







-
瓯柑Citrus suavissima优良新品种‘无子瓯柑’C. suavissima ‘Seedless’是通过芽变选种获得的,其果实无核,遗传性状稳定[1]。与瓯柑相比,两者的花粉量相当,但‘无子瓯柑’花药不易开裂,自然散发的花粉少[2]。形态学和细胞学研究表明:小孢子母细胞减数分裂异常是‘无子瓯柑’雄性不育的重要原因之一[2−3]。瓯柑及‘无子瓯柑’花粉发育早期的花药转录组与蛋白质组关联分析结果表明:差异代谢通路主要富集在苯丙素生物合成、黄酮类化合物生物合成和苯丙氨酸代谢通路[4]。CsRNF217基因是瓯柑与‘无子瓯柑’在小孢子母细胞时期与苯丙素生物合成途径密切关联的重要基因,与瓯柑相比,该基因在同时期的‘无子瓯柑’中显著上调表达;CsRNF217基因的氨基酸序列中存在RING-HC_RBR和IBR 2个结构域,属于典型的单亚基RING-HC E3亚家族,定位于细胞核[5]。
泛素化是真核生物中一种高度通用的翻译后修饰,它介导蛋白酶体降解、细胞运输、蛋白质相互作用和细胞蛋白质的功能激活[6−8]。泛素化级联需要3种不同的酶催化:E1泛素激活酶、E2泛素结合酶和E3泛素连接酶[9]。其中,E3泛素连接酶具有显著的多样性,它决定了底物的特异性,并作为调节细胞反应的枢纽[10−11]。E3泛素连接酶分为单亚基和多亚基两类。单亚基组由3个主要的亚家族组成:RING、HECT以及U-box结构域家族[11−13]。CsRNF217基因即属于单亚基RING结构域家族。E3泛素连接酶在植物的生长发育过程中起着关键的作用,包括细胞程序性死亡和抗病防御反应[14]、成花调控[15]、生物[16]和非生物应激反应[14]以及花粉发育[17−19]等。其中,在水稻Oryza sativa[17]、拟南芥Arabidopsis thaliana[18]、白菜Brassica campestris ssp. chinensis[19]中发现的RING型E3泛素连接酶基因DSNP1、DAF、Bra015092等在花粉发育过程中起重要作用。
本研究以过表达‘无子瓯柑’CsRNF217的转基因烟草Nicotiana tabacum植株为材料,通过对基因表达分析、转基因自交1代植株(T1)花粉活力、转基因植株自交及与野生型烟草正反交的结实率等参数的测定,分析过表达CsRNF217对转基因烟草植株育性的影响,为进一步揭示CsRNF217基因在‘无子瓯柑’雌雄败育过程中的重要作用提供参考。
-
以野生型烟草(WT)作为对照,采用农杆菌Agrobacterium tumefaciens介导法获得过表达CaMV 35S :: CsRNF217的阳性转基因烟草[5](T0)。将T0自交种子穴盘播种获得转基因烟草自交1代植株(T1)。采集T1阳性烟草植株6#、35#、63#株系的叶片及含苞待放花蕾的花药,立即置于液氮中速冻并于−80 ℃保存,用于半定量RT-PCR分析。
-
采集野生型烟草及T1烟草植株的叶片,采用CTAB法提取DNA,以pCAMBIA 2300s : CsRNF217质粒作为阳性对照,ddH2O为阴性对照,用35S_F及基因特异引物CsRNF217_R838_S[5](表1)进行PCR检测筛选阳性植株,PCR程序为94 ℃,5 min;94 ℃,30 s,55 ℃,30 s,72 ℃,1 min,35次循环;72 ℃,10 min;12 ℃保存。
表 1 引物信息
Table 1. Specific primers used in this study
引物名称 引物序列(5′→3′) CsRNF217_R838_S GTCGACTTGCATAGAGCCAATAAA 35S_F ACGCACAATCCCACTATCCTTC CsRNF217_qL2 ACGTGCGAGGGTATGAAAGA CsRNF217_qR2 TACCCTCCATGCCACTTCAG EF1α-F TGGTTGTGACTTTTGGTCCCA EF1α-R ACAAACCCACGCTTGAGATCC -
于晴天10:00采集烟草含苞待放的花蕾,采用亚历山大染色法[20]对野生型烟草及经PCR鉴定的T1阳性植株进行花粉染色活力检测,采用花粉离体培养法[21]对花粉染色活力显著下降的烟草单株进行花粉萌发试验,每个植株随机挑选3朵花作为生物学重复。花粉粒被染为紫红色的视为有活力的花粉粒,花粉管长度大于或等于花粉粒直径视为萌发。花粉染色活力=着色花粉粒数/视野中的花粉粒总数×100%,花粉萌发率=萌发花粉粒数/视野中的花粉粒总数×100%。
使用MiniBEST Plant RNA Extraction Kit (TaKaRa,日本)试剂盒,提取野生型烟草及花粉活力显著下降的T1烟草阳性植株花药RNA,采用EASYScript one-step gDNA removal and cDNA synthesis supermix (TransGen Biotech code#AE311- 03)试剂盒进行cDNA合成,使用CsRNF217基因对引物和烟草内参基因EF1α特异引物(表1)进行半定量RT-PCR分析,程序为94 ℃,5 min;94 ℃,30 s,55 ℃,30 s,72 ℃,1 min,29次循环;72 ℃,10 min;12 ℃保存。
-
于晴天10:00收集野生型和T1已开花,但未散粉植株的花药于离心管,4 ℃干燥保存备用。于傍晚用干净的小镊子摘去即将开放花蕾的花瓣和雄蕊。次日10:00蘸取花粉轻点已分泌黏液的柱头,套袋,每个植株进行3朵花的正反交重复,3 d后摘去袋子。正反交组合配置中,正交授粉以转基因株系为父本,野生型为母本;反交授粉以野生型为父本,转基因株系为母本。种子成熟时统计每个单株采收的蒴果数,测量蒴果横径、纵径及种子总质量。
-
采用SPSS软件对种子萌发率、花粉染色活力、花粉离体萌发率、蒴果横纵径及种子数量等进行了单因素方差分析(one-way ANOVA,LSD)。
-
T0烟草种子穴盘播种共获得137个单株,其中6#株系16株,35#株系80株,63#株系41株。以野生型及T0种子播种获得的烟草叶片DNA为模板,经PCR检测,共获得68株阳性植株(表2)。其中,6#株系10株,35#株系33株,63#株系25株,阳性率分别为62.5%、41.2%、61.5%。
表 2 野生型及T1烟草植株阳性率与花粉染色活力
Table 2. Positive rate and pollen viability of wild type and T1 tobacco plants
株系 株数/
株阳性
株数/株植株阳
性率/%花粉染色活力
均值/%6# 16 10 62.5 19.5±2.7 b 35# 80 33 41.2 31.9±2.8 b 63# 41 25 61.5 21.8±2.3 b WT 3 0 0 94.5±2.4 a 说明:同列不同字母表示不同株系间花粉染色活力差异显著(P<0.05)。WT为野生型株系。 -
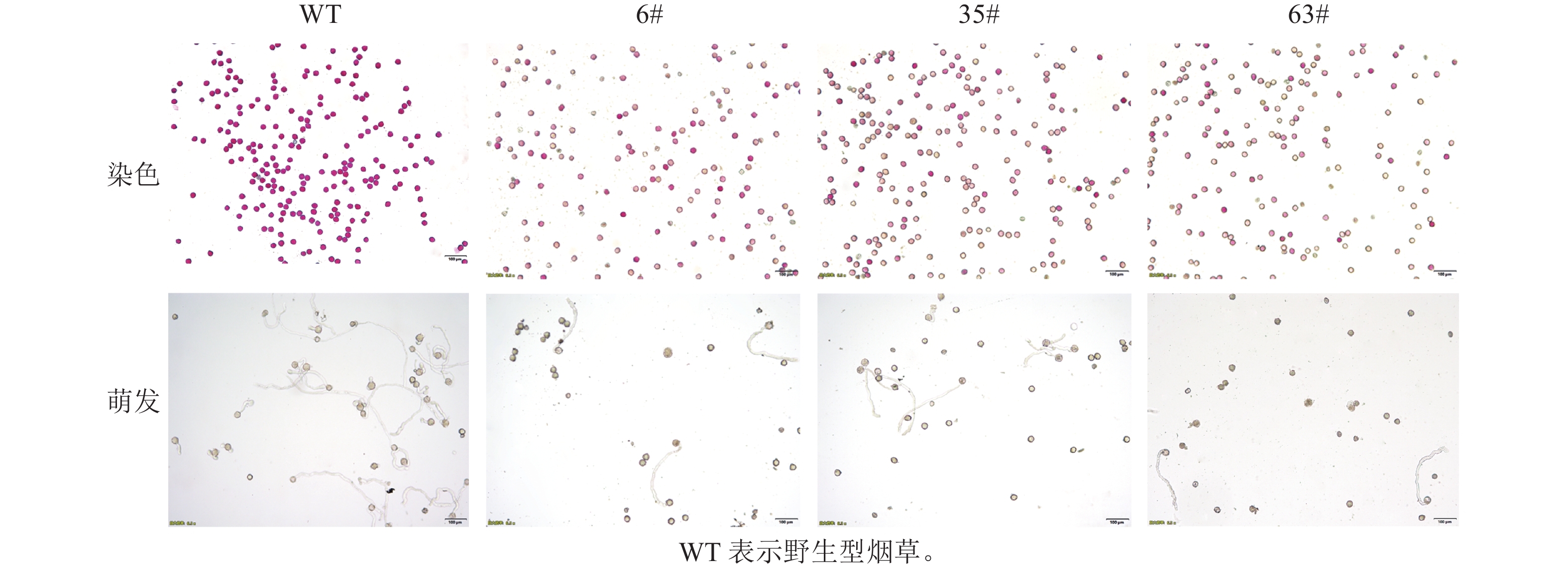
T1阳性烟草植株花粉的散粉量与野生型存在差异(图1)。野生型植株花药开裂后,散粉量多,可见大量花粉散布于花瓣及柱头;T1阳性烟草植株花药裂开后,散粉量少,几乎无可见花粉散出。经染色法和花粉离体萌发培养(图2):T1阳性烟草植株花粉染色活力显著低于野生型植株(图3,P<0.05)。有活力的花粉粒经亚历山大染色后呈紫红色,无活力的花粉粒呈黄褐色。野生型植株经亚历山大染色后花粉粒大多呈紫红色(94.5%),阳性植株呈紫红色的花粉粒数量显著少于野生型(图3,P<0.05)。其中6#株系的14号单株(6#14)、35#株系的4号单株(35#4)、63#株系的4号单株(63#4)的花粉经亚历山大染色后着色率最低,分别为9.6%、12.0%、9.7%。有活力的花粉粒能在适宜的离体条件下萌发,T1阳性植株花粉粒的萌发率显著低于野生型植株(60.3%)。其中6#株系的9号单株(6#9)、6#株系的14号单株(6#14)、63#株系的4号单株(63#4)的花粉粒萌发率在各株系中最低,分别为30.6%、29.0%、33.4%。半定量RT-PCR分析显示(图4):CsRNF217基因在过表达的各株系中均能表达,其中,外源基因在63#株系各单株花药中的表达量最高。

图 1 野生型与T1阳性植株成熟花药形态及散粉情况
Figure 1. Anther morphology and pollen release of wild type and T1 positive plants

图 2 野生型及T1阳性植株的花粉活力
Figure 2. Pollen viability of wild type and T1 positive plants

图 3 野生型及T1阳性植株的花粉活力
Figure 3. Pollen vitality of wild-type and T1 positive plants

图 4 野生型及T1阳性植株半定量RT-PCR分析
Figure 4. Semi-quantitative RT-PCR of wild type and T1 positive plants
-
对花粉染色活力、萌发率都显著低于野生型的T1转基因阳性单株进行授粉(表3,图5)表明:自交和与野生型进行正反交的各授粉组合均能结实,但蒴果大小及种子数量存在较大差异。蒴果横径的比较结果显示:转基因63#株系自交、与野生型进行正反交的蒴果横径均显著小于野生型自交的蒴果(P<0.05);35#株系自交蒴果的横径显著小于野生型自交(P<0.05)。蒴果纵径的比较结果显示:转基因63#株系自交和与野生型反交的蒴果纵径均显著小于野生型自交(P<0.05),其余组合与野生型自交无显著差异。根据烟草原始种的种子千粒重(0.087 g)[22],计算每个株系的单果种子数量。阳性株系自交、与野生型正反交获得的种子数量都显著低于野生型自交种子数(P<0.05),但转基因株系之间无显著差异。
表 3 野生型及阳性株系自交及杂交试验的蒴果大小和种子数量
Table 3. Capsule and seed number in selfing and hybridization test
处理 株系 蒴果横径/mm 蒴果纵径/mm 种子数量/(粒·果−1) 自交 WT 9.2±0.2 a 15.8±0.2 a 1 525.7±19.9 a 6# 8.9±0.1 ab 14.9±0.1 ab 1 030.4±105.2 b 35# 8.4±0.1 bcd 14.9±0.1 ab 948.5±37.3 b 63# 8.4±0.0 bcd 14.6±0.1 b 1 006.7±36.0 b 正交 WT (♀) × 6# (♂) 8.5±0.1 abcd 15.2±0.4 ab 845.5±69.4 b WT (♀) × 35# (♂) 8.6±0.2 abc 14.3±0.2 b 924.6±26.7 b WT (♀) × 63# (♂) 8.1±0.2 cd 15.0±0.4 ab 1 049.8±8.0 b 反交 6# (♀) × WT (♂) 8.6±0.2 abc 15.1±0.5 ab 1 103.4±119.8 b 35# (♀) × WT (♂) 9.1±0.3 ab 14.8±0.5 ab 992.3±185.9 b 63# (♀) × WT (♂) 7.8±0.3 d 14.4±0.4 b 914.4±90.7 b 说明:WT表示野生型烟草。同列不同字母表示株系间差异显著(P<0.05)。 
图 5 野生型与T1阳性植株自交及与野生型正反交授粉后的蒴果及种子
Figure 5. Capsules and seeds of wild type and T1 positive plants after self-crossing and cross-pollination with wild type
-
本研究表明:过表达CsRNF217的烟草植株在花粉染色活力、花粉萌发率、蒴果大小及种子数量上均显著低于野生型,说明过表达CsRNF217降低了转基因烟草的小孢子育性,甚至对胚囊育性也存在一定的影响。
近年来的研究表明:RING型E3泛素连接酶基因参与了植物花粉发育[19]、花药开裂[18]、胚囊发育[17]等生命过程。水稻中的RING型E3泛素连接酶DSNP1与水稻减数分裂过程中的联会复合体组装和同源重组关系密切,其突变体dsnp1中形成的稳定同源配对和重组交叉严重减少,并最终形成活性较低的花粉[17]。然而,白菜中过表达E3泛素连接酶基因Bra015092,也降低了转基因白菜的花粉染色活力和萌发率,并导致了花粉外部形态的畸形[19]。前期研究表明:‘无子瓯柑’成熟花粉的染色活力和离体萌发率均显著低于有籽瓯柑,其小孢子母细胞减数分裂异常[2−3],CsRNF217的表达在小孢子发育早期显著上调[5]。本研究中过表达CsRNF217的烟草花粉离体萌发率为野生型的一半,花粉染色活力则更低。推测CsRNF217基因可能通过负调控小孢子母细胞的减数分裂过程参与‘无子瓯柑’的花粉发育。拟南芥E3泛素连接酶基因DAF在雄蕊中特异表达,并通过正向调控茉莉酸生物合成途径来促进花药开裂,通过干涉实验抑制其表达的植株则表现为雄性不育[18]。‘无子瓯柑’的花药自然开裂难、散粉量低[2−3],实时定量PCR结果显示:CsRNF217基因在雄蕊的表达丰度显著高于其他花器官[5]。本研究中,过表达CsRNF217的转基因烟草也表现出自然散粉较弱的特性,推测CsRNF217可能参与了花药开裂的调控。
以野生型花粉对水稻不育突变体dsnp1进行授粉,突变体不能结实,表明该突变体既是雄性不育的,也表现出雌性不育[17],说明该RING型E3泛素连接酶DSNP1对水稻的雌雄育性都有影响。本研究表明:转基因烟草不仅花粉活力显著下降,其种子数量也显著减少,说明CsRNF217的超量表达在负调控花粉育性的同时,对胚囊的育性也存在一定的调控作用。在生产实践中,‘无子瓯柑’在同其他有籽柑橘品种混栽时也表现无籽,说明胚囊败育是‘无子瓯柑’果实无核的重要原因,因此CsRNF217对‘无子瓯柑’胚囊育性的影响值得进一步研究。
-
E3泛素连接酶基因CsRNF217对转基因烟草的育性存在显著影响,过表达‘无子瓯柑’CsRNF217的T1烟草阳性植株雌雄育性均显著下降,推测CsRNF217对‘无子瓯柑’的育性存在负调控作用。
Fertility effect of E3 ubiquitin ligase gene CsRNF217 in transgenic tobacco plants
-
摘要:
目的 为了验证‘无子瓯柑’Citrus suavissima ‘Seedless’ E3泛素连接酶基因CsRNF217对雄蕊育性的影响,采用异源转化获得过表达CsRNF217的转基因烟草Nicotiana tabacum,分析其对烟草育性的影响。 方法 采用亚历山大染色法、花粉离体培养法、杂交授粉后的结实率分析野生型烟草及转基因烟草自交1代(T1)阳性株系的花粉活力及胚囊育性,利用半定量RT-PCR分析基因CsRNF217在转基因烟草T1阳性株系花药中的表达强弱。 结果 ‘无子瓯柑’CsRNF217在转基因烟草自交T1株系中高效表达,转基因株系的花粉染色活力和离体萌发率、自交结实率、以及与野生型的正反交结实率均显著低于野生型(P<0.05)。 结论 过表达CsRNF217的烟草植株花粉育性显著下降,同时伴随胚囊育性下降的现象,推测CsRNF217基因对‘无子瓯柑’雌雄育性存在负向调控的作用。图5表3参22 Abstract:Objective This study, with an investigation into the transgenic plants of tobacco (Nicotiana tabacum) overexpressing CsRNF217, is aimed to verify the effect of the E3 ubiquitin ligase gene CsRNF217 of Citrus suavissima ‘Seedless’ on male sterility. Method First, an analysis was conducted of the pollen viability of wild type (WT) and the positive plants of the 1st generation of selfing of transgenic plants (T1) with the employment Alexander staining and pollen germination in vitro. Then, the seed setting rate was obtained via selfing and reciprocal crosses between transgenic plants and WT. Finally, semi-quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR) was used to analyze the relative expression of CsRNF217 in T1 positive tobacco plants. Result CsRNF217 derived from C. suavissima ‘Seedless’ was efficiently expressed in T1 lines. The pollen dyeing viability and in vitro germination rate of T1 lines, selfing seed setting rate of T1 lines, and seed setting rate of T1 lines reciprocal crosses with WT were significantly lower than those of WT (P<0.05). Conclusion T1 plants overexpressing CsRNF217 had a severe decline in pollen fertility and partial aberrant of embryo sac, suggesting that an up-regulation of CsRNF217 could play a negative regulatory role on male and female fertility of C. suavissima ‘Seedless’. [Ch, 5 fig. 3 tab. 22 ref.] -
Key words:
- Citrus /
- E3 ubiquitin ligase /
- male sterility /
- embryo sac abortion
-
图 1 野生型与T1阳性植株成熟花药形态及散粉情况
Figure 1 Anther morphology and pollen release of wild type and T1 positive plants
图 4 野生型及T1阳性植株半定量RT-PCR分析
Figure 4 Semi-quantitative RT-PCR of wild type and T1 positive plants
图 5 野生型与T1阳性植株自交及与野生型正反交授粉后的蒴果及种子
Figure 5 Capsules and seeds of wild type and T1 positive plants after self-crossing and cross-pollination with wild type
表 1 引物信息
Table 1. Specific primers used in this study
引物名称 引物序列(5′→3′) CsRNF217_R838_S GTCGACTTGCATAGAGCCAATAAA 35S_F ACGCACAATCCCACTATCCTTC CsRNF217_qL2 ACGTGCGAGGGTATGAAAGA CsRNF217_qR2 TACCCTCCATGCCACTTCAG EF1α-F TGGTTGTGACTTTTGGTCCCA EF1α-R ACAAACCCACGCTTGAGATCC  下载: 导出CSV
下载: 导出CSV
表 2 野生型及T1烟草植株阳性率与花粉染色活力
Table 2. Positive rate and pollen viability of wild type and T1 tobacco plants
株系 株数/
株阳性
株数/株植株阳
性率/%花粉染色活力
均值/%6# 16 10 62.5 19.5±2.7 b 35# 80 33 41.2 31.9±2.8 b 63# 41 25 61.5 21.8±2.3 b WT 3 0 0 94.5±2.4 a 说明:同列不同字母表示不同株系间花粉染色活力差异显著(P<0.05)。WT为野生型株系。
下载: 导出CSV
表 3 野生型及阳性株系自交及杂交试验的蒴果大小和种子数量
Table 3. Capsule and seed number in selfing and hybridization test
处理 株系 蒴果横径/mm 蒴果纵径/mm 种子数量/(粒·果−1) 自交 WT 9.2±0.2 a 15.8±0.2 a 1 525.7±19.9 a 6# 8.9±0.1 ab 14.9±0.1 ab 1 030.4±105.2 b 35# 8.4±0.1 bcd 14.9±0.1 ab 948.5±37.3 b 63# 8.4±0.0 bcd 14.6±0.1 b 1 006.7±36.0 b 正交 WT (♀) × 6# (♂) 8.5±0.1 abcd 15.2±0.4 ab 845.5±69.4 b WT (♀) × 35# (♂) 8.6±0.2 abc 14.3±0.2 b 924.6±26.7 b WT (♀) × 63# (♂) 8.1±0.2 cd 15.0±0.4 ab 1 049.8±8.0 b 反交 6# (♀) × WT (♂) 8.6±0.2 abc 15.1±0.5 ab 1 103.4±119.8 b 35# (♀) × WT (♂) 9.1±0.3 ab 14.8±0.5 ab 992.3±185.9 b 63# (♀) × WT (♂) 7.8±0.3 d 14.4±0.4 b 914.4±90.7 b 说明:WT表示野生型烟草。同列不同字母表示株系间差异显著(P<0.05)。
下载: 导出CSV
-
[1] 徐象华, 颜福花, 叶荣华, 等. 瓯柑研究进展[J]. 浙江林业科技, 2008, 28(3): 75 − 77. XU Xianghua, YAN Fuhua, YE Ronghua, et al. Advances in researches of Citrus reticulata cv. suavissima [J]. Journal of Zhejiang Forestry Science and Technology, 2008, 28(3): 75 − 77. [2] 曾燕如, 张炼炳, 斯金平, 等. 无籽瓯柑无核原因的研究[J]. 浙江林学院学报, 2005, 22(4): 359 − 362. ZENG Yanru, ZHANG Lianbing, SI Jinping, et al. Preliminary study on seedlessness of seedless Ougan [J]. Journal of Zhejiang Forestry College, 2005, 22(4): 359 − 362. [3] 张迟, 张敏, 朱铨, 等. ‘瓯柑’及其无子突变体花粉发育的细胞学观察[J]. 果树学报, 2014, 31(2): 265 − 269. ZHANG Chi, ZHANG Min, ZHU Quan, et al. Cytological observation of pollen development in ‘Ougan’ (Citrus suavissima Hort. ex Tanaka) and its seedless mutant [J]. Journal of Fruit Science, 2014, 31(2): 265 − 269. [4] ZHANG Chi, YU Dihu, KE Fuzhi, et al. Seedless mutant ‘Wuzi Ougan’ (Citrus suavissima Hort. ex Tanaka ‘seedless’) and the wild type were compared by iTRAQ-based quantitative proteomics and integratedly analyzed with transcriptome to improve understanding of male sterility [J/OL]. BMC Genetics, 2018, 19(1): 106[2022-11-10]. doi: 10.1186/s12863-018-0693-9. [5] 孙明艳, 叶潇铃, 蔡子涵, 等. 无子瓯柑E3泛素连接酶基因CsRNF217的克隆与功能分析[J]. 果树学报, 2022, 39(2): 845 − 855. SUN Mingyan, YE Xiaoling, CAI Zihan, et al. Cloning and functional analysis of E3 ubiquitin ligase gene CsRNF217 in Wuzi Ougan (Citrus suavissima) [J]. Journal of Fruit Science, 2022, 39(2): 845 − 855. [6] FOOT N, HENSHALL T, KUMAR S. Ubiquitination and the regulation of membrane proteins [J]. Physiological Reviews, 2017, 97(1): 253 − 281. [7] SASAKI A T, CARRACEDO A, LOCASALE J W, et al. Ubiquitination of K-Ras enhances activation and facilitates binding to select downstream effectors [J/OL]. Science Signaling, 2011, 4(163): ra13[2022-11-10]. doi: 10.1126/scisignal.2001518. [8] SHARMA B, JOSHI D, YADAV P K, et al. Role of ubiquitin-mediated degradation system in plant biology [J/OL]. Frontiers in Plant Science, 2016, 7: 806[2022-11-10]. doi: 10.3389/fpls.2016.00806. [9] GUERRA D D, CALLIS J. Ubiquitin on the move: the ubiquitin modification system plays diverse roles in the regulation of endoplasmic reticulum- and plasma membrane-localized proteins [J]. Plant Physiology, 2012, 160(1): 56 − 64. [10] BUETOW L, HUANG D T. Structural insights into the catalysis and regulation of E3 ubiquitin ligases [J]. Nature Reviews Molecular Cell Biology, 2016, 17(10): 626 − 642. [11] SHU Kai, YANG Wenyu. E3 ubiquitin ligases: ubiquitous actors in plant development and abiotic stress responses [J]. Plant and Cell Physiology, 2017, 58(9): 1461 − 1476. [12] DESHAIES R J, JOAZEIRO C A. RING domain E3 ubiquitin ligases [J]. Annual Review of Biochemistry, 2009, 78: 399 − 434. [13] MARÍN I. Evolution of plant HECT ubiquitin ligases [J/OL]. PLoS One, 2013, 8(7): e68536[2022-11-10]. doi: 10.1371/journal.pone.0068536. [14] SHIRSEKAR G S, VEGA-SANCHEZ M E, BORDEOS A, et al. Identification and characterization of suppressor mutants of spl11-mediated cell death in rice [J]. Molecular Plant-microbe Interactions, 2014, 27(6): 528 − 536. [15] LU Dongping, LIN Weiwei, GAO Xiquan, et al. Direct ubiquitination of pattern recognition receptor FLS2 attenuates plant innate immunity [J]. Science, 2011, 332(6036): 1439 − 1442. [16] LUO Hongli, LALUK K, LAI Zhibin, et al. The arabidopsis botrytis susceptible1 interactor defines a subclass of RING E3 ligases that regulate pathogen and stress responses [J]. Plant Physiology, 2010, 154(4): 1766 − 1782. [17] REN Lijun, ZHAO Tingting, ZHAO Yangzi, et al. The E3 ubiquitin ligase DESYNAPSIS1 regulates synapsis and recombination in rice meiosis [J/OL]. Cell Reports, 2021, 37(5): 109941[2022-11-10]. doi: 10.1016/j.celrep.2021.109941. [18] PENG Yanjhu, SHIH Chingfang, YANG Junyi, et al. A RING-type E3 ligase controls anther dehiscence by activating the jasmonate biosynthetic pathway gene DEFECTIVE IN ANTHER DEHISCENCE1 in Arabidopsis [J]. The Plant Journal, 2013, 74(2): 310 − 327. [19] TIAN Aimei, YU Hui, CUI Zhuoyue. Functional characterization of E3 ubiquity ligase Bra015092 in pollen development of Brassica campestris ssp. Chinensis [J/OL]. Physiologia Plantarum, 2022, 174(6): e13808[2022-11-10]. doi: 10.1111/ppl.13808. [20] PETERSON R, SLOVIN J P, CHEN Changbin. A simplified method for differential staining of aborted and nonaborted pollen grains [J/OL]. International Journal of Plant Biology, 2010, 1(2): e13[2022-11-10]. doi: 10.4081/pb.2010.e13. [21] 牛永志, 陈志芸, 索文龙, 等. 不同培养基组分对烟草花粉管生长的影响[J]. 云南农业大学学报, 2020, 35(1): 82 − 87. NIU Yongzhi, CHEN Zhiyun, SUO Wenlong, et al. The influence of medium composition on the growth of tobacco pollen tube [J]. Journal of Yunnan Agricultural University, 2020, 35(1): 82 − 87. [22] 李佛琳, 马宇雷, 张国锋. 烟草种子含钾量和千粒重的基因型差异[J]. 种子, 2000(6): 18 − 21. LI Folin, MA Yulei, ZHANG Guofeng. Genotypic differences in potassium contents and weight of tobacco seeds [J]. Seed, 2000(6): 18 − 21. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220710
 点击查看大图
点击查看大图

计量
- 文章访问数: 1382
- HTML全文浏览量: 567
- PDF下载量: 161
- 被引次数: 0
