




-
森林生态系统是地球上最重要的陆地生态系统,不仅直接提供人类生存所必需的各种资源,还在森林固碳和生物多样性保护等方面扮演至关重要的角色[1−3]。在全球气候变暖和降水格局变化的大背景下,森林群落的物种组成、群落结构甚至生态系统功能和生产力都发生了显著改变[1, 4]。落叶阔叶林是北半球一种极为重要的森林类型,对提供生态系统服务并缓解由人类活动引起的气候变化具有关键作用[5]。相较分布于中欧、西欧以及北美东部的落叶阔叶林,东亚地区的落叶阔叶林具有较高的生物多样性和特有物种数量[6]。中国落叶阔叶林是东亚地区落叶阔叶林的重要组成部分,涵盖了从半干旱地区、半湿润地区到湿润地区的广泛气候梯度[7]。一般来说,常绿阔叶林被认为是亚热带地区典型的地带性植被,但在中高海拔的亚热带区域也经常分布一定面积的落叶阔叶林[8]。以往关于亚热带森林的研究很少关注落叶阔叶林,研究中国亚热带地区典型落叶阔叶林群落组成及其结构特征,对于更全面地认识该地区森林生态系统的组成特点和优化森林群落保护策略具有一定的补充作用[9],对完善落叶阔叶林群落研究具有重要意义。
森林群落的物种组成及群落结构特征是群落生态学研究的基础,对阐释物种共存和多样性保护至关重要[10−12]。群落中大多数物种的多度较低,仅有少数物种为常见种[13]。作为森林群落物种组成的另一关键要素,物种的空间分布是物种本身及其与环境相互作用的结果,常被视为探究种群动态和生物多样性维持机制的关键[14]。此外,群落多样性也是生物群落的重要特征,反映了植物群落对环境变化的响应,多样性的变化影响群落结构、功能和演替动态[15]。同时,森林群落在水平和垂直方向上的结构在描述群落特征方面的作用也不容忽视[16],径级结构可以反映林木的胸径、干形等基本状况,以及群落的更新状态和物种与环境之间的相互关系[17],森林群落的垂直分层结构不仅促使光照等生境条件在群落内部发生分化,而且为众多物种的共存提供了丰富的生态位空间,从而影响群落的生物多样性[18−19]。
自巴拿马巴洛科罗拉多岛(Barro Colorado Island)热带雨林大样地建立以来,国内外对森林群落的组成和结构的研究取得了显著进展,在生物多样性维持、物种的空间分布格局以及群落的动态变化等方面展开了卓有成效的研究[20−22]。近20 a来,随着古田山亚热带常绿阔叶林24 hm2监测样地[23]、鼎湖山南亚热带常绿阔叶林20 hm2监测样地[24]和西双版纳热带雨林20 hm2监测样地[25]等的陆续建立,中国目前已建成了29个主样地和近60个面积1~5 hm2的辅助样地(来源于中国科学院生物多样性委员会),与此同时,对温带地区落叶阔叶林群落的研究也在逐步深入,如在中国百花山国家级自然保护区[26]、秦岭长青国家级自然保护区[27]、北京小龙门国家级森林公园[28]、浙江龙王山[29]等自然保护地,也建立了落叶阔叶林的动态监测样地。婺城南山省级自然保护区是目前浙江省内面积最大的省级自然保护区,其中落叶阔叶林是保护区重点保护的森林生态系统,对其中高海拔区域落叶阔叶林物种组成和群落结构的研究尚未见报道。本研究以婺城南山1 hm2固定样地为调查对象,分析样地内物种组成、区系分布、物种多样性、群落径级结构和群落垂直结构等特征,以期为婺城南山省级自然保护区的生物多样性和特色植被资源的保护和动态研究提供基础数据和科学依据。
-
浙江婺城南山省级自然保护区(28°43′56.021″~28°54′8.069″N,119°23′0.081″~119°30′45.113″E)成立于2020年,地处金华市婺城区南山白沙溪源头,以婺城区沙畈乡南部中山区为主,南部与武义县、遂昌县接壤,占地面积为9 532.60 hm2,是目前浙江省内面积最大的省级自然保护区,是保护中亚热带中高海拔落叶阔叶林及穿山甲Manis javanica、黄腹角雉Tragopan caboti和永瓣藤Monimopetalum chinense等珍稀濒危野生动植物的森林生态类型自然保护区。南山保护区的山体多为火山岩覆盖,岩性坚硬,地表残坡积层深厚。气候属于中亚热带湿润季风气候,干湿季分明,夏季高温多雨,冬季温和少雨,适宜林木生长,中高海拔处分布着大面积的落叶阔叶林及针阔混交林,森林覆盖率达94.4%[30]。
-
婺城南山1 hm2长期动态监测样地(以下简称婺城南山样地)选址于保护区内大西坑区域,位于金华婺城区南端,28°46′3.7″N,119°24′30.6″E,海拔为1 012~1 103 m。根据中国森林生物多样性监测网络(CForBio)[31]和《浙江省自然保护地生物多样性长期监测样地建设和调查技术规范(试行)》的建设规范,婺城南山样地以南北、东西方向为边界,用全站仪和森林罗盘仪将整个样地标定成25个20 m × 20 m的样方,在20 m交汇点处设置永久标桩(以西南角为原点编号为0000,向东和向北依次增加,东北角编号为2020,用每个样方东北角编号作为该样方的编号),每个样方划分成16个5 m × 5 m的小样方,在5 m交汇点处插入聚氯乙烯(PVC)管,依据PVC管和标桩对样地进行拉线分隔,于2023年完成样地建设和调查工作。
样地建设完成后,用红色油漆在1.3 m处标记该样地内所有胸径(DBH)≥1 cm的存活个体,以20 m × 20 m样方为基本调查单位,5 m × 5 m小样方为基本测树单元,对样地内所有DBH≥1 cm个体进行定位、物种鉴定、挂牌标记,并调查记录样方号、吊牌号、物种名称、胸径、坐标、树高、枝下高、生长状况等信息,以便进行长期监测。
-
重要值是反映物种在群落中重要性的定量指标,用于比较不同群落间某一物种在群落中的重要性[32],能够反映某一种在群落当中的地位和作用。对婺城南山样地群落中不同植物物种的重要值[33]进行计算,确定群落优势物种。参照HUBBELL等[34]的定义划分稀有种,在本研究中将物种数量为1的物种判定为稀有种。
-
采用Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)、Simpson多样性指数(D)3个多样性指数[35]来描述南山样地内群落的α多样性。
-
群落的年龄结构以径级结构代替[36],以1.0 cm为起测径级,根据婺城南山木本植物的基本情况,确定以2.5 cm为递增单元,划分成13个区间,最大区间为31.0 cm及以上。并以1.0 cm为作图单元,最大作图区间为33.0~34.0 cm。
测量群落内木本植物的树高,划分林层并统计,林层的具体划分参照天目山常绿落叶阔叶林群落[18],按照婺城南山实际情况将样地群落垂直结构划分为3层:灌木层(0~4.2 m)、亚冠层(4.3~7.6 m)、林冠层(7.7~13.4 m)。
-
根据调查结果统计,在婺城南山样地内DBH ≥ 1 cm的木本植物总计有39科65属97种。其中,裸子植物有三尖杉Cephalotaxus fortunei、杉木Cunninghamia lanceolata和黄山松Pinus taiwanensis 3种,被子植物有36科62属94种,以蔷薇科Rosaceae、杜鹃花科Ericaceae和樟科Lauraceae的木本植物物种数最多,3科种数占总种数的27.84%,其次为冬青科Aquifoliaceae、壳斗科Fagaceae、忍冬科Caprifoliaceae和山茶科Theaceae的木本植物,占总种数的18.56%,有17科含1属1种,占全部种数的17.53%。
样地内有71种落叶阔叶木本植物(73.20%),23种常绿阔叶木本植物(23.71%),3种常绿针叶树种(3.09%)。婺城南山样地内落叶阔叶木本植物物种的比例显著高于常绿阔叶木本植物和针叶类木本植物,样地物种的组成以落叶阔叶树种为主。
-
参考文献[37−39],在婺城南山样地内,木本植物科的热带成分略高于温带成分(热带成分18科,温带成分14科)。植物属的分布以温带成分为主,总计39属(60.00%),其中北温带20属(30.77%);热带植物成分24属(36.92%),其中泛热带11属(16.92%);在样地内有中国特有属和世界分布属各1属(表1)。
表 1 婺城南山1 hm2固定样地木本植物区系类型
Table 1. Flora types for woody plants in the 1 hm2 forest plot in Nanshan Nature Reserve
分布区类型 科数 属数 世界广布 1. 广布 7 1 热带 2. 泛热带 11 11 3. 东亚及热带南美间断 5 3 4. 旧世界热带 0 3 5. 热带亚洲至热带大洋洲 0 1 6. 热带亚洲至热带非洲 1 1 7. 热带亚洲 1 5 温带 8. 北温带 10 20 9. 东亚及北美间断 2 11 10. 旧世界温带 0 0 11. 温带亚洲 0 0 12. 地中海区、西亚至中亚 0 0 13. 中亚 0 0 14. 东亚 2 8 中国特有 15. 中国特有 0 1 总计 39 65 -
样地内DBH≥1 cm的木本植物有11 019株(不包括分枝和根萌),映山红Rhododendron simsii的个体数量达2 686株(占植物总株数的24.38%),石灰花楸Sorbus folgneri和短柄枹栎Quercus serrata的个体数量分别为1 371株(12.44%)和1 073株(9.74%)。由表2可见:多度前10位的物种株数总和为7 793 株(70.72%),多度前20位的物种株数总和为9 616株(87.27%)。另外在对样地物种的统计中发现,三尖杉、乌饭Vaccinium bracteatum、华东野胡桃Juglans mandshurica var. formosana、大叶冬青Ilex latifolia、野山楂Crataegus cuneata、杨梅Morella rubra、构树Broussonetia papyifera、棘茎楤木Aralia echinocaulis、楤木Aralia elata、短尾越橘Vaccinium carlesii、短梗冬青Ilex buergeri、石楠Photinia serratifolia、秀丽槭Acer cordatum、紫果槭Acer cordatum、美丽胡枝子Lespedeza thunbergii subsp. formosa、蓝果树Nyssa sinensis、豆腐柴Premna microphylla等物种在该样地中只有1株,是该群落的稀有种。
表 2 婺城南山1 hm2固定样地木本植物多度排序(前10位)
Table 2. Ranking of top ten woody plant abundance in the 1 hm2 forest plot in Nanshan Nature Reserve
种名 多度 显著度 频度 映山红 Rhododendron simsii 2 686 3.15 1.00 石灰花楸 Sorbus folgneri 1 371 16.93 0.88 短柄枹栎 Quercus serrata 1 073 44.52 1.00 山橿 Lindera reflexa 547 1.59 1.00 包石栎 Lithocarpus cleistocarpus 449 38.21 0.88 宜昌荚蒾 Viburnum erosum 399 2.24 0.92 黄檀 Dalbergia hupeana 350 6.84 0.96 化香 Platycarya strobilacea 322 84.47 0.96 山胡椒 Lindera glauca 307 6.17 1.00 粗枝绣球 Hydrangea robusta 289 7.89 0.64 婺城南山1 hm2样地落叶阔叶林群落中,重要值(表3)最高的是短柄枹栎,其值为0.115,其重要值受到相对显著度的影响较大,该树种在婺城南山落叶阔叶林群落中处于优势竞争位,其他重要值较高的物种有映山红、石灰花楸、化香Platycarya strobilacea和包石栎Lithocarpus cleistocarpus,重要值分别为0.104、0.086、0.061和0.048,短柄枹栎、映山红和石灰花楸3种落叶树种是该群落的优势木本植物;样地内所有落叶树种的重要值总和为0.709,常绿物种和针叶物种的重要值分别为0.180和0.111,落叶树种的重要值远大于常绿及针叶树种,可见该样地为典型落叶阔叶林。
表 3 婺城南山1 hm2固定样地木本植物重要值排序(前10位)
Table 3. Ranking of important values of top ten woody plants in the 1 hm2 forest plot in Nanshan Nature Reserve
种名 相对
多度相对显
著度相对
频度重要值 短柄枹栎 Quercus serrata 0.097 0.218 0.030 0.115 映山红 Rhododendron simsii 0.244 0.039 0.030 0.104 石灰花楸 Sorbus folgneri 0.124 0.106 0.026 0.086 化香 Platycarya strobilacea 0.029 0.124 0.029 0.061 包石栎 Lithocarpus cleistocarpus 0.041 0.078 0.026 0.048 钟花樱 Prunus campanulata 0.021 0.052 0.030 0.034 山合欢 Albizia kalkora 0.023 0.051 0.029 0.034 山橿 Lindera reflexa 0.050 0.004 0.030 0.028 杉木 Cunninghamia lanceolata 0.011 0.047 0.020 0.026 红果钓樟 Lindera erythrocarpa 0.014 0.034 0.029 0.025 -
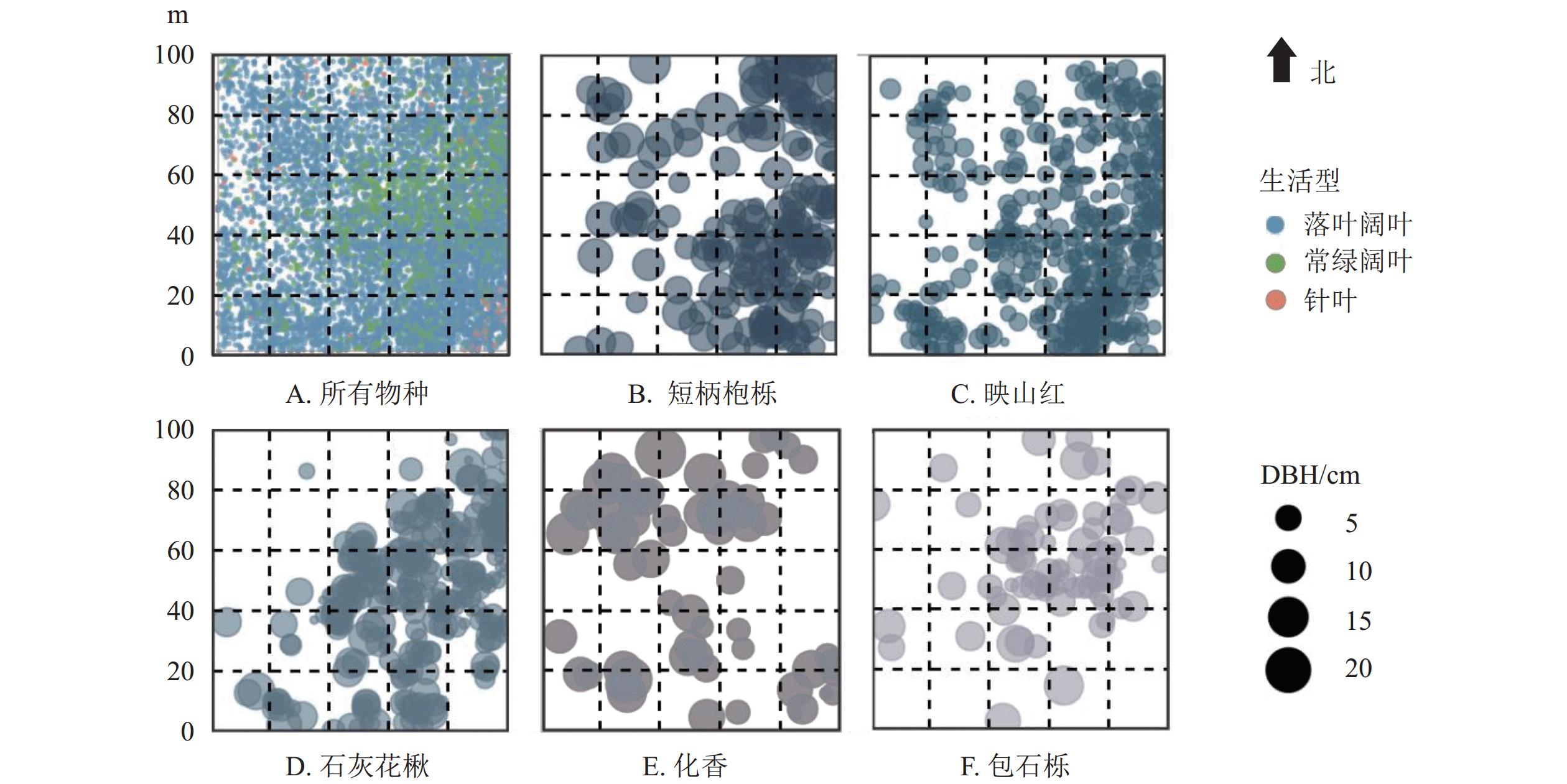
婺城南山样地植株空间分布如图1。图1A表示所有物种在样地内的空间分布格局,样地东南区域木本植物个体较为密集,样地西北角区域木本植物分布相对分散,聚集程度相对其他区域较低。从具体生活型看,落叶阔叶物种在各样方中分布范围广,在样地中较为常见;常绿阔叶树种分布范围小于落叶阔叶物种,在样地内分布比较分散,主要集中于样地中部和东部地区;针叶树种在样地中分布范围较小,主要集中于样地北侧和东南角。图1B~F为婺城南山样地中重要值前5位的物种的空间分布图,斑点位置代表该物种在样地中的位置,斑点大小代表植株的胸径大小。总体而言,5个优势种在样地内分布比较广泛,不同物种在样地分布的聚集区域具有一定的差别。除西侧略少见外,短柄枹栎和映山红在样地中总体较常见;石灰花楸和包石栎在样地内分布范围小于短柄枹栎和映山红,具有相似的分布范围;化香在样地中分布范围较小,在部分区域出现聚集分布。

图 1 婺城南山1 hm2固定样地物种空间分布
Figure 1. Spatial distribution of species in the 1 hm2 forest plot in Nanshan Nature Reserve
-
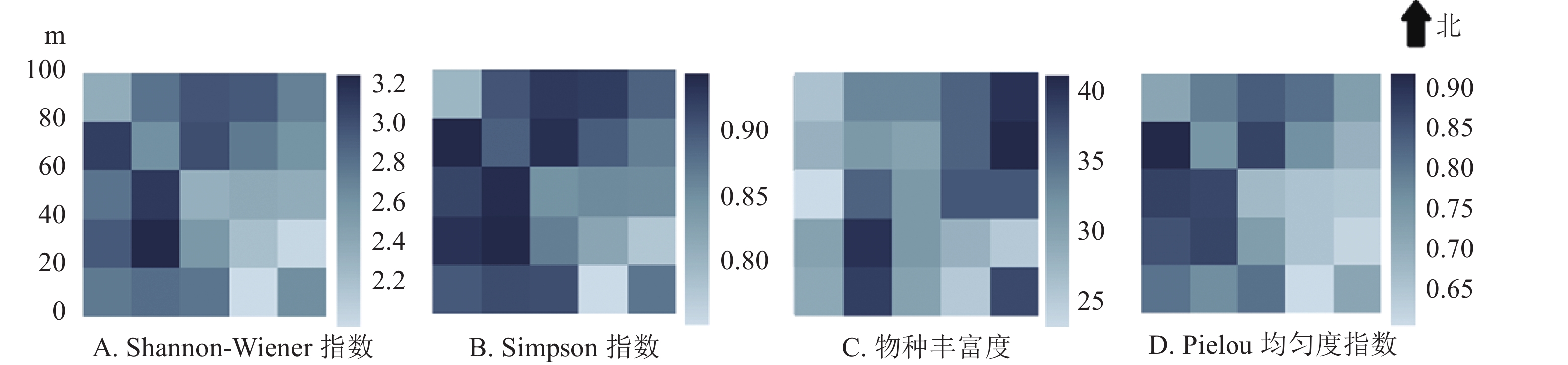
物种多样性指标如图2。Simpson多样性指数与Shannon-Wiener多样性指数在整体上具有一致性,在样地东南角的1604和2008样方数值最低,在样地西南方的0808样地数值最高,整体上样地东侧的物种多样性指数略低于西侧。Pielou均匀度指数在大样地的西南部相对较高。物种丰富度在2016样方达到最高(样地东北角),在0808样方和2004样方物种丰富度也相对较高。
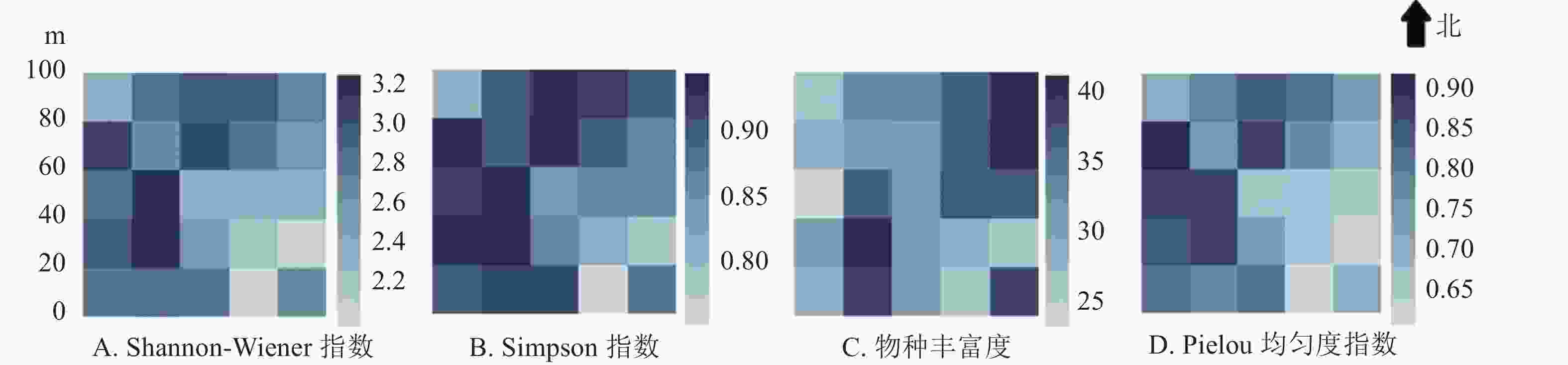
图 2 婺城南山1 hm2固定样地20 m × 20 m样方物种多样性指数热图
Figure 2. Heatmap of species diversity index in the 1 hm2 forest plot in Nanshan Nature Reserve
-
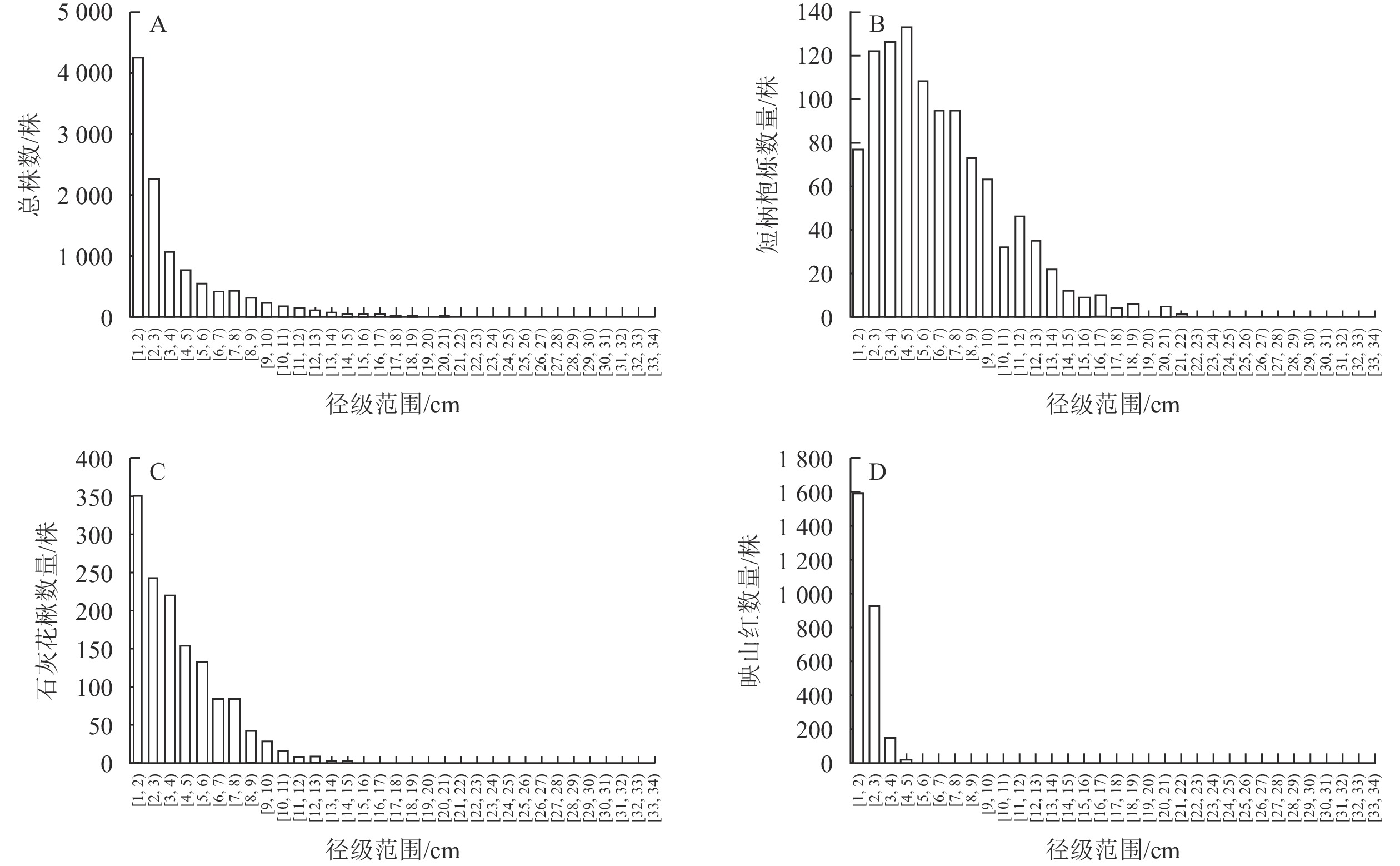
统计样地内植物胸径,发现样地木本植物胸径总体较小,植株平均胸径为3.66 cm,其中DBH<6 cm和DBH<11 cm的小径级和较小径级植株分别占个体总数的80.78%和95.16%,DBH≥16 cm的植株仅占个体总数的0.83%。
婺城南山样地内群落径级结构整体呈倒“J”型(图3A),最大的胸径数值为32.3 cm,该物种为杉木。物种数量在1~6 cm的胸径区间内存在较大程度的下降,但整体上并未出现明显的断层。3种优势物种中,短柄枹栎(图3B)和石灰花楸(图3C)在不同径级结构中都比较常见,97.80%的映山红胸径在3.5 cm以下(图3D)。短柄枹栎和石灰花楸在样地中表现出小乔木的特征,在群落中幼苗或幼树的储备量比较充足,较小或较大径级的植株都比较充裕,映山红则表现出灌木的性状,较大径级植株数量较少。
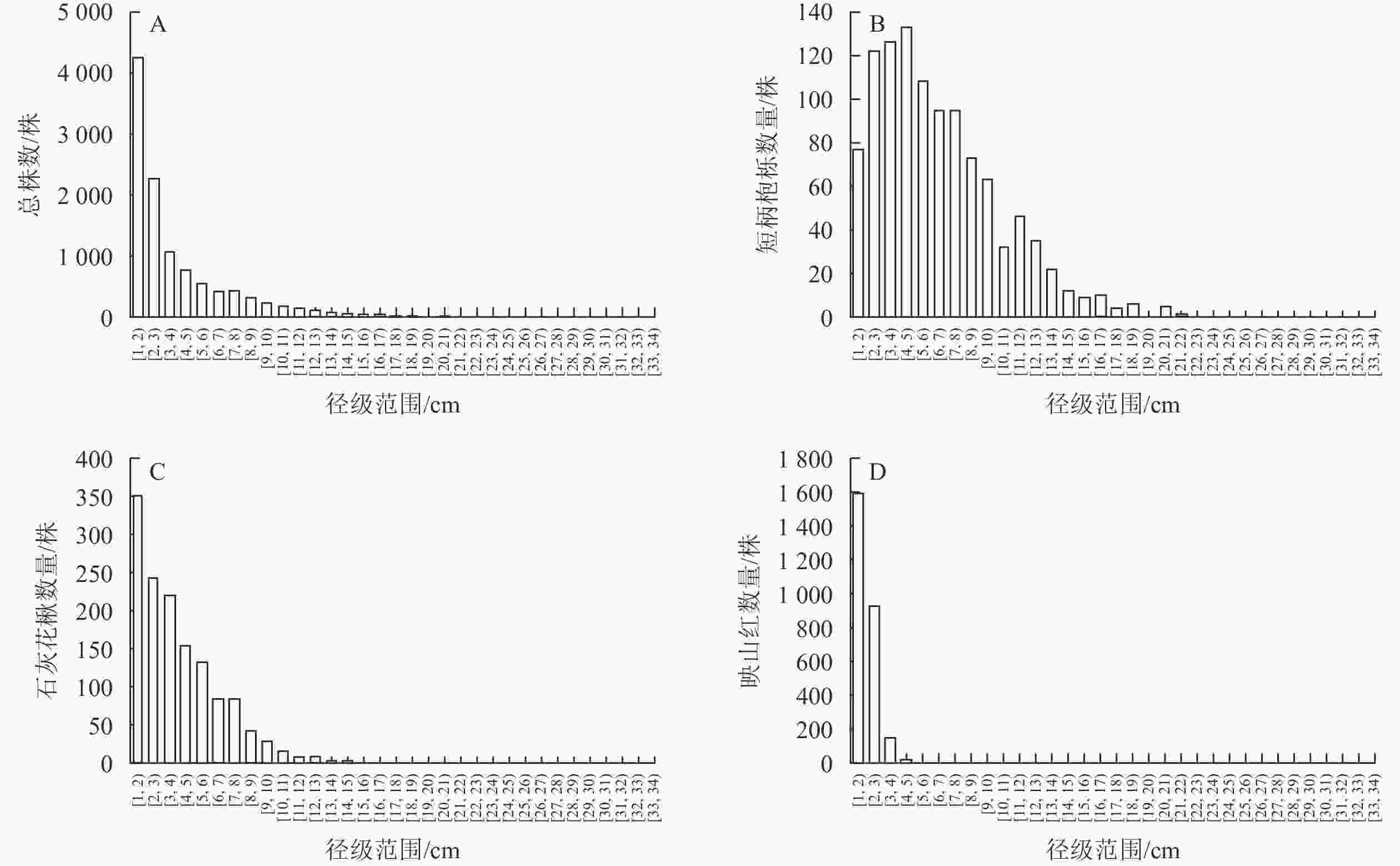
图 3 婺城南山1 hm2固定样地木本植物径级结构分布图
Figure 3. Distribution of species DBH class structure in the 1 hm2 forest plot in Nanshan Nature Reserve
-
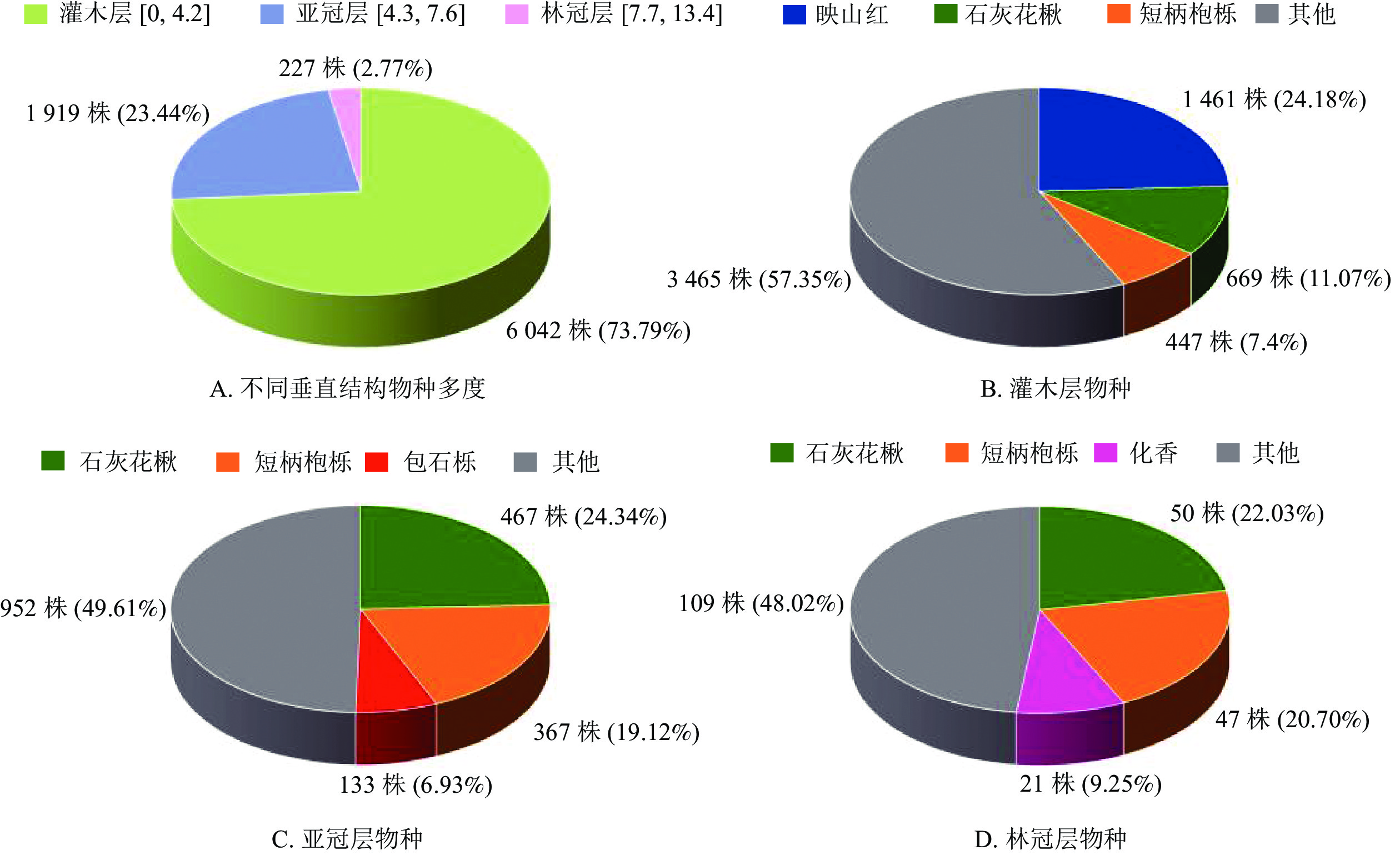
排除受地形或外力因素导致倾斜角度过大或倒伏的木本植物,用于垂直结构分析的植物共8 188株。婺城南山样地中树高范围在灌木层的植物有6 042株(73.79%),其中,映山红、石灰花楸和短柄枹栎的数目较多,分别占该区间植物数的24.18%、11.07%和7.40%(图4A和图4B)。婺城南山样地中树高范围在亚冠层的植物有1 919株(23.44%),其中,石灰花楸、短柄枹栎和包石栎分别占所在林层植株数量的24.34%、19.12%和6.93%,在亚冠层中数量较多(图4A和图4C)。处于林冠层的植物总计227株(2.77%),植株总数相对较少,其中,石灰花楸、短柄枹栎和化香3种落叶树种分别占所在林层植株数量的22.03%、20.70%和9.25%(图4A和图4D)。
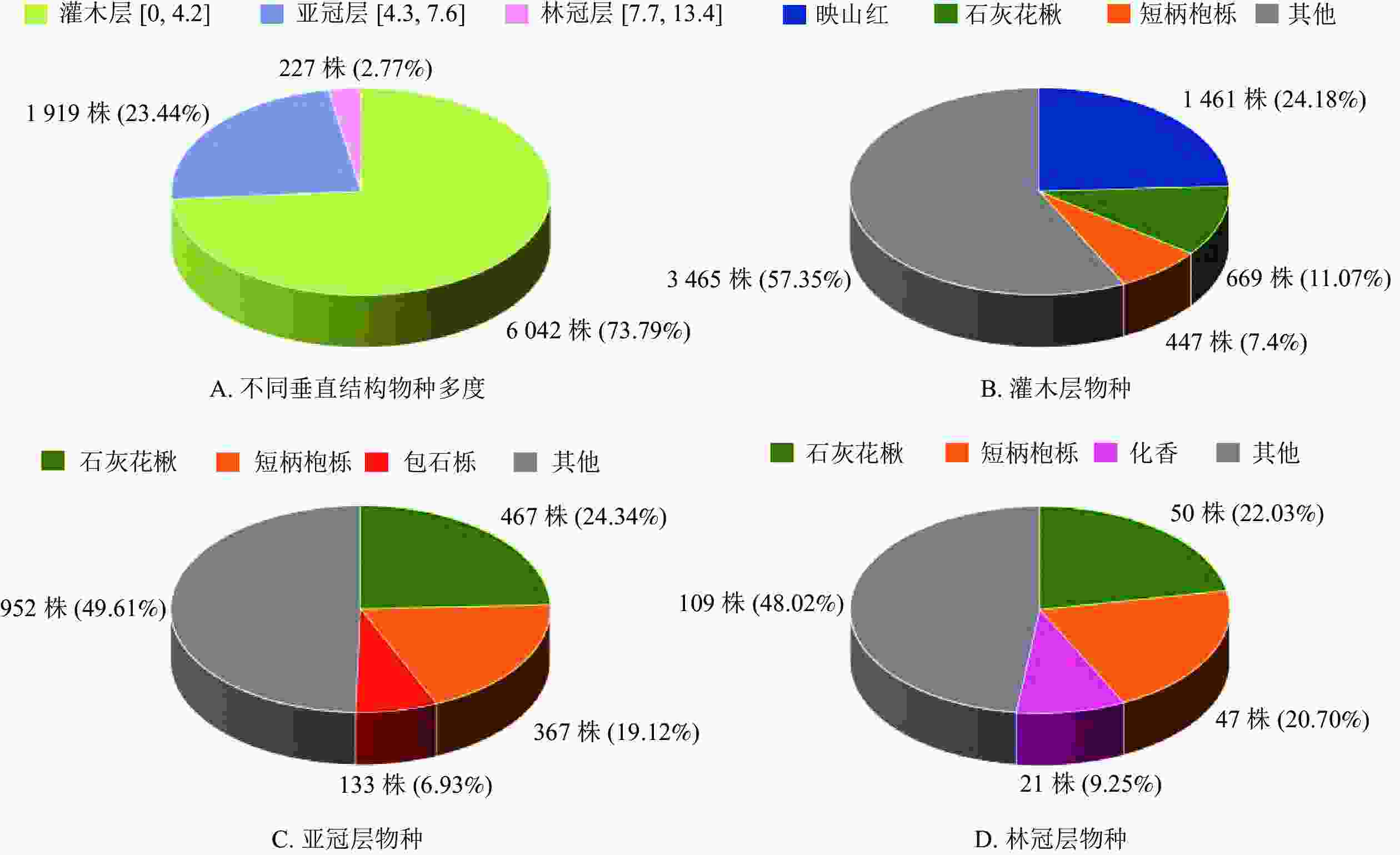
图 4 婺城南山1 hm2固定样地不同垂直结构主要树种及数量统计
Figure 4. Distribution of species vertical structure in the 1 hm2 forest plot in Nanshan Nature Reserve
婺城南山样地大部分木本植物处于灌木层,其中,映山红和短柄枹栎在灌木层中的数量较多,可认为是灌木层中的优势种;在该群落的亚冠层中,石灰花楸和短柄枹栎处于优势地位;群落的林冠层中,以石灰花楸、短柄枹栎和化香为主。调查发现,样地中下坡位还发现一定数量的林间层藤本植物,如密花崖豆藤Millettia congestiflora、中华猕猴桃Actinidia chinensis、葛藤Argyreia pierreana、大芽南蛇藤Celastrus gemmatus等,其中,密花崖豆藤相对较常见。
-
经调查统计发现:婺城南山样地内共有木本植物39科65属97种,表明该群落的生物多样性丰富。群落中落叶物种、常绿物种和针叶物种的重要值分别为70.91%、17.95%和11.14%,意味着该群落为典型的落叶阔叶林。与调查方法类似且纬度接近的湖北神农架25 hm2样地比较,神农架样地平均密度仅为2 442株·hm−2[9],而婺城南山样地的木本植物总计11 019 株,植株密度为神农架样地的4.5倍。从数据统计层面分析,神农架样地以大径级米心水青冈Fagus engleriana和锐齿槲栎Quercus aliena var. acuteserrata为主要建群种[40],群落发育时间较长,植株个体胸径较大,其竞争强度受胸径影响较大[41],进而呈现平均密度小于婺城南山样地的结果。此外,婺城南山样地生物多样性程度可能与样地地理位置与气候条件有关。金华南山整体位于东部季风区[42],样地降水丰富,地形、地貌受海拔影响复杂多样,生境异质性高,为维持生物多样性提供了基础条件。
从物种的区系成分来看,婺城南山样地内物种的温带成分是热带成分的1.63倍,温带分布属的比例显著高于热带,和纬度接近的古田山24 hm2大样地[43]的属级成分存在明显区别。这可能和2个样地的植被类型和物种组成有关。古田山样地植被为原生的常绿阔叶林,物种以常绿阔叶种类为主,且样地海拔相对较低,而南山1 hm2样地地处海拔为1 012~1 103 m 的山脊,较高的海拔使得婺城南山样地气温低于古田山样地,植被为典型山地落叶阔叶林。婺城南山样地物种区系成分以温带占优的现象,可能反映了植被分布的垂直地带性特征[44]。
木本植物的径级和垂直结构揭示了群落的年龄特征,也反映了可能的演替历程和影响因素[45]。婺城南山样地中,中小径级的个体在群落中占明显优势,和纬度相近的百山祖样地[46]相比,径级分布总体规律类似(均为倒“J”型),说明婺城南山群落在一定时间内具有良好的自我更新能力,但婺城南山样地的木本植物胸径普遍小于百山祖样地。
从垂直结构看,婺城南山样地73.79%的植株处于灌木层,23.44%的植株处于亚冠层,说明婺城南山样地木本植物树高总体偏矮小,这与浙江乌岩岭国家自然保护区的结果[47]存在差异,在乌岩岭保护区内,随着树木的增高,乔木树种占比先增加而后缓慢下降。径级结构与垂直结构均反映了婺城南山落叶阔叶林群落树龄结构偏小的特点,演替正处于初期阶段,这可能是受到人为因素干扰的结果。从婺城南山样地的周边情况看,样地南侧的遂昌郑岗岭林场育有大面积长势良好的大径级杉木林和少量的黄山松,北侧不远处有少量胸径大于20 cm的甜槠Castanopsis eyrei和木荷Schima superba,周边沟谷地带零星分布有杉木,推测婺城南山样地可能在早期是以甜槠和木荷为优势种的群落,原生林遭遇人为干扰破坏(人为砍伐)后,树龄结构较大的树种数量减少,落叶树种逐步占据优势,演变为落叶阔叶林。现阶段,婺城南山样地中,短柄枹栎、石灰花楸等落叶物种在林冠层、亚冠层和灌木层的重要值占比较高,推测该群落在一定时期内仍将继续维持落叶阔叶林原貌,未来是否有可能向常绿落叶阔叶混交林或其他植被类型演替,尚需进一步的动态观察和研究。
-
浙江婺城南山森林样地是典型的落叶阔叶林群落,群落物种多样性丰富。虽然科的分布温带与热带比例接近,但是属的分布温带成分占据明显优势。群落径级结构呈现倒“J”型,群落中物种径级总体偏小,植株密度较大。垂直方向上,灌木层和亚冠层占优势,分层不明显。婺城南山落叶阔叶林的物种组成和群落结构在中亚热带中高海拔地区有一定的代表性。
-
感谢婺城南山省级自然保护区管理中心,浙江大学于明坚、毛志斌,浙江省森林资源监测中心陈征海,以及浙江师范大学杨依丹、柴澎涛、金航锋、叶永强、沈彬宇、郑林尧、汪骋翼等在样地选择、建设和调查过程中给予帮助。
Species composition and community structure of deciduous broad-leaved forest in Wucheng Nanshan Nature Reserve, Zhejiang Province
-
摘要:
目的 浙江婺城南山自然保护区在植被分区属于中亚热带常绿阔叶林带,但在其中高海拔区域分布着典型的落叶阔叶林群落,是保护区重点保护的森林生态系统。分析浙江婺城南山1 hm2动态监测样地物种组成及群落结构等特征,为保护区的生物多样性和特色植被资源的保护和动态研究提供参考,也为研究中国亚热带地区落叶阔叶林群落提供基础数据。 方法 基于婺城南山1 hm2固定样地的调查数据,具体分析了该群落物种组成、区系特征、物种多样性指数、群落的径级结构与垂直结构等信息。 结果 ①样地内共有木本植物39科65属97种(裸子植物3科3属3种,被子植物36科62属94种),其中蔷薇科Rosaceae、杜鹃花科Ericaceae和樟科Lauraceae所占物种数最多;②样地内木本植物区系成分中,属级层面温带成分为热带成分的1.63倍,温带成分明显占优势;③样地内木本植物总计11 019株,短柄枹栎Quercus serrata、映山红Rhododendron simsii和石灰花楸Sorbus folgneri重要值居前3位,是该群落的优势物种;群落中落叶物种、常绿物种和针叶物种的重要值分别为0.709、0.180和0.111,意味着该群落为典型的落叶阔叶林;④样地内植株平均胸径3.66 cm,径级结构呈现倒“J”型,小径级和较小径级的植物数量占比达95.16%。⑤群落垂直结构上,灌木层、亚冠层和林冠层的植株占比分别为73.79%、23.44%和2.77%,这说明该群落灌木层植株比例较高且林冠层不明显。 结论 浙江婺城南山1 hm2固定样地为典型的落叶阔叶林,群落的物种多样性丰富,植株密度较大,但群落中植株径级总体较小,垂直结构分层不明显,在中亚热带中高海拔地区有一定的代表性。图4表3参47 Abstract:Objective The Wucheng Nanshan Nature Reserve in Zhejiang Province belongs to the mid-subtropical evergreen broad-leaved forest zone in the vegetation zone, while the typical deciduous broad-leaved forest community is distributed in medium and high altitude area, which is the forest ecosystem under the protection of the Reserve. By analyzing the species composition and community structure characteristics of the 1 hm2 forest plot in Nanshan Nature Reserve, this study provides scientific basis for the protection and dynamic research of biodiversity and characteristic vegetation resources in the nature reserve, and also contributes foundational data for the study of deciduous broad-leaved forest communities in the subtropical areas of China. Method Based on the survey data from 1 hm2 forest plot in the Nanshan Nature Reserve, species composition, flora characteristics, the species diversity index, diameter class structure and vertical structure of the community were analyzed. Result (1) There were 97 woody species belong to 39 families, 65 genera (3 gymnosperms species belong to 3 families and 3 genera, 94 angiosperms species belong to 36 families and 62 genera), among which Rosaceae, Ericaceae and Lauraceae accounted for the largest number of species. (2) From a perspective of the flora, the temperate component was approximately 1.63 times as much as the tropical component at the generic level, and the temperate component was dominant. (3) A total of 11 019 woody individuals were found in the plot. Quercus serrata, Rhododendron simsii and Sorbus folgneri were the top three dominant species in the community. The important values of the deciduous species, evergreen species and coniferous species were 0.709, 0.180 and 0.111, respectively, indicating that the community was a typical deciduous broad-leaved forest. (4) The DBH size-class distribution followed a reverse “J” shape with an average diameter of only 3.66 cm, and the individuals at the minor diameter class accounted for 95.16% of the total population. (5) The proportion of plants in shrub layer, sub-canopy layer and canopy layer was 73.79%, 23.44% and 2.77%, respectively, indicating that shrub plants occupied a higher proportion in the vertical structure and the forest canopy layer was not obvious. Conclusion Nanshan forest plot in Zhejiang Province is a typical deciduous broad-leaved forest with rich species diversity and high plant density, but the plant diameter class in the community is generally small and the vertical structure is not obvious. This community is representative in the middle and high altitudes of the mid-subtropical regions. [Ch, 4 fig. 3 tab. 47 ref.] -
图 1 婺城南山1 hm2固定样地物种空间分布
Figure 1 Spatial distribution of species in the 1 hm2 forest plot in Nanshan Nature Reserve
图 2 婺城南山1 hm2固定样地20 m × 20 m样方物种多样性指数热图
Figure 2 Heatmap of species diversity index in the 1 hm2 forest plot in Nanshan Nature Reserve
图 3 婺城南山1 hm2固定样地木本植物径级结构分布图
Figure 3 Distribution of species DBH class structure in the 1 hm2 forest plot in Nanshan Nature Reserve
图 4 婺城南山1 hm2固定样地不同垂直结构主要树种及数量统计
Figure 4 Distribution of species vertical structure in the 1 hm2 forest plot in Nanshan Nature Reserve
表 1 婺城南山1 hm2固定样地木本植物区系类型
Table 1. Flora types for woody plants in the 1 hm2 forest plot in Nanshan Nature Reserve
分布区类型 科数 属数 世界广布 1. 广布 7 1 热带 2. 泛热带 11 11 3. 东亚及热带南美间断 5 3 4. 旧世界热带 0 3 5. 热带亚洲至热带大洋洲 0 1 6. 热带亚洲至热带非洲 1 1 7. 热带亚洲 1 5 温带 8. 北温带 10 20 9. 东亚及北美间断 2 11 10. 旧世界温带 0 0 11. 温带亚洲 0 0 12. 地中海区、西亚至中亚 0 0 13. 中亚 0 0 14. 东亚 2 8 中国特有 15. 中国特有 0 1 总计 39 65  下载: 导出CSV
下载: 导出CSV
表 2 婺城南山1 hm2固定样地木本植物多度排序(前10位)
Table 2. Ranking of top ten woody plant abundance in the 1 hm2 forest plot in Nanshan Nature Reserve
种名 多度 显著度 频度 映山红 Rhododendron simsii 2 686 3.15 1.00 石灰花楸 Sorbus folgneri 1 371 16.93 0.88 短柄枹栎 Quercus serrata 1 073 44.52 1.00 山橿 Lindera reflexa 547 1.59 1.00 包石栎 Lithocarpus cleistocarpus 449 38.21 0.88 宜昌荚蒾 Viburnum erosum 399 2.24 0.92 黄檀 Dalbergia hupeana 350 6.84 0.96 化香 Platycarya strobilacea 322 84.47 0.96 山胡椒 Lindera glauca 307 6.17 1.00 粗枝绣球 Hydrangea robusta 289 7.89 0.64
下载: 导出CSV
表 3 婺城南山1 hm2固定样地木本植物重要值排序(前10位)
Table 3. Ranking of important values of top ten woody plants in the 1 hm2 forest plot in Nanshan Nature Reserve
种名 相对
多度相对显
著度相对
频度重要值 短柄枹栎 Quercus serrata 0.097 0.218 0.030 0.115 映山红 Rhododendron simsii 0.244 0.039 0.030 0.104 石灰花楸 Sorbus folgneri 0.124 0.106 0.026 0.086 化香 Platycarya strobilacea 0.029 0.124 0.029 0.061 包石栎 Lithocarpus cleistocarpus 0.041 0.078 0.026 0.048 钟花樱 Prunus campanulata 0.021 0.052 0.030 0.034 山合欢 Albizia kalkora 0.023 0.051 0.029 0.034 山橿 Lindera reflexa 0.050 0.004 0.030 0.028 杉木 Cunninghamia lanceolata 0.011 0.047 0.020 0.026 红果钓樟 Lindera erythrocarpa 0.014 0.034 0.029 0.025
下载: 导出CSV
-
[1] KING A W, HAYES D J, HUNTZINGER D N, et al. North American carbon dioxide sources and sinks: magnitude, attribution, and uncertainty [J]. Frontiers in Ecology and the Environment, 2012, 10(10): 512−519. [2] WU Anchi, XIONG Xin, ZHOU Guoyi, et al. Climate change-related biodiversity fluctuations and composition changes in an old-growth subtropical forest: a 26-yr study [J/OL]. Science of the Total Environment, 2024, 914 : 169899[2024-07-01]. DOI: 10.1016/j.scitotenv.2024.169899. [3] LIU Meng, TRUGMAN A T, PEÑUELAS J, et al. Climate-driven disturbances amplify forest drought sensitivity [J]. Nature Climate Change, 2024, 14(7): 746−752. [4] WU Yu, WANG Dongya, QIAO Xiujuan, et al. Forest dynamics and carbon storage under climate change in a subtropical mountainous region in central China [J/OL]. Ecosphere, 2020, 11 (3): e03072[2024-07-01]. DOI: 10.1002/ecs2.3072. [5] XU Tingting, ZHANG Anzhi, XU Xiyan, et al. Synchronized slowdown of climate warming and carbon sink enhancement over deciduous broadleaf forests based on FLUXNET analysis [J/OL]. Ecological Indicators, 2023, 155 : 111042[2024-07-01]. DOI: 10.1016/j.ecolind.2023.111042. [6] CHEN Guoping, CAI Qiong, MA Suhui, et al. Climate and forest attributes influence above-ground biomass of deciduous broadleaf forests in China [J]. Journal of Ecology, 2023, 111(2): 495−508. [7] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980. WU Zhengyi. Chinese Vegetation [M]. Beijing: Science Press, 1980. [8] HAI Lisi, LI Xiaoqian, ZHANG Jingbo, et al. Assembly dynamics of East Asian subtropical evergreen broadleaved forests: new insights from the dominant Fagaceae trees [J]. Journal of Integrative Plant Biology, 2022, 64(11): 2126−2134. [9] 魏嘉欣, 姜治国, 杨林森, 等. 湖北神农架中亚热带山地落叶阔叶林25 ha动态监测样地群落物种组成与结构特征[J]. 生物多样性, 2024, 32(3): 5−15. WEI Jiaxin, JIANG Zhiguo, YANG Linsen, et al. Community composition and structure in a 25 ha mid-subtropical mountain deciduous broad-leaved forest dynamics plot in Shennongjia, Hubei, China [J]. Biodiversity Science, 2024, 32(3): 5−15. [10] HOOPER D U, CHAPIN F S, EWEL J J, et al. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge [J]. Ecological Monographs, 2005, 75(1): 3−35. [11] 马克平. 森林动态大样地是生物多样性科学综合研究平台[J]. 生物多样性, 2017, 25(3): 227−228. MA Keping. Forest dynamics plot is a crosscutting research platform for biodiversity science [J]. Biodiversity Science, 2017, 25(3): 227−228. [12] 马克平, 徐学红. 中国森林生物多样性监测网络有力支撑生物群落维持机制研究[J]. 中国科学: 生命科学, 2020, 50(4): 359−361. MA Keping, XU Xuehong. Chinese forest biodiversity monitoring network well developed for community assembly studies [J]. Scientia Sinica Vitae, 2020, 50(4): 359−361. [13] 王世雄, 赵亮, 李娜, 等. 稀有种和常见种对植物群落物种丰富度格局的相对贡献[J]. 生物多样性, 2016, 24(6): 658−664. WANG Shixiong, ZHAO Liang, LI Na, et al. The relative contributions of rare and common species to the patterns of species richness in plant communities [J]. Biodiversity Science, 2016, 24(6): 658−664. [14] 温智峰, 魏识广, 李林, 等. 亚热带不同纬度森林群落物种空间分布格局[J]. 生态学报, 2022, 42(22): 9153−9164. WEN Zhifeng, WEI Shiguang, LI Lin, et al. Spatial distribution patterns of forest community species at different latitudes in the subtropics [J]. Acta Ecologica Sinica, 2022, 42(22): 9153−9164. [15] 林阳, 李时轩, 周伟龙, 等. 百山祖国家公园植物群落α和β多样性对海拔梯度的响应[J]. 生态学报, 2024, 44(17): 7700−7712. LIN Yang, LI Shixuan, ZHOU Weilong, et al. α and β diversity patterns of woody plant communities along an elevation gradient in Baishshanzu National Park [J]. Acta Ecologica Sinica, 2024, 44(17): 7700−7712. [16] 李建, 李晓宇, 曹静, 等. 长白山次生针阔混交林群落结构特征及群落动态[J]. 生态学报, 2020, 40(4): 1195−1206. LI Jian, LI Xiaoyu, CAO Jing, et al. Community structure and dynamics of secondary coniferous and broad-leaved mixed forest in Changbai Mountains [J]. Acta Ecologica Sinica, 2020, 40(4): 1195−1206. [17] 王永琪, 秦佳双, 马姜明, 等. 广西马尾松人工林林下木本植物径级结构特征[J]. 广西科学, 2020, 27(6): 630−637. WANG Yongqi, QIN Jiashuang, MA Jiangming, et al. Diameter structure characteristics of understory woody species in Pinus massoniana plantations in Guangxi [J]. Guangxi Sciences, 2020, 27(6): 630−637. [18] 楼一恺, 范忆, 戴其林, 等. 天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系[J]. 生态学报, 2021, 41(21): 8568−8577. LOU Yikai, FAN Yi, DAI Qilin, et al. Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of Tianmu Mountain Natural Reserve [J]. Acta Ecologica Sinica, 2021, 41(21): 8568−8577. [19] CHAI Pengtao, XIE Jiajie, YANG Lisheng, et al. Community vertical stratification drives temporal taxonomic and phylogenetic beta diversity in a mixed broadleaf-conifer forest [J/OL]. Frontiers in Ecology and Evolution, 2023, 11 : 1170197[2024-07-01]. DOI: 10.3389/fevo.2023.1170197. [20] CONDIT R. Research in large, long-term tropical forest plots [J]. Trends in Ecology & Evolution, 1995, 10(1): 18−22. [21] MI Xiangcheng, FENG Gang, HU Yibo, et al. The global significance of biodiversity science in China: an overview [J]. National Science Review, 2021, 8(7): 34−58. [22] 冯晓娟, 米湘成, 肖治术, 等. 中国生物多样性监测与研究网络建设及进展[J]. 中国科学院院刊, 2019, 34(12): 1389−1398. FENG Xiaojuan, MI Xiangcheng, XIAO Zhishu, et al. Overview of Chinese biodiversity observation network (Sino BON) [J]. Bulletin of Chinese Academy of Sciences, 2019, 34(12): 1389−1398. [23] 金毅, 于明坚. 古田山24 ha常绿阔叶林样地林窗幼苗更新动态2011—2016[C]//浙江省科学技术学会. 第七届浙江省生物多样性保护研讨会摘要集. 庆元: 第七届浙江省生物多样性保护研讨会, 2017: 17. JIN Yi, YU Mingjian. Seedling regeneration from 2011 to 2016 in 24 ha of evergreen broad-leaved forest sample plot in Gutianshan [C]//Zhejiang Institute of Science and Technology. Abstracts of the Seventh Symposium on Biodiversity Conservation in Zhejiang Province. Qingyuan: The Seventh Symposium on Biodiversity Conservation in Zhejiang Province, 2017: 17. [24] 裴男才. 利用植物DNA条形码构建亚热带森林群落系统发育关系——以鼎湖山样地为例[J]. 植物分类与资源学报, 2012, 34(3): 263−270. PEI Nancai. Building a subtropical forest community phylogeny based on plant DNA barcodes from Dinghushan Plot [J]. Plant Diversity and Resources, 2012, 34(3): 263−270. [25] 邓志云, 李玉武. 热带雨林生物多样性监测与研究样地网络建设实践[J]. 林业调查规划, 2019, 44(4): 60−64. DENG Zhiyun, LI Yuwu. Establishment of biodiversity monitoring and research plot network in tropical rainforest [J]. Forest Inventory and Planning, 2019, 44(4): 60−64. [26] 高永龙, 孙艳丽, 徐铭泽, 等. 北京百花山落叶阔叶林群落内木本植物的叶片功能性状变异特征[J]. 北京林业大学学报, 2024, 46(4): 40−51. GAO Yonglong, SUN Yanli, XU Mingze, et al. Variation characteristics in leaf functional traits of woody plants in deciduous broadleaved forest community in Baihua Mountain of Beijing [J]. Journal of Beijing Forestry University, 2024, 46(4): 40−51. [27] 何春梅, 李雨姗, 尹秋龙, 等. 秦岭皇冠暖温性落叶阔叶林优势树种的径级结构和数量特征[J]. 植物生态学报, 2023, 47(12): 1658−1667. HE Chunmei, LI Yushan, YIN Qiulong, et al. Diameter structure and quantitative characteristics of dominant tree species in warm temperate deciduous broadleaf forest plot in Huangguan of Qinling Mountains, China [J]. Chinese Journal of Plant Ecology, 2023, 47(12): 1658−1667. [28] 邓婷婷, 魏岩, 任思远, 等. 北京东灵山暖温带落叶阔叶林地形和林分结构对林下草本植物物种多样性的影响[J]. 生物多样性, 2023, 31(7): 18−29. DENG Tingting, WEI Yan, REN Siyuan, et al. Effects of topography and stand structure of warm temperate deciduous broad-leaved forest on understory herb diversity in Donglingshan, Beijing [J]. Biodiversity Science, 2023, 31(7): 18−29. [29] 徐建, 韦新良, 王敬, 等. 龙王山落叶阔叶林优势树种的种内种间竞争[J]. 浙江农林大学学报, 2014, 31 (6): 868−876. XU Jian, WEI Xinliang, WANG Jing, et al. Intraspecific and interspecific competition of dominant species in a deciduous, broadleaf forest of Longwang Mountain [J]. Journal of Zhejiang A&F University, 2014, 31 (6): 868−876. [30] 问青青, 叶锋. 自然保护生态南山[J]. 浙江林业, 2020(12): 24−25. WEN Qingqing, YE Feng. Nature conservation, ecological Nanshan [J]. Zhejiang Forestry, 2020(12): 24−25. [31] 徐学红, 王巍伟, 米湘成, 等. 中国森林生物多样性监测网络(CForBio): 20年进展与展望[J]. 生物多样性, 2023, 31(12): 132−146. XU Xuehong, WANG Weiwei, MI Xiangcheng, et al. The Chinese forest biodiversity monitoring network: 20-year achievements and outlook [J]. Biodiversity Science, 2023, 31(12): 132−146. [32] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533−548. FANG Jingyun, WANG Xiangping, SHEN Zehao, et al. Methods and protocols for plant community inventory [J]. Biodiversity Science, 2009, 17(6): 533−548. [33] LINARES-PALOMINO R, PONCE-ALVAREZ S I. Tree community patterns in seasonally dry tropical forests in the Cerros de Amotape Cordillera, Tumbes, Peru [J]. Forest Ecology and Management, 2005, 209(3): 261−272. [34] HUBBELL S, FOSTER R. Commonness and rarity in a neotropical forest: implications for tropical tree conservation [J]. Plant Ecology, 1986, 8 : 205−231. [35] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ: α多样性的测度方法[J]. 生物多样性, 1994, 2(4): 231−239. MA Keping, LIU Yuming. Methods for measuring biodiversity of community: measurement of α diversity [J]. Biodiversity Science, 1994, 2(4): 231−239. [36] 任毅华, 周尧治, 侯磊, 等. 色季拉山急尖长苞冷杉种群不同龄级立木的空间分布格局[J]. 生态学报, 2021, 41(13): 5417−5424. REN Yihua, ZHOU Yaozhi, HOU Lei, et al. Spatial distribution patterns of standing trees at different ages in Abies georgei var. smithii forests in Sejila Mountain [J]. Acta Ecologica Sinica, 2021, 41(13): 5417−5424. [37] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245−257. WU Zhengyi, ZHOU Zhekun, LI Dezhu, et al. The areal-types of the world families of seed plants [J]. Acta Botanica Yunnanica, 2003, 25(3): 245−257. [38] 吴征镒. 《世界种子植物科的分布区类型系统》的修订[J]. 云南植物研究, 2003, 25(5): 535−538. WU Zhengyi. Revision of the distribution types of seed plants in the world [J]. Acta Botanica Yunnanica, 2003, 25(5): 535−538. [39] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13 (增刊4): 1−139. WU Zhengyi. The areal-types of Chinese genuses of seed plants [J]. Acta Botanica Yunnanica, 1991, 13 (suppl 4): 1−139. [40] 王向雨, 胡东, 贺金生. 神农架地区米心水青冈林和锐齿槲栎林生物量的研究[J]. 首都师范大学学报(自然科学版), 2007, 28(2): 62−67. WANG Xiangyu, HU Dong, HE Jinsheng. Biomass research of Fagus engleriana and Quercus aliena var. acuteserrata forest in Shennongjia forest District [J]. Journal of Capital Normal University (Natural Science Edition), 2007, 28(2): 62−67. [41] 刘耀凤, 董利虎, 郝元朔, 等. 竞争对天然针阔混交林树木直径生长的影响[J]. 东北林业大学学报, 2023, 51(9): 1−7. LIU Yaofeng, DONG Lihu, HAO Yuanshuo, et al. Effect of competition on tree growth in natural coniferous and broad-leaved mixed forest [J]. Journal of Northeast Forestry University, 2023, 51(9): 1−7. [42] 张文江. 中国地理大势分析[J]. 学术月刊, 1995, 27(3): 78−82. ZHANG Wenjiang. Analysis of Chinese geographical trend [J]. Academic Monthly, 1995, 27(3): 78−82. [43] 祝燕, 赵谷风, 张俪文, 等. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构[J]. 植物生态学报, 2008, 32 (2): 262−273. ZHU Yan, ZHAO Gufeng, ZHANG Liwen, et al. Community composition and structure of Gutianshan forest dynamic pot in a mid-subtropical evergreen broad-leaved forest, East China [J]. Chinese Journal of Plant Ecology, 2008, 32 (2): 262−273. [44] 刘华训. 我国山地植被的垂直分布规律[J]. 地理学报, 1981, 36(3): 267−279. LIU Huaxun. The vertical zonation of mountain vegetation in China [J]. Acta Geographica Sinica, 1981, 36(3): 267−279. [45] 张芳, 杜虎, 曾馥平, 等. 西南喀斯特峰丛洼地木本植物群落结构与多样性变化[J]. 生态学报, 2020, 40(12): 4094−4104. ZHANG Fang, DU Hu, ZENG Fuping, et al. Changes of woody community structure and diversity in karst peak-cluster depressions in Southwest China [J]. Acta Ecologica Sinica, 2020, 40(12): 4094−4104. [46] 刘啸林, 吴友贵, 张敏华, 等. 浙江百山祖25 ha亚热带森林动态监测样地群落组成与结构特征[J]. 生物多样性, 2024, 32(2): 25−35. LIU Xiaolin, WU Yougui, ZHANG Minhua, et al. Community composition and structure of a 25 ha forest dynamics plot of subtropical forest in Baishanzu, Zhejiang Province [J]. Biodiversity Science, 2024, 32(2): 25−35. [47] 江彦苹, 刘西, 潘向东, 等. 浙江乌岩岭国家级自然保护区毛果青冈群落结构特征及物种多样性[J]. 福建农业科技, 2023, 54(12): 1−6. JIANG Yanping, LIU Xi, PAN Xiangdong, et al. Community structure characteristics and species diversity of Cyclobalanopsis pachyloma in Wuyanling National Nature Reserve of Zhejiang [J]. Fujian Agricultural Science and Technology, 2023, 54(12): 1−6. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240442
 点击查看大图
点击查看大图
计量
- 文章访问数: 1671
- HTML全文浏览量: 478
- PDF下载量: 62
- 被引次数: 0
