




-
森林是地球上最庞大最重要的陆地生态系统,物种多样性及其保护问题一直是生态学领域的研究热点[1]。虽然草本层生物量仅占整个森林生物量的约1%,但其包含的物种数量却高达林内总量的90%以上[2],且其多样性在保持水土平衡、促进养分循环和维持生物多样性等方面具有重要意义[3]。然而,确定林下植物群落物种多样性的关键驱动因素是森林生态学研究领域的一大挑战[4]。尽管已有研究表明[5−8]:林分结构和土壤养分均与林下草本层植物的多样性密切相关,然而考虑到众多的影响因子以及森林作为一个复杂生态系统的特点,目前仍未能明确揭示出主导天然次生白桦Betula platyphylla林下草本植物多样性的关键因素,因此,这一领域的科学研究仍需进一步深入展开。
近年来,许多学者已经开展了林下草本植物的多样性研究。闫玮明等[9]发现:粤北阔叶人工林和次生林植物多样性主要受土壤氮、磷影响;王景玺等[10]研究了辽宁章古台樟子松Pinus sylvestris var. Mongolica人工林死亡率、树高、胸径、冠幅和密度对林下草本多样性的影响;邓婷婷等[11]探究了不同地形因子和林分结构对暖温带落叶阔叶林草本植物物种多样性的影响;邹星晨等[12]采用冗余分析研究了生物因子和非生物因子对青海云杉Picea crassifolia不同演替阶段林下草本多样性的影响。然而,以往研究多侧重于郁闭度、林分密度、林龄等林分非空间结构因子和海拔、坡度等环境因子方面,难以全面揭示对草本植物多样性影响的真实机理。林分空间结构是林分结构重要的组成部分,不仅决定林分的内部竞争态势以及各物种空间生态位,而且在森林结构中相对易于调控[13],在分析和调整林分结构与其功能之间的关系上起到至关重要的作用[14]。
地处黄土高原和青藏高原交界地带的青海省大通县宝库林场,生态环境相当敏感,森林资源迫切需要针对性的管理。白桦是该地区现存天然次生林中的先锋树种,但由于树种单一,导致林下草本多样性低,森林生态系统脆弱、生态功能低下,不利于持续发挥其生态效益[15]。基于此,本研究以天然次生白桦林为研究对象,调查和分析其林分的空间结构、非空间结构、土壤养分状况以及草本植物多样性等指标,运用冗余分析方法试图揭示影响天然次生白桦林下草本植物多样性的主要驱动因素,以期为该区域次生林空间结构调控、草本多样性改善以及生态效益最大化提供科学依据。
-
研究区位于青海省大通县宝库林场, 36°55′~37°32′N, 100°52′~101°56′E,海拔为2 660~4 600 m,地势西北高东南低。该区属高原大陆性气候,在垂直方向上变化显著,冷暖干湿随海拔高度的不同而有很大差异。年平均气温为4.9 ℃,≥5 ℃的年积温为1 908 ℃,无霜期为60~130 d,年平均降水量为549.9 mm。该地区的土壤以黄土母质上形成的山地棕褐土和栗钙土为主,土层厚实。森林平均覆盖率为79.0%。研究区主要针叶树种为青海云杉Picea crassifolia、祁连圆柏Juniperus przewalskii、油松Pinus tabuliformis等;落叶阔叶树种有白桦、山杨Populus davidiana、青杨Populus cathayana等;灌木树种有峨眉蔷薇Rosa omeiensis、锦鸡儿Caragana sinica、金露梅Potentilla fruticosa等。白桦林主要分布在宝库河、东峡河和黑林河中上游的大部分区域,具有复杂的立地结构,树种相对单一,但亦与青海云杉、华北落叶松Larix gmelinii var. principis-rupprechtii、青杨等树种形成混交林。
-
2023年6—7月,在宝库林场现存天然次生白桦林中选取林龄约80 a,坡向为阴坡,立地条件基本一致的20个24 m×24 m标准地,设置宽为2 m的缓冲区以消除边缘效应。详细记录各样地的坡位、坡向、海拔和植被类型等信息,并对胸径>5 cm的林木进行逐一检尺,包括树高、胸径、冠幅、枝下高以及郁闭度等关键林分指标。同时,在各样地四角和中心位置,设置5个1 m×1 m的草本小样方,并调查和记录草本植物的种类、数量、高度以及覆盖度(表1)。
表 1 样地基本信息
Table 1. Basic information table of sample plots
样地编号 海拔/m 纬度(N) 经度(E) 林分密度/(株·hm−2) 郁闭度 平均胸径/cm 平均树高/m 1 2783 37°13′50″ 101°56′17″ 295 0.59 30.52 12.92 2 2936 37°14′47″ 101°62′43″ 312 0.59 14.46 6.54 3 2888 37°12′73″ 101°59′93″ 329 0.67 17.68 8.90 4 2881 37°11′86″ 101°51′20″ 381 0.62 29.50 14.88 5 2894 37°11′83″ 101°51′35″ 399 0.55 32.93 16.91 6 2884 37°14′74″ 101°62′69″ 399 0.61 9.30 9.87 7 2788 37°13′53″ 101°56′05″ 416 0.65 23.94 15.90 8 2847 37°11′90″ 101°51′37″ 434 0.61 28.59 15.93 9 2803 37°12′03″ 101°50′95″ 434 0.6 23.72 12.44 10 2801 37°13′47″ 101°56′17″ 468 0.64 23.36 13.88 11 2855 37°11′90″ 101°51′08″ 503 0.68 28.75 14.27 12 2940 37°14′46″ 101°62′39″ 538 0.69 9.64 6.46 13 2850 37°11′92″ 101°51′20″ 572 0.57 28.22 14.64 14 2940 37°14′69″ 101°62′75″ 572 0.64 17.27 15.92 15 2773 37°13′51″ 101°56′24″ 736 0.63 21.11 13.48 16 2793 37°13′02″ 101°59′50″ 850 0.74 9.53 8.41 17 2946 37°14′47″ 101°62′41″ 850 0.76 8.16 6.01 18 2965 37°14′78″ 101°62′67″ 896 0.78 9.83 13.18 19 2853 37°12′85″ 101°59′60″ 873 0.68 11.21 15.29 20 2868 37°12′78″ 101°59′72″ 638 0.69 10.62 11.19 -
为全面评估林分的结构特征,选取了角尺度、大小比数、Hegyi竞争指数和开敞度等指标来表征林分的空间结构;林分非空间结构则通过测量林分密度、郁闭度、平均树高以及平均胸径等参数来全面描述。应用Voronoi图确定样地林木空间结构单元,参考惠刚盈等[16]计算空间结构参数的方法,计算不同林木的角尺度、大小比数、Hegyi竞争指数、开敞度等林分空间结构参数。通过统计单位面积林地上林木株数计算得出林分密度;使用鱼眼相机(WinSCANOPYTM 2010a)获取冠层照片,运用XLScanopy软件处理照片,计算得出郁闭度;分别使用胸径尺和测高仪测定[16] 林分平均胸径和平均树高。
-
在各个标准地中,用土钻分别在5个草本小样方的0~20 cm深度范围内进行土壤采样。为确保土样的代表性,将同层土壤样本混匀为1个综合土样,带回实验室在自然条件下风干,筛除颗粒较大的砂粒和根系后进行土壤样品测定。碱解氮采用靛酚蓝比色法测定;速效钾采用火焰光度法测定;有机质采用重铬酸钾容量法-消煮法测定;全氮采用凯氏定氮法测定;全磷和速效磷通过钼锑抗显色紫外分光光度法测定[17−18]。
-
本研究中,采用物种重要值作为衡量物种重要性的标准,并利用物种丰富度、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数对草本植物的多样性水平进行全面评估[19]。
-
利用Excel 2019和SPSS 22.0进行数据整理和统计分析,采用Pearson相关系数分析林下草本多样性与影响因子的相关性。应用CANOCO 5.0中的冗余分析(RDA)排序,探讨影响草本多样性变化的主要因素。采用Origin 2021绘图。
-
由表2可知:研究区白桦林下草本植物共47种,分属24科41属。伞形科在白桦林下的草本层中属于优势科, 6属7种,占总种数的14.89%;其次为毛茛科含5属5种,占总种数的10.64%;菊科含3属4种,占总种数的8.51%;茜草科含2属4种,占总种数的8.51%;蔷薇科含3属3种,占总种数的6.38%;蓼科含2属3种,占总种数的6.38%;紫草科含2属2种、禾本科含2属2种、石竹科含1属2种,分别占总种数的4.26%;其余16科为单属单种。其中,白桦林下草本植物以黑麦草Lolium perenne、野草莓Fragaria vesca、珠芽蓼Polygonum viviparum、四叶葎Galium bungei、老鹳草Geranium wilfordii为优势种,其重要值均值分别为19.52%、18.91%、10.53%、6.95%、5.27%。
表 2 林下草本植物科内种的组成
Table 2. Composition of species within the understory herb family
种数 科数 科名(属数/种数) ≥7 1 伞形科Apiaceae (6/7) 4~5 3 茜草科Rubiaceae (2/4)、菊科Asteraceae (3/4)、毛茛科Ranunculaceae (5/5) 2~3 5 石竹科Caryophyllaceae (1/2)、紫草科Boraginaceae (2/2)、禾本科Gramineae (2/2)、蓼科(2/3) Polygonaceae、蔷薇科Rosaceae (3/3) 1 16 牻牛儿苗科Geraniaceae (1/1)、瓶尔小草科Ophioglossaceae (1/1)、豆科Fabaceae (1/1)、木贼科Equisetaceae (1/1)、五福花科Adoxaceae (1/1)、唇形科Lamiaceae (1/1)、堇菜科Violaceae (1/1)、芍药科Paeoniaceae (1/1)、玄参科Scrophulariaceae (1/1)、天门冬科Asparagaceae (1/1)、真藓科Bryaceae (1/1)、报春花科Primulaceae (1/1)、十字花科Brassicaceae (1/1)、桔梗科Campanulaceae (1/1)、莎草科Cyperaceae (1/1) -
由表3可知:研究区林分大小比数均值为0.50,说明林分整体呈中庸状态;林分角尺度均值为0.48,属于理想的随机分布状态;林分竞争指数均值为3.25,说明林木竞争程度较弱;林分开敞度均值为0.37,表明林内透光基本充足。其中,物种丰富度的变异系数为24.89%,Pielou均匀度指数的变异系数为15.17%,这2个指数的变异系数相对较低;Simpson多样性指数和Shannon-Wiener多样性指数的变异系数则相对较高,分别为39.54%、33.48%。数据显示各样地林下草本植物水平的差异性,同时进一步表明各样地乔木在林分结构和土壤特性上存在明显不同。
表 3 样地各变量基本特征
Table 3. Basic characteristics of the variables in the sample plots
项目 非空间结构 空间结构 土壤养分 林分密度
(株·hm−2)郁闭度 平均胸
径/cm平均树
高/m大小比 角尺度 竞争
指数开敞度 pH 有机质/
(g·kg−1)均值 516.15 0.57 16.97 12.50 0.50 0.48 3.25 0.37 7.65 149.78 标准差 250.54 0.07 5.98 2.80 0.07 0.04 0.83 0.12 0.21 13.86 变异系数/% 43.99 12.93 35.22 22.36 13.29 8.26 25.51 31.75 3.19 9.26 项目 土壤养分 草本多样性 速效钾/
(mg·kg−1)速效磷/
(mg·kg−1)碱解氮/
(mg·kg−1)全磷/
(mg·kg−1)全氮/
(mg·kg−1)物种丰
富度Simpson多样性
指数(D)Shannon-Wiener
多样性指数(H)Pielou均匀度
指数(J)均值 246.56 12.04 0.80 0.68 5.83 12.60 0.96 2.01 0.77 标准差 76.74 9.12 0.09 0.11 1.11 3.14 0.38 0.67 0.12 变异系数/% 31.13 75.74 11.69 15.67 18.98 24.89 39.54 33.48 15.17 -
如图1所示:物种丰富度与平均树高、平均胸径呈显著负相关(P<0.05),与开敞度、速效钾、全磷、速效磷、碱解氮和全氮存在显著正相关(P<0.05);Simpson多样性指数与林分密度、郁闭度、平均树高、大小比数、竞争指数呈显著负相关(P<0.05),与开敞度、有机质、速效钾、碱解氮存在显著正相关(P<0.05);Shannon-Wiener多样性指数与郁闭度、平均胸径、平均树高、大小比数呈显著负相关(P<0.05),与开敞度、速效钾、碱解氮、有机质、速效磷呈显著相关(P<0.05);Pielou均匀度指数与林分密度、大小比数、竞争指数呈显著正相关(P<0.05),与开敞度、有机质质量分数存在显著负相关(P<0.05)。角尺度和土壤pH与4个草本多样性指标均无显著相关性。
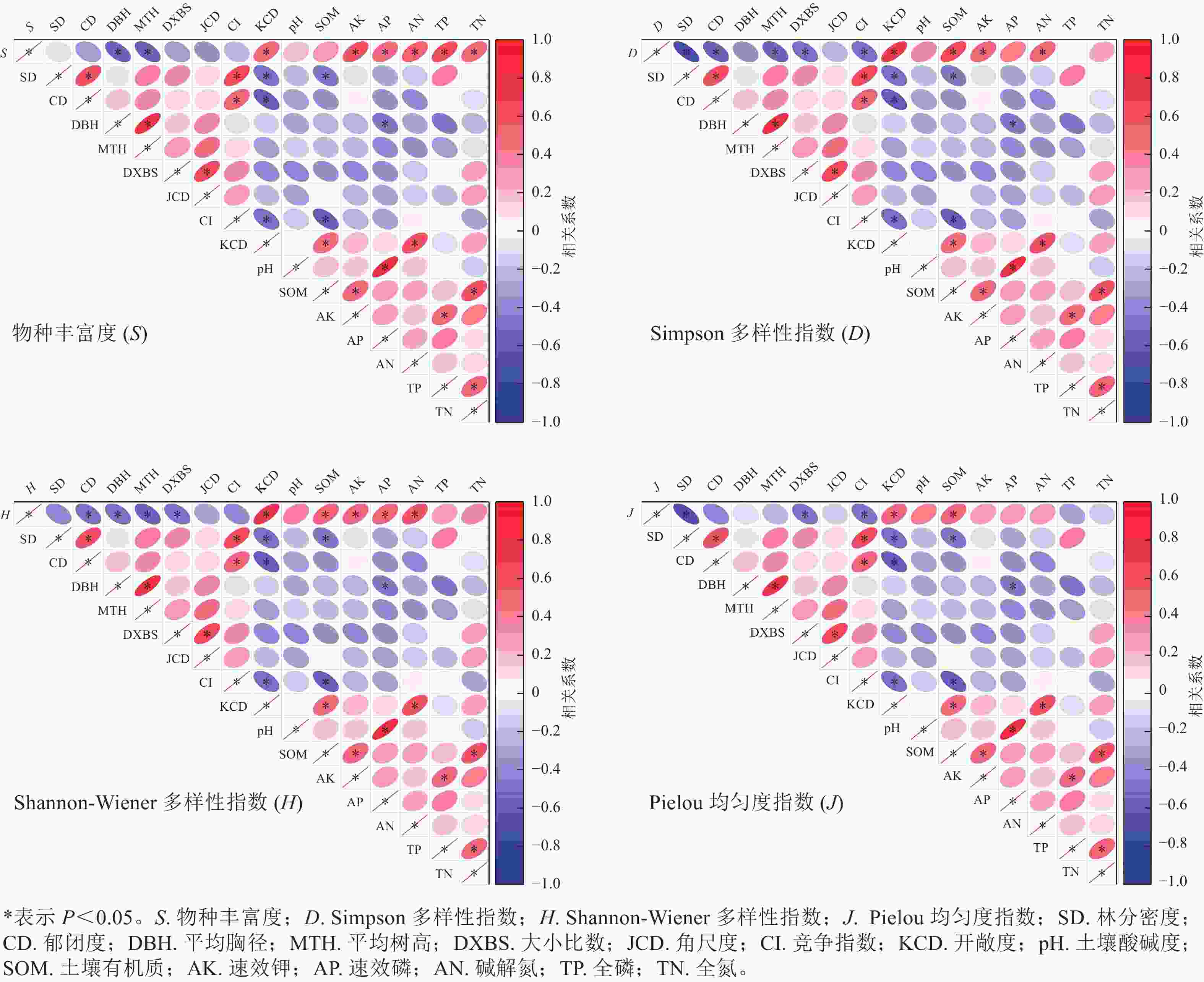
图 1 林分结构因子和土壤养分因子与多样性指数的相关性
Figure 1. Correlations between forest structure factors and soil nutrient factors and diversity index
-
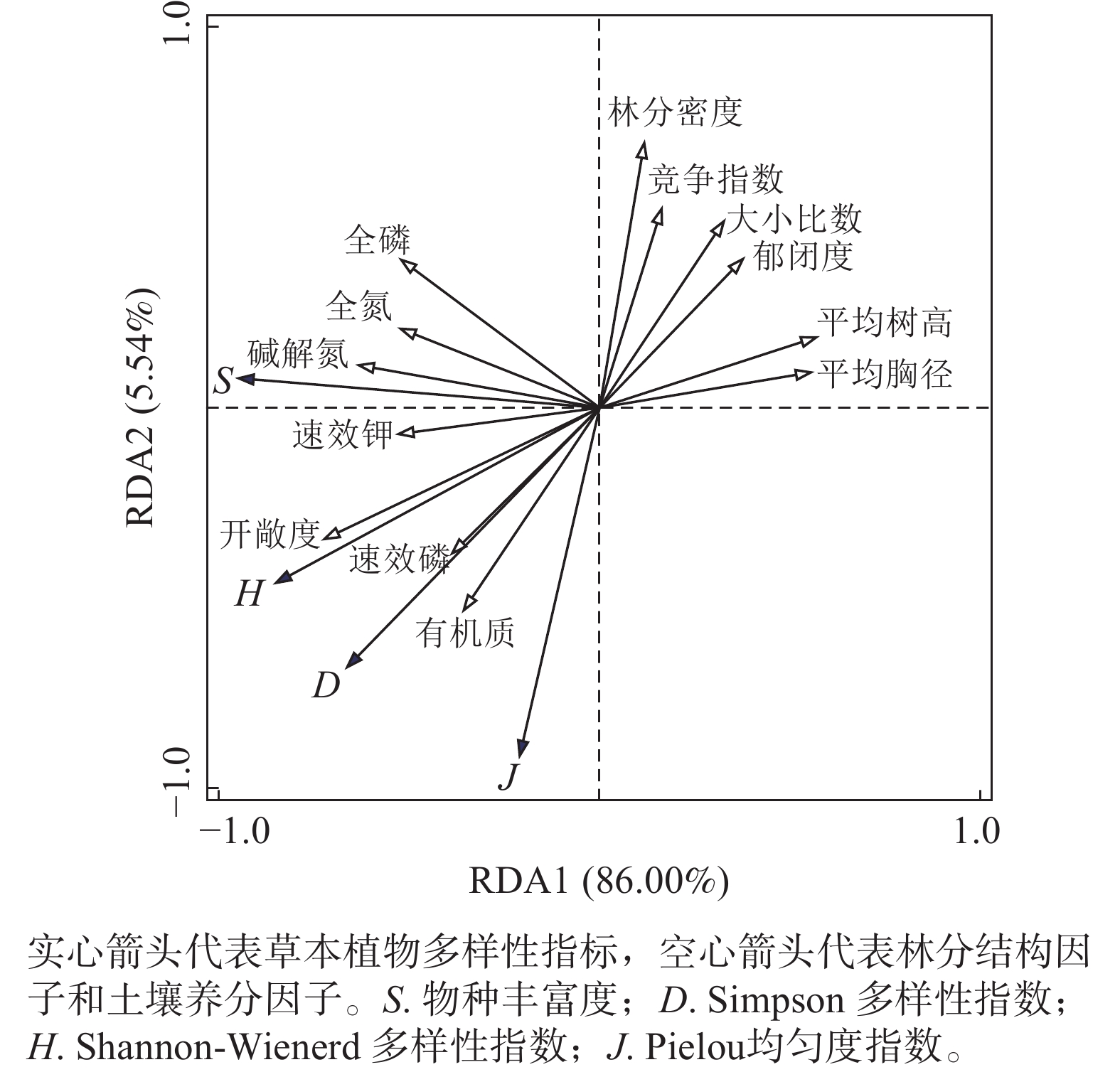
冗余分析排序结果(图2)显示:前2个排序轴累计包含了草本植物多样性指数与林分结构因子和土壤养分因子关系91.54%的解释率,其中第1主轴和第2主轴解释变量分别为86.00%和5.54%。实心箭头代表草本植物多样性指标,空心箭头代表林分结构因子和土壤养分因子。箭头长度反映的是各因子与多样性指数之间变化关系的强弱,箭头连线和排序轴夹角揭示某一因子与排序轴的相关性。夹角越小相关性越强,夹角小于90°表示正相关,夹角大于90°则表示负相关[20]。林分空间结构因子中开敞度与大小比数贡献度较大,贡献率分别为47.2%和9.6%;非空间结构因子中平均胸径贡献度最大,为17.4%;且开敞度与草本植物多样性变化呈极显著正相关(P=0.002,F=14.8);平均胸径和大小比数与草本植物多样性变化呈显著负相关(P=0.010,F=7.8;P=0.042,F=4.2)。土壤养分因子中碱解氮和速效钾与草本植物多样性指数夹角均为锐角且连线长度较长,较其他土壤养分因子贡献度较大,分别为10.1%和8.1%,均与草本植物多样性变化呈显著正相关(表4,P=0.036,F=5.4;P=0.05,F=4.8)。

图 2 林分结构因子、土壤因子与草本植物多样性RDA分析结果
Figure 2. RDA analysis results of stand structure factors, soil factors and herbaceous plant diversity
表 4 林分结构因子、 土壤养分因子解释度与蒙特卡洛检验
Table 4. Explanatory degree of stand structure factors and soil nutrient factors and Monte Carlo test
影响因素 解释度/% 贡献率/% F检验 P 影响因素 解释度/% 贡献率/% F检验 P 开敞度 44.1 47.2 14.8 0.002 有机质 2.1 2.3 0.7 0.462 平均胸径 16.8 17.4 7.8 0.010 全氮 1.5 1.6 0.5 0.496 碱解氮 7.9 10.1 5.4 0.036 全磷 0.6 0.7 0.6 0.590 大小比数 8.2 9.6 4.2 0.042 林分密度 0.5 0.6 0.4 0.596 速效钾 7.6 8.1 4.8 0.050 平均树高 0.3 0.3 0.2 0.726 郁闭度 1.1 1.2 1.1 0.322 竞争指数 0.1 0.1 0.1 0.988 速效磷 0.7 0.8 0.6 0.454 -
研究结果表明:白桦林下草本植物较少,以单科单种、单属单种为主,分布较不均匀,草本多样性存在差异,其中黑麦草、野草莓、珠芽蓼等植物是林下主要优势种,这可能与它们独特的生物学特性有关。具体表现为:黑麦草具有快速分蘖的能力、高度的逆境抗性以及较强的再生能力[21];野草莓具细长匍匐茎,当接触湿润土壤后,可在节点生根形成分株,其繁殖速度快且繁殖能力强[22];珠芽蓼是典型高山草甸植物,耐寒性强,对异质性生境具有较强的适应能力[23]。优势种大多为喜光植物,这可能是白桦自身的喜光习性及白桦林的林冠层较为开放,阳光可以穿透林冠层照射到地面,为其提供光照导致的[24]。
-
本研究表明:开敞度和大小比数是影响林下草本多样性变化的主要空间结构因子,这与刘红民等[25]和马刘乐等[26]研究结果相似。开敞度能直接决定林下的有效光强度。光照被认为是限制林下草本多样性的主要因素[27],在一定程度上影响了林下的耐荫草本植物、高光需求的非耐荫植物以及普通草本植物的存活率,进而影响林下植物多样性[28]。此外,在高开敞度林分中,草本植物之间竞争较小,有更多的生长空间扩展其根系和繁殖器官,从而优化林分竞争态势,提高林下草本植物多样性。大小比数反映树木的大小分化程度。本研究发现:大小比数与草本植物多样性之间存在显著负相关关系。在林木大小分化明显的林分中,大型乔木冠层盖度增加,且其根系也更加发达,可能会导致草本层在光照、养分和水分的获取上受到一定限制,进而影响草本层植物的生存、繁衍和物种组成[29]。相反,在林木分化较小的林分中,林下光照条件更为优越,有利于草本植物的生长和繁衍。此外,研究表明乔木的平均胸径与草本植物多样性之间存在显著负相关关系。这可能是因为乔木树高和胸径越大,其冠幅也越大,抑制了林下草本植物对光照的吸收能力,导致喜阳草本植物的数量逐渐减少[30]。另外,乔木对土壤养分的充分吸收加剧了林下草本植物生态位竞争[31],进而影响林下物种丰富度和多样性。
-
本研究表明:土壤因子中的碱解氮质量分数与林下物种丰富度及草本植物多样性指数呈显著正相关。这可能是碱解氮作为植物可以直接吸收和利用的土壤养分,在林地生态系统中发挥着重要作用。林地内的家畜粪便和动植物残体通过自然分解增加了土壤中的碱解氮。这些氮素被草本植物直接吸收利用,进而促进了其生长和发育。因此,在碱解氮质量分数较高的林地中,不仅草本植物的数量显著增加,还使得草本层各物种的数量分布也变得更加均匀,从而提高林下物种的丰富度和整体的草本多样性[32]。速效钾是土壤中易被植物吸收利用的钾素形态,主要经过土壤中的微生物和植物代谢作用转化而来,对植物的根系生长和根毛发育具有显著促进作用。这不仅扩大了植物的吸收面积,还增加了吸收深度,从而提高植物对其他养分的吸收能力[33]。本研究发现:速效钾对林下草本多样性有显著正向影响,高速效钾质量分数的土壤能够满足草本植物对钾元素的需求,促进植物体内酶的活化,增强光合作用,从而提高草本植物的生长速度和生长量[34]。此外,速效钾可提高植物抗逆性,使植物能够更好地适应不利环境条件。因此,速效钾质量分数高的样地中草本植物多样性显著提高[35]。王树梅等[36]认为:林下植被多样性受磷元素显著影响,但本研究显示:速效磷质量分数对草本物种多样性的影响相对较小。一方面可能是由于研究区土壤磷元素质量分数低[13],未能展现出明显差异性;另一方面可能是研究区的大部分优势种对磷的需求量较小,导致整体上对草本多样性的影响有限[37]。
-
林分空间结构、林分非空间结构和土壤养分对林下草本多样性均有显著影响,林分结构因子中开敞度、平均胸径、大小比数解释度较高,是影响草本植物多样性的主导林分因子;土壤养分因子中碱解氮和速效钾解释度较高,是主要的土壤养分特征。因此,对林下草本植物多样性低下的天然次生白桦林可适当调整其林分开敞度和大小比数,科学优化林分空间结构,提升土壤肥力,以维持和提高林下草本多样性整体水平,促进生态系统的稳定和可持续发展。
Impact of stand structure and soil nutrients on herbaceous diversity in natural secondary Betula platyphylla forests in eastern Qinghai Province
-
摘要:
目的 探究林分结构和土壤养分对天然次生白桦Betula platyphylla林下草本多样性影响的主导因子,旨在为青海天然次生白桦林林分结构优化与提高草本多样性提供理论依据。 方法 以青海省大通县宝库林场天然次生白桦林为研究对象,通过典型取样法在该研究区内选取20个24 m×24 m的标准样地,利用相关性分析和冗余分析法探究多重因素(林分空间结构、林分非空间结构、土壤养分)对草本多样性的影响。 结果 ①白桦林下共发现草本植物24科41属47种,其中黑麦草Lolium perenne、野草莓Fragaria vesca、珠芽蓼Polygonum viviparum、四叶葎Galium bungei、老鹳草Geranium wilfordii为优势种,其重要值均值分别为19.52%、18.91%、10.53%、6.95%、5.27%。②冗余分析结果显示:开敞度、平均胸径、大小比数分别可解释44.1%、16.8%、8.2%的草本植物多样性变化格局,是影响草本多样性的主要林分因子;碱解氮和速效钾分别可解释7.9%和7.6%的草本植物多样性变化格局,是主要的土壤养分因子。 结论 林分结构和土壤养分均显著影响林下草本多样性。可通过调整林分空间结构、优化林分竞争态势和提升土壤肥力等方式提高林下草本多样性整体水平。图2表4参37 Abstract:Objective This study is to explore the dominant factors in stand structure and soil nutrients that affect the understory herbaceous diversity of natural secondary Betula platyphylla forest, so as to provide theoretical basis for optimizing the stand structure and improving herbaceous diversity of natural secondary B. platyphylla forest in Qinghai Province. Method Taking the natural secondary B. platyphylla forest in Baoku Forest Farm of Datong County, Qinghai Province as the research object, 20 standard plots (24 m×24 m) were selected by typical sampling method. The effects of multiple factors (stand spatial structure, stand non-spatial structure, and soil nutrient) on herbaceous diversity were investigated by correlation analysis and redundancy analysis. Result (1) A total of 47 species of herbaceous plants belonging to 41 genera and 24 families were found under the B. platyphylla forest, among which Lolium perenne, Fragaria vesca, Polygonum viviparum, Galium bungei and Geranium wilfordii were the dominant species, with mean important values of 19.52%, 18.91%, 10.53%, 6.95% and 5.27%, respectively. (2) Redundancy analysis showed that openness, mean DBH, and size ratio could explain 44.1%, 16.8% and 8.2% of the variation patterns in herbaceous diversity, respectively, and were the main stand factors affecting herbaceous diversity. Alkali-hydrolyzed nitrogen and available potassium could explain 7.9% and 7.6% of herbaceous diversity pattern, respectively, and were the main soil nutrient factors. Conclusion Both stand structure and soil nutrients significantly affect understory herbaceous diversity. The overall level of understory herbaceous diversity can be improved by adjusting stand spatial structure, optimizing stand competition and improving soil fertility. [Ch, 2 fig. 4 tab. 37 ref.] -
Key words:
- Betula platyphylla /
- herbaceous diversity /
- stand structure /
- soil nutrients /
- redundancy analysis
-
图 1 林分结构因子和土壤养分因子与多样性指数的相关性
Figure 1 Correlations between forest structure factors and soil nutrient factors and diversity index
图 2 林分结构因子、土壤因子与草本植物多样性RDA分析结果
Figure 2 RDA analysis results of stand structure factors, soil factors and herbaceous plant diversity
表 1 样地基本信息
Table 1. Basic information table of sample plots
样地编号 海拔/m 纬度(N) 经度(E) 林分密度/(株·hm−2) 郁闭度 平均胸径/cm 平均树高/m 1 2783 37°13′50″ 101°56′17″ 295 0.59 30.52 12.92 2 2936 37°14′47″ 101°62′43″ 312 0.59 14.46 6.54 3 2888 37°12′73″ 101°59′93″ 329 0.67 17.68 8.90 4 2881 37°11′86″ 101°51′20″ 381 0.62 29.50 14.88 5 2894 37°11′83″ 101°51′35″ 399 0.55 32.93 16.91 6 2884 37°14′74″ 101°62′69″ 399 0.61 9.30 9.87 7 2788 37°13′53″ 101°56′05″ 416 0.65 23.94 15.90 8 2847 37°11′90″ 101°51′37″ 434 0.61 28.59 15.93 9 2803 37°12′03″ 101°50′95″ 434 0.6 23.72 12.44 10 2801 37°13′47″ 101°56′17″ 468 0.64 23.36 13.88 11 2855 37°11′90″ 101°51′08″ 503 0.68 28.75 14.27 12 2940 37°14′46″ 101°62′39″ 538 0.69 9.64 6.46 13 2850 37°11′92″ 101°51′20″ 572 0.57 28.22 14.64 14 2940 37°14′69″ 101°62′75″ 572 0.64 17.27 15.92 15 2773 37°13′51″ 101°56′24″ 736 0.63 21.11 13.48 16 2793 37°13′02″ 101°59′50″ 850 0.74 9.53 8.41 17 2946 37°14′47″ 101°62′41″ 850 0.76 8.16 6.01 18 2965 37°14′78″ 101°62′67″ 896 0.78 9.83 13.18 19 2853 37°12′85″ 101°59′60″ 873 0.68 11.21 15.29 20 2868 37°12′78″ 101°59′72″ 638 0.69 10.62 11.19  下载: 导出CSV
下载: 导出CSV
表 2 林下草本植物科内种的组成
Table 2. Composition of species within the understory herb family
种数 科数 科名(属数/种数) ≥7 1 伞形科Apiaceae (6/7) 4~5 3 茜草科Rubiaceae (2/4)、菊科Asteraceae (3/4)、毛茛科Ranunculaceae (5/5) 2~3 5 石竹科Caryophyllaceae (1/2)、紫草科Boraginaceae (2/2)、禾本科Gramineae (2/2)、蓼科(2/3) Polygonaceae、蔷薇科Rosaceae (3/3) 1 16 牻牛儿苗科Geraniaceae (1/1)、瓶尔小草科Ophioglossaceae (1/1)、豆科Fabaceae (1/1)、木贼科Equisetaceae (1/1)、五福花科Adoxaceae (1/1)、唇形科Lamiaceae (1/1)、堇菜科Violaceae (1/1)、芍药科Paeoniaceae (1/1)、玄参科Scrophulariaceae (1/1)、天门冬科Asparagaceae (1/1)、真藓科Bryaceae (1/1)、报春花科Primulaceae (1/1)、十字花科Brassicaceae (1/1)、桔梗科Campanulaceae (1/1)、莎草科Cyperaceae (1/1)
下载: 导出CSV
表 3 样地各变量基本特征
Table 3. Basic characteristics of the variables in the sample plots
项目 非空间结构 空间结构 土壤养分 林分密度
(株·hm−2)郁闭度 平均胸
径/cm平均树
高/m大小比 角尺度 竞争
指数开敞度 pH 有机质/
(g·kg−1)均值 516.15 0.57 16.97 12.50 0.50 0.48 3.25 0.37 7.65 149.78 标准差 250.54 0.07 5.98 2.80 0.07 0.04 0.83 0.12 0.21 13.86 变异系数/% 43.99 12.93 35.22 22.36 13.29 8.26 25.51 31.75 3.19 9.26
下载: 导出CSV
项目 土壤养分 草本多样性 速效钾/
(mg·kg−1)速效磷/
(mg·kg−1)碱解氮/
(mg·kg−1)全磷/
(mg·kg−1)全氮/
(mg·kg−1)物种丰
富度Simpson多样性
指数(D)Shannon-Wiener
多样性指数(H)Pielou均匀度
指数(J)均值 246.56 12.04 0.80 0.68 5.83 12.60 0.96 2.01 0.77 标准差 76.74 9.12 0.09 0.11 1.11 3.14 0.38 0.67 0.12 变异系数/% 31.13 75.74 11.69 15.67 18.98 24.89 39.54 33.48 15.17
下载: 导出CSV
表 4 林分结构因子、 土壤养分因子解释度与蒙特卡洛检验
Table 4. Explanatory degree of stand structure factors and soil nutrient factors and Monte Carlo test
影响因素 解释度/% 贡献率/% F检验 P 影响因素 解释度/% 贡献率/% F检验 P 开敞度 44.1 47.2 14.8 0.002 有机质 2.1 2.3 0.7 0.462 平均胸径 16.8 17.4 7.8 0.010 全氮 1.5 1.6 0.5 0.496 碱解氮 7.9 10.1 5.4 0.036 全磷 0.6 0.7 0.6 0.590 大小比数 8.2 9.6 4.2 0.042 林分密度 0.5 0.6 0.4 0.596 速效钾 7.6 8.1 4.8 0.050 平均树高 0.3 0.3 0.2 0.726 郁闭度 1.1 1.2 1.1 0.322 竞争指数 0.1 0.1 0.1 0.988 速效磷 0.7 0.8 0.6 0.454
下载: 导出CSV
-
[1] 曹小玉, 李际平, 委霞. 亚热带典型林分空间结构与林下草本物种多样性的差异特征分析及其关联度[J]. 草业科学, 2019, 36(10): 2466−2475. CAO Xiaoyu, LI Jiping, WEI Xia. Analysis of the difference and correlation between the spatial structure and understory herbaceous species diversity of typical subtropical forests [J]. Acta Prataculturae Sinica, 2019, 36(10): 2466−2475. [2] 龚艳宾, 郭建斌, 赵秀海, 等. 吉林蛟河天然阔叶红松林草本植物多样性及其与土壤因子的关系[J]. 浙江农林大学学报, 2016, 33(4): 620−628. GONG Yanbin, GUO Jianbin, ZHAO Xiuhai, et al. Herbaceous species diversity as related to soil factors in a Korean pine broadleaved forest of Jiaohe, Jilin Province [J]. Journal of Zhejiang A&F University, 2016, 33(4): 620−628. [3] 胡尔查, 王铮, 李梓豪, 等. 毛乌素沙地不同林龄樟子松人工林林下植物多样性和生物量的动态变化[J/OL]. 生态学杂志, 2024, 43 (11): 3246−3254. HU Ercha, WANG Zheng, LI Zihao, et al. Dynamic changes of understory plant diversity and biomass of Pinus sylvestris var. mongolica plantations at different ages in Mu Us Sandy Land [J/OL]. Chinese Journal of Ecology, 2024, 43 (11): 3246−3254. [4] GILLIAM S F. The ecological significance of the herbaceous layer in temperate forest ecosystems [J]. BioScience, 2007, 57(10): 845−858. [5] 谢政锠, 曹小玉, 赵文菲, 等. 不同龄组杉木公益林林分空间结构与灌木物种多样性[J]. 生态学杂志, 2022, 41(8): 1466−1473. XIE Zhengchang, CAO Xiaoyu, ZHAO Wenfei, et al. Spatial structure and shrub species diversity of different aged stands of Chinese fir public welfare forests [J]. Chinese Journal of Ecology, 2022, 41(8): 1466−1473. [6] 张泽鑫, 蔡有柱, 赵丽娟, 等. 祁连山东部森林林分结构和环境因素对草本物种多样性的影响[J]. 生态学报, 2024, 44(5): 2089−2099. ZHANG Zexin, CAI Youzhu, ZHAO Lijuan, et al. Effects of stand structure and environmental factors on understory herbaceous plant diversity in eastern Qilian Mountains [J]. Acta Ecologica Sinica, 2024, 44(5): 2089−2099. [7] YANG Yan, ZHOU Zhengli, SHEN Liuji, et al. Effects of stand structure of artificial shelter forest on understory herb diversity in desert-oasis ecotone [J/OL]. Diversity, 2023, 15 (10): 1083[2024-05-24]. DOI: 10.3390/d15101083. [8] 陈笑, 李远航, 左亚凡, 等. 林分特征和土壤养分对林下草本物种多样性的影响[J]. 西北植物学报, 2022, 42(8): 1396−1407. CHEN Xiao, LI Yuanhang, ZUO Yafan, et al. Effects of stand characteristics and soil nutrient characteristics on herbaceous diversity [J]. Acta Botanica Boreali-Occidentalia Sinica, 2022, 42(8): 1396−1407. [9] 闫玮明, 孙冰, 裴男才, 等. 粤北阔叶人工林和次生林植物多样性与土壤理化性质相关性研究[J]. 生态环境学报, 2019, 28(5): 898−907. YAN Weiming, SUN Bing, PEI Nancai, et al. Correlation analyses on plant diversity and soil physico-chemical properties between evergreen broad-leaved plantations and natural secondary forest in north Guangdong, China [J]. Ecology and Environmental Sciences, 2019, 28(5): 898−907. [10] 王景玺, 赵浩唱, 宋启民, 等. 林分因子对辽宁章古台樟子松人工林草本植物多样性的影响[J]. 防护林科技, 2022(4): 17−20. WANG Jingxi, ZHAO Haochang, SONG Qimin, et al. Effects of stand factors on herbaceous species diversity in Pines sylvestris var. Mongolica plantations in Zhanggutai area, Liaoning Province [J]. Protection Forest Science and Technology, 2022(4): 17−20. [11] 邓婷婷, 魏岩, 任思远, 等. 北京东灵山暖温带落叶阔叶林地形和林分结构对林下草本植物物种多样性的影响[J]. 生物多样性, 2023, 31(7): 18−29. DENG Tingting, WEI Yan, REN Siyuan, et al. Effects of topography and stand structure of warm temperate deciduous broad-leaved forest on understory herb diversity in Donglingshan, Beijing [J]. Biological Science, 2023, 31(7): 18−29. [12] 邹星晨, 王欣苗, 左亚凡, 等. 青海云杉不同演替阶段林下草本多样性特征及其环境解释[J]. 生态学报, 2023, 43(24): 10285−10294. ZOU Xingchen, WANG Xinmiao, ZUO Yafan, et al. Characteristics of herbaceous diversity and envionmental interpretation of Picea crassifolia at different succession stages [J]. Acta Ecologica Sinica, 2023, 43(24): 10285−10294. [13] 张亚昊, 佃袁勇, 黄光体, 等. 不同演替阶段马尾松林林分空间结构对物种多样性的影响[J]. 生态学杂志, 2021, 40(8): 2357−2365. ZHANG Yahao, DIAN Yuanyong, HUANG Guangti, et al. Effects of spatial structure on species diversity in Pinus massoniana plantation of different succession stages [J]. Chinese Journal of Ecology, 2021, 40(8): 2357−2365. [14] 曹小玉, 李际平. 林分空间结构指标研究进展[J]. 林业资源管理, 2016(4): 65−73. CAO Xiaoyu, LI Jiping. Research progress on indicators of stand spatial structure [J]. Forest Resources Management, 2016(4): 65−73. [15] 赵竹梅. 大通白桦林改造措施及成效分析[J]. 青海农林科技, 2010(3): 86−87. ZHAO Zhumei. Reconstruction measure and effect of birch forest in Datong [J]. Science and Technology of Qinghai Agriculture and Forestry, 2010(3): 86−87. [16] 惠刚盈, 胡艳波. 混交林树种空间隔离程度表达方式的研究[J]. 林业科学研究, 2001, 14(1): 23−27. HUI Gangying, HU Yanbo. Mearsuring species spatial isolation in mixed forests [J]. Forest Research, 2001, 14(1): 23−27. [17] 惠刚盈, 张连金, 胡艳波, 等. 林分拥挤度及其应用[J]. 北京林业大学学报, 2016, 38(10): 1−6. HUI Gangying, ZHANG Lianjin, HU Yanbo, et al. Stand crowding degree and its application [J]. Journal of Beijing Forestry University, 2016, 38(10): 1−6. [18] 南国卫, 王静慧, 秦淑莹, 等. 不同恢复年限刺槐林林下草本层的物种多样性[J]. 浙江农林大学学报, 2024, 41(5): 978−985. NAN Guowei, WANG Jinghui, QIN Shuying, et al. Diversity characteristics of herbaceous species under Robinia pseudoacacia forest in different years [J]. Journal of Zhejiang A&F University, 2024, 41(5): 978−985. [19] 刘灿然, 马克平. 生物群落多样性的测度方法(Ⅴ)生物群落物种数目的估计方法[J]. 生态学报, 1997, 17(6): 39−48. LIU Canran, MA Keping. Measurement of biotic community diversity (Ⅴ) methods for estimating the number of species in a community [J]. Acta Ecologica Sinica, 1997, 17(6): 39−48. [20] 刘俊廷, 张建军, 王恒星, 等. 晋西黄土区不同退耕年限油松林草本多样性与土壤养分的关系[J]. 干旱区研究, 2020, 37(2): 400−409. LIU Junting, ZHANG Jianjun, WANG Hengxing, et al. Relationship between species diversity at the herbaceous stratum and soil nutrients in Pinus tabulaeformis plantations of various age on the loess plateau of western Shanxi Province, China [J]. Arid Zone Research, 2020, 37(2): 400−409. [21] 许崇强, 吴强. 黑麦草应用研究进展及其对石漠化治理的启示[J]. 中国野生植物资源, 2021, 40(10): 66−72. XU Chongqiang, WU Qiang. Research progress on application of ryegrass and its enlightenment to rocky desertification control [J]. Chinese Wild Plant Resources, 2021, 40(10): 66−72. [22] 陈劲松, 董鸣, 于丹. 青藏高原东缘匍匐茎草本野草莓的分株种群特征及其沿海拔梯度的变化[J]. 生态学报, 2003, 23(3): 428−435. CHEN Jinsong, DONG Ming, YU Dan. The characteristics of stoloniferous herb Fragaria vesca L. ramet population and their variation along an altitudinal gradient in the eastern edge of the Qing-Zang Plateau in China [J]. Acta Ecologica Sinica, 2003, 23(3): 428−435. [23] 王玉萍, 高会会, 张峰, 等. 珠芽蓼叶片对海拔变化的表型可塑性[J]. 应用生态学报, 2021, 32(6): 2070−2078. WANG Yuping, GAO Huihui, ZHANG Feng, et al. Altitudinal phenotypic plasticity of leaf characteristics of Polygonum viviparum [J]. Chinese Journal of Applied Ecology, 2021, 32(6): 2070−2078. [24] 边更战, 朱光玉. 八大公山栎类林分空间结构对林下灌木物种多样性影响分析[J]. 中国农学通报, 2020, 36(19): 39−43. BIAN Gengzhan, ZHU Guangyu. Effect of spatial structure of Quercus on species diversity of undergrowth shrubs in Badagong Mountain [J]. Chinese Agricultural Science Bulletin, 2020, 36(19): 39−43. [25] 刘红民, 董莉莉, 高英旭, 等. 辽东山区典型次生林空间结构对草本物种多样性的影响[J]. 沈阳农业大学学报, 2020, 51(6): 670−679. LIU Hongmin, DONG Lili, GAO Yingxu, et al. Effects of spatial structure on herbaceous diversity of typical secondary forest in mountainous area of eastern Liaoning [J]. Journal of Shenyang Agricultural University, 2020, 51(6): 670−679. [26] 马刘乐, 张梦弢. 不同干扰强度下林分空间结构对灌草层物种多样性的影响[J]. 西北林学院学报, 2023, 38(3): 63−69. MA Liule, ZHANG Mengtao. Effects of stand spatial structure on species diversity in shrub grass layer under different disturbance intensities [J]. Journal of Northwest Forestry University, 2023, 38(3): 63−69. [27] 黎芳, 潘萍, 宁金魁, 等. 飞播马尾松林林分空间结构对林下植被多样性的影响[J]. 东北林业大学学报, 2016, 44(11): 31−35, 40. LI Fang, PAN Ping, NING Jinkui, et al. Effects of stand spatial structure on understory vegetation diversity of aerial seeding Pinus massoniana plantations [J]. Journal of Northeast Forestry University, 2016, 44(11): 31−35, 40. [28] 曹小玉, 李际平, 赵文菲, 等. 基于结构方程模型分析林分空间结构对草本物种多样性的影响[J]. 生态学报, 2020, 40(24): 9164−9173. CAO Xiaoyu, LI Jiping, ZHAO Wenfei, et al. Effects of stand spatial structure on herbaceous species diversity in forests based on structural equation modeling [J]. Acta Ecologica Sinica, 2020, 40(24): 9164−9173. [29] 牛一迪, 蔡体久. 大兴安岭北部次生林演替过程中物种多样性的变化及其影响因子[J]. 植物生态学报, 2024, 48(3): 349−363. NIU Yidi, CAI Tijiu. Changes in species diversity and influencing factors in secondary forest succession in northern Da Hinggan Mountains [J]. Chinese Journal of Plant Ecology, 2024, 48(3): 349−363. [30] 张勇强, 李智超, 厚凌宇, 等. 林分密度对杉木人工林下物种多样性和土壤养分的影响[J]. 土壤学报, 2020, 57(1): 239−250. ZHANG Yongqiang, LI Zhichao, HOU Lingyu, et al. Effects of stand density on understory species diversity and soil nutrients in Chinese fir plantation [J]. Acta Pedologica Sinica, 2020, 57(1): 239−250. [31] 张林, 齐实, 周飘, 等. 北京山区侧柏林下草本植物多样性的影响因素分析[J]. 草地学报, 2022, 30(8): 2199−2206. ZHANG Lin, QI Shi, ZHOU Piao, et al. Analysis of influencing factors on the understory herbaceous plant diversity of Platycladus orientalis forest in Beijing mountainous areas [J]. Acta Agrestia Sinica, 2022, 30(8): 2199−2206. [32] 许丽, 丰菲, 刘莹, 等. 煤矸石山植物物种多样性与土壤化学因子的关系: 以灵武矿区生态修复初期为例[J]. 煤炭科学技术, 2020, 48(4): 97−104. XU Li, FENG Fei, LIU Ying, et al. Relationship between plant species diversity and soil chemical properties in coal gangue dump: early stage of ecological restoration in Lingwu Mining Area [J]. Coal Science and Technology, 2020, 48(4): 97−104. [33] 高晟, 王磊, 薛建辉, 等. 贵州喀斯特地区草本植被盖度与土壤养分的相互关系[J]. 南京林业大学学报(自然科学版), 2012, 36(1): 79−83. GAO Sheng, WANG Lei, XUE Jianhui, et al. The relationship between coverage of herbaceous vegetation communities and soil nutrients in the karst area of Guizhou province [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2012, 36(1): 79−83. [34] ARQUERO O, BARRANCO D, BENLLOCH M. Potassium starvation increases stomatal conductance in olive trees [J]. HortScience, 2006, 41(2): 433−436. [35] CAKMAK I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants [J]. Journal of Plant Nutrition and Soil Science, 2005, 168(4): 521−530. [36] 王树梅, 庞元湘, 宋爱云, 等. 基于林龄的滨海盐碱地杨树刺槐混交林土壤理化性质及草本植物多样性动态[J]. 生态学报, 2018, 38(18): 6539−6548. WANG Shumei, PANG Yuanxiang, SONG Aiyun, et al. Soil physicochemical properties and diversity of herbaceous plants dynamic on the different ages mixed forests of Populus × Euramercana ‘Neva’ and Robinia pseucdoacacia in coastal saline-alkail area [J]. Acta Ecologica Sinica, 2018, 38(18): 6539−6548. [37] PONTIGO S, PARRA-ALMUNA L, LUENGO-ESCOBAR A, et al. Biochemical and molecular responses underlying the contrasting phosphorus use efficiency in ryegrass cultivars [J/OL]. Plants, 2023, 12 (6): 1224[2024-05-24]. DOI: 10.3390/plants12061224. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240383
 点击查看大图
点击查看大图
计量
- 文章访问数: 1340
- HTML全文浏览量: 418
- PDF下载量: 30
- 被引次数: 0
