




-
种群的空间分布格局是指在一定时间内,个体在其生活空间中的位置、状态和布局[1]。研究种群空间分布格局是揭示植物群落演替动态的基础,是合理安排森林经营活动的基本前提[2],一直以来都是生态理论研究的核心[3−4],判定种群的空间分布类型和空间关联性为其主要的研究内容[5]。空间分布格局的研究方法众多,主要有扩散系数/方差平均值比法、平均拥挤度法、最近邻体法等,但这些方法只能分析特定尺度环境下的种群分布格局。由于种群空间分布格局和关联性对空间尺度的依赖性较强,因此这些方法在一定程度上存在局限性[6]。点格局分析方法以种群个体的空间位置坐标为基础,能最大限度地利用点与点间的距离,分析任意尺度下种群的空间格局特征及关联性,因而得到了广泛应用,已成为国际上分析种群格局的主要方法之一[7]。
辽东栎Quercus mongolica林是暖温带落叶阔叶林区域典型的地带性植被类型之一,在山西分布广泛,是山西森林的主要组成部分[8]。目前,对于辽东栎的研究集中在生长模型构建[9]、生物多样性[10]、林分空间结构[11]、生态位特征[12]等方面,对辽东栎林空间分布格局的研究主要在宁夏的六盘山[13]、山西的灵空山[14]和芦芽山[15]等地,且这些研究多以辽东栎与油松Pinus tabuliformis形成的共优种为研究对象。有研究表明:种群的空间分布格局在不同生境和不同群落类型中表现不同[16],因此对以辽东栎为主要优势种的天然林进行研究,有助于了解同一树种在不同群落中空间分布格局形成过程的差异。本研究以吕梁山西侧城庄沟林场的辽东栎天然林为研究对象,分析其优势种群结构和空间分布格局,探讨不同径级优势种的空间分布格局,不同径级优势种的种内和种间空间关联性,以便了解该地区辽东栎天然林优势植物的分布形态及其与环境之间的关系,对比不同地区辽东栎林空间分布格局的差异性。研究结果有助于认识辽东栎天然林内物种的共存机制,可为辽东栎天然林的合理经营和人工辽东栎林的改造提供参考。
-
研究区位于山西省黑茶山国有林管理局城庄沟林场(37°58′~38°09′N,111°11′~111°17′E),吕梁山西侧,海拔为1 400~1 900 m,林地多分布于阴坡,属中高山天然林区。主要天然植被为针阔混交林和落叶阔叶林,乔木主要为辽东栎、油松、山杨Populus davidiana、白桦Betula platyphylla、茶条槭Acer ginnala等,灌木主要有绣线菊Spiraea pubescens、黄刺玫Rosa xanthina、虎榛子Ostryopsis davidiana等,草本主要为薹草Carex lanceolata等。气候属暖温带大陆性气候,年平均气温为8.8 ℃,≥10 ℃年平均积温为3 212.0 ℃,年无霜期为160.0 d,年平均降水量为518.8 mm。
-
2021年在海拔1 700 m的半阴坡无人为干扰的辽东栎天然林设立样地,面积为160 m×60 m,采用相邻格子法将样地划分为24个20 m×20 m的小样方,调查并记录样地内所有胸径≥2.5 cm的乔木树种及胸径、坐标等详细信息。
-
以物种重要值确定样地内的优势种。重要值计算公式为:重要值=(相对多度+相对频度+相对显著度)/3[6],其中相对多度指某物种株数占所有种总株数的百分比,相对频度指某物种在20 m×20 m样方中出现的次数占所有种出现次数的百分比,相对显著度指某物种胸高断面积占所有种胸高断面积的百分比。
-
结合文献[14],确定起测胸径(DBH)为2.5 cm,径级划分为:Ⅰ级(2.5 cm≤DBH<7.5 cm)、Ⅱ级(7.5 cm≤DBH<12.5 cm)、Ⅲ级(12.5 cm≤DBH<17.5 cm)、Ⅳ级(17.5 cm≤DBH<22.5 cm)、Ⅴ级(22.5 cm≤DBH<27.5 cm)、Ⅵ级(27.5 cm≤DBH<32.5 cm)、Ⅶ级(32.5 cm≤DBH<37.5 cm)、Ⅷ级(37.5 cm≤DBH<42.5 cm)。
-
优势种群的空间分布格局采用Ripley的L函数计算,公式为:$ \mathop L\limits^{} (t) = \sqrt {\mathop K\limits^{} (t)/{\text{π}} } - t $[17−18]。不同径级间的关联性采用多元点格局分析计算,公式为:$ {\mathop L_{12} }(t) = \sqrt {\mathop {{K_{12}}}\limits^{} (t)/{\text{π}}} - t $[18−19]。其中:$ \mathop K\limits^{} (t) = \dfrac{A}{{{n^2}}}\displaystyle\sum\limits_{i = 1}^n {\displaystyle\sum\limits_{j = 1}^n {w_{ij}^{ - 1}{I_t}({u_{ij}})} } (i \ne 1) $,$ \mathop {{K_{12}}}\limits^{} (t) = \dfrac{A}{{{n_1}{n_2}}}\displaystyle\sum\limits_{i = 1}^{{n_1}} {\displaystyle\sum\limits_{j = 1}^{{n_2}} {w_{ij}^{ - 1}{I_t}({u_{ij}})} } (i \ne j) $, t为空间尺度,A为样地面积,n为林木株数,n1和n2为样地内2个不同径级的林木株数,K(t)公式中i和j分别为1个种群的任意2株林木,K12(t)公式中i和j分别为2个径级的任意1株林木,$ {u_{ij}} $为2株林木i和j间的距离,$ {I_t}({u_{ij}}) $为指示函数,当$ u_{i j} $≤t时,$ I_{t}\left(u_{i j}\right) $=1,$ u_{i j} $>t时,$ I_{t}\left(u_{i j}\right) $=0,wij为权重值,用于边缘矫正。采用Monte Carlo拟合检验,随机拟合99次,计算上下包迹线,得到99%的置信区间,L(t)值和L12(t)值若在包迹线以内,则种群符合随机分布,不同径级间无关联;在上包迹线以上则种群属于聚集分布,不同径级间正关联;在下包迹线以下则种群呈均匀分布,不同径级间负关联。
优势种群的空间分布格局和不同径级间的关联性均在Programita软件中完成分析。
-
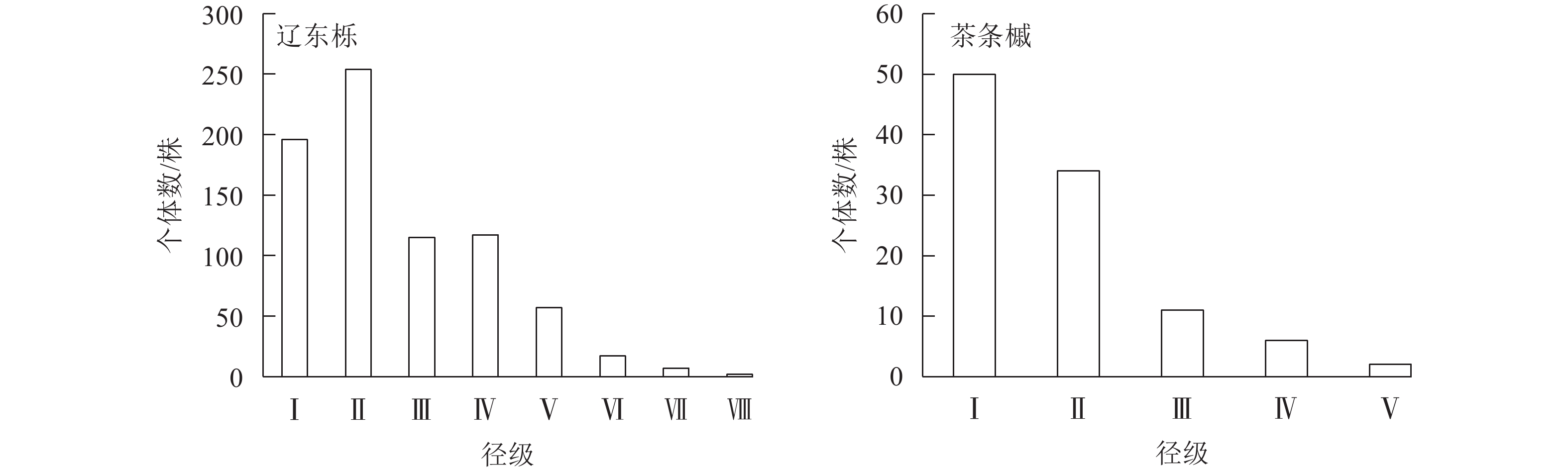
从表1可看出:辽东栎天然林中共出现8种乔木植物,其中辽东栎在所有样方中均出现,重要值最大,达58.76%,且个体数(相对多度)和胸径在乔木层中最大,为样地乔木层的建群种。茶条槭重要值次之,为11.06%,其个体数和出现的样方数仅次于辽东栎,小胸径林木数量较多,导致相对显著度小于山杨,为样地的主要伴生种。其余6种乔木的重要值均较小,故将辽东栎和茶条槭作为优势种。
表 1 辽东栎天然林群落各树种的重要值
Table 1. Importance values of tree species in Q. mongolica community
树种 相对多度/% 相对频度/% 相对显著度/% 重要值/% 树种 相对多度/% 相对频度/% 相对显著度/% 重要值/% 辽东栎 69.80 25.81 80.66 58.76 山丁子 3.83 12.90 2.78 6.51 茶条槭 9.67 18.28 5.24 11.06 山杏 5.57 12.90 2.30 6.92 白桦 1.64 7.53 1.09 3.42 油松 1.19 8.60 0.68 3.49 山杨 6.84 7.53 5.88 6.75 鹅耳枥 1.46 6.45 1.37 3.09 说明:山丁子Malus baccata;山杏Prunus sibirica;鹅耳枥Carpinus turczaninowii。 群落中2种优势种的径级结构相似,表现为随径级增加个体数减少的趋势,径级分布呈倒“J”型特性(图1),即小径级个体(DBH<12.5 cm)较多。林内更新较好,处于增长阶段。
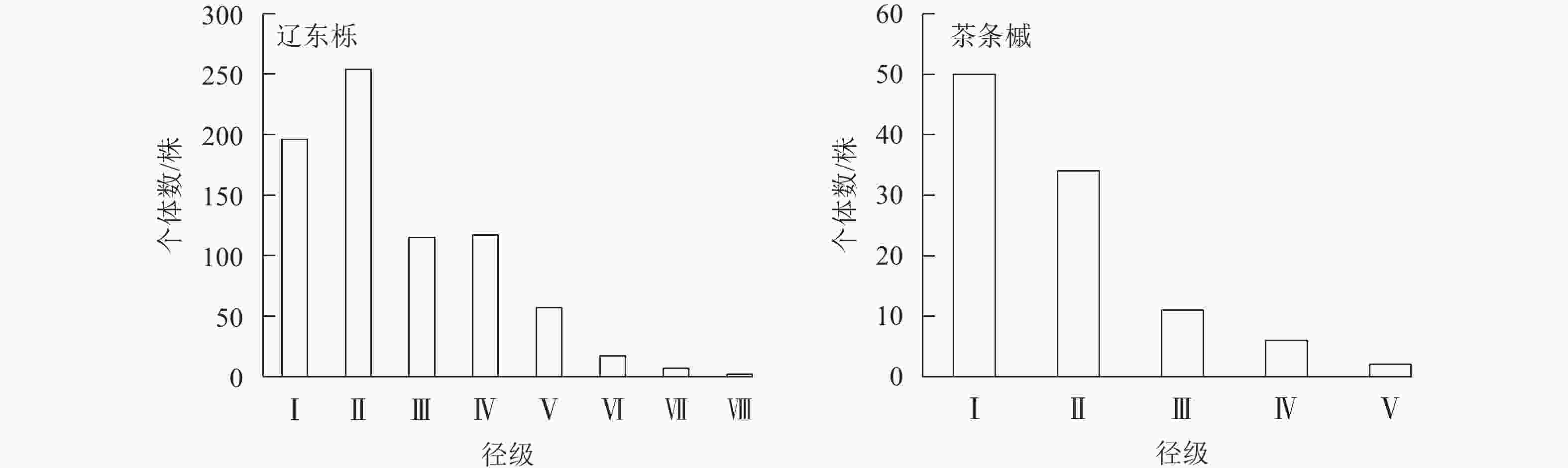
图 1 优势种的径级结构
Figure 1. Diameter class structure of dominant species
-
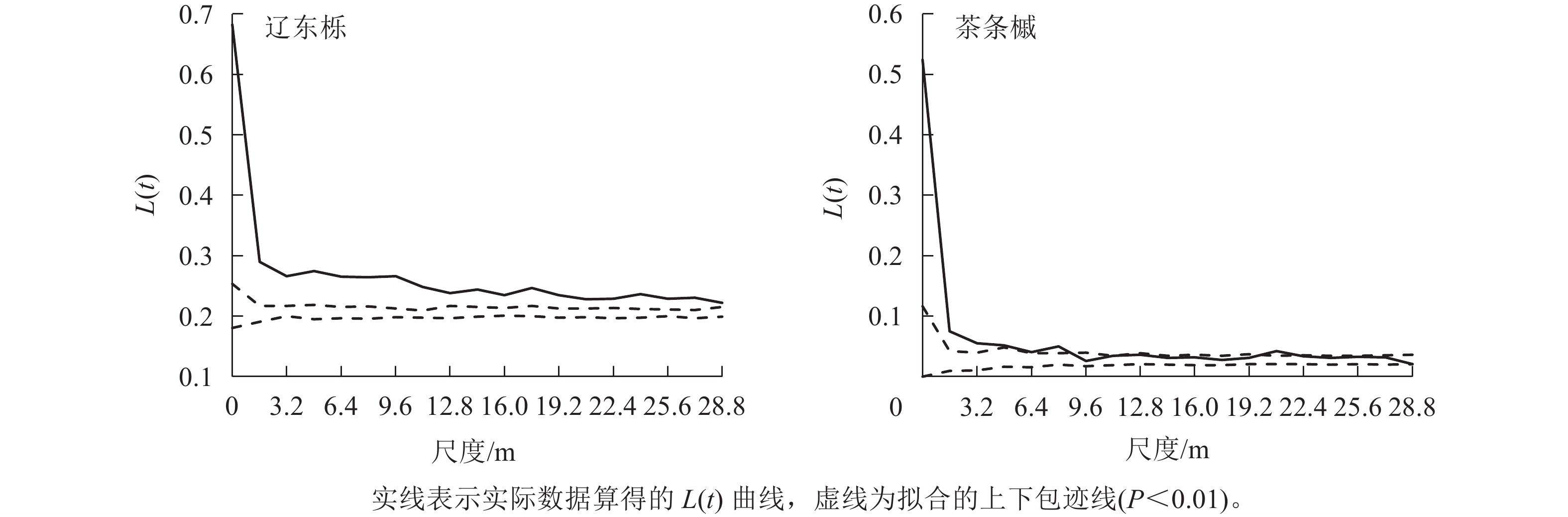
辽东栎和茶条槭种群的空间分布格局不尽相同。辽东栎种群在整个研究尺度均呈聚集分布,但随尺度增大,聚集程度逐渐减小。茶条槭种群在0~8.0 m尺度内表现为聚集分布,但随尺度增大聚集程度逐渐减小,在20.8 m尺度上呈程度很小的聚集分布,其余尺度上均呈随机分布(图2)。
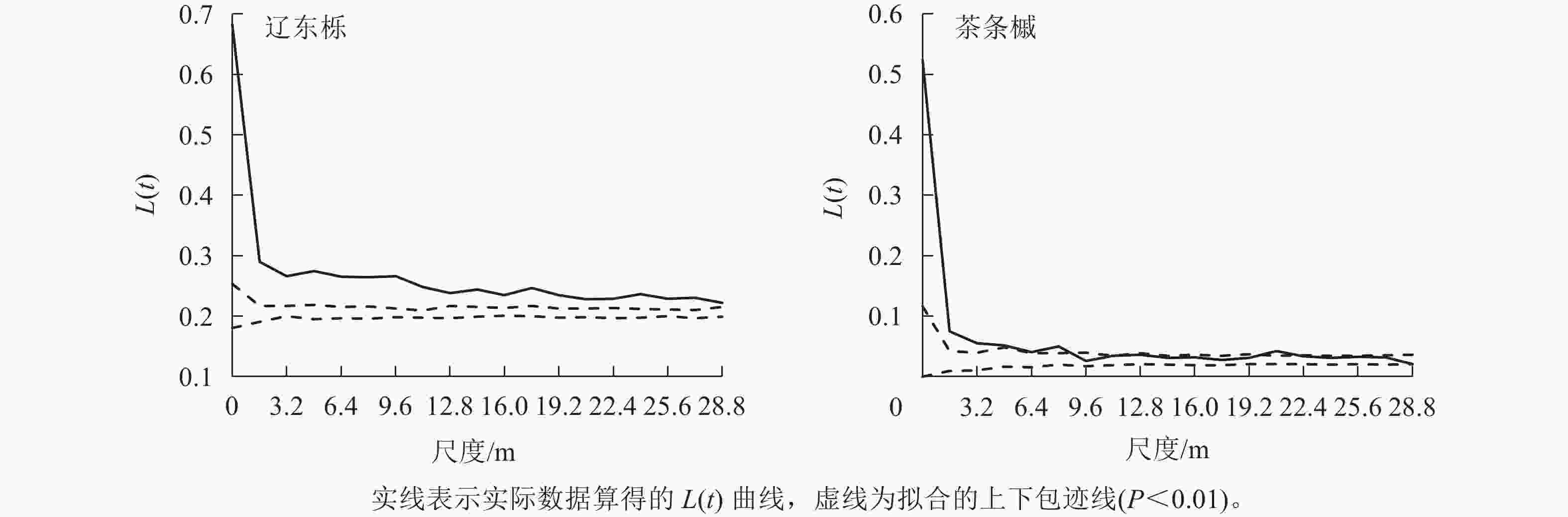
图 2 优势种群的空间分布格局
Figure 2. Spatial distribution pattern of dominant species
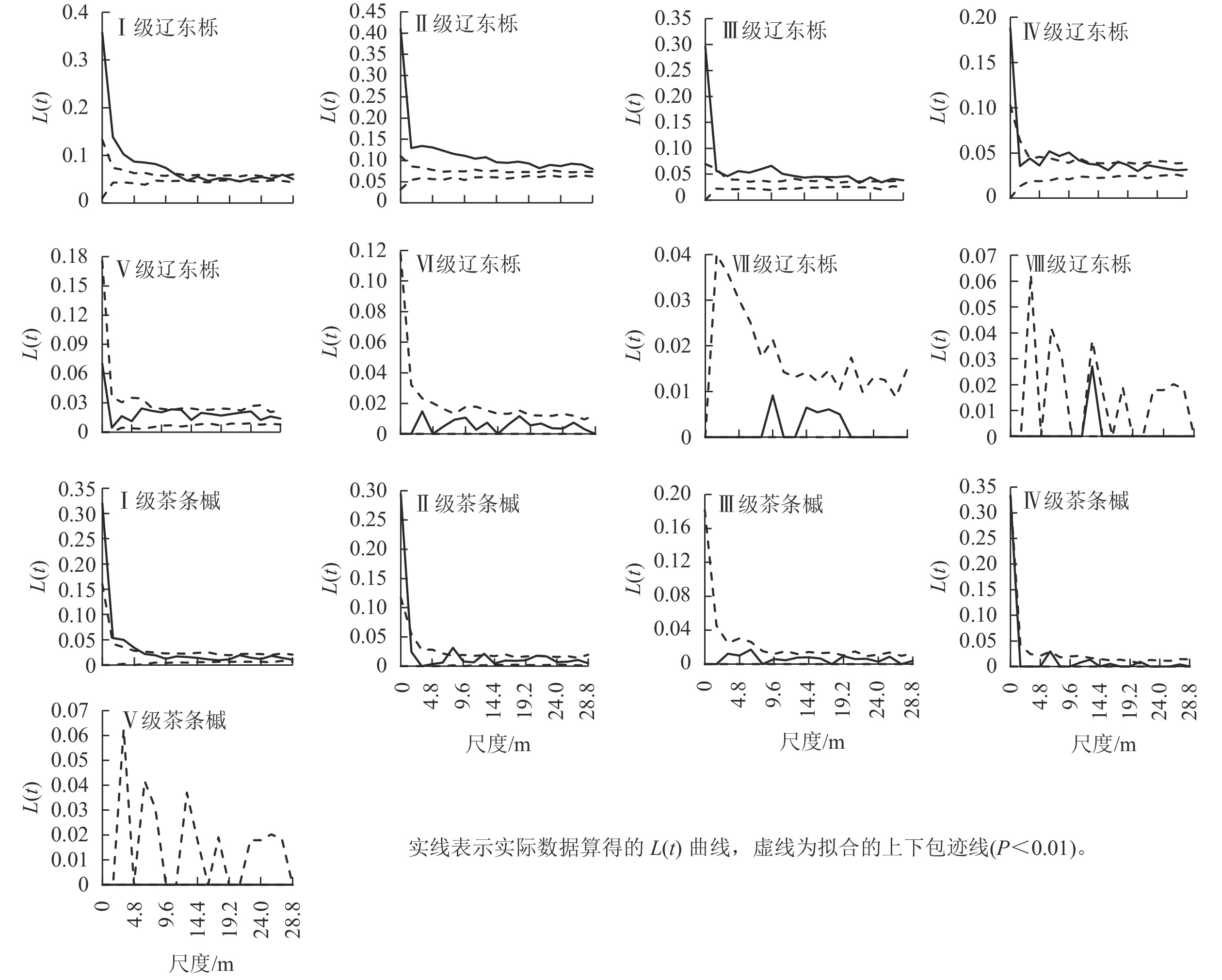
优势种群不同径级的空间分布格局如图3,其中Ⅰ~Ⅳ级辽东栎(DBH<22.5 cm)在研究尺度上呈不同程度的聚集分布与随机分布的组合,Ⅰ级辽东栎在0~11.2 m尺度上呈聚集分布,其余尺度为随机分布,Ⅱ、Ⅲ级辽东栎在整个研究尺度上几乎均为聚集分布,且聚集程度随研究尺度增大而减小,Ⅳ级辽东栎在0~1.6和6.4~9.6 m尺度上有聚集,其余尺度为随机分布。Ⅴ级及以上辽东栎在整个研究尺度均呈随机分布。可见,辽东栎的聚集分布仅表现为中小径级林木。
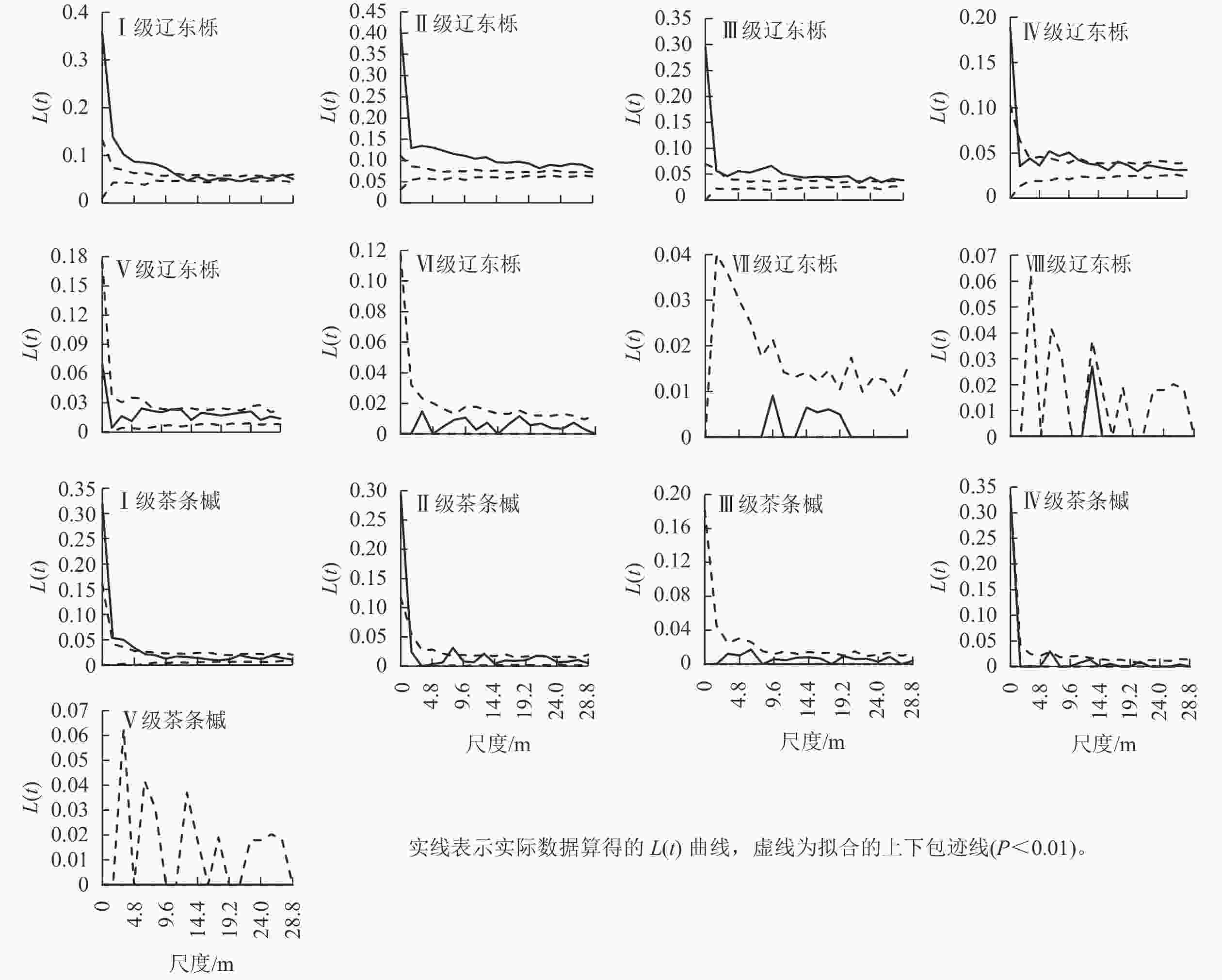
图 3 优势种群不同径级的空间分布格局
Figure 3. Spatial distribution pattern of dominant species at different diameter class
Ⅰ、Ⅱ级茶条槭在小尺度上为聚集分布,随尺度增大其分布格局变为随机分布。其中Ⅰ级茶条槭在0~4.8 m尺度上为聚集分布,Ⅱ级茶条槭在0~1.6和8.0 m尺度上为聚集分布,其余尺度和其他径级在整个研究尺度上均为随机分布。
-
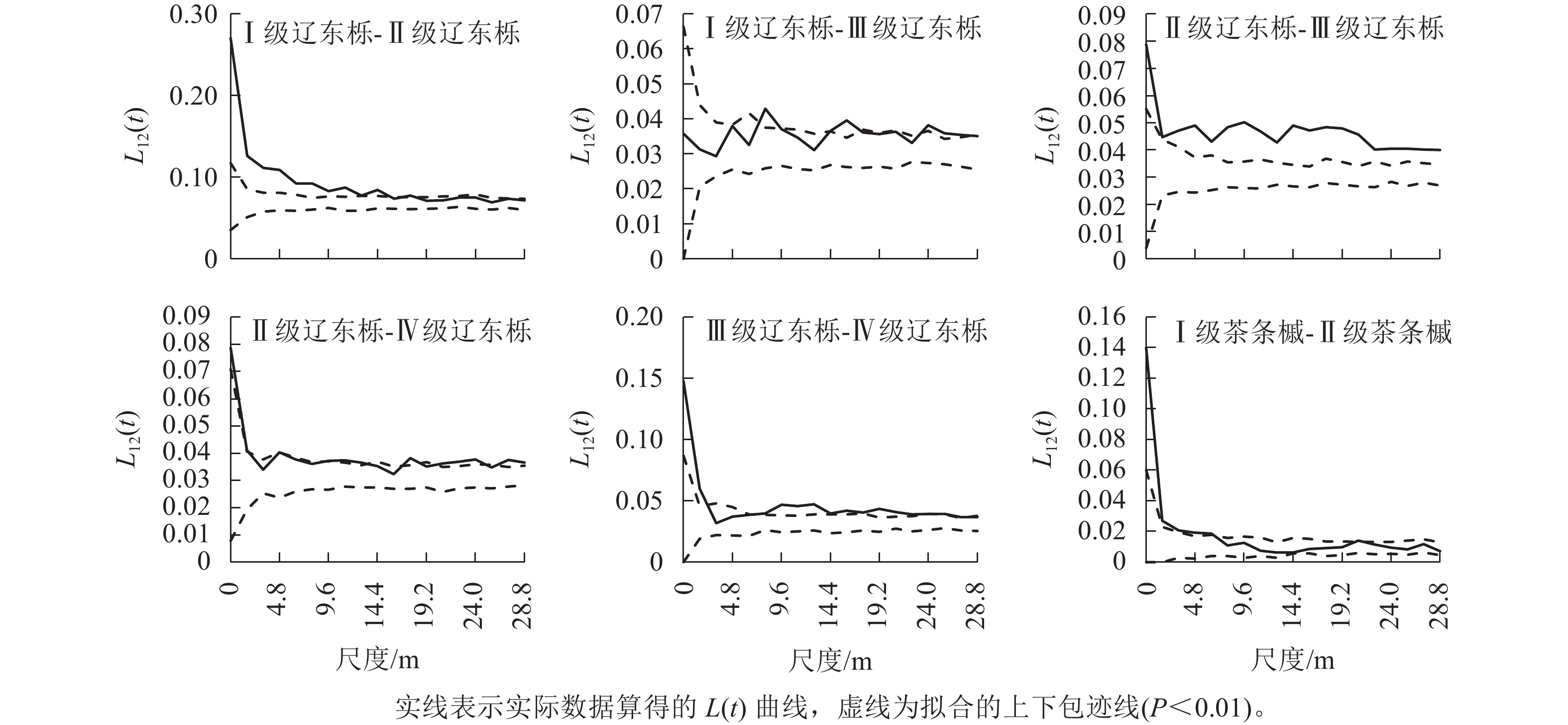
本研究中优势种径级较多,得到的空间关联性图数量大。为便于说明,仅图4和图5展示了辽东栎和茶条槭种内和种间存在空间正关联和负关联的径级。辽东栎种内的关联性主要出现在Ⅰ~Ⅳ级林木间,其中Ⅰ级林木与Ⅱ级林木在空间尺度0~14.4 m呈正关联,其余尺度呈无关联,Ⅰ级林木与Ⅲ级林木在8.0、14.4~16.0、24.0~27.2 m尺度上呈正关联,其余尺度为无关联,Ⅱ级林木与Ⅲ级林木在整个研究尺度上均呈正关联,且关联程度较大,Ⅱ级林木与Ⅳ级林木在研究尺度内以正关联为主,但关联性较弱,在3.2、6.4~8.0、14.4~16.0、19.2和25.6 m尺度上表现为无关联,Ⅲ级林木与Ⅳ级林木仅在3.2~6.4 m尺度上为无关联,其余尺度均为正关联。可见,辽东栎种内关联性仅存在于中小径级林木间,且以正关联为主,径级差距越小,正关联的程度和空间尺度范围越大。
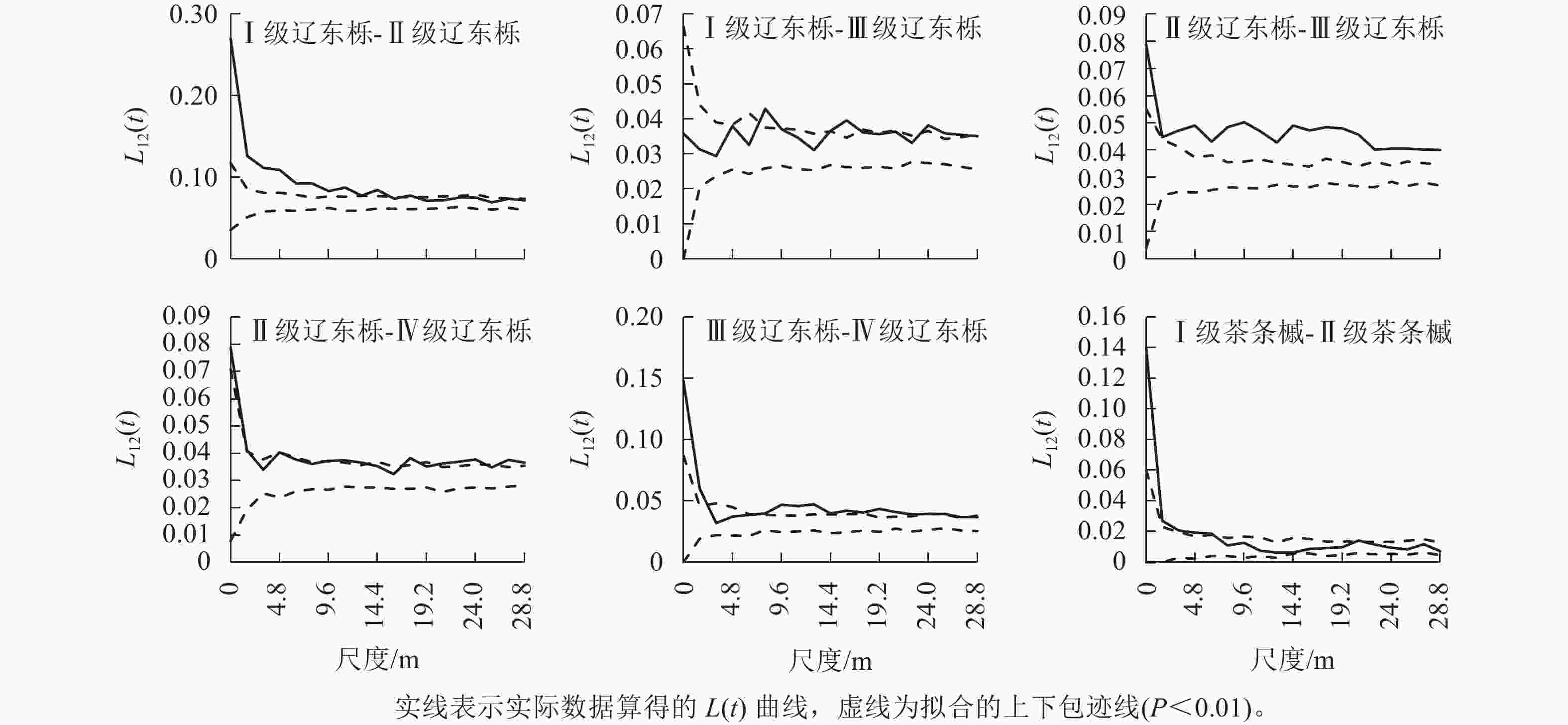
图 4 优势种的种内空间关联性
Figure 4. Intraspecies spatial correlation of dominant species
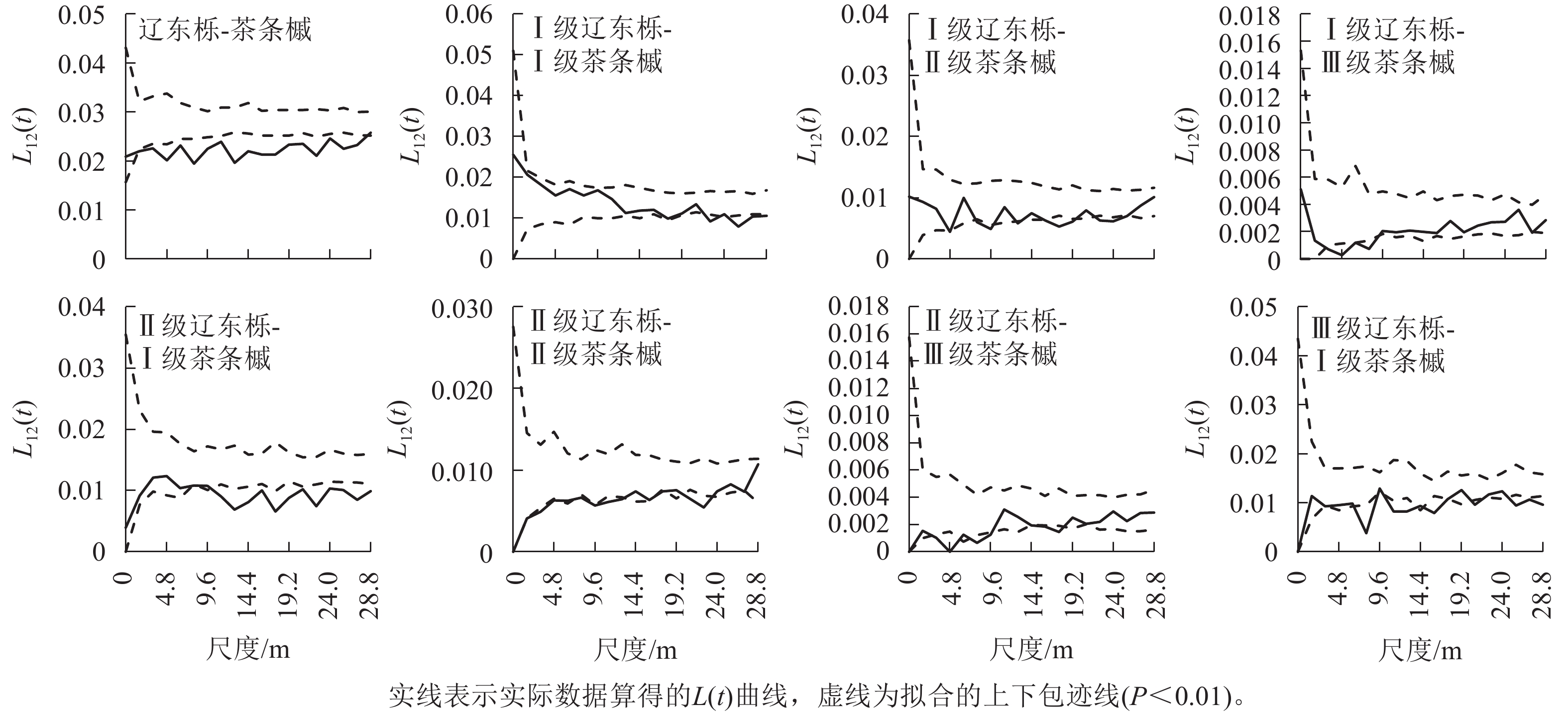
茶条槭种内关联性仅出现在Ⅰ级林木与Ⅱ级林木间,且L12(t)值仅在0~6.4 m尺度内位于置信区间上方,呈正关联,但其与上包迹线接近,关联性较弱,其余尺度上表现为无关联。
辽东栎与茶条槭种群间以负关联为主(图5),L12(t)值仅在0~1.6 m尺度内及28.8 m处位于置信区间内呈无关联,其余尺度均为负关联,说明辽东栎与茶条槭种群间对于资源的竞争较为激烈。
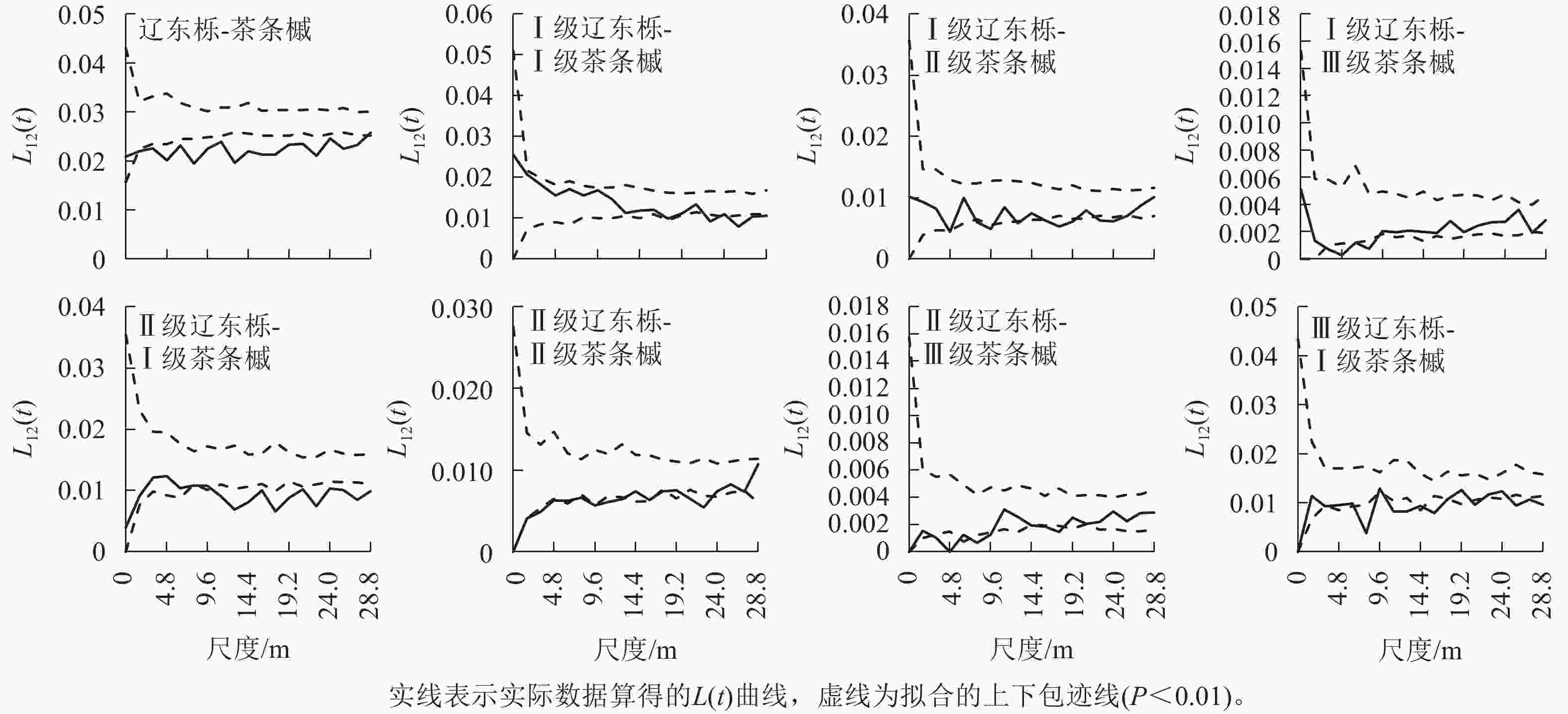
图 5 优势种的种间空间关联性
Figure 5. Interspecies spatial correlation of dominant species
辽东栎与茶条槭不同径级的种间关联性主要出现在不同种群的Ⅰ~Ⅲ级林木间,且为负关联,较大径级种间均无关联性,Ⅰ级辽东栎与Ⅰ级茶条槭的负关联出现在较大空间尺度(22.4~28.8 m),在小尺度上表现为无关联,与Ⅱ级茶条槭在4.8、8.0~9.6、12.8、16.0~19.2、22.4~25.6 m尺度上均呈负关联,与Ⅲ级茶条槭仅在3.2~8.0 m尺度上呈负关联;Ⅱ级辽东栎与Ⅰ级茶条槭在小尺度上无关联,在较大尺度(11.2~28.8 m)上呈负关联,与Ⅱ级茶条槭在1.6~6.4、8.0~12.8、20.8~22.4 m尺度上呈负关联,与Ⅲ级茶条槭在3.2~4.8、8.0~9.6、14.4~17.6 m尺度上呈负关联;Ⅲ级辽东栎与Ⅰ级茶条槭在3.2、8.0、11.2~12.8、16.0~17.6、20.8、25.6~28.8 m尺度上呈负关联。可见,优势种群间的关联性仅出现在中小径级间,且随茶条槭径级的增大负关联的尺度由大变小,随辽东栎径级的增大负关联的尺度增大。
-
径级结构能反映种群生长发育状况和植物群落的稳定性[17],幼龄个体数量是维持种群稳定性的基础[20]。本研究中优势种群辽东栎和茶条槭的幼龄个体数量均较大,种群后备资源丰富,均为增长型,具有较强的持续发育能力。这与它们自身的生物学特性有关。辽东栎在林下荫蔽湿润的环境下更利于萌发[21],茶条槭结实量大,种子可通过风媒散布而产生大量幼苗[22]。本研究中Ⅰ级辽东栎个体数量较Ⅱ级稍少,这与前人的研究结果[21]类似,原因可能与环境过滤有关。辽东栎从种子萌发到生长至Ⅰ级时会经历对光、水和土壤养分等有限资源的强烈竞争,出现死亡高峰,但辽东栎具有很强的萌生更新能力,其种群顺利渡过这一更新瓶颈之后保持相对稳定[23]。采用黄维等[24]在研究油松飞播林林分空间结构中使用的熵权法确定权重,进而构建空间结构综合指数的方法,对辽东栎天然林林分空间结构进行分级评价,其空间结构综合指数为0.603 5,属于4级,即空间结构较好。王志鸣等[25]认为:林分空间结构越好,其稳定性越高。由此可见,本研究中的辽东栎天然林的稳定性较高。
种群分布格局受植物生物学特性和生境因子的共同影响[16]。辽东栎种子较大,下落位置集中在母树周围,这一生物学特性使其种群在研究尺度内呈聚集分布。这一结果与王燕等[14]对太岳山针阔混交林中辽东栎的研究结果一致,但与张中惠等[13]在六盘山辽东栎林的研究结果不同,从而验证了生境和群落类型不同种群的空间分布格局不尽相同的观点[16]。茶条槭为翅果,易随风飘落,种子特性并不会造成聚集分布,而且茶条槭为喜光树种,林下更新受控于上层林隙和枯立木冠层空隙[22],因此林下光照条件是限制茶条槭更新的主要生境因子,也是茶条槭聚集分布格局形成的主导因子。种群的空间分布格局随尺度而变化,具有强烈的尺度依赖性[26],因此茶条槭种群总体呈现出小尺度上聚集而大尺度随机分布的格局,辽东栎种群随尺度增加聚集程度降低也与此有关。种群的空间分布格局受径级的影响,不同径级的空间分布格局不同[27]。辽东栎和茶条槭种群均表现为小径级个体在小尺度上以聚集分布为主,大径级个体为随机分布格局,这与温带地区的许多研究结果一致[18, 28]。原因可能是小径级个体生命力脆弱,生长过程中需要多个个体聚集起来应对外界不利因素,以此来提高存活率。
空间关联性反映了物种对有限资源的竞争结果,或者是恶劣条件下物种在生存中相互促进的结果[29]。本研究中,辽东栎种群种内仅在中小径级林木间表现为正关联,且径级差距越小,正关联的程度和空间尺度范围越大,茶条槭种群仅在小尺度的Ⅰ级林木和Ⅱ林木间表现为正关联。可见,径级差距与种内空间关联性密切相关,这与陈列等[30]对长白山北坡红松Pinus koraiensis和紫椴Tilia amurensis种群的研究结果及涂洪润等[17]对桂林岩溶石山檵木Loropetalum chinense种群的研究结果一致。辽东栎和茶条槭均表现为小径级个体数量多,分布密度大,在不同尺度范围内聚集分布。小径级个体间竞争资源的能力较低,更需要彼此相互协调来抵抗不利因素[1],故正关联的尺度和程度较大。随着林木生长,在资源限制的条件下,与大径级林木相比,中、小径级林木对资源的竞争力较小,它们只有相互庇护才有利于发挥群体效应,增大存活机会,以适应长期的自然选择,故中、小径级林木间也表现为正关联。当林木生长到较大径级后,在长期的发育过程中适应了当地环境,占据了适宜的生态位,对资源可以相互分享和协调,故种内中小径级林木间互不干扰,表现出无关联的现象。辽东栎与茶条槭种群间总体关联性表现为随尺度增加,呈无关联转变为负关联再到无关联的特点,不同径级种群间的关联性仅出现在中小径级林木间,且随茶条槭径级的增大负关联的空间尺度变小,随辽东栎径级的增大负关联的空间尺度范围增大。原因可能与这2个优势种更新所需要的生境条件不同有关,但随径级增大种群间对资源的竞争更加激烈,故其负关联尺度会由大尺度向小尺度靠近。这也说明了种群间的空间关联性是植物与环境和资源利用相适应的结果。
一般认为,群落处于演替初级阶段时,物种间的关联程度较低,甚至产生较大的负关联。本研究中优势种群辽东栎与茶条槭间以负关联为主的种间关系,表明该群落仍处于演替初级阶段。优势种群更新状态良好,且种内在一定尺度范围内均表现为正关联,说明优势种群处于相对稳定的状态。邹书珍等[31]在子午岭对4个不同演替阶段的油松种群进行研究,发现演替初级阶段种群径级结构表现为增长型,王艳等[14]依据种群稳定增长且老龄个体数量较少而确定群落处于演替初级阶段。本研究中优势种群相对稳定,且增长型的径级结构进一步证实了该群落处于演替初级阶段。
-
本研究基于吕梁山西侧0.96 hm2辽东栎天然林样地的调查数据,分析了辽东栎和茶条槭2个优势种群的径级结构和空间分布格局。研究发现:2个优势种群幼龄个体数量均较大,后备资源丰富,处于稳定增长状态。辽东栎种群在研究尺度内呈聚集分布,茶条槭种群则在小尺度为聚集分布,大尺度为随机分布。辽东栎种群中小径级林木间表现为正关联,茶条槭种群仅在小尺度的Ⅰ级和Ⅱ级林木间表现为正关联,优势种群间总体关联性随尺度增加,由无关联转变为负关联再转变为无关联,中小径级种群间表现为负关联。该地辽东栎天然林处于较稳定的演替初级阶段,优势种群结构稳定,更新状态良好。为加快演替进程,可适当辅以人工干预,促进林分向顶级群落发展。
Point pattern analysis of dominant populations in Quercus mongolica natural forest on the west side of Lüliang Mountain
-
摘要:
目的 探究山西主要森林类型辽东栎Quercus mongolica天然林的树种空间分布格局,掌握其生长发育现状,为该林分的合理经营和人工辽东栎林的改造提供参考。 方法 以吕梁山西侧城庄沟林场2021年调查的0.96 hm2辽东栎天然林为研究对象,用径级结构代替年龄结构,并运用点格局分析方法分析优势种群不同径级个体的空间分布格局及空间关联性。 结果 在该林分中,辽东栎和茶条槭Acer ginnala为主要优势种,其径级结构均呈倒“J”型,为增长型,种群结构稳定、更新良好。在研究尺度内辽东栎种群呈聚集分布,茶条槭种群则表现为小尺度聚集分布,大尺度随机分布的格局。2个优势种群的小径级个体在小尺度上以聚集分布为主,大径级个体为随机分布格局。辽东栎种群种内关联性仅在中小径级林木间表现为正关联,且径级差距越小,正关联的程度和空间尺度范围越大,茶条槭种群仅在小尺度的Ⅰ级和Ⅱ级林木间表现为正关联。随尺度增加,优势种群间总体关联性由无关联转变为负关联再转变为无关联,不同径级种群间的空间关联性仅出现在中小径级林木间,且为负关联。 结论 该辽东栎天然林仍处于相对稳定的演替初级阶段,优势种群结构稳定,更新状态良好。为加快演替进程,可适当辅以人工干预,促进林分向顶级群落发展。图5表1参31 Abstract:Objective This study aims to explore the spatial distribution pattern of tree species in Quercus mongolica natural forest, a major forest type in Shanxi Province, and understand its growth and development status, so as to provide reference for the rational management of the forest and conversion of Q. mongolica plantation. Method The research object was a 0.96 hm2 plot of Q. mongolica natural forest investigated in 2021 in Chengzhuanggou Forest Farm on the west side of Lüliang Mountain. The diameter class structure was used instead of the age structure. The spatial distribution pattern and spatial correlation of individuals with different diameter levels in the dominant population were analyzed by point pattern analysis. Result Q. mongolica and Acer ginnala were dominant species, and their diameter structure was an inverted “J ” type, indicating a growth type with stable population structure and good regeneration. The population of Q. mongolica showed a cluster distribution within the research scale, while that of A. ginnala exhibited a pattern of small-scale cluster distribution and large-scale random distribution. The small diameter individuals of the two dominant populations were mainly clustered in small scale, while the large diameter individuals were randomly distributed. The intraspecies association of Q. mongolica was positive only among middle and small size trees, and the smaller the diameter difference, the greater the degree and spatial scale range of positive association. The population of A. ginnala showed positive correlation only between small-scale GradeⅠand GradeⅡindividuals. With the increase of the scale, the overall correlation between the dominant populations changed from no correlation to negative correlation and then to no correlation. The spatial correlation between populations of different diameter classes only appeared among trees of medium and small diameter classes, and the correlation was negative. Conclusion Q. mongolica natural forest in the study area is at the early stage of succession, with a stable dominant population structure and good renewal status. To accelerate the succession process, artificial intervention should appropriately supplemented to promote the development of forest stands towards top-level communities. [Ch, 5 fig. 1 tab. 31 ref.] -
图 3 优势种群不同径级的空间分布格局
Figure 3 Spatial distribution pattern of dominant species at different diameter class
表 1 辽东栎天然林群落各树种的重要值
Table 1. Importance values of tree species in Q. mongolica community
树种 相对多度/% 相对频度/% 相对显著度/% 重要值/% 树种 相对多度/% 相对频度/% 相对显著度/% 重要值/% 辽东栎 69.80 25.81 80.66 58.76 山丁子 3.83 12.90 2.78 6.51 茶条槭 9.67 18.28 5.24 11.06 山杏 5.57 12.90 2.30 6.92 白桦 1.64 7.53 1.09 3.42 油松 1.19 8.60 0.68 3.49 山杨 6.84 7.53 5.88 6.75 鹅耳枥 1.46 6.45 1.37 3.09 说明:山丁子Malus baccata;山杏Prunus sibirica;鹅耳枥Carpinus turczaninowii。  下载: 导出CSV
下载: 导出CSV
-
[1] 邱婧, 韩安霞, 何春梅, 等. 秦岭优势乔木锐齿槲栎的空间分布格局及种内关联[J]. 应用生态学报, 2022, 33(8): 2035−2042. QIU Jing, HAN Anxia, HE Chunmei, et al. Spatial distribution pattern and intraspecific association of dominant species Quercus aliena var. acutiserrata in Qinling Mountains, China[J]. Chinese Journal of Applied Ecology, 2022, 33(8): 2035−2042. [2] 张晓红, 张会儒, 卢军, 等. 长白山蒙古栎次生林群落结构特征及优势树种空间分布格局[J]. 应用生态学报, 2019, 30(5): 1571−1579. ZHANG Xiaohong, ZHANG Huiru, LU Jun, et al. Community structure characteristics and spatial distribution of dominant species of secondary Quercus mongolica forest in Changbai Mountains, China[J]. Chinese Journal of Applied Ecology, 2019, 30(5): 1571−1579. [3] NATHAN R. Long-distance dispersal of plants[J]. Science, 2006, 313(5788): 786−788. [4] HOU J H, MI X C, LIU C R, et al. Spatial patterns and associations in a Quercus-Betula forest in northern China[J]. Journal of Vegetation Science, 2004, 15(3): 407−414. [5] 王静轩, 张佳鑫, 向泽宇, 等. 庐山25 hm2森林样地乔木层优势种空间分布格局及关联性[J]. 应用生态学报, 2023, 34(6): 1491−1499. WANG Jingxuan, ZHANG Jiaxin, XIANG Zeyu, et al. Spatial distribution pattern and correlation of dominant species in the arbor layer at a 25 hm2 forest plot in Lushan Mountain, China[J]. Chinese Journal of Applied Ecology, 2023, 34(6): 1491−1499. [6] 王丽萍, 乌俊杰, 柴勇, 等. 高黎贡山中山湿性常绿阔叶林优势种空间分布格局及其关联性[J]. 植物生态学报, 2024, 48(2): 180−191. WANG Liping, WU Junjie, CHAI Yong, et al. Spatial patterns and associations of dominant species in a subtropical mid-mountain moist evergreen broadleaf forest in Gaoligong Mountains, Southwest China[J]. Chinese Journal of Plant Ecology, 2024, 48(2): 180−191. [7] BEN-SAID M. Spatial point-pattern analysis as a powerful tool in identifying pattern-process relationships in plant ecology: an updated review[J/OL]. Ecological Processes, 2021, 10(1): 56[2024-08-10]. DOI: 10.1186/s13717-021-00314-4. [8] 伊力塔. 山西灵空山辽东栎林群落特征研究[D]. 北京: 北京林业大学, 2008. YI Lita. Community Characteristics of Quercus liaotungensis Koidz. in the Lingkong Mountain in Shanxi Province, China[D]. Beijing: Beijing Forestry University, 2008. [9] 宁鹏. 中条山优势树种包含气候因子的生长模型构建与动态预测[D]. 太谷: 山西农业大学, 2023. NING Peng. Study on the Growth Models Including Climate Dactors and Dynamic Prediction of Dominant Tree Species in Zhongtiao Mountains[D]. Taigu: Shanxi Agricultural University, 2023. [10] 任达. 采伐强度对辽东栎林分生长和林下物种多样性的短期影响[J]. 山西林业科技, 2021, 50(4): 14−17. REN Da. Short term effects of cutting intensity on stand growth and understory species diversity of Quercus Liaotungensis[J]. Shanxi Forestry Science and Technology, 2021, 50(4): 14−17. [11] 刘铭波, 韩海荣, 程小琴, 等. 山西灵空山典型天然林空间结构特征及其对生产力的影响[J]. 西北林学院学报, 2022, 37(1): 33−40. LIU Mingbo, HAN Hairong, CHENG Xiaoqin, et al. Spatial structure characteristics and their influences on productivity of typical natural forests in the Lingkong Mountain, Shanxi Province[J]. Journal of Northwest Forestry University, 2022, 37(1): 33−40. [12] 王晶, 毕润成, 张钦弟, 等. 山西太岳山辽东栎群落优势种生态位研究[J]. 生态科学, 2016, 35(4): 62−70. WANG Jing, BI Runcheng, ZHANG Qindi, et al. Niche characteristics of dominant plant species in a Quercus wutaishanica community in Taiyue Mountain Shanxi Province[J]. Ecological Science, 2016, 35(4): 62−70. [13] 张中惠, 郭建斌, 王彦辉, 等. 六盘山辽东栎林种群结构和空间分布格局[J]. 浙江农林大学学报, 2021, 38(6): 1091−1099. ZHANG Zhonghui, GUO Jianbin, WANG Yanhui, et al. Populaion structure and spatial distribution pattern of Quercus wutaishanica in Liupan Mountains[J]. Journal of Zhejiang A&F University, 2021, 38(6): 1091−1099. [14] 王艳, 毕润成, 许强. 山西太岳山南部针阔混交林群落特征及空间分布格局[J]. 广西植物, 2017, 37(7): 901−911. WANG Yan, BI Runcheng, XU Qiang. Community characteristics and spatial distribution of mixed conifer and broad-leaved forest in south Taiyue Mountain, Shanxi Province of China[J]. Guihaia, 2017, 37(7): 901−911. [15] 张金屯, 孟东平. 芦芽山油松-辽东栎林优势树种空间分布格局研究[J]. 西北植物学报, 2006, 26(8): 1682−1685. ZHANG Jintun, MENG Dongping. Spatial distribution patterns of dominant tree species in Pinus tabulaeformis-Quercus liaotungensis forest in Luyashan Mountain, China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2006, 26(8): 1682−1685. [16] 胡尔查, 王晓江, 张文军, 等. 乌拉山自然保护区白桦种群的年龄结构和点格局分析[J]. 生态学报, 2013, 33(9): 2867−2876. HU Ercha, WANG Xiaojiang, ZHANG Wenjun, et al. Age structure and point pattern of Butula platyphylla in Wulashan Natural Reserve of Inner Mongolia[J]. Acta Ecologica Sinica, 2013, 33(9): 2867−2876. [17] 涂洪润, 李姣凤, 刘润红, 等. 桂林岩溶石山檵木种群空间格局及其关联性[J]. 应用生态学报, 2019, 30(8): 2621−2630. TU Hongrun, LI Jiaofeng, LIU Runhong, et al. Spatial distribution patterns and association of Loropetalum chinense population in karst hills of Guilin, Southwest China[J]. Chinese Journal of Applied Ecology, 2019, 30(8): 2621−2630. [18] 曹羚, 金晟康, 叶尔江·拜克吐尔汉, 等. 额敏县新疆野苹果种群空间分布格局及其关联性[J]. 浙江农林大学学报, 2023, 40(2): 390−397. CAO Ling, JIN Shengkang, Yeerjiang Baiketuerhan, et al. Spatial pattern and correlation of Malus sieversii population in Emin County[J]. Journal of Zhejiang A&F University, 2023, 40(2): 390−397. [19] 金亚宁, 管增艳, 石松林, 等. 川西云杉人工林与天然林群落空间分布格局及种间关联性[J]. 浙江农林大学学报, 2022, 39(3): 495−504. JIN Yaning, GUAN Zengyan, SHI Songlin, et al. Spatial distribution pattern and interspecific correlation between plantation and natural forest of Picea likiangensis var. rubescens in western Sichuan, China[J]. Journal of Zhejiang A&F University, 2022, 39(3): 495−504. [20] 董灵波, 马榕, 田栋元, 等. 大兴安岭天然林不同演替阶段共优势种种群结构与动态[J]. 应用生态学报, 2022, 33(8): 2077−2087. DONG Lingbo, MA Rong, TIAN Dongyuan, et al. Structure and dynamics of co-dominant species in different succession stages of natural forests in Daxing’an Mountains, China[J]. Chinese Journal of Applied Ecology, 2022, 33(8): 2077−2087. [21] 董丽, 郭东罡, 段毅豪, 等. 灵空山辽东栎-油松林更新空间分布格局及其与地形因子的关系[J]. 应用与环境生物学报, 2013, 19(6): 914−921. DONG Li, GUO Donggang, DUAN Yihao, et al. Spatial distribution patterns of regenerating Quercus wutaishanica-Pinus tabulaeformis in relation to topographic factors in the Lingkong Mountain[J]. Chinese Journal of Applied and Environmental Biology, 2013, 19(6): 914−921. [22] 王祎玲, 张钦弟, 郝晓杰, 等. 山西七里峪茶条槭的种群结构与空间分布格局[J]. 西北植物学报, 2012, 32(5): 1027−1035. WANG Yiling, ZHANG Qindi, HAO Xiaojie, et al. Structure and spatial distribution of Acer ginnala population in Qiliyu, Shanxi[J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(5): 1027−1035. [23] 冯云, 马克明, 张育新, 等. 北京东灵山地区辽东栎(Quercus liaotungensis)种群生活史特征与空间分布[J]. 生态学杂志, 2009, 28(8): 1443−1448. FENG Yun, MA Keming, ZHANG Yuxin, et al. Life history characteristics and spatial pattern of Quercus liaotungensis population in Dongling Mountain of Beijing, China[J]. Chinese Journal of Ecology, 2009, 28(8): 1443−1448. [24] 黄维, 薛卫鹏, 陈周娟, 等. 油松飞播林林分空间结构特征分析与评价[J]. 森林与环境学报, 2022, 42(4): 425−433. HUANG Wei, XUE Weipeng, CHEN Zhoujuan, et al. Analysis and evaluation of stand spatial structure characteristics of Pinus tabuliformis aerial seeding forest[J]. Journal of Forest and Environment, 2022, 42(4): 425−433. [25] 王志鸣, 郭秋菊, 艾训儒, 等. 恩施州不同海拔马尾松天然次生林林分结构特征研究[J]. 西南林业大学学报(自然科学), 2019, 39(1): 114−122. WANG Zhiming, GUO Qiuju, AI Xunru, et al. Structure characteristics of Pinus massoniana natural secondary forest at different altitude of Enshi[J]. Journal of Southwest Forest University (Natural Science), 2019, 39(1): 114−122. [26] 杜华强, 汤孟平, 周国模, 等. 天目山物种多样性尺度依赖及其与空间格局关系的多重分形[J]. 生态学报, 2007, 27(12): 5038−5049. DU Huaqiang, TANG Mengping, ZHOU Guomo, et al. Linking species diversity with spatial scale dependence to spatial patterns using multifractal analysis[J]. Acta Ecologica Sinica, 2007, 27(12): 5038−5049. [27] 解传奇, 田民霞, 赵忠瑞, 等. 西藏色季拉山急尖长苞冷杉种群点格局分析[J]. 应用生态学报, 2015, 26(6): 1617−1624. XIE Chuanqi, TIAN Minxia, ZHAO Zhongrui, et al. Spatial point pattern analysis of Abies georgei var. smithii in forest of Sygera Mountains in southeast Tibet, China[J]. Chinese Journal of Applied Ecology, 2015, 26(6): 1617−1624. [28] 兰航宇, 段文标, 陈立新, 等. 小兴安岭天然针阔混交林主要树种空间格局及其关联性[J]. 生态学报, 2019, 39(18): 6660−6669. LAN Hangyu, DUAN Wenbiao, CHEN Lixin, et al. Spatial point patterns and associations of populations in the coniferous and deciduous broadleaved mixed forest in Xiaoxing’an Mountain[J]. Acta Ecologica Sinica, 2019, 39(18): 6660−6669. [29] BROOKER R W, MAESTRE F T, CALLAWAY R M, et al. Facilitation in plant communities: the past, the present, and the future[J]. Journal of Ecology, 2008, 96(1): 18−34. [30] 陈列, 赵秀海, 张赟. 长白山北坡椴树红松林空间分布及其空间关联[J]. 北京林业大学学报, 2009, 31(3): 6−10. CHEN Lie, ZHAO Xiuhai, ZHANG Yun. Spatial distribution patterns and associations of Pinus koraiensis and Tilia amurensis in Tilia-Korean pine forest on the north slope of Changbai Mountain, Northeastern China[J]. Journal of Beijing Forestry University, 2009, 31(3): 6−10. [31] 邹书珍, 尹才佳, 杨茜, 等. 子午岭次生油松林立木竞争格局研究[J]. 植物研究, 2023, 43(1): 140−149. ZOU Shuzhen, YIN Caijia, YANG Qian, et al. Competition pattern of standing trees in secondary Pinus forest in the Ziwuling Mountains, China[J]. Bulletin of Botanical Research, 2023, 43(1): 140−149. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240535
 点击查看大图
点击查看大图
图(5) / 表(1)
计量
- 文章访问数: 1141
- HTML全文浏览量: 397
- PDF下载量: 45
- 被引次数: 0
