




-
在群落构建过程中,物种共现或排斥的格局一直吸引着生态学家[1]。虽然共存理论一直在发展,新的假说不断出现,但此问题尚未完全解决[2]。径级结构是种群最基本的特征,反映了特定时间内种群中个体在不同径级下的配置状况和发展趋势[3]。空间格局是植物个体在空间中所处的位置和分布状态,其形成与种群生物学特性、个体间相互作用和生境因子影响等诸多因素密切相关[4]。对不同种群的径级结构及空间格局进行研究不仅可以反映种群的分布特征,更有助于揭示维持物种共存的潜在机制[5−6]。
以往对种群空间格局的研究多以群落中少数几个常见种或优势种作为主要研究对象[7−8],但对群落中大多数物种的空间格局及种间相关性的分析较少。由于物种自身的属性不同,其生态学特性也会存在差异,但同一类群的物种可能会存在一些相似的生态学特性。安璐等[9]研究发现:冠层木和灌木的成熟个体表现出聚集分布,而亚林层木在20 m处由聚集分布转为随机分布。郭屹立等[10]发现:不同多度类群(稀有种、常见种和优势种)或不同生活型之间的聚集度存在显著差异。偶见种和稀有种作为群落的重要组分,常以较少的个体数贡献较大的物种多样性[4, 11]。但由于其个体数较少,通常较难单独分析。而对类群整体的研究可以解决单个物种个体数不足的问题,从而反映类群整体的一些生态学特征[12−13]。那么不同类群或物种在径级结构以及资源利用上是否存在差异,生境异质性以及生态位分化是否会影响这些类群或物种的共存?鉴于此,本研究在浙江九龙山国家级自然保护区内岩坪1 hm2的固定样地内,基于所有胸径≥1 cm木本植物个体的调查数据,根据物种的多度等属性划分类群(优势种、常见种、偶见种与稀有种),研究了不同种群或类群的径级结构,探究其空间关联以及生境偏好,以期进一步认识群落中的物种共存机制,并为制定相关的保护措施提供科学依据。
-
九龙山国家级自然保护区(28°19′10″~28°24′43″N,118°49′38″~118°55′03″E)地处浙、闽、赣三省毗邻地带的浙江遂昌县西南部,为武夷山系仙霞岭山脉的一部分,总面积为55.25 hm2[14]。保护区处于中亚热带季风气候带,冬冷夏凉,四季分明,雨量充沛。年均气温为16.2 ℃,极端最高气温为36.5 ℃,极端最低气温为−10.5 ℃,年降水量为1 855.6 mm,相对湿度为80%,年日照时数为1 925.0 h[15]。
-
参照美国热带森林研究中心(CTFS)的方法[16],在九龙山国家级自然保护区内岩坪(28°23′N,118°53′E,海拔为707~758 m)建立一个1 hm2 (100 m × 100 m)的常绿阔叶林长期监测样地。对样地内胸径(DBH)≥1 cm的所有木本植物个体进行调查,记录个体的种名、空间坐标、DBH、高度及生活状态。样地平均坡度为32.77°,坡向东南,平均岩石裸露率为27.33%,平均土层厚度为15.93 cm。样地所在位置曾为杉木Cunninghamia lanceolata林,在20世纪60年代前后经过中等程度的杉木采伐活动,随后禁止人为干扰,自然演替为常绿阔叶林。
将1 hm2样地均匀划分为20 m × 20 m的样方,在各样方的顶点及中心点于土层0~20 cm位置取土样,共计61份土壤样品。样品除去根系,混合后装入贴编号的封口袋中,将土壤样品风干后过0.075 mm筛,使用元素分析仪测定土壤的全碳和全氮,采用原子吸收光谱测定全磷等化学参数[17]。通过罗盘仪和卷尺在每个取样点测量坡度、坡向和岩石裸露率等地形因子。
-
以往研究将1 hm2内个体数≤1株的物种定义为稀有种,>1株且≤10株的物种为偶见种,>10株的物种称为常见种[18]。由于优势种通常对群落内的其他物种产生较大影响,本研究把样地内重要值最大的物种——木荷Schima superba从常见种中独立出来作为优势种,然后对优势种、常见种、偶见种与稀有种之间的关系和差异进行研究。
采用点格局分析中的g(r)函数分析群落内不同类群的空间格局。g(r)函数以中心树为圆心,半径为r,环宽为w的圆环中个体密度的期望,再除以格局的聚集强度λ计算得到[19]。为排除生境异质性效应对物种空间分布的影响,本研究采用完全空间随机(CSR)模型和异质性泊松过程(HP)分析不同类群的空间格局[20]。其中CSR模型假设个体在空间中任一位置出现的概率相同,其分布不受任何生物或非生物过程影响。而HP模型是依据空间异质性强度函数λ(x, y)决定个体的分布,可以排除大尺度上生境异质性的影响。本研究采用高斯核函数进行密度估计,选取25 m作为标准差排除大尺度上生境异质性对空间格局的影响[21]。不同类群间的空间关联分析采用双变量g12(r)函数分析,由于种间竞争是非对称的,需分别检验物种1对物种2的影响和物种2对物种1的影响,为此采用先决条件零假设和环向位移零假设检验不同类群之间的空间关联[8]。
通过199次Monte Carlo随机模拟计算99%的置信区间,当某一尺度上的函数值高于上包迹线表示该类群在该尺度上为聚集分布或2种类群间存在显著正相关,低于下包迹线表示该类群在该尺度上为均匀分布或2种类群间存在显著负相关,在上下包迹线之间表示该类群在该尺度上为随机分布或2种类群间无显著相关[22]。拟合优度检验用于检验模型对实际格局的拟合能力。计算0~25 m尺度内观测值和理论值的离差平方和,用秩检验检测模型的显著性,当观测值的秩<5或>195时,说明观测值在P=0.05水平上显著偏离理论值[23]。
将样方内5个原始环境数据的平均值作为该样方自变量,样方内某一类群的个体数作为因变量,构建全模型。使用具有特定空间相关结构的广义最小二乘模型消除空间自相关的影响[24]。通过向后选择法逐步回归,根据赤池信息量准则(AIC)筛选最优模型,模型间的AIC差值≥2时认为模型具有显著差异[25]。使用R 4.0.2软件[26]完成上述分析和制图。
-
本研究群落中共有132种植物,5 015株个体,其中个体数≥30株的物种有25种,个体数总计4 364株,占群落中总个体数的87.02%。除优势种木荷外的24种植物的径级结构见表1。其中:短尾越橘Vaccinium carlesii、峨眉鼠刺Itea omeiensis、格药柃Eurya muricata、海桐山矾Symplocos heishanensis、红楠Machilus thunbergii、檵木Loropetalum chinense、马银花Rhododendron ovatum、毛花连蕊茶Camellia fraterna、拟赤杨Alniphyllum fortunei、青冈Quercus glauca、杉木、微毛柃Eurya hebeclados、细枝柃Eurya loquaiana、窄基红褐柃Eurya rubiginosa var. attenuata和浙江新木姜子Neolitsea aurata var. chekiangensis的径级结构呈“L”型分布。麂角杜鹃Rhododendron latoucheae、山槐Albizia kalkora和小果冬青Ilex micrococca的径级结构近似正态分布。杜鹃Rhododendron simsii、石斑木Rhaphiolepis indica和石木姜子Litsea elongata var. faberi的个体胸径集中在2~4 cm,径级结构体呈现单柱型结构。南酸枣Choerospondias axillaris、石栎Lithocarpus glaber和甜槠Castanopsis eyrei的径级结构为间歇型,小树较多,大径级个体间断分布。
表 1 24种植物的空间格局
Table 1. Spatial distribution patterns of 24 species
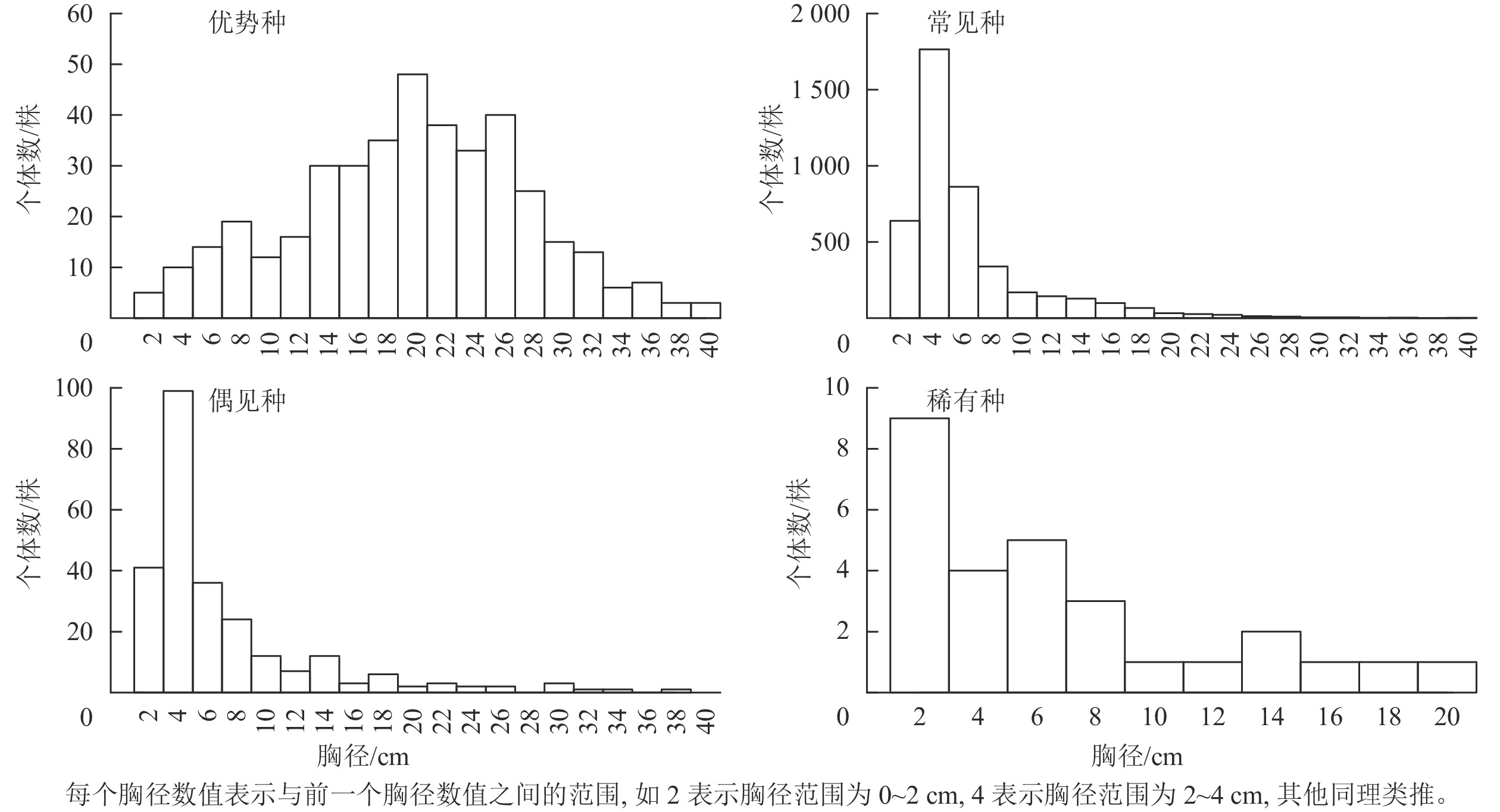
物种 径级结构 完全空间随机模型 异质性泊松过程 聚集分布 随机分布 均匀分布 P 聚集分布 随机分布 均匀分布 P 杜鹃 Rhododendron simsii 单柱型 20 5 0 0.005 4 21 0 0.045 短尾越橘 Vaccinium carlesii “L”型 1 24 0 0.040 0 25 0 0.010 峨眉鼠刺 Itea omeiensis “L”型 5 20 0 0.005 1 24 0 0.005 格药柃 Eurya muricata “L”型 24 1 0 0.005 6 19 0 0.005 海桐山矾 Symplocos heishanensis “L”型 0 25 0 0.545 0 25 0 0.240 红楠 Machilus thunbergii “L”型 25 0 0 0.005 3 21 1 0.005 麂角杜鹃 Rhododendron latoucheae 稳定型 21 4 0 0.005 0 25 0 0.060 檵木 Loropetalum chinense “L”型 19 6 0 0.005 11 6 8 0.005 马银花 Rhododendron ovatum “L”型 20 4 1 0.005 18 6 1 0.005 毛花连蕊茶 Camellia fraterna “L”型 25 0 0 0.005 6 19 0 0.005 南酸枣 Choerospondias axillaris 间歇型 19 6 0 0.005 0 25 0 0.095 拟赤杨 Alniphyllum fortunei “L”型 9 15 1 0.005 7 16 2 0.010 青冈 Quercus glauca “L”型 18 7 0 0.005 3 22 0 0.040 山槐 Albizia kalkora 稳定型 0 25 0 0.480 0 25 0 0.365 杉木 Cunninghamia lanceolata “L”型 19 6 0 0.005 10 14 1 0.005 石斑木 Rhaphiolepis indica 单柱型 1 24 0 0.005 0 25 0 0.490 石栎 Lithocarpus glaber 间歇型 1 24 0 0.005 1 24 0 0.005 石木姜子 Litsea elongata var. faberi 单柱型 4 21 0 0.005 1 24 0 0.010 甜槠 Castanopsis eyrei 间歇型 2 23 0 0.040 0 25 0 0.985 微毛柃 Eurya hebeclados “L”型 22 3 0 0.005 13 10 2 0.005 细枝柃 Eurya loquaiana “L”型 17 8 0 0.005 6 19 0 0.005 小果冬青 Ilex micrococca 稳定型 0 25 0 0.305 0 25 0 0.330 窄基红褐柃 Eurya rubiginosa var. attenuata “L”型 25 0 0 0.005 14 9 2 0.005 浙江新木姜子 Neolitsea aurata var. chekiangensis “L”型 1 24 0 0.050 0 25 0 0.475 说明:聚集分布、随机分布和均匀分布列下的数字为相应格局的尺度数之和。 群落内优势种木荷有402株;常见种有45种,4 330株;偶见种有58种,255株;稀有种有28种,28株。优势种的径级结构近似正态分布,其胸径<10 cm和>30 cm的树种较少,而胸径20 cm的树种较多。常见种和偶见种的小径级个体较多,径级结构为“L”型分布。稀有种的径级结构也呈“L”型分布,但缺少胸径>20 cm的个体(图1)。
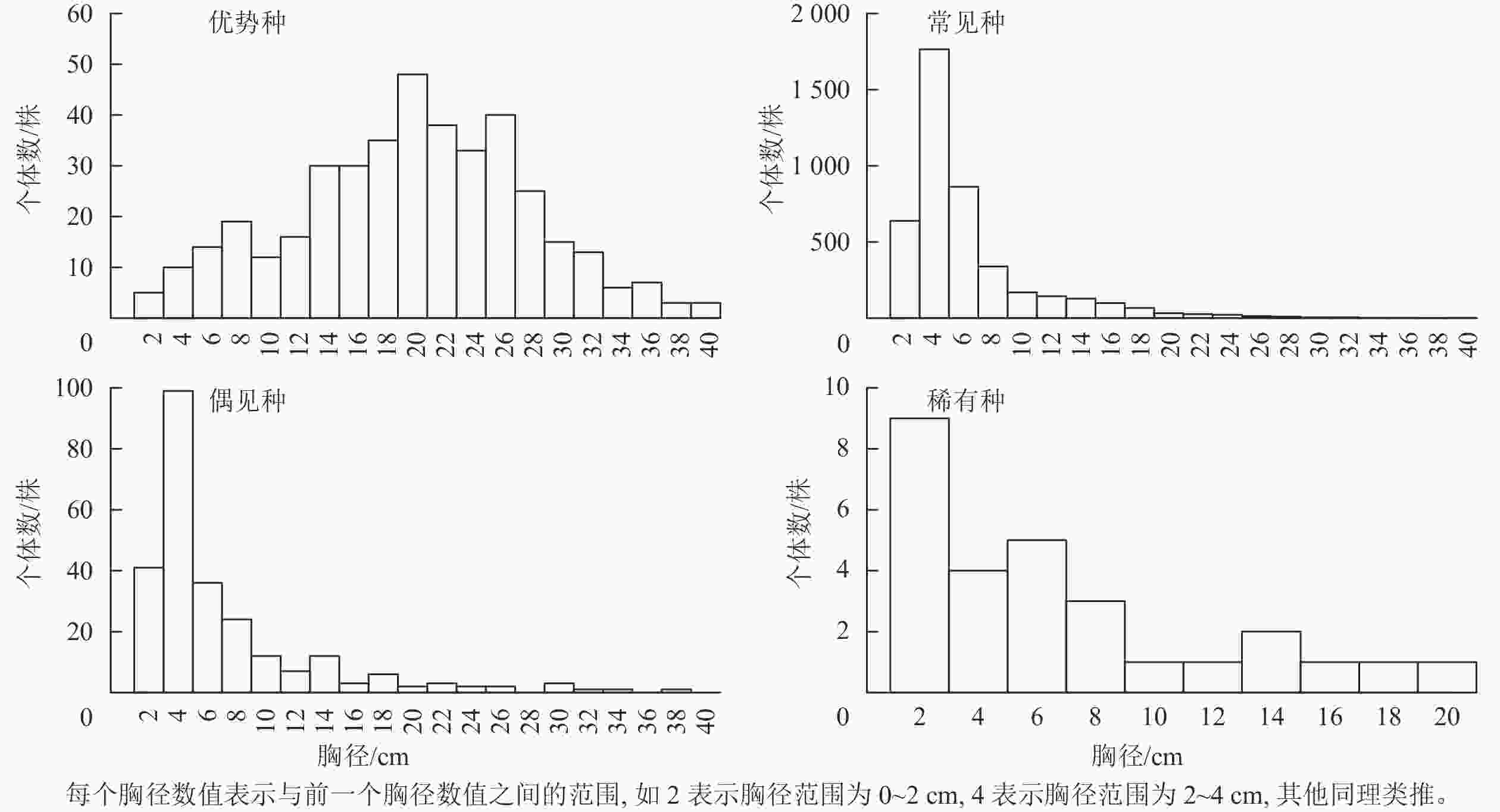
图 1 不同类群的径级结构
Figure 1. Size structure of different groups
-
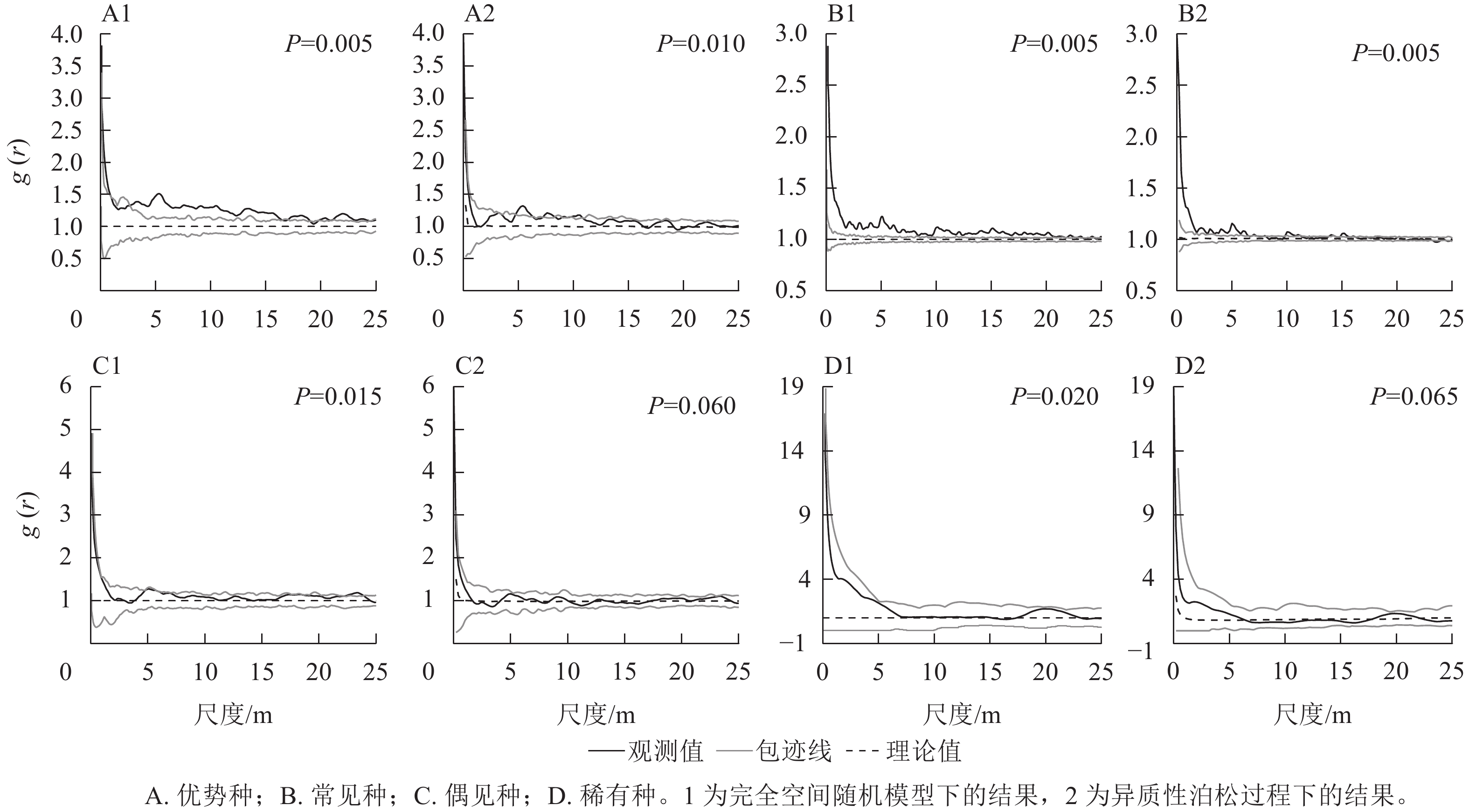
如图2所示:在完全空间随机模型下,除了稀有种类群,其他3个类群均表现出聚集分布;在排除生境异质性后,优势种在<10 m的大多数尺度上以聚集分布为主,在>10 m的尺度上变为均匀分布;常见种在大多数尺度上表现为聚集分布,偶见种和稀有种均表现为随机分布。拟合优度检验结果(图2)显示:所有类群的空间格局在0~25 m的尺度上均显著偏离完全空间随机模型,表现为聚集分布;在异质性泊松过程下,优势种和常见种显著偏离模型,表现为聚集分布,而偶见种和稀有种符合模型,表现为随机分布。
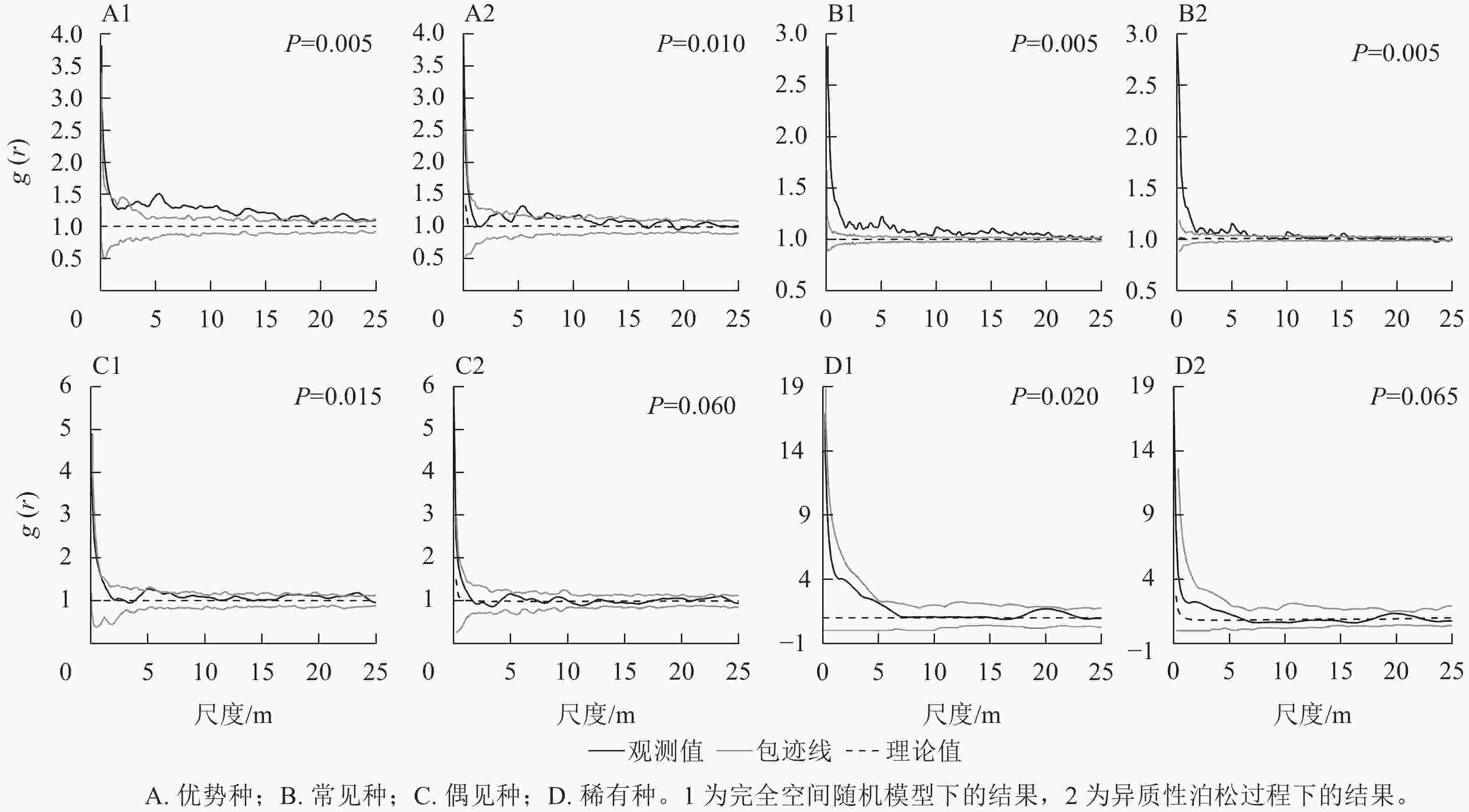
图 2 不同类群的空间格局变化
Figure 2. Change of spatial distribution pattern of different groups
对样地中除优势种外的个体数≥30株的24种植物分析(表1)发现:海桐山矾、山槐和小果冬青在CSR和HP模型下均表现为随机分布,其他21种植物在CSR模型下均表现出聚集分布。但在HP模型下,短尾越橘、麂角杜鹃、南酸枣、石斑木、甜槠和浙江新木姜子变为随机分布,其他18种植物表现出聚集的尺度数明显减少。拟合优度检验结果显示,仅有海桐山矾、山槐和小果冬青符合CSR模型,表现为随机分布,其余21种植物均为聚集分布。而在HP模型下,除了海桐山矾、麂角杜鹃、南酸枣、山槐、石斑木、甜槠、小果冬青和浙江新木姜子表现为随机分布外,其余16种植物均为聚集分布。
由于径级结构可能会对物种的空间格局产生较大影响,本研究进一步比较了不同径级结构的物种空间格局差异。结果发现:“L”型分布的物种在CSR模型下有93.33%的物种表现为显著的聚集分布(P<0.05),单柱型、间歇型的物种均表现为显著的聚集分布,而稳定型的物种呈显著聚集分布的物种百分比仅为33.33%。在排除了生境异质性后,“L”型分布的物种中仍有86.67%的物种表现为聚集分布,而稳定型的物种则均表现为随机分布。
-
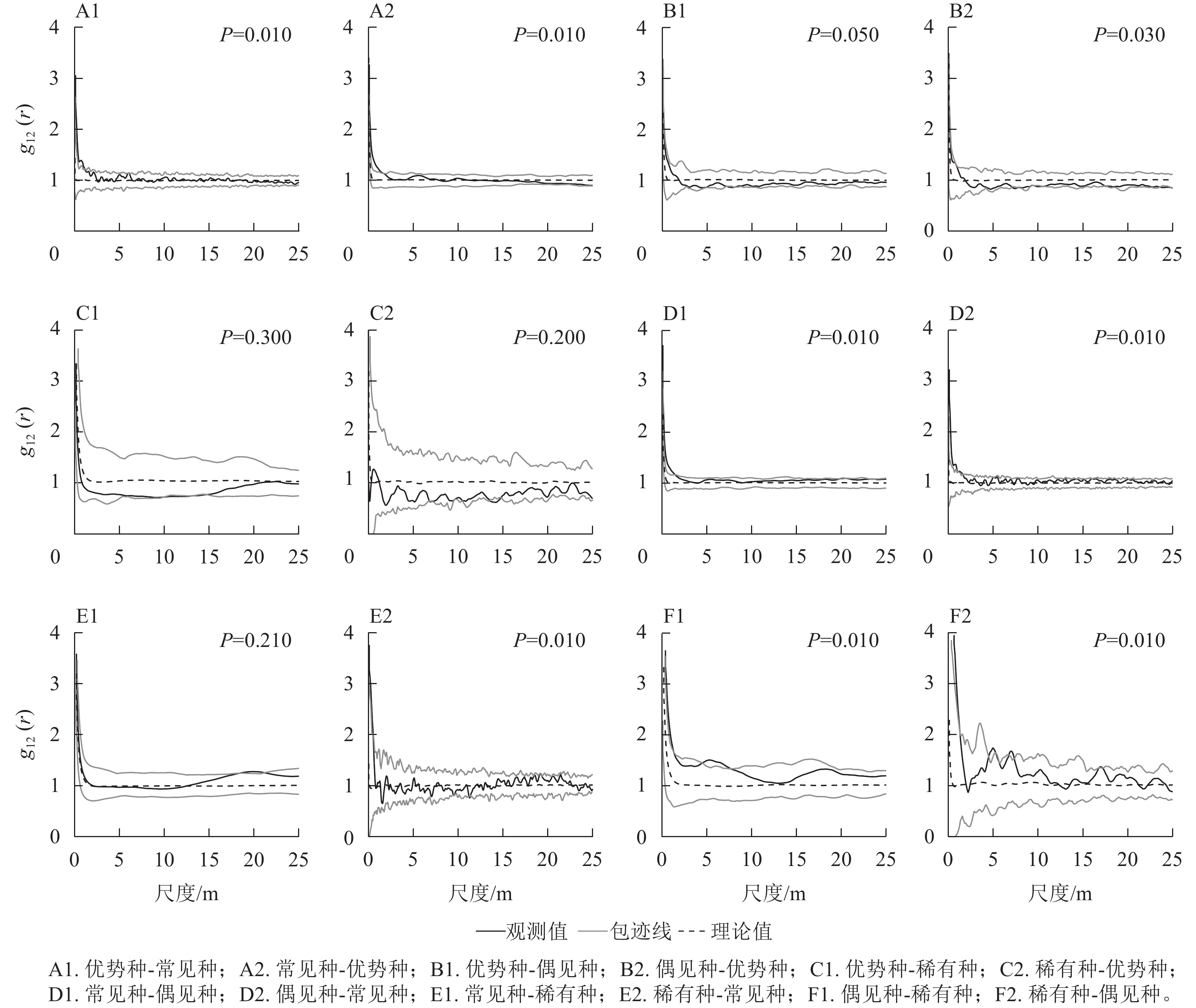
如图3所示:优势种与常见种均在<3 m的尺度上存在正相关,与偶见种均在7 m的尺度上呈现负相关,与稀有种均在10~15 m的部分尺度上表现出负相关。常见种与偶见种均在<3 m的尺度上存在正相关,常见种与稀有种、稀有种与常见种分别在20 m以及0~2 m尺度上存在正相关。偶见种与稀有种互相均在<10 m以及15~20 m的部分尺度上存在正相关。拟合优度检验结果显示:优势种与稀有种为不显著负相关,常见种与稀有种为不显著正相关,其他类群间的空间关联均达到显著水平。
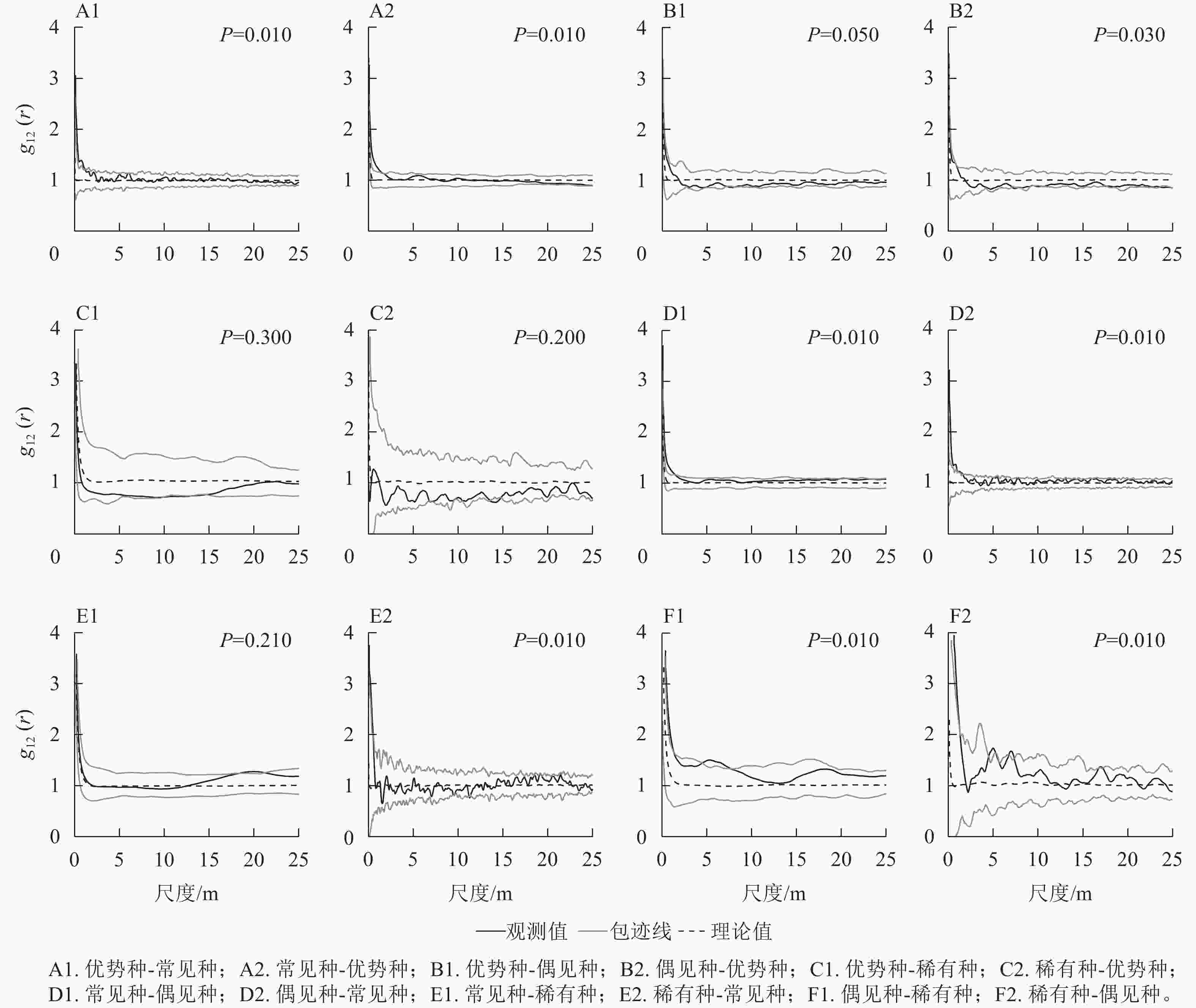
图 3 不同类群之间的空间关联变化
Figure 3. Change of spatial association between each two different groups
不同类群与24种植物的相关性存在较大差异(表2)。总体上,不同类群之间表现出显著的相关性,并以正相关为主,同时小尺度(0~10 m)上的负相关几乎没有。全部尺度(0~25 m)与小尺度上的相关趋势相同,正相关占比最高的是优势种与24种植物,其次为偶见种与24种植物以及24种植物之间,而稀有种与24种植物之间的正相关占比最低。不同类群之间的负相关集中在大尺度(11~25 m)上,负相关占比最高的为优势种与24种植物,其次为稀有种与24种植物,偶见种与24种植物以及24种植物之间的负相关占比最低。
表 2 不同类群与24种植物的空间关联
Table 2. Spatial association between different groups and 24 species
物种间的空间关联 不同尺度下显著空间关联的物种对占总物种对的比例/% 总物种对 0~10 m 11~25 m 0~25 m 负相关 正相关 负相关 正相关 负相关 正相关 优势种与24种植物 0.00 25.00 12.50 4.17 12.50 27.08 48 偶见种与24种植物 0.00 16.67 2.08 4.17 2.08 16.67 48 稀有种与24种植物 0.00 2.08 4.17 4.17 4.17 6.25 48 24种植物之间 1.63 9.96 1.99 7.79 2.90 13.41 552 -
不同类群与生境因子的相关性存在差异(表3)。优势种与岩石裸露率之间呈显著负相关,与碳含量的正相关达到边缘显著水平。常见种与磷含量呈显著正相关,与岩石裸露率的负相关达到边缘显著水平。偶见种与坡向呈显著正相关,与磷含量呈显著负相关。稀有种与碳含量呈显著负相关。
表 3 不同类群与生境因子的相关性
Table 3. Correlation between different groups and environmental factors
不同类群 最优模型参数 估计值 标准误 P 赤池信息量准则(AIC) 模型P 优势种 截距 32.056 13.688 0.030 170.128 <0.001 坡度 0.847 2.160 0.699 坡向 −1.915 1.676 0.267 岩石裸露率 −4.143 1.987 0.050 碳含量 3.915 2.194 0.090 常见种 截距 294.240 54.339 <0.001 244.723 <0.001 坡度 0.777 14.567 0.958 坡向 −15.223 12.143 0.224 岩石裸露率 −26.870 13.016 0.052 磷含量 33.214 13.896 0.027 偶见种 截距 9.612 1.049 <0.001 148.895 0.001 坡向 3.451 1.078 0.004 岩石裸露率 −1.294 1.020 0.219 磷含量 −2.532 1.122 0.035 稀有种 截距 2.120 0.407 <0.001 110.297 0.005 碳含量 −1.252 0.415 0.006 -
种群的径级结构可以反映其更新类型和发展趋势[3]。本研究样地原为杉木林,在20世纪60年代前后受到杉木采伐活动,随后自然演替为常绿阔叶林,目前属于演替中期向后期过渡的阶段。本研究中,优势种木荷以及麂角杜鹃、山槐和小果冬青的径级结构呈近似正态分布,这些物种的个体集中在中间径级,种群结构比较稳定[27]。常见种和偶见种的径级结构均为“L”型分布,同时24种植物中有21种的径级结构呈“L”型分布、单柱型结构或间歇型结构,这些径级结构都是小径级个体较多,表明这些类群的小树较多,更新状况良好,种群数量会保持稳定或继续增长[3, 11]。稀有种的径级结构为“L”型分布,但没有胸径>20 cm的个体,可见稀有种类群的小径级个体数量较多,类群发展属于增长型。群落中除了优势种外的其他3个类群以小径级个体较多,表明群落中物种总体的更新状况良好,多数物种在稳定增长。
周赛霞等[28]对后河自然保护区珍稀植物的研究表明:稀有濒危植物寿命长,并且大多物种幼苗更新良好,由此推测稀有濒危植物能长期稳定地存在于群落中。但就本研究稀有种类群中的28种植物而言,每个物种仅有1株个体,极易由于个体的死亡导致物种丢失。需加强对稀有种的保护,营造适合稀有种更新生长的生境。
-
种群的空间分布格局通常是物种与生物和非生物因子长期相互作用的结果[4]。在CSR模型下,优势种和常见种均表现出聚集分布,在排除生境异质性后,优势种和常见种在小尺度上仍然表现聚集分布,表明这2个类群在大尺度上的聚集可能是生境异质性所致,而小尺度上的聚集可能主要与种子扩散限制等生物因素有关,这与前人的研究结果一致[7]。而偶见种则由聚集分布完全变为随机分布,这表明偶见种偏向于在特定生境中聚集生存。稀有种在2种模型下均表现为随机分布,但拟合优度检验显示:稀有种在CSR模型下为聚集分布,而在HP模型下为随机分布,这表明稀有种在一定程度上受到生境异质性的影响,同时稀有种的随机分布也可能是类群的衰退所致。李文良等[29]研究发现:随着连香树Cercidiphyllum japonicum种群衰退加剧,格局由聚集分布向随机分布转变。
对24种常见种进一步分析发现:有21种在CSR模型下表现为聚集分布,排除了生境异质性后,仍有多数物种表现为聚集分布,但聚集程度明显减弱。这表明大多数常见种物种都为聚集分布,并且受到生境异质性的影响较大,这与常见种类群的结果较为一致。同时也表明对常见种类群整体空间格局的分析在一定程度上反映了多数常见种的格局。
在CSR和HP模型下,“L”型、单柱型和间歇型等具有众多小径级个体的物种比小径级个体较少的稳定型物种更容易表现出聚集分布。表明小径级个体通常更容易聚集分布,而这可能也是造成种群总体聚集分布的原因之一。前人研究同样发现聚集程度随径级增大而明显减弱[6, 22]。这可能是由于扩散限制等原因使得小树聚集分布于母树附近[7];而大径级个体经历了较强的种内竞争以及天敌等影响后,聚集程度有所减弱[19]。
-
优势种作为群落中优势度最大的物种,占据了大量的生存空间。优势种与偶见种、稀有种均呈现负相关,这说明稀有种和偶见种均在特定尺度上受到优势种的竞争排斥,在群落中的发展处于劣势地位[30]。但从0~25 m整体尺度来看,优势种与稀有种之间的相关性并不显著,说明稀有种受到优势种的影响较为有限。偶见种与稀有种呈正相关,表明这2个类群存在一定的种间互助,同时也可能因为都被优势种排斥而均生长在优势种较少的位置。常见种与其他几个类群均存在正相关,一方面表明常见种与其他几个类群间没有明显竞争关系,而且可能存在一定互助作用[31];另一方面可能与其本身数量较大,在群落中随处可见有关。
对24种常见种分析发现:这些常见种与其他类群的空间关联以正相关为主,这与常见种类群的结果较为一致。这些常见种与优势种的正相关和负相关的占比均最高,表明群落中24种植物与优势种之间的相互作用比与偶见种或稀有种的相互作用更加强烈。同时24种植物与其他类群在小尺度上的正相关表明群落中竞争较小,物种间存在互利作用。这可能与环境中资源较为丰富有关[6]。同时也可能是较强的种内竞争或密度制约减弱了种间竞争所致[32]。24种植物与其他类群在大尺度上的负相关表明这些类群对生境利用存在分化,这进一步减弱了可能存在的种间竞争,同时也表明各种群暂时处于稳定发展的状态。
样地中不同类群均至少与一种生境因子呈显著相关性,表明这些类群的分布均受到不同程度生境因素的影响。优势种和常见种的大多数个体偏向于生长在岩石较少的地方。此外优势种会生长在碳含量较高的地方,而常见种多生长在磷含量较高的环境中。偶见种主要生长在少磷的地方,而且在西坡的分布比东坡略多。而稀有种主要生长在少碳的环境中。不同类群对环境因子的偏好存在较大差异。同一群落中,不同类群在资源的利用上存在一定的相似性,但经过长时间的相互作用也产生一定的生态位分化,进而有利于物种共存[6, 33]。
-
本研究发现:样地中多数物种或类群为聚集分布,并受到生境异质性的影响。相比小径级个体较少的稳定型物种,具有众多小径级个体的“L”型等径级结构的种群更容易表现出聚集分布。不同类群的结构和空间分布格局具有一些其特有的生态学性质,并且它们对资源利用的分化有利于其共存。光环境对物种空间分布也存在较大影响,今后应深入研究不同环境因子对不同生活型物种的影响。本研究仅使用了1次的调查数据,在一定程度上反映了该类群组成物种的共性,还需进行长期观测以及大尺度的研究。
-
浙江大学的张田田、吴倩倩、王莹等同学以及浙江师范大学的刘晓彤、谭斌、樊海东、袁泉、王国敏、曹嘉瑜、徐佳参加了野外调查工作,谢雨初、王云泉、刘立斌和仲磊对论文的修改提供了意见和建议,一并致谢!
Size structure and spatial association of different species of an evergreen broad-leaved forest in Jiulong Mountain, Zhejiang Province
-
摘要:
目的 不同植物物种或类群(优势种、常见种、偶见种与稀有种)往往具有不同的生态学特性。通过对不同物种或类群的径级结构和空间格局进行研究,有助于揭示物种共存机制。 方法 基于浙江九龙山国家级自然保护区1 hm2常绿阔叶林长期监测样地,通过点格局分析等方法,对24种植物和不同类群的径级结构以及空间关联性进行分析。 结果 ①15种植物以及常见种、偶见种和稀有种为小径级个体居多的“L”型分布,3种植物为单柱型分布,3种植物为间歇型分布,3种植物和优势种的径级结构为近似正态分布;②多数物种(21/24)或类群以聚集分布格局为主,并受到生境异质性的影响;③优势种与偶见种、稀有种均呈负的空间关联,常见种与其他类群以正的空间关联为主,24种植物对其他类群也以正的空间关联为主。 结论 群落中多数物种和类群受到生境异质性的影响,同时生态位分化减弱了物种或类群之间的竞争,在其共存中发挥着重要作用,群落暂时处于一个稳定的状态。图3表3参33 Abstract:Objective Different plant species and groups, with dominant species, common species, occasional species and rare species included, often demonstrate various ecological characteristics. Therefore, this study, with an investigation of the size structure and spatial pattern of different species and groups, is aimed to further explore mechanisms of species coexistence. Method With the long-term monitoring 1 hm2 plot of evergreen broad-leaved forest in Jiulong Mountain National Nature Reserve of Zhejiang Province, China selected as the subject, point pattern analysis was conducted of the size structure of 24 common species and different groups and their association with other groups and habitat factors. Result (1) The size structure of 15 species and common species, occasional species, and rare species was mainly distributed in an “L” shape with more small-diameter individuals. The size structure of 3 species was distributed in sporadic type and 3 species was distributed in unibar type. The size structure of 3 species and dominant species was close to the normal distribution; (2) The spatial distribution pattern of most species (21 out of 24) or groups was in aggregation and was affected by habitat heterogeneity; (3) Dominant species had negative spatial association with occasional species and rare species, and common species had positive spatial association with other groups whereas the significant spatial associations of 24 common species with other groups were mainly positive. Conclusion Most species and groups in the community were affected by habitat heterogeneity. At the same time, niche differentiation weakened the competition between species or groups, playing an important role in their coexistence. The community is temporarily in a stable state. [Ch, 3 fig. 3 tab. 33 ref.] -
Key words:
- size structure /
- spatial distribution pattern /
- spatial association /
- groups /
- dominant species /
- common species /
- occasional species /
- rare species
-
图 3 不同类群之间的空间关联变化
Figure 3 Change of spatial association between each two different groups
表 1 24种植物的空间格局
Table 1. Spatial distribution patterns of 24 species
物种 径级结构 完全空间随机模型 异质性泊松过程 聚集分布 随机分布 均匀分布 P 聚集分布 随机分布 均匀分布 P 杜鹃 Rhododendron simsii 单柱型 20 5 0 0.005 4 21 0 0.045 短尾越橘 Vaccinium carlesii “L”型 1 24 0 0.040 0 25 0 0.010 峨眉鼠刺 Itea omeiensis “L”型 5 20 0 0.005 1 24 0 0.005 格药柃 Eurya muricata “L”型 24 1 0 0.005 6 19 0 0.005 海桐山矾 Symplocos heishanensis “L”型 0 25 0 0.545 0 25 0 0.240 红楠 Machilus thunbergii “L”型 25 0 0 0.005 3 21 1 0.005 麂角杜鹃 Rhododendron latoucheae 稳定型 21 4 0 0.005 0 25 0 0.060 檵木 Loropetalum chinense “L”型 19 6 0 0.005 11 6 8 0.005 马银花 Rhododendron ovatum “L”型 20 4 1 0.005 18 6 1 0.005 毛花连蕊茶 Camellia fraterna “L”型 25 0 0 0.005 6 19 0 0.005 南酸枣 Choerospondias axillaris 间歇型 19 6 0 0.005 0 25 0 0.095 拟赤杨 Alniphyllum fortunei “L”型 9 15 1 0.005 7 16 2 0.010 青冈 Quercus glauca “L”型 18 7 0 0.005 3 22 0 0.040 山槐 Albizia kalkora 稳定型 0 25 0 0.480 0 25 0 0.365 杉木 Cunninghamia lanceolata “L”型 19 6 0 0.005 10 14 1 0.005 石斑木 Rhaphiolepis indica 单柱型 1 24 0 0.005 0 25 0 0.490 石栎 Lithocarpus glaber 间歇型 1 24 0 0.005 1 24 0 0.005 石木姜子 Litsea elongata var. faberi 单柱型 4 21 0 0.005 1 24 0 0.010 甜槠 Castanopsis eyrei 间歇型 2 23 0 0.040 0 25 0 0.985 微毛柃 Eurya hebeclados “L”型 22 3 0 0.005 13 10 2 0.005 细枝柃 Eurya loquaiana “L”型 17 8 0 0.005 6 19 0 0.005 小果冬青 Ilex micrococca 稳定型 0 25 0 0.305 0 25 0 0.330 窄基红褐柃 Eurya rubiginosa var. attenuata “L”型 25 0 0 0.005 14 9 2 0.005 浙江新木姜子 Neolitsea aurata var. chekiangensis “L”型 1 24 0 0.050 0 25 0 0.475 说明:聚集分布、随机分布和均匀分布列下的数字为相应格局的尺度数之和。  下载: 导出CSV
下载: 导出CSV
表 2 不同类群与24种植物的空间关联
Table 2. Spatial association between different groups and 24 species
物种间的空间关联 不同尺度下显著空间关联的物种对占总物种对的比例/% 总物种对 0~10 m 11~25 m 0~25 m 负相关 正相关 负相关 正相关 负相关 正相关 优势种与24种植物 0.00 25.00 12.50 4.17 12.50 27.08 48 偶见种与24种植物 0.00 16.67 2.08 4.17 2.08 16.67 48 稀有种与24种植物 0.00 2.08 4.17 4.17 4.17 6.25 48 24种植物之间 1.63 9.96 1.99 7.79 2.90 13.41 552
下载: 导出CSV
表 3 不同类群与生境因子的相关性
Table 3. Correlation between different groups and environmental factors
不同类群 最优模型参数 估计值 标准误 P 赤池信息量准则(AIC) 模型P 优势种 截距 32.056 13.688 0.030 170.128 <0.001 坡度 0.847 2.160 0.699 坡向 −1.915 1.676 0.267 岩石裸露率 −4.143 1.987 0.050 碳含量 3.915 2.194 0.090 常见种 截距 294.240 54.339 <0.001 244.723 <0.001 坡度 0.777 14.567 0.958 坡向 −15.223 12.143 0.224 岩石裸露率 −26.870 13.016 0.052 磷含量 33.214 13.896 0.027 偶见种 截距 9.612 1.049 <0.001 148.895 0.001 坡向 3.451 1.078 0.004 岩石裸露率 −1.294 1.020 0.219 磷含量 −2.532 1.122 0.035 稀有种 截距 2.120 0.407 <0.001 110.297 0.005 碳含量 −1.252 0.415 0.006
下载: 导出CSV
-
[1] SFENTHOURAKIS S, TZANATOS E, GIOKAS S. Species co-occurrence: the case of congeneric species and a causal approach to patterns of species association [J]. Global Ecology and Biogeography, 2006, 15(1): 39 − 49. [2] 储诚进, 王酉石, 刘宇, 等. 物种共存理论研究进展[J]. 生物多样性, 2017, 25(4): 345 − 354. CHU Chengjin, WANG Youshi, LIU Yu, et al. Advances in species coexistence theory [J]. Biodiversity Science, 2017, 25(4): 345 − 354. [3] 达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J]. 植物生态学报, 2004, 28(3): 376 − 384. DA Liangjun, YANG Yongchuan, SONG Yongchang. Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park, Zhejiang Province, eastern China [J]. Acta Phytoecologica Sinica, 2004, 28(3): 376 − 384. [4] LI Lin, HUANG Zhongliang, YE Wanhui, et al. Spatial distributions of tree species in a subtropical forest of China [J]. Oikos, 2009, 118(4): 495 − 502. [5] JOHNSON C N, BALMFORD A, BROOK B W, et al. Biodiversity losses and conservation responses in the Anthropocene [J]. Science, 2017, 356(6335): 270 − 274. [6] ZHU Yan, GETZIN S, WIEGAND T, et al. The relative importance of Janzen-Connell effects in influencing the spatial patterns at the Gutianshan subtropical forest [J/OL]. PLoS One, 2013, 8(9): e74560[2022-08-01]. doi: 10.1371/journal.pone.0074560. [7] NGUYEN H H, URIA-DIEZ J, WIEGAND K. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam [J]. Journal of Vegetation Science, 2016, 27(2): 318 − 327. [8] WIEGAND T, GUNATILLEKE S, GUNATILLEKE N. Species associations in a heterogeneous Sri Lankan dipterocarp forest [J]. American Naturalist, 2007, 170(4): E77 − E95. [9] 安璐, 吴兆飞, 范春雨, 等. 长白山次生杨桦林种群空间点格局及密度制约效应[J]. 生态学报, 2021, 41(4): 1461 − 1471. AN Lu, WU Zhaofei, FAN Chunyu, et al. Spatial point patterns and effects of density dependence in secondary poplar-birch forest, Changbai Mountains, China [J]. Acta Ecologica Sinica, 2021, 41(4): 1461 − 1471. [10] 郭屹立, 王斌, 向悟生, 等. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析[J]. 生物多样性, 2015, 23(2): 183 − 191. GUO Yili, WANG Bin, XIANG Wusheng, et al. Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China [J]. Biodiversity Science, 2015, 23(2): 183 − 191. [11] 王云泉, 田磊, 仲磊, 等. 东白山自然保护区木荷-马尾松群落结构及物种多样性分析[J]. 浙江大学学报(理学版), 2015, 42(1): 38 − 46. WANG Yunquan, TIAN Lei, ZHONG Lei, et al. Community structure and species diversity of Schima superba-Pinus massoniana communities in Dongbaishan Nature Reserve [J]. Journal of Zhejiang University (Science Edition), 2015, 42(1): 38 − 46. [12] LAROCHE F, VIOLLE C, TAUDIERE A, et al. Analyzing snapshot diversity patterns with the Neutral Theory can show functional groups’ effects on community assembly [J/OL]. Ecology, 2020, 101(4): e2977[2022-08-01]. doi: 10.1002/ecy.2977. [13] 马志波, 黄清麟, 庄崇洋, 等. 典型中亚热带天然阔叶林群落各乔木亚层的空间格局与关联性[J]. 林业科学, 2017, 53(12): 12 − 19. MA Zhibo, HUANG Qinglin, ZHUANG Chongyang, et al. Spatial pattern and association among sub-layers of typical mid-subtropical natural broad-leaved forest community [J]. Scientia Silvae Sinicae, 2017, 53(12): 12 − 19. [14] 李皓, 姜景民, 邵文豪, 等. 浙江九龙山自然保护区粉叶柿群落的结构特征[J]. 林业科学研究, 2009, 22(3): 360 − 366. LI Hao, JIANG Jingmin, SHAO Wenhao, et al. Community structure of Diospyros glaucifolia in Jiulongshan Nature Reserve, Zhejiang, China [J]. Forest Research, 2009, 22(3): 360 − 366. [15] 王昌腾. 浙江省九龙山国家级自然保护区珍稀濒危植物调查研究[J]. 安徽农业科学, 2005, 33(2): 250 − 251. WANG Changteng. Rare and endangered plants of Jiulongshan Natural Reserve in Zhejiang Province [J]. Journal of Anhui Agricultural Sciences, 2005, 33(2): 250 − 251. [16] CONDIT R. Tropical Forest Census Plots Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots [M]. Berlin Heidelberg: Springer-Verlag, 1998. [17] 戎福仁, 王祖华, 吴初平, 等. 生境片段化对千岛湖次生马尾松林土壤理化性质的影响[J]. 浙江大学学报(理学版), 2018, 45(6): 748 − 755, 764. RONG Furen, WANG Zuhua, WU Chuping, et al. Effects of habitat fragmentation on soil properties of the secondary Masson pine (Pinus massoniana) forests of the Thousand Island Lake region [J]. Journal of Zhejiang University (Science Edition), 2018, 45(6): 748 − 755, 764. [18] 丁晖, 方炎明, 杨青, 等. 武夷山中亚热带常绿阔叶林样地的群落特征[J]. 生物多样性, 2015, 23(4): 479 − 492. DING Hui, FANG Yanming, YANG Qing, et al. Community characteristics of a mid-subtropical evergreen broad-leaved forest plot in the Wuyi Mountains, Fujian Province, southeastern China [J]. Biodiversity Science, 2015, 23(4): 479 − 492. [19] WIEGAND T, MOLONEY K A. Rings, circles, and null-models for point pattern analysis in ecology [J]. Oikos, 2004, 104(2): 209 − 229. [20] GU Li, O’HARA K L, LI Weizhong, et al. Spatial patterns and interspecific associations among trees at different stand development stages in the natural secondary forests on the Loess Plateau, China [J]. Ecology and Evolution, 2019, 9(11): 6410 − 6421. [21] 王进, 朱江, 艾训儒, 等. 湖北木林子保护区山矾属植物空间分布格局及其关联性[J]. 生态学报, 2020, 40(21): 7709 − 7720. WANG Jin, ZHU Jiang, AI Xunru, et al. Spatial distribution pattern and intraspecific and interspecific relationships of genus Symplocosin Mulinzi Nature Reserve, Hubei Province [J]. Acta Ecologica Sinica, 2020, 40(21): 7709 − 7720. [22] ZHANG Lingyu, DONG Lingbo, LIU Qiang, et al. Spatial patterns and interspecific associations during natural regeneration in three types of secondary forest in the central part of the greater Khingan Mountains, Heilongjiang Province, China [J/OL]. Forests, 2020, 11(2): 152[2022-08-01]. doi: 10.3390/f11020152. [23] WANG Xugao, WIEGAND T, HAO Zhanqing, et al. Species associations in an old-growth temperate forest in north-eastern China [J]. Journal of Ecology, 2010, 98(3): 674 − 686. [24] CRAWLEY M J. The R Book[M]. 2nd ed. West Sussex: John Wiley & Sons, 2012. [25] 刘海丰, 薛达元, 桑卫国. 地形因子对暖温带森林群落物种丰富度-地上生物量关系的影响[J]. 生态环境学报, 2012, 21(8): 1403 − 1407. LIU Haifeng, XUE Dayuan, SANG Weiguo. Effect of topographic factors on the relationship between species richness and aboveground biomass in a warm temperate forest [J]. Ecology and Environmental Sciences, 2012, 21(8): 1403 − 1407. [26] R Development Core Team. R: A Language and Environment for Statistical Computing [M]. Vienna: R Foundation for Statistical Computing, 2017. [27] ZHAO Bingqing, GUO Donggang, SHAO Hongbo, et al. Investigating the population structure and spatial pattern of restored forests in an opencast coal mine, China [J/OL]. Environmental Earth Sciences, 2017, 76(20): 679[2022-08-01]. doi: 10.1007/s12665-017-7020-z. [28] 周赛霞, 江明喜, 鲍大川, 等. 后河自然保护区珍稀植物群落结构及更新特性[J]. 广西植物, 2011, 31(2): 209 − 216. ZHOU Saixia, JIANG Mingxi, BAO Dachuan, et al. Population structure and regenerative characteristics of major tree species of rare plant community in Houhe Nature Reserve [J]. Guihaia, 2011, 31(2): 209 − 216. [29] 李文良, 张小平, 郝朝运, 等. 湘鄂皖连香树种群的年龄结构和点格局分析[J]. 生态学报, 2009, 29(6): 3221 − 3230. LI Wenliang, ZHANG Xiaoping, HAO Chaoyun, et al. Age structure and point pattern analysis of four Cercidiphyllum japonicum populations in three provinces of Hunan, Hubei and Anhui [J]. Acta Ecologica Sinica, 2009, 29(6): 3221 − 3230. [30] 刘春生, 刘鹏, 张志祥, 等. 九龙山南方铁杉群落物种多样性及乔木种种间联结性[J]. 生态环境, 2008, 17(4): 1533 − 1540. LIU Chunsheng, LIU Peng, ZHANG Zhixiang, et al. Species diversity and interspecific association of trees of Tsuga tchekiangensis communities in Jiulong Mountain [J]. Ecology and Environment, 2008, 17(4): 1533 − 1540. [31] 黄阶华, 吴林芳, 钟慧聪, 等. 广东南山保护区常绿阔叶林主要树种空间分布格局和空间关联[J]. 林业与环境科学, 2021, 37(1): 36 − 42. HUANG Jiehua, WU Linfang, ZHONG Huicong, et al. Spatial distribution patterns and interspecific relationships of tree species of evergreen broad-leaved forest at Nanshan Natural Reserve of Guangdong [J]. Forestry and Environmental Science, 2021, 37(1): 36 − 42. [32] ZHOU Quan, SHI Hang, SHU Xiao, et al. Spatial distribution and interspecific associations in a deciduous broad-leaved forest in north-central China [J]. Journal of Vegetation Science, 2019, 30(6): 1153 − 1163. [33] 杨秀清, 史婵, 王旭刚, 等. 关帝山云杉次生林不同生活型物种与生境相关性[J]. 生态学杂志, 2017, 36(6): 1481 − 1487. YANG Xiuqing, SHI Chan, WANG Xugang, et al. Correlation between different life-form species and habitat in secondary Picea forest [J]. Chinese Journal of Ecology, 2017, 36(6): 1481 − 1487. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220521
 点击查看大图
点击查看大图
计量
- 文章访问数: 1595
- HTML全文浏览量: 383
- PDF下载量: 60
- 被引次数: 0
