







-
森林生物量是森林生态系统的基本特征之一,是研究森林生态系统结构和功能的基础[1],也是评估森林碳收支的主要参数。森林生物量是衡量森林碳储存能力的关键指标,对减缓全球气候变化具有重要的作用[2]。开展森林生物量及其驱动机制研究不仅有助于深入了解森林生态系统结构、功能及动态变化,而且对于提升森林经营管理水平,增强碳汇功能具有重要意义。
近年来,关于森林生物量驱动因素的研究逐渐成为生态学家关注的热点问题。众多研究表明:物种多样性的增加有助于促进森林地上生物量的积累[3−6]。生态位互补假说为这一现象提供了理论解释,认为随着物种多样性的增加,物种间相互作用趋于复杂化,促进了生态位的分化和物种在资源利用上的竞争,提高了生态系统对资源的利用效率[7],进而促进了森林生物量的积累。然而,也有研究指出:物种多样性与生产力的关系可能因生态位重叠和竞争压力的增加而变得不显著甚至呈负相关[8−9]。可见,关于物种多样性对森林生物量的影响机制仍存在较大的争议。
结构多样性为林木个体在空间分布、物种组成和树木大小(树木高度和直径)上异质性的指标[10−11],能够反映森林空间结构(水平和垂直结构)的复杂性[12]。复杂的林分结构能够提供更丰富的生境空间和资源,提高资源的利用效率,减轻竞争压力,进而促进地上生物量的积累[13−14]。有研究认为:相较于物种多样性,结构多样性对森林地上生物量的影响更为显著,物种多样性通过结构多样性间接作用于森林地上生物量[15−16]。
然而,物种多样性和结构多样性并非森林地上生物量的唯一驱动因素,它同时受土壤、地形、气候等非生物因素的影响[17−19]。HOFHANSL等[20]研究指出:在局域尺度上土壤养分是影响森林地上生物量的重要因素。何潇等[21]对退化生态系统林分碳储量的研究发现:气候是林分碳储量变化的主要影响因素。贾勃等[22]对东北针阔混交林的研究表明:海拔和年均气温是影响林分生物量变化的关键非生物因素。综上所述,在研究森林生物量驱动因素时,不能仅考虑单一因素的影响,需要综合考虑生物和非生物因素的相互作用,全面衡量森林地上生物量的多元驱动因素[21, 23]。
蒙古栎Querus mongolica林是中国东北地区典型的植被类型,面积占东北有林地面积的15%~20%[24],在水源涵养、生物多样性保护以及木材供应等方面发挥着重要作用。然而,由于长期的不合理经营,蒙古栎林的生产力和生态功能受到了严重影响。因此,本研究以辽东山区蒙古栎林为研究对象,采用线性回归和结构方程模型,探讨了物种多样性、结构多样性以及土壤养分等对林分乔木生物量的影响,以期为辽东山区蒙古栎林的可持续经营和碳汇功能提升提供理论依据。
-
研究区位于辽宁省抚顺市新宾县境内(41°40′~41°35′N,124°26′~124°20′E),该区属北温带大陆性季风气候区,年均气温为7 ℃,年均降水量为750~850 mm,≥10 ℃的年有效积温为2 300~2 500 ℃,无霜期为150 d。土壤类型以暗棕壤和棕壤为主。主要森林类型为蒙古栎林、阔叶混交林、针阔混交林以及人工林。主要乔木树种有蒙古栎、色木槭Acer mono、胡桃楸Juglans mandshurica、水曲柳Fraxinus mandshurica、落叶松Larix gmelimii、红松Pinus koraiensis、花曲柳F. rhynchophylla、紫椴Tilia amurensis、榆树Ulmus pumila等。主要灌木树种有长白忍冬Lonicera ruprechtiana、卫矛Evonymus alatus、绣线菊Spiraea salicifolia、胡枝子Lespedeza bicolor等[25]。
-
分别于2020和2023年的6—8月,在全面踏查的基础上,按照龄组、地形因子等差异性,选择具有代表性的蒙古栎林,设置25 m×25 m的样地28块,30 m×30 m的样地19块,共计47块样地。对样地内所有胸径(DBH)≥5 cm的树木进行每木检尺,记录树种的名称、位置(x,y)、胸径、树高等。同时利用手持全球定位系统(GPS)测定样地的海拔、坡向、坡位、坡度等信息。样地基本信息见表1。
表 1 样地基本信息
Table 1. Basic information of the sample plots
龄组 胸径/cm 树高/m 林分密度/(株·hm−2) 林分断面积/(m2·hm−2) 幼龄林 11.5~13.5 9.7~11.8 1 256~2 289 20.10~27.06 中龄林 12.9~15.9 9.8~11.0 1 089~1 578 22.39~28.47 近熟林 18.9~24.6 11.1~15.0 688~1 040 20.15~33.03 成熟林 16.2~19.2 10.7~14.7 1 000~1 222 29.23~34.79 过熟林 18.3~20.7 11.4~15.9 733~956 31.69~44.69 -
采用异速生长方程计算样地内所有乔木的生物量。不同异速生长方程模型及其参数设定见《中国主要林木生物量模型手册》[26]。
-
采用物种丰富度、物种香农指数和均匀度指数代表林分物种多样性,计算公式详见参考文献[12]。采用胸径的香农指数、均匀度指数和变异系数代表结构多样性,计算公式详见参考文献[13]。
-
在样方的四角和中心采集0~20 cm土层土壤样品500 g,于实验室内测定全氮、全磷、速效氮、速效磷以及有机质含量。土壤全氮采用凯氏定氮法测定;速效氮采用氢氧化钠水解-碱解扩散法测定;全磷和速效磷采用钼锑抗吸光光度法测定;有机质采用重铬酸钾容量法测定。
-
由于土壤养分因子较多,结构方程模型不容易收敛,因此,采用主成分分析法对土壤养分进行主成分提取。主成分分析结果表明:第1主成分解释了总体变异的89.7%,第2主成分解释了总体变异的7.6%,由于第1主成分的方差贡献率大于70%,因此,选用第1主成分代表土壤养分(图1)。在结构多样性和物种多样性指标的筛选上,选择与林分乔木生物量相关性最大的指标来代表林分结构的复杂性和物种多样性。

图 1 土壤养分主成分分析
Figure 1. Principal component analysis of soil nutrient
利用结构方程模型检验土壤养分、林分密度、结构多样性和物种多样性对林分乔木层生物量的直接和间接效应。首先,假设土壤养分、林分密度、结构多样性和物种多样性直接影响乔木生物量,土壤养分也可通过物种多样性、结构多样性间接调控乔木生物量,建立初始模型。
在构建结构方程模型前,使用Z-score标准化方法对模型中的自变量和响应变量进行标准化处理,以消除不同变量在量纲上的差异。利用方差膨胀因子(VIF)来检验自变量之间的多重共线性。若VIF>10,则表明自变量之间存在较为严重的多重共线性,若VIF<10,则表明自变量之间不存在显著的共线性[13]。验证结果显示:VIF<10,因此可以确认自变量之间不存在多重共线性。
采用最大似然法对结构方程模型进行拟合,在评价模型优度时,利用卡方检验比较拟合优度指数(CFI)、渐进残差均方和平方根(RMSEA)以及显著性概率(P)。拟合优度的临界值一般为CFI>0.9,RMSEA<0.08,P>0.05[27]。
-
主成分分析采用SPSS 21.0,结构方程模型构建采用AMOS 23.0,绘图采用Origin 2019。
-
辽东山区蒙古栎林乔木生物量为276.49 t·hm−2。其中,优势树种蒙古栎的生物量占比最大(83.67%),为231.34 t·hm−2,其次为花曲柳(6.46%)、紫椴(3.81%)、胡桃楸(1.69%)和色木槭(1.42%),这5个树种共占林分乔木总生物量的97.05%,其他树种仅占林分乔木生物量的2.95%(图2A)。可见,优势树种蒙古栎对林分乔木生物量起决定作用。从不同龄组蒙古栎林乔木层生物量特征来看,幼龄林的生物量最低,为178.18 t·hm−2,其次为中龄林,近熟林和成熟林乔木层生物量相差不大,过熟林最高,为409.62 t·hm−2,为近熟林和成熟林的1.4倍(图2B)。表明随着龄级的增加,乔木层生物量呈增加趋势。
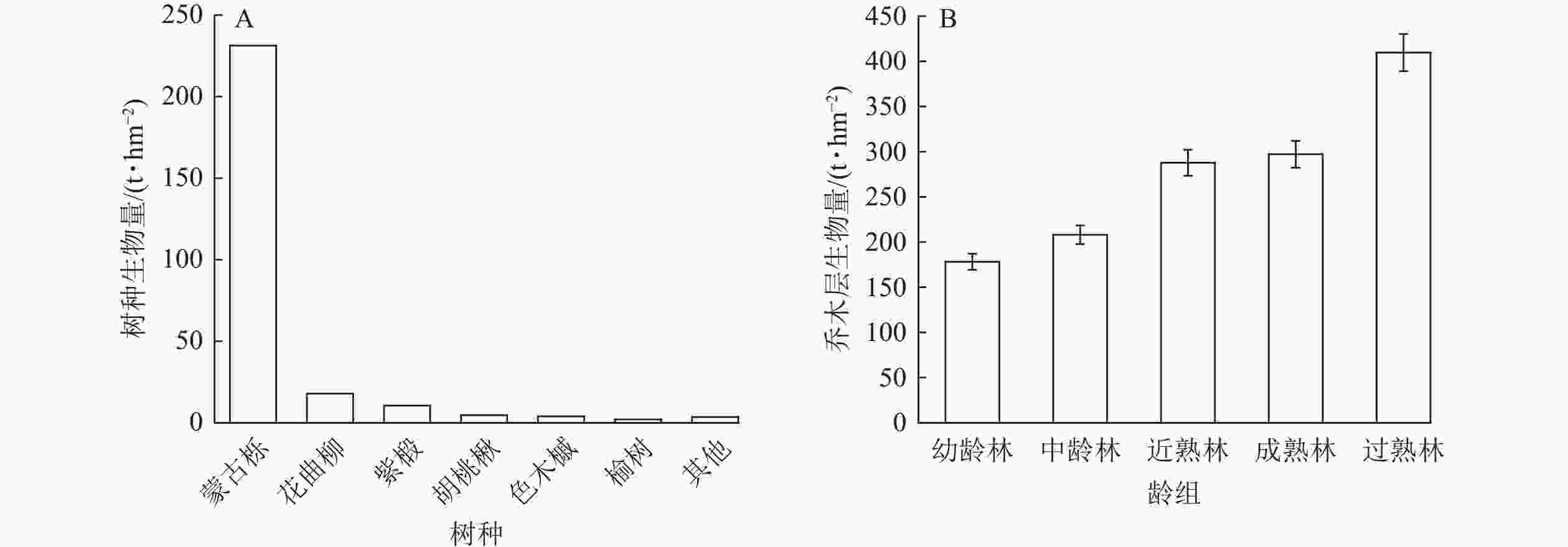
图 2 辽东山区蒙古栎林优势树种和不同龄组乔木层生物量分布特征
Figure 2. Biomass distribution characteristics of dominant tree species and different age-class trees in Q. mongolica forest in the mountainous region of eastern Liaoning Province
-
通过对物种多样性、结构多样性指数与林分乔木生物量做一元线性回归分析(图3)可知:代表物种多样性的丰富度和香农指数均与林分乔木生物量呈极显著负相关(相关系数r=−0.503,P<0.01;r=−0.417,P<0.01),代表结构多样性的胸径香农指数与林分乔木生物量呈极显著正相关(r=0.405,P<0.01),而物种均匀度指数、胸径均匀度指数和胸径变异系数与林分乔木生物量相关性不显著。这表明物种多样性越高,林分乔木生物量越低,同时林分胸径结构多样性越高,林分乔木生物量越高。
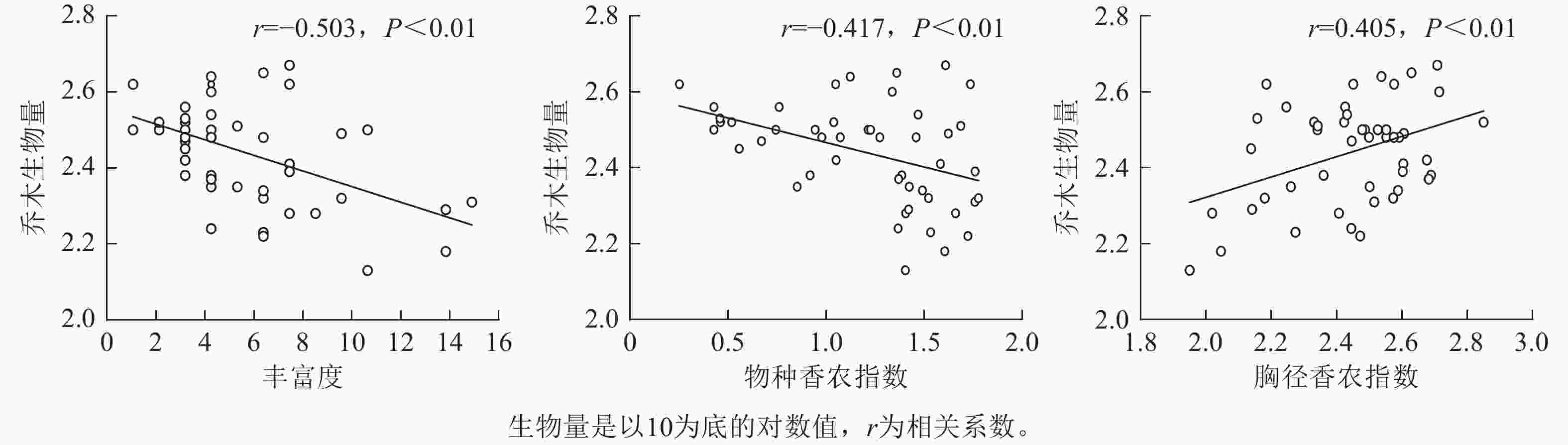
图 3 物种多样性和结构多样性与林分乔木生物量的线性回归分析
Figure 3. Linear regression analysis of species diversity, structural diversity, and stand tree biomass
-
以林分乔木生物量为因变量,物种多样性、结构多样性、土壤养分和林分密度为潜变量构建结构方程模型,最优模型结果(图4和表2)表明:各变量共解释了林分乔木生物量变化的27.7%。林分的物种多样性、结构多样性对乔木生物量均具有显著的直接影响,路径系数分别为−0.394 (P<0.01)和0.281 (P<0.05),土壤养分对林分乔木生物量的直接影响并不显著,主要是通过结构多样性和林分密度对林分乔木生物量产生间接正影响。林分密度对林分乔木生物量的影响也是通过物种多样性和结构多样性产生间接的负影响。
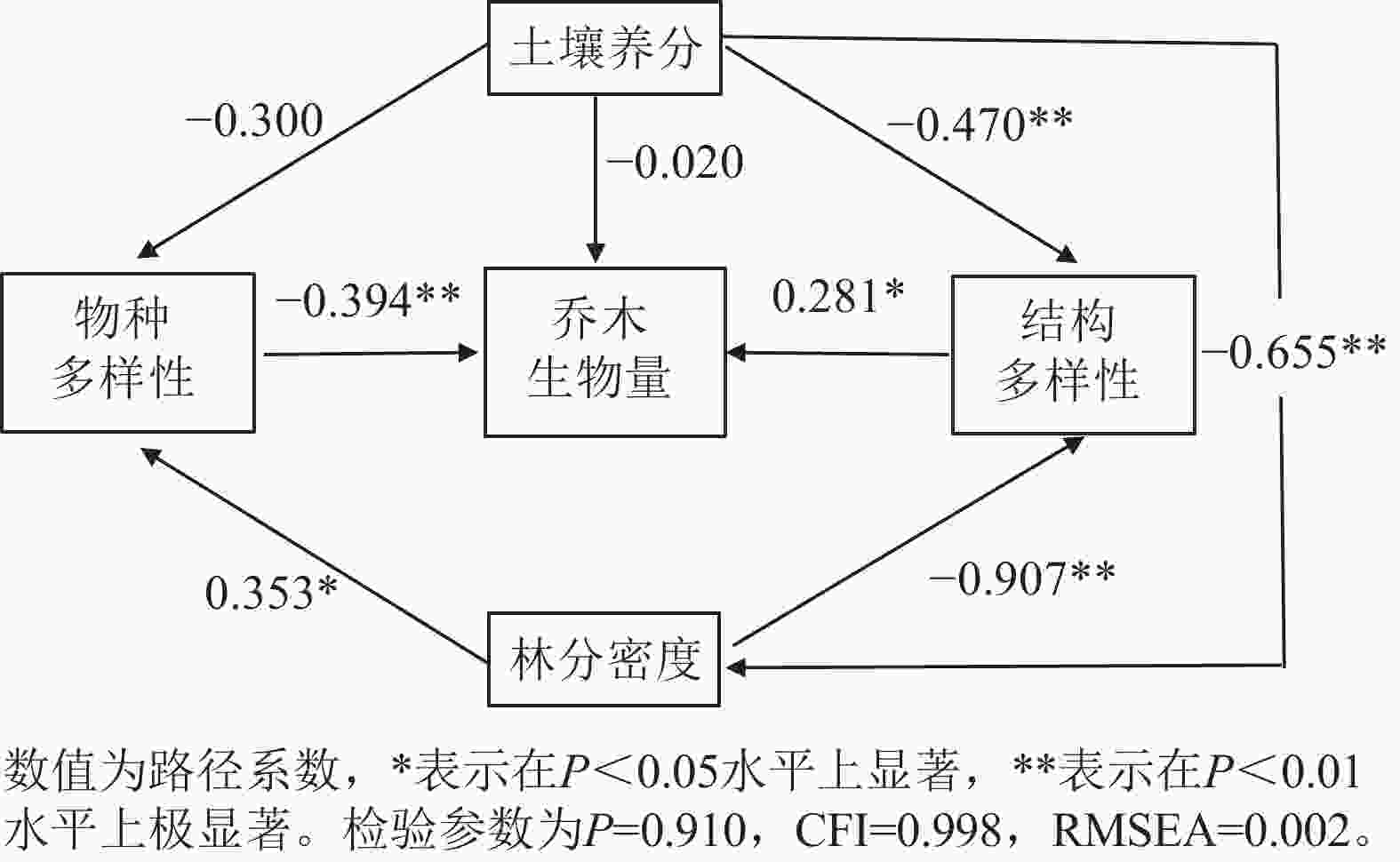
图 4 生物和非生物因素对林分乔木生物量影响的结构方程模型图
Figure 4. Structural equation model diagram of the impact of biotic and abiotic factors
表 2 最优结构方程模型中各解释变量对林分乔木生物量的标准化效益
Table 2. Standardized effects of each explanatory variable in the optimal structural equation model on the biomass of forest stand trees
项目 效应值 结构多样性 物种多样性 土壤养分 林分密度 直接效应 0.281 −0.394 −0.025 0 间接效应 0 0 0.244 −0.393 总效应 0.281 −0.394 0.219 −0.393 -
林分结构多样性是反映树木个体大小差异的关键指标,主要来源于种内或种间个体的非对称竞争机制,同时也受到物种固有生物学特性差异的影响[28−29]。大量研究表明:复杂的林分结构对于促进林分生物量的积累具有显著的正效应[15−16, 22, 30]。本研究也得出相同的结论。一方面,林分结构多样性的增加促进了光资源分配的优化,使生长在中下层的林木通过冠层间隙获得更多的光照,提高了森林对光资源的利用效率,从而有利于林分生物量的积累[27, 31]。另一方面,复杂的林分结构促进了物种的生态位分化,使得不同层次的植物能够更加有效地利用资源,减少竞争压力,进而有利于林分生物量的提高。此外,林分结构多样性增加的同时提高了森林生态系统的稳定性和抗逆性[32],从而有助于维持较高水平的生物量。总之,林分结构可通过合理利用资源、促使生态位分化以及维持生态系统的稳定性和抗逆性等,促进林分生物量的增加。
-
物种多样性与林分生物量之间的关系一直是研究的热点。已有研究普遍认为物种多样性的增加能够通过生态位互补效应促进林分生物量的积累[3−4]。不同物种在资源利用上的差异性,通过生态位分化降低了物种间的竞争,从而提高了林分对资源的整体利用效率[13, 30, 33]。然而,本研究结果与以上结论并不一致。本研究发现:随着物种多样性的增加,林分乔木生物量呈降低趋势。这一结果与朱杰等[11]对南亚热带常绿阔叶林的研究以及谭珊珊等[23]对巴拿马热带雨林的研究结果相一致。造成这一现象的原因可能与研究对象所处的演替阶段有关。前者的研究对象主要处于演替前期,林分结构相对简单,物种间的竞争较弱,物种多样性的增加有利于生态位的分化,从而促进了林分生物量的积累。此外,演替前期,林分中物种数量较少,多以高生产力、低碳储量的演替先锋物种为主,随着物种多样性的增加,群落中包含高碳储量物种的机会增多,从而促进了林分生物量的增加[3, 34]。相反,在演替的中后期(或近成熟林),与生态位互补效应相比,非对称性竞争效应更为显著,林分生物量主要取决于高碳储量的树种,然而高碳储量树种的竞争作用抑制了其他树种对资源的利用[11, 35],从而间接限制了物种多样性的增加,导致物种多样性与生物量呈负相关。此外,森林类型也是影响物种多样性与林分地上生物量关系的重要因素。在蒙古栎林中,蒙古栎作为优势树种,具有较高的生产力和较强的竞争能力,不仅促进了林分生物量的积累,同时也抑制了其他竞争能力较弱的树种,导致物种多样性降低。物种多样性与林分生物量之间的关系还与环境条件、资源可获得性以及研究尺度等因素密切相关[11, 33−39]。吴初平等[3]通过对亚热带次生林的研究指出:在不考虑环境因子时,物种丰富度与林分生物量呈显著的线性正相关,当考虑土壤、地形、光照等环境因子时,物种多样性与林分生物量之间的关系受多种环境因素的共同影响而变得复杂。同样,物种多样性与林分生物量之间的关系还表现出尺度依赖性,即随着空间尺度的增加,物种多样性与林分生物量之间的关系逐渐由正相关转变为不相关,甚至负相关[6, 23]。总之,物种多样性与林分乔木生物量之间的关系具有多维度,且十分复杂,这是演替阶段、森林类型、环境因素、研究尺度等多种因素共同作用的结果。
-
土壤养分作为森林生态系统中非生物因素的重要组成部分,对林分生物量具有显著影响[40−42]。土壤中含有树木生长的必需元素,这些元素有效性的增加对林分生物量的积累具有积极的正效应[9, 43],但也有研究表明:随着土壤养分的增加,林分生物量呈降低趋势[39],这可能是由于土壤养分的增加,加剧了种间竞争,导致树木死亡率提高或树木生长受到抑制[44],从而使林分生物量减少。土壤通过影响树木的生长和存活状况,对林分生物量的累积产生直接影响。同时,它也可能通过影响生物因素,如物种组成、林分结构和功能性状等,间接影响林分生物量的变化[45]。本研究表明:土壤养分对林分乔木生物量的直接作用并不显著,而是通过林分结构(结构多样性和林分密度)对林分乔木生物量产生间接正影响。在蒙古栎林中,相对于土壤养分等环境因子而言,光照可能是林木生长的主要限制因素[46],林木通过增加结构的异质性,增强对光资源的捕获和利用效率,从而间接促进林分生物量的积累。
-
本研究表明:林分结构多样性和物种多样性对林分乔木生物量均有直接影响,林分结构多样性与林分乔木生物量呈正相关,而物种多样性与林分乔木生物量呈负相关,土壤养分对林分乔木生物量的直接影响不显著,而是通过林分结构(林分密度、结构多样性)间接作用于林分乔木生物量。因此,在森林经营管理实践中,调整和优化森林的树种组成,增加森林结构的异质性对提高森林碳汇功能具有重要的意义。
Influencing factors of tree biomass in Quercus mongolica forest in the mountainous region of eastern Liaoning Province
-
摘要:
目的 探讨物种多样性、结构多样性以及土壤养分对林分乔木生物量的影响及其驱动机制,为森林的科学经营和碳汇功能提升提供科学依据。 方法 以辽东山区蒙古栎Quercus mongolica林为研究对象,利用线性回归模型分析了物种多样性、结构多样性以及土壤养分对林分乔木生物量的影响,在此基础上选择物种多样性和结构多样性的最优指标,结合土壤养分因子构建结构方程模型,定量分析物种多样性、结构多样性和土壤养分对林分乔木生物量的直接和间接影响。 结果 ①物种多样性、结构多样性均对林分乔木生物量有显著影响。其中,物种多样性中的丰富度(相关系数r=−0.503,P<0.01)和香农指数(r=−0.417,P<0.01)与乔木生物量呈极显著负相关,结构多样性中的胸径香农指数与乔木生物量呈极显著正相关(r=0.405,P<0.01)。②通过最优结构方程模型结果可知:物种多样性、结构多样性对林分乔木生物量均有显著的直接效应,路径系数分别为−0.394 (P<0.01)和0.280 (P<0.05);土壤养分对乔木生物量的直接影响并不显著,主要通过结构多样性和林分密度间接作用于乔木生物量,路径系数分别为−0.470 (P<0.01)和−0.655 (P<0.01)。 结论 物种多样性和结构多样性直接影响辽东山区蒙古栎林乔木生物量,而土壤养分则通过林分结构(结构多样性和林分密度)间接影响其乔木生物量。图4表2参46 Abstract:Objective This study aimed to elucidate the impacts of species diversity, structural diversity, and soil nutrients on tree biomass in forest stands, thereby providing a scientific basis for sustainable forest management and enhanced carbon sequestration capacity. Method The study focuses on Quercus mongolica forest in the mountainous region of eastern Liaoning Province. A Linear Regression Model was employed to analyze the effects of species diversity, structural diversity, and soil nutrients on tree biomass of forest stands. Based on this analysis, the most representative measures of species and structural diversity were identified. Subsequently, a structural equation model was constructed to quantitatively evaluate the direct and indirect effects on tree biomass. Result (1) Both species diversity and structural diversity significantly influenced tree biomass of forest stands. Species richness (r=−0.503, P<0.01) and Shannon index (r=−0.417, P<0.01) within species diversity were significantly negatively correlated with tree biomass of forest stands, while the Shannon index of diameter at breast height within structural diversity was significantly positively correlated with tree biomass of forest stands (r=0.405, P<0.01). (2) The optimal structural equation model indicated significant direct effects of species diversity and structural diversity on tree biomass of forest stands, with path coefficients of −0.394 (P<0.01) and 0.280 (P<0.05) respectively. Although soil nutrients did not have a significant direct impact on tree biomass, they influenced it indirectly through their effects on structural diversity and stand density, with path coefficients of −0.470 (P<0.01) and −0.655 (P<0.01) respectively. Conclusion These findings emphasize the primary role of species and structural diversity as direct drivers of tree biomass in Q. mongolica forest, while soil nutrients indirectly contribute to tree biomass by shaping forest stand structure. [Ch, 4 fig. 2 tab. 46 ref.] -
Key words:
- tree biomass /
- species diversity /
- structural diversity /
- soil nutrition /
- Querus mongolica forest
-
图 2 辽东山区蒙古栎林优势树种和不同龄组乔木层生物量分布特征
Figure 2 Biomass distribution characteristics of dominant tree species and different age-class trees in Q. mongolica forest in the mountainous region of eastern Liaoning Province
图 3 物种多样性和结构多样性与林分乔木生物量的线性回归分析
Figure 3 Linear regression analysis of species diversity, structural diversity, and stand tree biomass
图 4 生物和非生物因素对林分乔木生物量影响的结构方程模型图
Figure 4 Structural equation model diagram of the impact of biotic and abiotic factors
表 1 样地基本信息
Table 1. Basic information of the sample plots
龄组 胸径/cm 树高/m 林分密度/(株·hm−2) 林分断面积/(m2·hm−2) 幼龄林 11.5~13.5 9.7~11.8 1 256~2 289 20.10~27.06 中龄林 12.9~15.9 9.8~11.0 1 089~1 578 22.39~28.47 近熟林 18.9~24.6 11.1~15.0 688~1 040 20.15~33.03 成熟林 16.2~19.2 10.7~14.7 1 000~1 222 29.23~34.79 过熟林 18.3~20.7 11.4~15.9 733~956 31.69~44.69  下载: 导出CSV
下载: 导出CSV
表 2 最优结构方程模型中各解释变量对林分乔木生物量的标准化效益
Table 2. Standardized effects of each explanatory variable in the optimal structural equation model on the biomass of forest stand trees
项目 效应值 结构多样性 物种多样性 土壤养分 林分密度 直接效应 0.281 −0.394 −0.025 0 间接效应 0 0 0.244 −0.393 总效应 0.281 −0.394 0.219 −0.393
下载: 导出CSV
-
[1] 胡宇美, 马理辉, 李蕊, 等. 黄土高原地区森林生态系统地下生物量影响因素[J]. 生态学报, 2021, 41(21): 8643−8653. HU Yumei, MA Lihui, LI Rui, et al. Factor analysis of underground biomass in forest ecosystem on the Loess Plateau [J]. Acta Ecologica Sinica, 2021, 41(21): 8643−8653. [2] 张志, 田昕, 陈尔学, 等. 森林地上生物量估测方法研究综述[J]. 北京林业大学学报, 2011, 33(5): 144−150. ZHANG Zhi, TIAN Xin, CHEN Erxue, et al. Review of methods on estimating forest above ground biomass [J]. Journal of Beijing Forestry University, 2011, 33(5): 144−150. [3] 吴初平, 韩文娟, 江波, 等. 浙江定海次生林内物种丰富度与生物量和生产力关系的环境依赖性[J]. 生物多样性, 2018, 26(6): 545−553. WU Chuping, HAN Wenjuan, JIANG Bo, et al. Relationships between species richness and biomass/productivity depend on environmental factors in secondary forests of Dinghai, Zhejiang Province [J]. Biodiversity Science, 2018, 26(6): 545−553. [4] CAVANAUGH K C, GOSNELL J S, DAVIS S L, et al. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale [J]. Global Ecology and Biogeography, 2014, 23(5): 563−573. [5] VELLEND M, BAETEN L, BECKER-SCARPITTA A, et al. Plant biodiversity change across scales during the anthropocene [J]. Annual Review of Plant Biology, 2017, 68: 563−586. [6] POORTER L, van der SANDE M T, THOMPSON J, et al. Diversity enhances carbon storage in tropical forests [J]. Global Ecology and Biogeography, 2015, 24(11): 1314−1328. [7] 范秀华, 张宝权, 范春雨. 长白山典型天然林不同演替阶段物种多样性和结构多样性对生产力影响[J]. 北京林业大学学报, 2021, 43(12): 1−8. FAN Xiuhua, ZHANG Baoquan, FAN Chunyu. Effects of species diversity and structural diversity on productivity in different succession stages of typical natural forest in Changbai Mountains of northeastern China [J]. Journal of Beijing Forestry University, 2021, 43(12): 1−8. [8] 张萌, 范秀华, 岳庆敏, 等. 吉林蛟河针阔混交林生物与非生物因素对生产力的影响[J]. 林业科学, 2023, 59(12): 71−77. ZHANG Meng, FAN Xiuhua, YUE Qingmin, et al. Effects of biotic and abiotic factors on productivity of coniferous and broad-leaved mixed forest in Jiaohe, Jilin Province [J]. Scientia Silvae Sinicae, 2023, 59(12): 71−77. [9] PRADO-JUNIOR J A, SCHIAVINI I, VALE V S, et al. Conservative species drive biomass productivity in tropical dry forests [J]. Journal of Ecology, 2016, 104(3): 817−827. [10] LEI Xiangdong, WANG Weifeng, PENG Changhui. Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick, Canada [J]. Canadian Journal of Forest Research, 2009, 39(10): 1835−1847. [11] 朱杰, 吴安驰, 邹顺, 等. 南亚热带常绿阔叶林树木多样性与生物量和生产力的关联及其影响因素[J]. 生物多样性, 2021, 29(11): 1435−1446. ZHU Jie, WU Anchi, ZOU Shun, et al. Relationships between tree diversity and biomass/productivity and their influence factors in a lower subtropical evergreen broad-leaved forest [J]. Biodiversity Science, 2021, 29(11): 1435−1446. [12] 庞荣荣, 彭潔莹, 闫琰. 太白山次生锐齿栎林地上生物量影响因素[J]. 林业科学, 2021, 57(10): 157−165. PANG Rongrong, PENG Jieying, YAN Yan. Factors influencing aboveground biomass in the secondary forest of Quercus aliena var. acutiserrata in Taibai mountain [J]. Scientia Silvae Sinicae, 2021, 57(10): 157−165. [13] 鲁君悦, 吴兆飞, 张春雨, 等. 吉林蛟河针阔混交林林层结构对生产力的影响[J]. 生态学报, 2021, 41(5): 2024−2032. LU Junyue, WU Zhaofei, ZHANG Chunyu, et al. Influence of forest strate structure on productivity of coniferous and broad-leaved mixed forest in Jiaohe, Jilin [J]. Acta Ecologica Sinica, 2021, 41(5): 2024−2032. [14] YACHI S, LOREAU M. Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities [J]. Ecology Letters, 2007, 10(1): 54−62. [15] ALI A, YAN Enrong, CHEN H Y H, et al. Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in eastern China [J]. Biogeosciences, 2016, 13(16): 4627−4635. [16] DĂNESCU A, ALBRECHT A T, BAUHUS J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany [J]. Oecologia, 2016, 182(2): 319−333. [17] EMMETT DUFFY J, GODWIN C M, CARDINALE B J. Biodiversity effects in the wild are common and as strong as key drivers of productivity [J]. Nature, 2017, 549(7671): 261−264. [18] 魏安然, 张雨秋, 谭凌照, 等. 抚育采伐对针阔混交林林分结构及物种多样性的影响[J]. 北京林业大学学报, 2019, 41(5): 148−158. WEI Anran, ZHANG Yuqiu, TAN Lingzhao, et al. Effects of tending felling on stand structure and species diversity of mixed coniferous and broadleaved forest [J]. Journal of Beijing Forestry University, 2019, 41(5): 148−158. [19] 魏安然. 采伐干扰对吉林蛟河针阔混交林结构与生长的影响[D]. 北京: 北京林业大学, 2020. WEI Anran. Effect of Cutting Disturbance on Structure and Growth of Coniferous and Broad-leaved Mixed Forest in Jiaohe, Jilin Province[D]. Beijing: Beijing Forestry University, 2020. [20] HOFHANSL F, CHACÓN-MADRIGAL E, FUCHSLUEGER L, et al. Climatic and edaphic controls over tropical forest diversity and vegetation carbon storage[J/OL]. Scientific Reports, 2020, 10(1): 5066[2024-08-01]. DOI: 10.1038/s41598-020-61868-5. [21] 何潇, 李海奎, 曹磊, 等. 退化森林生态系统中林分碳储量的驱动因素——以内蒙古大兴安岭为例[J]. 林业科学研究, 2020, 33(2): 69−76. HE Xiao, LI Haikui, CAO Lei, et al. The factors affecting carbon storage in degraded forest ecosystem: a case study from Daxing’anling areas of Inner Mongolia [J]. Forest Research, 2020, 33(2): 69−76. [22] 贾勃, 王新杰. 东北针阔混交林生物量动态过程及稳定性研究[J]. 林业科学研究, 2023, 36(5): 41−49. JIA Bo, WANG Xinjie. Dynamics and stability of biomass of coniferous and broad-leaved mixed forests in northeast China [J]. Forest Research, 2023, 36(5): 41−49. [23] 谭珊珊, 王忍忍, 龚筱羚, 等. 群落物种及结构多样性对森林地上生物量的影响及其尺度效应: 以巴拿马BCI样地为例[J]. 生物多样性, 2017, 25(10): 1054−1064. TAN Shanshan, WANG Renren, GONG Xiaoling, et al. Scale dependent effects of species diversity and structural diversity on aboveground biomass in a tropical forest on Barro Colorado Island, Panama [J]. Biodiversity Science, 2017, 25(10): 1054−1064. [24] 陈科屹, 张会儒, 雷相东. 天然次生林蒙古栎种群空间格局[J]. 生态学报, 2018, 38(10): 3462−3470. CHEN Keyi, ZHANG Huiru, LEI Xiangdong. Spatial pattern of Quercus mongolica in natural secondary forest [J]. Acta Ecologica Sinica, 2018, 38(10): 3462−3470. [25] 董莉莉, 汪成成, 赵济川, 等. 辽东山区蒙古栎次生林天然更新特征及其影响因子分析[J]. 西北农林科技大学学报(自然科学版), 2023, 51(1): 94−101,109. DONG Lili, WANG Chengcheng, ZHAO Jichuan, et al. Natural regeneration characteristics and influencing factors of Quercus mongolica secondary forest in the mountainous area of eastern Liaoning [J]. Journal of Northwest A&F University (Natural Science Edition), 2023, 51(1): 94−101,109. [26] 罗云建, 王效科, 逯非. 中国主要林木生物量模型手册[M]. 北京: 中国林业出版社, 2015. LUO Yunjian, WANG Xiaoke, LU Fei. Comprehensive Database of Biomass Regressions for China’s Tree Species[M]. Beijing: China Forestry Publishing House, 2015. [27] GRACE J B, ANDERSON T M, SEABLOOM E W, et al. Integrative modelling reveals mechanisms linking productivity and plant species richness [J]. Nature, 2016, 529: 390−393. [28] 吴兆飞, 张雨秋, 张忠辉, 等. 东北温带森林林分结构与生产力关系研究[J]. 北京林业大学学报, 2019, 41(5): 48−55. WU Zhaofei, ZHANG Yuqiu, ZHANG Zhonghui, et al. Study on the relationship between forest structure and productivity of temperate forests in northeast China [J]. Journal of Beijing Forestry University, 2019, 41(5): 48−55. [29] BOSELA M, IGOR Š, PETRÁŠ R, et al. The effects of climate warming on the growth of European beech forests depend critically on thinning strategy and site productivity [J]. Agricultural and Forest Meteorology, 2016, 222: 21−31. [30] 谭凌照, 范春雨, 范秀华. 吉林蛟河阔叶红松林木本植物物种多样性及群落结构与生产力的关系[J]. 植物生态学报, 2017, 41(11): 1149−1156. TAN Lingzhao, FAN Chunyu, FAN Xiuhua. Relationships between species diversity or community structure and productivity of woody-plants in a broad-leaved Korean pine forest in Jiaohe, Jilin, China [J]. Chinese Journal of Plant Ecology, 2017, 41(11): 1149−1156. [31] WRIGHT I J, REICH P B, ATKIN O K, et al. Irradiance, temperature and rainfall influence leaf dark respiration in woody plants: evidence from comparisons across 20 sites [J]. New Phytologist, 2006, 169(2): 309−319. [32] 张雪儿, 乔雪涛, 张春雨, 等. 吉林蛟河针阔混交林生物量时间稳定性驱动机制[J]. 生态学杂志, 2023, 42(4): 812−819. ZHANG Xueer, QIAO Xuetao, ZHANG Chunyu, et al. The driving mechanism of biomass temporal stability of mixed coniferous and broad-leaved forest in Jiaohe, Jilin [J]. Chinese Journal of Ecology, 2023, 42(4): 812−819. [33] ZHANG Yu, CHEN H Y H. Individual size inequality links forest diversity and above-ground biomass [J]. Journal of Ecology, 2015, 103(5): 1245−1252. [34] CASPERSEN J P, PACALA S W. Successional diversity and forest ecosystem function [J]. Ecological Research, 2001, 16(5): 895−903. [35] OUYANG Shuai, XIANG Wenhua, WANG Xiangping, et al. Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south China [J]. Forest Ecology and Management, 2016, 372: 291−302. [36] LIANG Jingjing, CROWTHER T W, PICARD N, et al. Positive biodiversity-productivity relationship predominant in global forests[J/OL]. Science, 2016, 354(6309): aaf8957[2024-08-01]. DOI: 10.1126/science.aaf8957. [37] RATCLIFFE S, WIRTH C, JUCKER T, et al. Biodiversity and ecosystem functioning relations in European forests depend on environmental context [J]. Ecology Letters, 2017, 20(11): 1414−1426. [38] WU Xian, WANG Xiangping, TANG Zhiyao, et al. The relationship between species richness and biomass changes from boreal to subtropical forests in China [J]. Ecography, 2015, 38(6): 602−613. [39] ALI A, YAN Enrong. The forest strata-dependent relationship between biodiversity and aboveground biomass within a subtropical forest [J]. Forest Ecology and Management, 2017, 401: 125−134. [40] QUESADA C A, PHILLIPS O L, SCHWARZ M, et al. Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate [J]. Biogeosciences, 2012, 9(6): 2203−2246. [41] 刘晨, 张春雨, 赵秀海. 采伐干扰对吉林蛟河针阔混交林生产力稳定性的影响[J]. 林业科学, 2022, 58(3): 1−9. LIU Chen, ZHANG Chunyu, ZHAO Xiuhai. Effects of disturtance by thinning on productivity stability of conifer-broadleaf mixed forest in Jiaohe, Jilin Province [J]. Scientia Silvae Sinicae, 2022, 58(3): 1−9. [42] 金超, 李领寰, 吴初平, 等. 浙江省公益林生物多样性和立地对生物量的影响[J]. 浙江农林大学学报, 2021, 38(6): 1083−1090. JIN Chao, LI Linghuan, WU Chuping, et al. Impact of biodiversity and site factors on biomass of public welfare forests in Zhejiang Province [J]. Journal of Zhejiang A&F University, 2021, 38(6): 1083−1090. [43] 王冰怡, 张勇, 吴翠蓉, 等. 不同造林年限马尾松林碳密度结构特征及其影响因素[J]. 浙江农林大学学报, 2025, 42(2): 291−301. WANG Bingyi, ZHANG Yong, WU Cuirong, et al. Characteristics and influencing factors of carbon density structure in Pinus massoniana forests with different afforestation years[J]. Journal of Zhejiang A&F University, 2025, 42(2): 291−301. [44] FERRY B, MORNEAU F, BONTEMPS J D, et al. Higher treefall rates on slopes and waterlogged soils result in lower stand biomass and productivity in a tropical rain forest [J]. Journal of Ecology, 2010, 98(1): 106−116. [45] 田立新, 丁易, 臧润国. 热带次生林树种多样性和地上生物量恢复动态的研究进展[J]. 陆地生态系统与保护学报, 2022, 2(6): 88−96. TIAN Lixin, DING Yi, ZANG Runguo. Advances in tree species diversity and aboveground biomass recovery dynamics in tropical secondary forests [J]. Terrestrial Ecosystem and Conservation, 2022, 2(6): 88−96. [46] PRETZSCH H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures [J]. Forest Ecology and Management, 2014, 327: 251−264. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240507
 点击查看大图
点击查看大图

计量
- 文章访问数: 1102
- HTML全文浏览量: 287
- PDF下载量: 29
- 被引次数: 0
