重要值,同时计算Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数[16]。采用Jaccard相似性指数分析不同群落间物种组成的相似性[17]。采用SPSS 27.0进行独立样本t检验,并利用R 4.3.3软件进行冗余分析(RDA)。采用Origin 2024进行绘图。
-
大陈岛共记录植物34种,隶属于26科32属。在科水平上,山茶科Theaceae出现频次最高,占总科数的44.0%;其次是桑科Moraceae和木麻黄科Casuarinaceae,分别占总科数的19.5%和10.0%。在属水平上,柃属Eurya出现频次最高,占总属数的44.0%;其次是榕属Ficus和木麻黄属Casuarina,分别占总属数的13.8%和10.0%。从表1可见:柃木、木麻黄和天仙果为大陈岛的3个主要优势物种,重要值之和为62.70%。从表2可见:以木麻黄-柃木为优势物种的群落Margalef丰富度指数最高(2.56),以木麻黄-海州常山Clerodendrum trichotomum为优势物种的群落Margalef丰富度指数最低(1.04)。以柃木-天仙果为优势物种的群落Simpson优势度指数最高(0.94),以木麻黄-海州常山为优势物种的群落Simpson优势度指数最低(0.55)。以木麻黄-柃木为优势物种的群落Shannon-Wiener多样性指数最高(1.91),以柃木-天仙果为优势物种的群落Shannon-Wiener多样性指数最低(0.70)。以海州常山-朴树Celtis sinensis为优势物种的群落Pielou均匀度指数最高(0.82),以柃木-天仙果为优势物种的群落Pielou均匀度指数最低(0.27)。
表 1 大陈岛群落物种重要性
Table 1. Importance of community species in Dachen Island
物种 个体数/
株平均胸
径/cm平均树
高/m物种重
要值/%柃木Eurya japonica 1 318 5.28 4.54 35.91 木麻黄Casuarina equisetifolia 299 8.93 6.54 17.85 天仙果Ficus erecta 414 3.16 3.46 8.94 樟Camphora officinarum 21 25.45 10.98 4.46 海州常山Clerodendrum
trichotomum181 3.57 4.28 4.36 海岛桑Morus maritima 141 4.52 4.56 4.33 朴树Celtis sinensis 51 14.13 6.51 4.10 野桐Mallotus japonicus 138 4.42 4.45 3.81 女贞Ligustrum lucidum 37 8.64 9.08 3.51 表 2 大陈岛群落物种多样性
Table 2. Community species diversity on Dachen Island
样方号 群落 Margalef
丰富度
指数Simpson
优势度
指数Shannon-
Wiener
多样性指数Pielou
均匀度
指数样方号 群落 Margalef
丰富度
指数Simpson
优势度
指数Shannon-
Wiener
多样性指数Pielou
均匀度
指数S01 木麻黄 1.33 0.57 1.47 0.71 S09 柃木-天仙果 1.73 0.78 1.06 0.44 S02 木麻黄-野桐 1.11 0.62 1.32 0.68 S10 柃木-天仙果 2.02 0.94 0.70 0.27 S03 木麻黄-海州常山 1.04 0.74 0.98 0.55 S11 柃木-樟 1.23 0.81 0.99 0.51 S04 天仙果 1.51 0.61 1.44 0.65 S12 柃木-野桐 1.76 0.68 1.40 0.61 S05 木麻黄-天仙果 1.21 0.74 1.02 0.57 S13 海州常山-朴树 1.62 0.56 1.71 0.82 S06 木麻黄-海州常山 1.72 0.55 1.78 0.81 S14 柃木-海州常山 2.01 0.67 1.58 0.62 S07 柃木-黄檀 1.75 0.77 1.21 0.51 S15 柃木-天仙果 2.37 0.76 1.39 0.53 S08 木麻黄-柃木 2.56 0.58 1.91 0.77 -
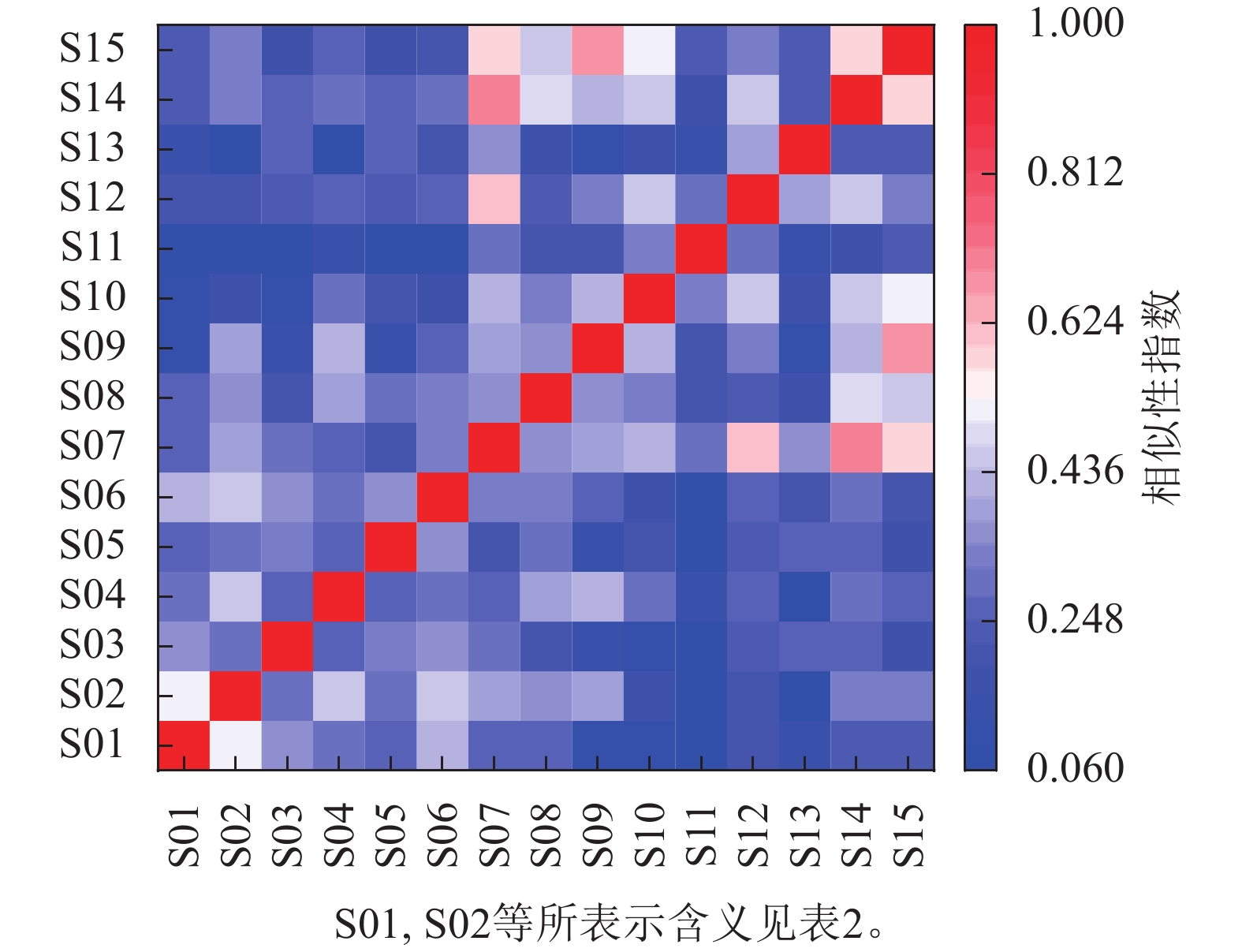
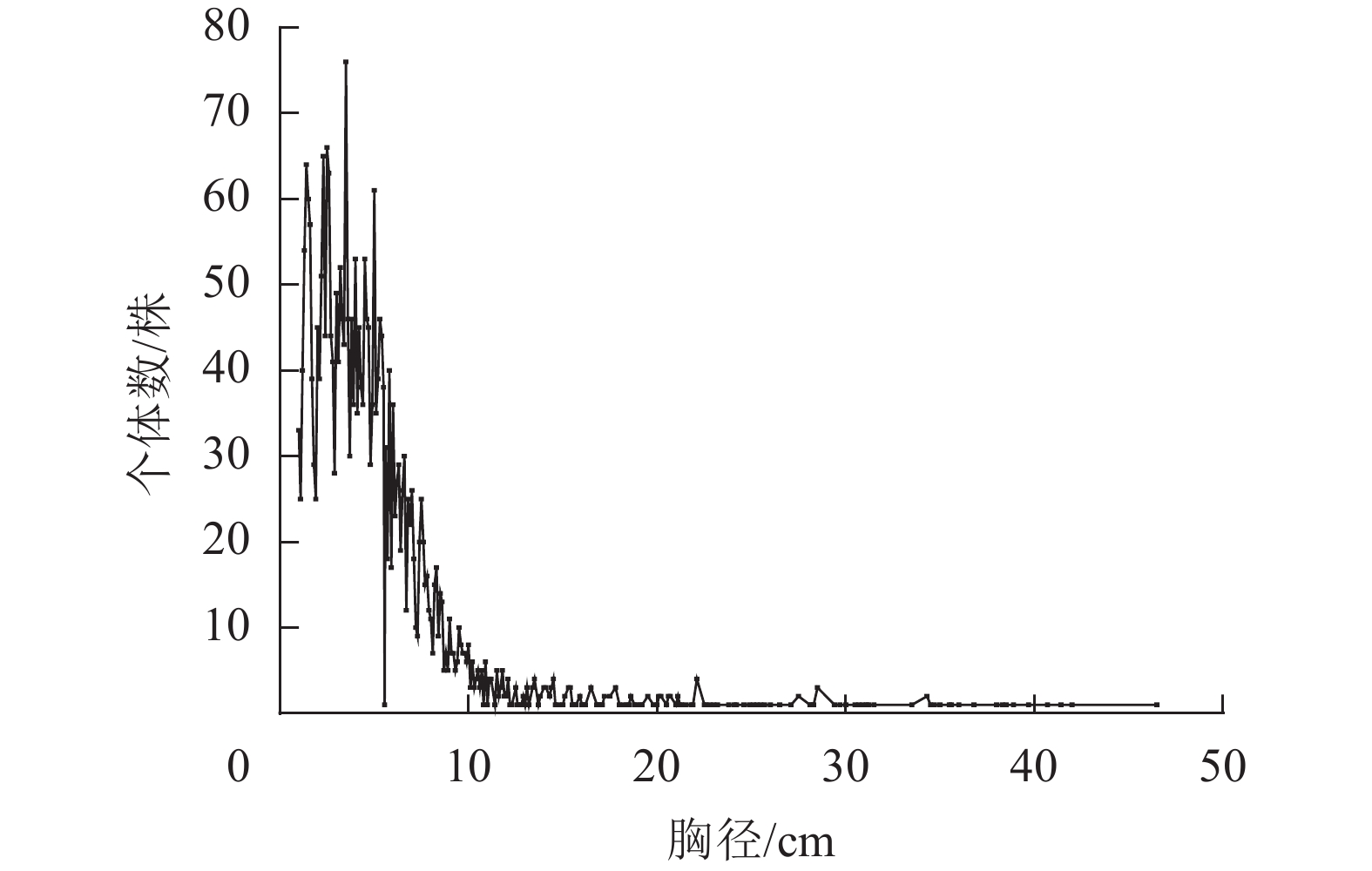
如图1所示:大陈岛乔木物种总径级结构呈倒“J”型分布,1 cm≤DBH<5 cm的植物有1 851株,占总个体数的61.78%,5 cm≤DBH<20 cm的植物有1 069株,占总个体数的35.68%,DBH≥20 cm的植物有76株,占总个体数的2.54%。
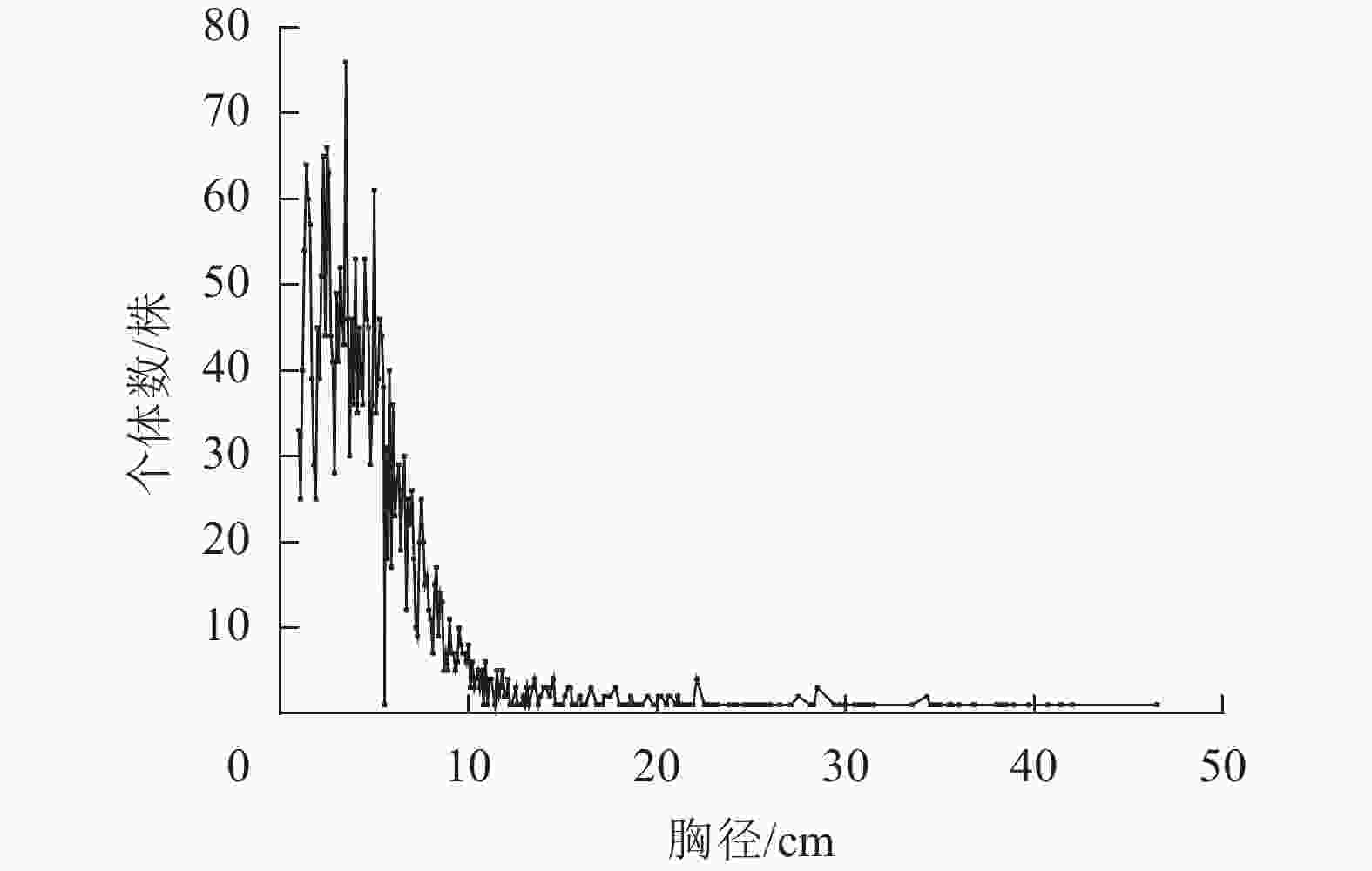
图 1 大陈岛样地乔木径级分布
Figure 1. Distribution of tree diameter classes in Dachen Island sample plots
大陈岛群落中,柃木、木麻黄和天仙果是3个重要的优势种,植株数量分别占总个体数的43.99%、13.82%和9.98%。对大陈岛3个重要的优势种进行径级结构(图2)分析发现:柃木径级结构主要呈现偏正态分布,个体数集中在1 cm≤DBH≤10 cm内;天仙果径级结构同样呈现偏正态分布,个体数集中在1 cm≤DBH≤5 cm内;木麻黄径级结构主要呈现倒“J”型分布,个体数在1 cm≤DBH≤5 cm内居多,DBH>5 cm的个体数径级分布比较均匀。
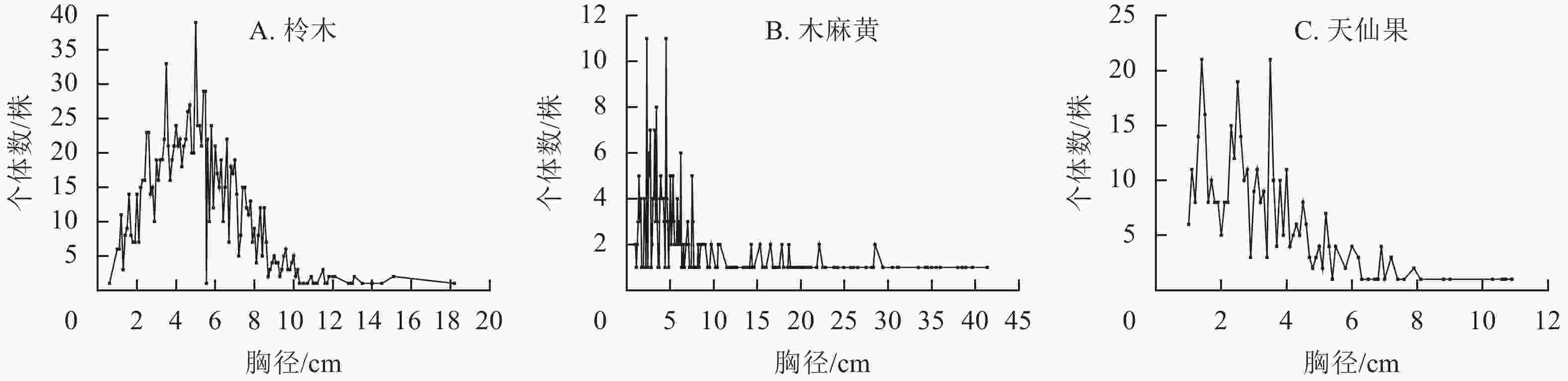
图 2 大陈岛样地优势种径级分布
Figure 2. Diameter class distribution of dominant species in Dachen Island sample plots
-
如图3所示:以柃木-黄檀为优势物种的群落与以柃木-海州常山为优势物种的群落之间Jaccard相似性指数最高,为0.71。以天仙果为优势物种的群落和以海州常山-朴树为优势物种的群落之间Jaccard相似性指数最低,为0.06。多数群落间Jaccard相似性指数小于0.50。群落间Jaccard相似性指数与全磷相似性指数呈负相关,其他土壤环境因子与Jaccard相似性指数均没有显著的相关性。
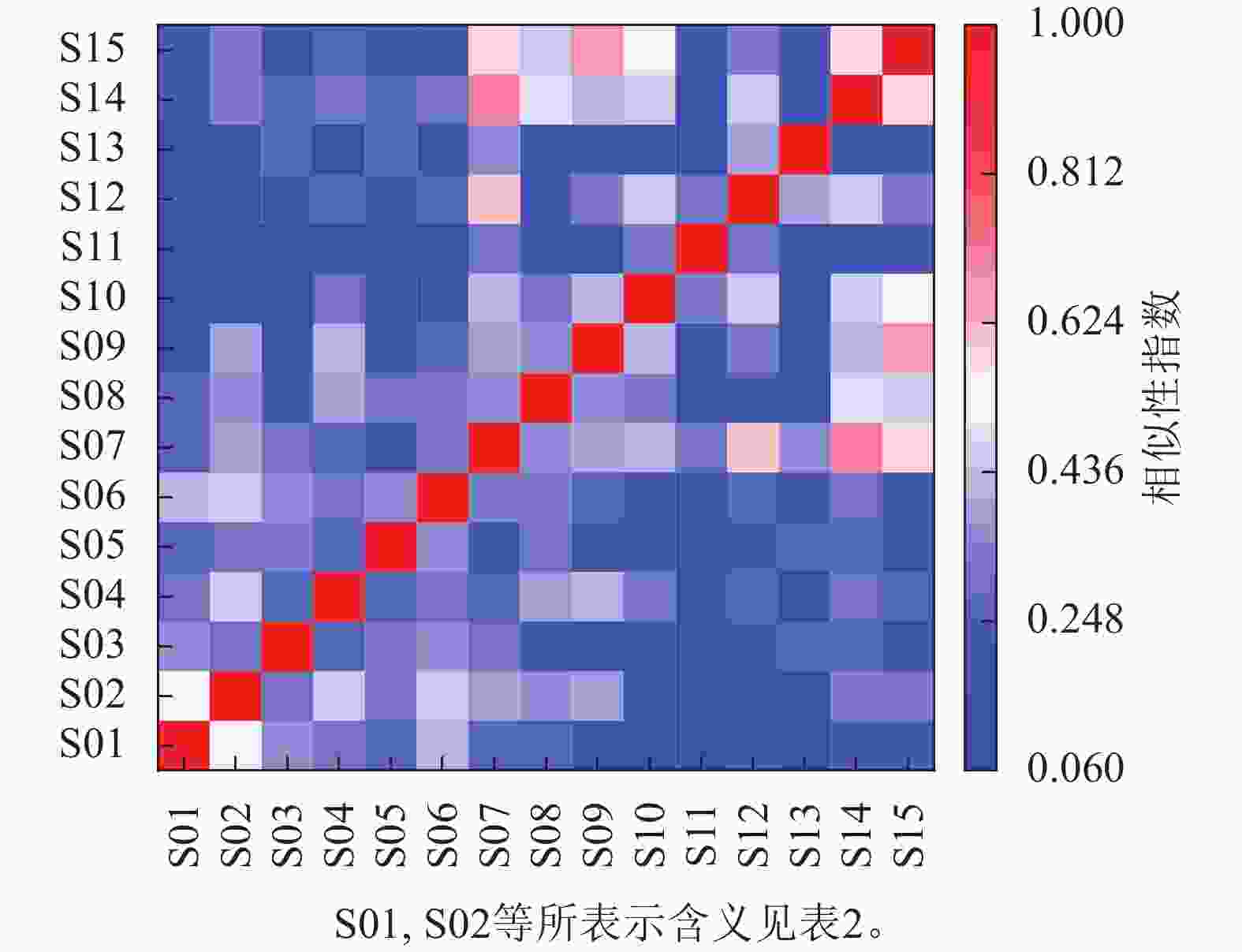
图 3 大陈岛植物群落相似性分析矩阵图
Figure 3. Species similarity analysis matrix of plant community on Dachen Island
-
大陈岛群落物种多样性与环境因子的冗余分析(图4)结果显示:排序轴1解释了总变量的69.35%,主要反映了全磷和全氮的影响,排序轴2解释了总变量的30.35%,主要反映了硝态氮、铵态氮、有效磷和全磷的影响。大陈岛群落的Simpson优势度指数与pH、有机碳、全碳、全氮呈正相关关系。Shannon-Wiener多样性指数与硝态氮、铵态氮和有效磷呈正相关关系,与有机碳、全碳和全氮均呈负相关关系。Pielou均匀度指数与硝态氮、铵态氮、有效磷和全磷呈正相关关系,与pH呈负相关关系。Margalef丰富度指数与pH呈正相关关系,与硝态氮、铵态氮、有效磷和全磷呈负相关关系。
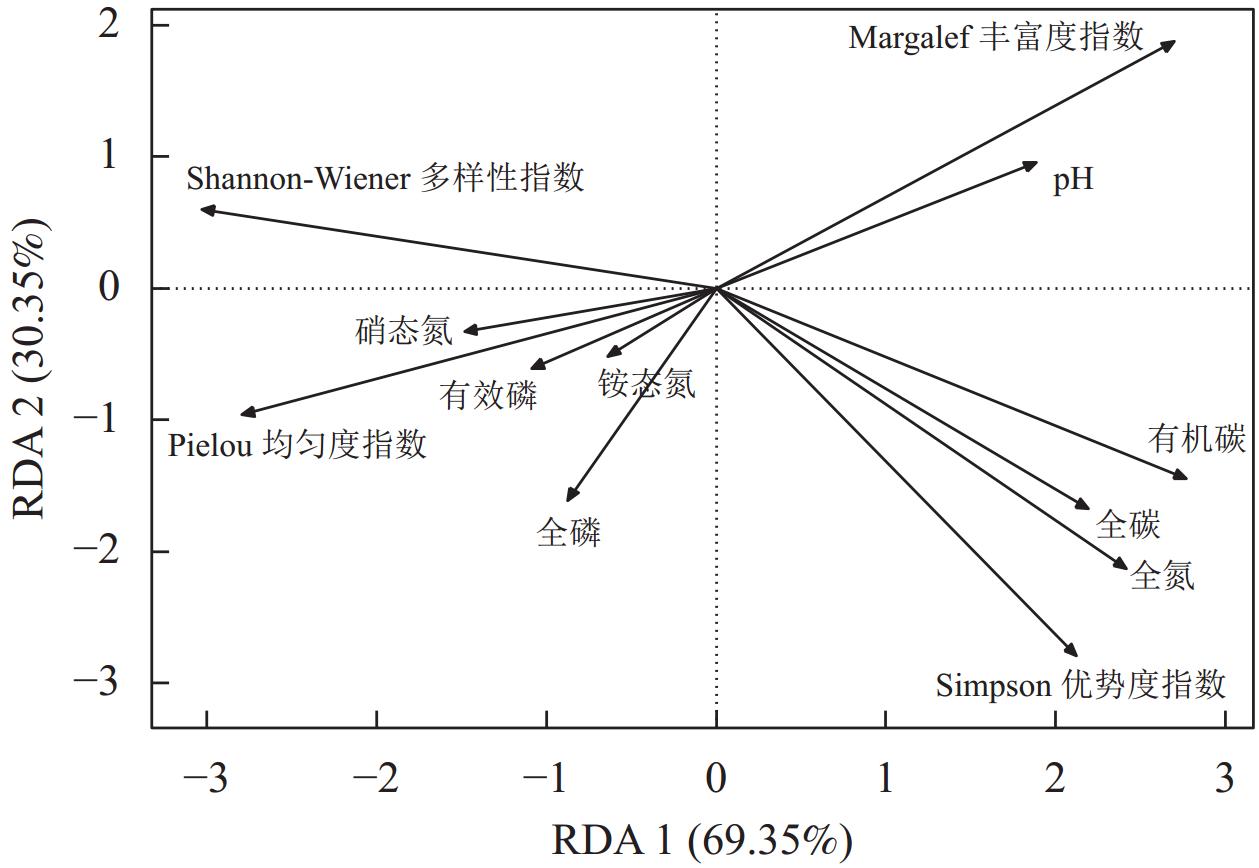
图 4 大陈岛群落物种多样性与环境因子的RDA排序
Figure 4. RDA ranking of species diversity and environmental factors in Dachen Island sample plots
-
物种组成是决定群落性质的重要因素,也是了解生态系统维持机制的重要途径[18]。大陈岛群落共记录海岛植物26科32属34种,以针叶林和阔叶林为主,森林覆盖率达56%,以山茶科、桑科、木麻黄科、唇形科Lamiaceae和大戟科Euphorbiaceae为优势科。柃木、木麻黄和天仙果是大陈岛群落的主要优势物种。柃木适宜生长在喜湿、耐水的海岛群落环境中。高大海等[19]研究发现:柃木主要分布在浙江台州、舟山等沿海岛屿,且生长在阴湿处,是海岛群落的主要优势物种。郑俊鸣等[20]对外马廊山岛进行研究时也发现柃木为优势物种。与柃木不同,木麻黄成为大陈岛植物群落的优势物种与长期人工引种存在紧密的关系,人工群落逸散改变了自然群落的物种组成。大陈岛群落中,以木麻黄-柃木为优势物种的群落和以木麻黄-海州常山为优势物种的群落Shannon-Wiener多样性指数最高,而且以木麻黄-海州常山为优势物种的群落Pielou均匀度指数最高。木麻黄为针叶树种,冠层透光较好,导致林下灌草植物生长旺盛,这与王芳等[21]在平潭岛的研究结果相似。本研究发现:以柃木-天仙果为优势物种的群落Shannon-Wiener多样性指数最低,Pielou均匀度指数也最低,这可能由于柃木和天仙果属于小乔木,冠层密闭,林下光照较少,林下植物被明显抑制,这与李应等[22]的研究结果相似。以柃木-天仙果或者柃木-樟为优势物种的群落Simpson优势度指数较高,但是以木麻黄-海州常山或者海州常山-朴树为优势物种的群落Simpson优势度指数较低。这主要由于柃木、天仙果、樟等优势物种形成密闭冠层,导致非优势物种受到强烈抑制,优势度指数则较高,木麻黄、海州常山、朴树等优势物种冠层较为稀疏,非优势物种得到一定的生长空间,优势度指数则较低,这与管诗敏等[23]在平潭岛的研究结果一致。本研究还发现:木麻黄为优势物种的群落Margalef丰富度指数较低,这与大陈岛以木麻黄为优势种的群落处于演替后期,物种较少有关,而柃木、天仙果等矮小乔木为优势物种的群落多处于演替中期,物种数量明显较多。金山岛植物群落在演替中期种群呈现间歇性发展格局,伴生、偶见种较多,群落物种多样性同样较为丰富[24],而演替后期青冈Quercus glauca、天竺桂Cinnamomum japonicum、红楠Machilus thunbergii为优势物种,许多物种被竞争抑制,导致物种多样性降低[25]。本研究结果表明:大陈岛植物群落主要分为2类:①柃木、天仙果为优势物种的原生植物群落,其优势物种具有绝对的竞争力,群落物种多样性和均匀度指数均较低;②以木麻黄为优势物种的植物群落,其优势物种竞争力相对较弱,群落多样性指数和均匀度指数相对较高。因此,海岛防护和生态林人工种植,对于海岛的植物类型及群落结构可能产生重要的影响。可见,木麻黄人工林种植对于海岛植物多样性维持具有一定的积极作用。
-
大陈岛植物群落所有物种的径级结构呈现倒“J”型分布,表明大陈岛群落幼株个体数较多,树种更新能力较强,但是同时反映了大陈岛群落物种对光照、土壤、水分等资源的竞争强度相对较高,导致少数个体存活并发育成大径级个体,大多数植株在竞争过程中被淘汰。同样,雷州半岛[7]和舟山群岛[26]植物群落的优势物种径级结构均呈倒“J”型分布。但是天仙果和柃木优势物种呈现偏正态分布,是由于这2个物种属于小乔木,受到其他大乔木抑制,因此大径级个体数相对较少,这与大汉三墩岛和蜈支洲岛植物优势种径级分布的结果相同[13]。木麻黄径级呈现倒“J”型分布,且个体数在中大径级分布较为均匀。木麻黄是大陈岛主要人工引入的生态防护物种,该结果表明木麻黄比较适应大陈岛生长环境,更新良好,在大陈岛今后的植物演替中可能起到重要的作用。
-
相似性分析结果显示:以柃木-黄檀为优势物种的群落和以柃木-海州常山为优势物种的群落之间Jaccard相似性指数最高。柃木为2类群落之间共同优势种,群落植物生长环境相似,柃木形成的密闭冠层导致2类群落共有较多的林下喜荫物种;以天仙果为优势物种的群落和以海州常山-朴树为优势物种的群落之间Jaccard相似性指数最低。天仙果喜排水良好的沙壤土,而海州常山和朴树喜荫,适宜较湿润土壤,群落生境不同可能导致其群落物种间存在较大差异。群落的Jaccard相似性指数跟土壤全磷呈显著的负相关,表明土壤全磷质量分数与群落物种组成存在紧密的关系。同样在涠洲岛群落土壤中,磷限制更为明显,磷素的增加会极大地提高海岛植物的生态适应性[13]。因此,磷可能是海岛植物对土壤生态适应的重要指示因子,磷的变化对群落内物种组成可能产生重要的影响。
-
Shannon-Wiener多样性指数、Pielou均匀度指数与硝态氮、铵态氮和有效磷呈正相关关系。何雅琴等[27]在福建平潭大练岛同样发现:土壤中氮磷的提高有助于提升植物群落的多样性。海岛由于常年高温多雨,且土壤遭风化严重,养分大量流失,土壤氮磷严重缺乏[28],有效性增加氮磷有助于促进植物生长,提升群落物种多样性。有机碳和全碳增加导致土壤碳氮比增加,已有研究表明:土壤有机碳等增加会促进岛屿优势物种生长[29],本研究同样发现了此现象。Margalef丰富度指数与硝态氮、铵态氮、有效磷和全磷呈负相关,这可能与大陈岛人工植被和原生植被的不同特点有关。以木麻黄为优势物种的人工植物群落由于营造时外源添加养分,导致土壤氮磷丰富,群落物种丰富度不高,但是柃木、天仙果等天然植物群落由于土壤氮磷增加,生长受到促进,导致该类植物Shannon-Wiener多样性指数和Pielou均匀度指数较高,Margalef丰富度指数反而较低;而以柃木、天仙果为优势物种的原生植被,优势物种优势度明显,但是物种数量较多,多数物种处于小树状态,导致群落Shannon-Wiener多样性指数和Pielou均匀度指数较低,物种数量相对较多。大陈岛土壤pH平均为6.2,呈微酸性,已有研究发现:酸性土壤pH升高会促进优势物种生长[30],进而导致均匀度指数降低;pH与Margalef丰富度指数呈正相关,与Pielou均匀度指数呈负相关。本研究表明:除了木麻黄优势物种自身特征影响外,人工种植及添加外源氮磷也是群落多样性指数和均匀度指数增加的原因。而土壤氮磷元素缺乏仍是大陈岛等海岛原生植物的生长常态,增加土壤氮磷可能会促进原生植物群落中竞争力低下的物种生长,提高群落多样性指数和均匀度指数。
-
大陈岛群落物种以柃木、木麻黄和天仙果为主要优势种,群落物种径级主要呈倒“J”型分布,土壤全磷与群落物种相似性存在紧密的关系,同时土壤硝态氮、铵态氮和有效磷的增加与群落物种多样性提高具有一定联系。
Community structure and species composition in Dachen Island
-
摘要:
目的 研究大陈岛植物群落结构、物种组成及其与土壤因子的关系,为大陈岛海岛植物维护与管理提供科学依据。 方法 基于大陈岛15个20 m×20 m的海岛植物监测样地,结合冗余分析(RDA)等方法,对群落物种组成、径级结构、群落相似性及其环境因子进行研究。 结果 大陈岛群落共记录木本植物34种,隶属26科32属。物种个体径级分布总体呈倒“J”型。以木麻黄Casuarina equisetifolia为优势物种的群落多样性指数较高,以柃木Eurya japonica、天仙果Ficus erecta为优势物种的群落多样性指数较低。以柃木-黄檀Dalbergia hupeana为优势物种的群落和以柃木-海州常山Clerodendrum trichotomum为优势物种的群落之间相似度最高,以天仙果为优势物种的群落和以海州常山-朴树Celtis sinensis为优势物种的群落之间相似度最低,而且土壤全磷质量分数与群落相似性系数呈负相关。土壤硝态氮、铵态氮和有效磷质量分数与群落物种多样性呈现正相关。 结论 大陈岛植物群落主要以木麻黄、柃木、天仙果为优势物种,土壤全磷质量分数与群落的物种组成存在紧密的联系。图4表2参30 Abstract:Objective This study, with an exploration of the structure and species composition of the plant community of Dachen Island and its relationships with soil factors, is aimed to provide a scientific basis for its maintenance and management. Method Based on the investigation of 15 plots (20 m × 20 m) on Dachen Island and combining with the method of Redundancy Analysis (RDA), a study was conducted of its species composition, diameter structure, community similarity and environmental factors. Result The Dachen Island community boasts a record of 34 species of woody plants, which belong to 26 families and 32 genera. The size-class distribution of species individuals was generally inverse J-shaped. The communities dominated by Casuarina equisetifolia had higher value of diversity indexes, while those dominated by Eurya japonica and Ficus erecta had lower value diversity indexes. The communities dominated by E. japonica and Dalbergia odorifera and those dominated by E. japonica and Clerodendrum trichotomum had the highest similarity coefficient, while the community dominated by F. erecta and those dominated by Clerodendrum trichotomum and Celtis sinensis had the lowest similarity coefficient. Furthermore, there was a negative correlation between soil total phosphorus content and community similarity coefficient whereas soil nitrate nitrogen, ammonium nitrogen, and available phosphorus showed a positive correlation with species diversity of communities. Conclusion The communities in Dachen Island are primarily dominated by Casuarina equisetifolia, E. japonica and F. erecta, and the soil total phosphorus had a close link with species composition. [Ch, 4 fig. 2 tab. 30 ref.] -
图 1 大陈岛样地乔木径级分布
Figure 1 Distribution of tree diameter classes in Dachen Island sample plots
图 2 大陈岛样地优势种径级分布
Figure 2 Diameter class distribution of dominant species in Dachen Island sample plots
图 3 大陈岛植物群落相似性分析矩阵图
Figure 3 Species similarity analysis matrix of plant community on Dachen Island
图 4 大陈岛群落物种多样性与环境因子的RDA排序
Figure 4 RDA ranking of species diversity and environmental factors in Dachen Island sample plots
表 1 大陈岛群落物种重要性
Table 1. Importance of community species in Dachen Island
物种 个体数/
株平均胸
径/cm平均树
高/m物种重
要值/%柃木Eurya japonica 1 318 5.28 4.54 35.91 木麻黄Casuarina equisetifolia 299 8.93 6.54 17.85 天仙果Ficus erecta 414 3.16 3.46 8.94 樟Camphora officinarum 21 25.45 10.98 4.46 海州常山Clerodendrum
trichotomum181 3.57 4.28 4.36 海岛桑Morus maritima 141 4.52 4.56 4.33 朴树Celtis sinensis 51 14.13 6.51 4.10 野桐Mallotus japonicus 138 4.42 4.45 3.81 女贞Ligustrum lucidum 37 8.64 9.08 3.51  下载: 导出CSV
下载: 导出CSV
表 2 大陈岛群落物种多样性
Table 2. Community species diversity on Dachen Island
样方号 群落 Margalef
丰富度
指数Simpson
优势度
指数Shannon-
Wiener
多样性指数Pielou
均匀度
指数样方号 群落 Margalef
丰富度
指数Simpson
优势度
指数Shannon-
Wiener
多样性指数Pielou
均匀度
指数S01 木麻黄 1.33 0.57 1.47 0.71 S09 柃木-天仙果 1.73 0.78 1.06 0.44 S02 木麻黄-野桐 1.11 0.62 1.32 0.68 S10 柃木-天仙果 2.02 0.94 0.70 0.27 S03 木麻黄-海州常山 1.04 0.74 0.98 0.55 S11 柃木-樟 1.23 0.81 0.99 0.51 S04 天仙果 1.51 0.61 1.44 0.65 S12 柃木-野桐 1.76 0.68 1.40 0.61 S05 木麻黄-天仙果 1.21 0.74 1.02 0.57 S13 海州常山-朴树 1.62 0.56 1.71 0.82 S06 木麻黄-海州常山 1.72 0.55 1.78 0.81 S14 柃木-海州常山 2.01 0.67 1.58 0.62 S07 柃木-黄檀 1.75 0.77 1.21 0.51 S15 柃木-天仙果 2.37 0.76 1.39 0.53 S08 木麻黄-柃木 2.56 0.58 1.91 0.77
下载: 导出CSV
-
[1] 池源, 郭振, 石洪华, 等. 北长山岛人工林健康状况评价[J]. 生态科学, 2017, 36(1): 215−223. CHI Yuan, GUO Zhen, SHI Honghua, et al. Health condition assessment of plantation on Beichangshan Island, Shandong, China [J]. Ecological Science, 2017, 36(1): 215−223. [2] WANG Chen, ZHANG Hui, LIU Hui, et al. Application of a trait-based species screening framework for vegetation restoration in a tropical coral island of China [J]. Functional Ecology, 2020, 34(6): 1193−1204. [3] 石娇星. 舟山群岛植被分类与制图[D]. 上海: 华东师范大学, 2021. SHI Jiaoxing. Classification and Mapping of Vegetation in Zhoushan Islands [D]. Shanghai: East China Normal University, 2021. [4] 俞意, 吴燕生. 珠海市海岛林地植被恢复技术及效果研究[J]. 绿色科技, 2023, 25(23): 144−151. YU Yi, WU Yansheng. Research on key points of vegetation restoration technology for island deforested land in Zhuhai City [J]. Journal of Green Science and Technology, 2023, 25(23): 144−151. [5] 姚兰, 艾训儒, 易咏梅, 等. 鄂西南亚热带常绿落叶阔叶混交林优势种群的结构与动态特征[J]. 林业科学, 2017, 53(2): 10−18. YAO Lan, AI Xunru, YI Yongmei, et al. Structure and dynamics of dominant populations in the mixed forest of subtropical evergreen and deciduous broad-leaved tree species in the southwest of Hubei Province [J]. Scientia Silvae Sinicae, 2017, 53(2): 10−18. [6] 盘远方, 梁志慧, 李嘉宝, 等. 桂林岩溶石山常绿落叶阔叶混交林群落结构与物种多样性[J]. 生态学报, 2021, 41(6): 2451−2459. PAN Yuanfang, LIANG Zhihui, LI Jiabao, et al. Community structure and species diversity of evergreen deciduous broad-leaved mixed forest in Karst hills of Guilin [J]. Acta Ecologica Sinica, 2021, 41(6): 2451−2459. [7] 陈材, 唐光大, 董晓全, 等. 雷州半岛风水林灌木层优势种群空间分布格局与关联性[J]. 应用生态学报, 2024, 35(2): 371−380. CHEN Cai, TANG Guangda, DONG Xiaoquan, et al. Spatial distribution pattern and correlation of dominant populations in the shrub layer of Fengshui forest in Leizhou Peninsula, China [J]. Chinese Journal of Applied Ecology, 2024, 35(2): 371−380. [8] 金乐薇, 郭超, 许洺山, 等. 上海大金山岛植被主要组成种的种群年龄结构和存活特征[J]. 应用与环境生物学报, 2023, 29(5): 1132−1141. JIN Lewei, GUO Chao, XU Mingshan, et al. Population age structure and survivorship of vegetation component species on Dajinshan Island, Shanghai [J]. Chinese Journal of Applied and Environmental Biology, 2023, 29(5): 1132−1141. [9] 张川英, 李婷婷, 龚笑飞, 等. 遂昌乌溪江流域山蜡梅生境群落特征与物种多样性[J]. 浙江农林大学学报, 2023, 40(4): 848−858. ZHANG Chuanying, LI Tingting, GONG Xiaofei, et al. Habitat community characteristics and species diversity of Chimonanthus nitens in Wuxi River Basin, Suichang [J]. Journal of Zhejiang A&F University, 2023, 40(4): 848−858. [10] 陈越琳, 黄阿青, 肖集泓, 等. 福建连江县5个无居民海岛种子植物区系及地理分布研究[J]. 热带作物学报, 2024, 45(2): 424−434. CHEN Yuelin, HUANG Aqing, XIAO Jihong, et al. Seed plant flora and distribution on five uninhabited islands in Lianjiang, Fujian Province [J]. Chinese Journal of Tropical Crops, 2024, 45(2): 424−434. [11] 孔昊, 杨薇. 气候变化背景下海岛生态环境脆弱性分析及其应对措施——以南澳岛为例[J]. 海洋开发与管理, 2016, 33(10): 72−77. KONG Hao, YANG Wei. Analysis and measures of island ecological vulnerability under the background of climate change: a case study on Nan’ao Island [J]. Ocean Development and Management, 2016, 33(10): 72−77. [12] 汤坤贤, 陈慧英, 陈玉珍, 等. 干旱胁迫对3种海岛植被修复植物生长与生理指标的影响[J]. 厦门大学学报(自然科学版), 2017, 56(6): 845−851. TANG Kunxian, CHEN Huiying, CHEN Yuzhen, et al. Effects of drought stress on morphological and physiological characteristics of three island vegetation restoration plants [J]. Journal of Xiamen University (Natural Science), 2017, 56(6): 845−851. [13] 赵定蓉, 陆梅, 赵旭燕, 等. 土壤细菌群落对纳帕海高原湿地退化的响应[J]. 浙江农林大学学报, 2024, 41(2): 406−418. ZHAO Dingrong, LU Mei, ZHAO Xuyan, et al. Response of soil bacterial community to wetland degradation in the Napahai Plateau [J]. Journal of Zhejiang A&F University, 2024, 41(2): 406−418. [14] 闫潍虹. 庙岛群岛之小钦岛植物群落结构及群落谱系构建研究[D]. 烟台: 鲁东大学, 2018. YAN Weihong. Study on Plant Community Structure and Community Pedigree Construction of Xiaoqin Island in Miaodao Islands [D]. Yantai: Ludong University, 2018. [15] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533−548. FANG Jingyun, WANG Xiangping, SHEN Zehao, et al. Methods and protocols for plant community inventory [J]. Biodiversity Science, 2009, 17(6): 533−548. [16] 张蔷, 李令军, 陈韵如, 等. 北京山地森林群落物种多样性、生物量及其关系分析[J/OL]. 中国环境科学, 2024-03-26[2024-07-01]. https://doi.org/10.19674/j.cnki.issn1000-6923.20240322.011. ZHANG Qiang, LI Lingjun, CHEN Yunru, et al. Analysis on species diversity and biomass of mountain forest communities in Beijing and their relationships [J/OL]. China Environmental Science, 2024-03-26[2024-07-01]. https://doi.org/10.19674/j.cnki.issn1000-6923.20240322.011. [17] 张恒庆, 许爽, 朱鹏宇, 等. 长海县大耗岛植物多样性研究[J]. 辽宁大学学报(自然科学版), 2020, 47(1): 12−19. ZHANG Hengqing, XU Shuang, ZHU Pengyu, et al. Plant diversity of Dahao Island in Changhai County [J]. Journal of Liaoning University (Natural Science Edition), 2020, 47(1): 12−19. [18] 于世勇, 张信杰, 刘波, 等. 我国寒温带典型森林群落特征及生物多样性研究[J]. 温带林业研究, 2023, 6(1): 1−6. YU Shiyong, ZHANG Xinjie, LIU Bo, et al. Research on the characteristics and biodiversity of typical cold-temperate forest communities in China [J]. Journal of Temperate Forestry Research, 2023, 6(1): 1−6. [19] 高大海, 陈斌, 贺位忠, 等. 舟山海岛柃木的分布和生境群落学初步研究[J]. 浙江林业科技, 2013, 33(6): 32−36. GAO Dahai, CHEN Bin, HE Weizhong, et al. Investigation on distribution and community of Eurya japonica in Zhoushan [J]. Journal of Zhejiang Forestry Science and Technology, 2013, 33(6): 32−36. [20] 郑俊鸣, 方笑, 朱雪平, 等. 外马廊山岛植被特性与植物多样性[J]. 广西植物, 2017, 37(3): 271−279. ZHENG Junming, FANG Xiao, ZHU Xueping, et al. Vegetation characteristics and plant diversity of Waimalangshan Island, Zhoushan [J]. Guihaia, 2017, 37(3): 271−279. [21] 王芳, 刘阳, 马晓迪, 等. 平潭岛木麻黄群落灌木层优势物种生态位及种间联结分析[J]. 山东林业科技, 2024, 54(3): 64−71. WANG Fang, LIU Yang, MA Xiaodi, et al. Analysis on the ecological niche and interspecific linkage of dominant species in the shrub layer of Casuarina equisetifolia community in Pingtan Island [J]. Journal of Shandong Forestry Science and Technology, 2024, 54(3): 64−71. [22] 李应, 詹洪平, 万炜, 等. 中国柃木属植物资源概述[J]. 南方林业科学, 2024, 52(1): 56−59, 66. LI Ying, ZHAN Hongping, WAN Wei, et al. Overview of Eurya plant resources in China [J]. South China Forestry Science, 2024, 52(1): 56−59, 66. [23] 管诗敏, 郑心炫, 吴雅华, 等. 平潭岛海岸自然植被物种和功能多样性特征[J]. 西南林业大学学报(自然科学), 2021, 41(6): 71−78. GUAN Shimin, ZHENG Xinxuan, WU Yahua, et al. Species diversity and functional diversity of primary vegetation on the coast of Pingtan Island [J]. Journal of Southwest Forestry University (Natural Sciences), 2021, 41(6): 71−78. [24] 达良俊, 杨永川, 陈燕萍. 上海大金山岛的自然植物群落多样性[J]. 中国城市林业, 2004, 2(3): 22−25. DA Liangjun, YANG Yongchuan, CHEN Yanping. The diversity of plant community on Dajinshan Island, Shanghai [J]. Journal of Chinese Urban Forestry, 2004, 2(3): 22−25. [25] CAPDEVILA P, STOTT I, CANT J, et al. Life history mediates the trade-offs among different components of demographic resilience [J]. Ecology Letters, 2022, 25(6): 1566−1579. [26] 邓力豪. 舟山群岛常绿阔叶林群落特征研究[D]. 长沙: 中南林业科技大学, 2022. DENG Lihao. Study on Community Characteristics of Evergreen Broad-leaved Forest in Zhoushan Islands [D]. Changsha: Central South University of Forestry & Technology, 2022. [27] 何雅琴, 曾纪毅, 陈国杰, 等. 福建平潭大练岛典型森林群落特征及物种多样性[J]. 应用与环境生物学报, 2022, 28(3): 759−769. HE Yaqin, ZENG Jiyi, CHEN Guojie, et al. Characteristics and species diversity of typical forest community in Dalian Island of Pingtan, Fujian, China [J]. Chinese Journal of Applied and Environmental Biology, 2022, 28(3): 759−769. [28] 梁楚欣, 范弢, 陈培云. 滇东石漠化坡地不同恢复模式下云南松林土壤碳氮磷化学计量特征及其影响因子[J]. 浙江农林大学学报, 2023, 40(3): 511−519. LIANG Chuxin, FAN Tao, CHEN Peiyun. Stoichiometric characteristics and influencing factors of soil C, N and P in Pinus yunnanensis forests under different restoration modes on rocky desertification slope land in eastern Yunnan [J]. Journal of Zhejiang A&F University, 2023, 40(3): 511−519. [29] 童珊, 曹广超, 张卓, 等. 土壤全碳全氮空间异质性及影响因素分析——以祁连山南坡黑河上游为例[J]. 土壤, 2022, 54(5): 1064−1072. TONG Shan, CAO Guangchao, ZHANG Zhuo, et al. Analysis of spatial heterogeneity and influencing factors of soil total carbon and nitrogen: take the upper reaches of Heihe River on southern slope of Qilian Mountain as an example [J]. Soils, 2022, 54(5): 1064−1072. [30] 李其斌, 张春雨, 赵秀海. 长白山不同演替阶段针阔混交林群落物种多样性及其影响因子[J]. 生态学报, 2022, 42(17): 7147−7155. LI Qibin, ZHANG Chunyu, ZHAO Xiuhai. Species diversity and influencing factors of coniferous and broad-leaved mixed forest communities in different successional stages of Changbai Mountain [J]. Acta Ecologica Sinica, 2022, 42(17): 7147−7155. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240423
 点击查看大图
点击查看大图
计量
- 文章访问数: 1122
- HTML全文浏览量: 395
- PDF下载量: 35
- 被引次数: 0
