





-
“景观”和“游憩”是城市森林的重要功能[1-3]。发挥城市森林“景观”功能的主要载体是林冠的“绿量”,而“游憩”功能则通过建立林下空间来实现。国家林业和草原局于2019年提出GB T37342−2019《国家森林城市评价指标》,旨在建立城乡一体化森林生态系统建设考核指标,并形成标准进行约束[4]。其中,涉及森林“绿量”的评价指标有市域森林覆盖率、新造林面积、城区绿化覆盖率、城区人均公园绿地面积、城区街道绿化(树冠覆盖率)、城区地面停车场绿化(乔木树冠覆盖率)等,用以表现城市总体绿化数量,但在推算城市绿化质量时会出现很大偏差。研究发现:2块面积形状相同、乔木数量和冠幅也相同的绿地,乔木高度不同时,其三维空间层面的绿量有显著差异,从而影响城市森林景观功能的发挥[5]。目前,中国城市森林建设中普遍存在过于讲求增加森林覆盖率的现象,对森林质量的要求相对忽视,树木人为矮化、垂直结构不合理、修剪严重等现象时常出现,城市森林绿量较小。城市森林不同于自然林,它具有一定的区位特殊性[6-8]和功能侧重性[9-13],“游憩”是城市森林的重要功能之一,需要通过构建一定数量的林下空间来实现。林下空间是城市森林空间占有量的有机组成部分,是人们利用最充分的活动场所。城市中人口密集,休闲活动空间有限,且多被硬化场地、草坪等所占据,利用率极低;同时部分林木枝下高过低,通透性差,严重影响了林下空间,限制了林内的游憩行为。本研究以广东省珠海市的城市公园风景林为研究对象,测算城市风景游憩林的林冠三维绿量(3DGQ)和林下游憩空间量(FRS),分析城市森林空间分配策略,从而引导城市森林生态服务功能的高效发挥和健康发展。
-
珠海市地处广东省南部(21°48′~22°27′N,113°03′~114°19′E),珠江出海口西岸,是珠江三角洲中心城市之一、珠江口西岸的核心城市、粤港澳大湾区重要节点城市。属南亚热带季风海洋性气候,气候湿润,年平均气温为22.5 ℃,年平均相对湿度为80%,年平均降水量为2 061.9 mm。选定的风景游憩林均为城市人工营建的常绿阔叶林,其中前山公园位于城市近郊区(CUA),林地面积约3.78 万m2;北理工珠海学院位于城市次核心区(SUA),林地面积约4.40 万m2;梅华城市公园位于城市核心区(UFA),林地面积约8.07 万m2。
-
采用森林群落生态学研究方法,参照周红敏等[14]方法选取样方,在考虑调查时间和成本情况下,设置样方大小为30 m×30 m,每1 万m2抽取4个样方以准确表达森林群落的空间分布格局。其中前山公园风景游憩林群落样地设置调查样方13个,北理工珠海学院14个,梅华城市公园26个。所有样方随机选取,尽可能互不相邻,尽量选择树木种类、配植和体量有一定差异的样方,水面、建筑、纯草地占比面积不超过1/3。所有样方的植被进行每木检尺,记录胸径≥5.0 cm的树种名称,测定胸径、树高、冠幅、树冠净空高度等指标。
-
三维绿量的计算参考郭雪艳等[15],不同树冠形态树种树冠体积计算公式不同。以x表示平均冠径(m),即东西和南北树冠长度的平均值,以y表示冠高(m),通过文献查阅确定不同树种的树冠形态[16-22],分别计算树冠体积(V,m3),确定树冠三维绿量(表1)。
表 1 树冠形态分类及树冠体积
Table 1. Classification of crown morphology and calculation of crown volume
树冠形状 树种 体积公式 球形 四季桂Osmanthus fragrans、黄金榕Ficus microcarpa V=πx2y/6 卵形 白兰Michelia × alba、垂柳Salix babylonica、翻白叶树Pterospermum heterophyllum、 红苞木Rhodoleia championii、黄金香柳Melaleuca bracteata、黄皮Clausena lansium、木荷 Schima superba、桑Morus alba、铁冬青Ilex rotunda、土蜜树Bridelia tomentosa、樟树 Cinnamomum camphora、海南蒲桃Syzygium hainanense、蒲葵Livistona chinensis V=πx2y/6 圆锥形 罗汉松Podocarpus macrophyllus、竹柏Nageia nagi、马占相思Acacia mangium、台湾相思Acacia confusa、马尾松Pinus massoniana、南洋杉Araucaria columnaris、鱼尾葵Caryota maxima、红 车Syzygium rehderianum V=πx2y/12 半球形/扁圆形 紫薇Lagerstroemia indica、黄槿Hibiscus tiliaceus、大花紫薇Lagerstroemia speciosa、鸡蛋花 Plumeria rubra、黄槐决明Senna surattensis、小叶榕Ficus microcarpa、潺槁木姜子Litsea glutinosa、 刺桐Erythrina variegata、非洲楝Khaya senegalensis、莲雾Syzygium samarangense、蒲桃 Sterculia monosperma、阴香Cinnamomum burmannii、印度紫檀Pterocarpus indicus、高山榕 Ficus altissima、荔枝Litchi chinensis V=πx2y/6 球扇形 大叶相思Acacia auriculiformis、洋紫荆Bauhinia variegata、红花紫荆Bauhinia×blakeana、蓝花 楹Jacaranda mimosifolia、柳叶榕Ficus binnendijkii、窿缘桉Eucalyptus exserta、麻楝 Chukrasia tabularis、杧果Mangifera indica、美丽针葵Phoenix canariensis、秋枫Bischofia javanica、盆架子Alstonia rostrata、散尾葵Chrysalidocarpus lutescens、小叶榄仁Terminalia neotaliala、凤凰木Delonix regia、番木瓜 Carica papaya、美丽异木棉Ceiba speciosa、木棉 Bombax ceiba、黄花风铃木Handroanthus chrysanthus V=π[2y3−y2(4y2−x2)1/2]/3 球缺形 澳洲火焰木Brachychiton acerifolius 、澳洲鸭脚木Schefflera actinophylla、大王椰子Roystonea regia、假槟榔Archontophoenix alexandrae、老人葵Coccothrinax crinita V=π(3xy2−2y3)/6 圆柱形 火焰木Spathodea campanulata、苹婆Sterculia monosperma、白千层Melaleuca cajuputi V=πx2y/4 说明:V为树冠体积(m3);x为平均冠层(m);y为冠高(m) -
林下游憩空间量反映风景游憩林为人们提供通行和游憩空间的大小。根据GB 51192−2016《城市公园设计规范》规定[23],公园集散场地种植的树木枝下净空应大于2.2 m。本研究定义林下游憩空间量为树冠垂直投影覆盖面积与乔木枝下净空乘积(枝下净空大于2.2 m),具体公式为:
$$ V_{\rm{C}} = \sum\limits_{i = 1}^n {{M_i}} {H_i}{\text{。}} $$ 其中:VC表示林下游憩空间量(m3),n表示枝下净空大于2.2 m的乔木数量;Mi表示第i株乔木树冠垂直投影覆盖面积(m2),Hi表示第i株乔木的枝下净空(m)。
-
研究可知:前山公园风景游憩林总绿量为4 442.60 m3,单位面积绿量0.38 m3·m−2,其中单木绿量最大的树种为白兰(262.00 m3),绿量排名前5%的单木绿量合计2 259.23 m3,占总绿量的50.85%,主要树种为白兰、小叶榕和樟树。北理工珠海学院风景游憩林总绿量为8 122.39 m3,单位面积绿量0.64 m3·m−2,其中单木绿量最大的树种为荔枝(261.67 m3),绿量排名前5%的单木绿量合计2 183.33 m3,占总绿量的26.88%,主要树种为荔枝、非洲楝、樟树。梅华城市公园风景游憩林总绿量为18 986.74 m3,单位面积绿量0.81 m3·m−2,其中单木绿量最大的树种为小叶榕(283.58 m3),绿量排名前5%的单木绿量合计4 855.09 m3,占总绿量的25.57%,主要树种为小叶榕、高山榕、凤凰木、阴香。
从3个研究林地的三维绿量核密度(KDE)分布图来看(图1),0~100 m3绿量级内,前山公园风景游憩林的三维绿量分布极不均衡,主要分布于0~25 m3,北理工珠海学院和梅华城市公园风景游憩林的三维绿量主要分布于0~50 m3。100~200 m3绿量级内,前山公园风景游憩林的绿量分布极不均衡,主要集中于100~150 m3,北理工珠海学院和梅华城市公园的三维绿量分布较为均匀,分布密度均小于0.015。200~300 m3绿量级内,前山公园和北理工珠海学院风景游憩林的三维绿量分布相对集中,在210~275 m3内分布密度较高,梅华城市公园分布较为均匀,分布密度小于0.015。
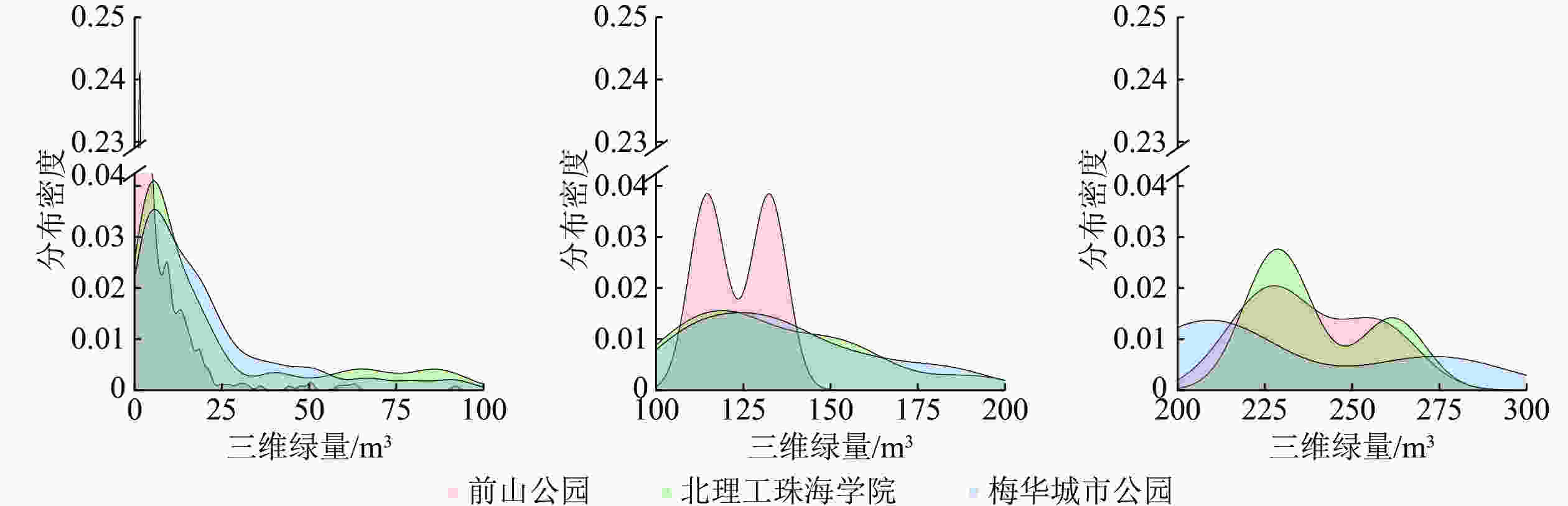
图 1 风景游憩林三维绿量核密度分布
Figure 1. KDE of 3DGQ distribution in study forests
-
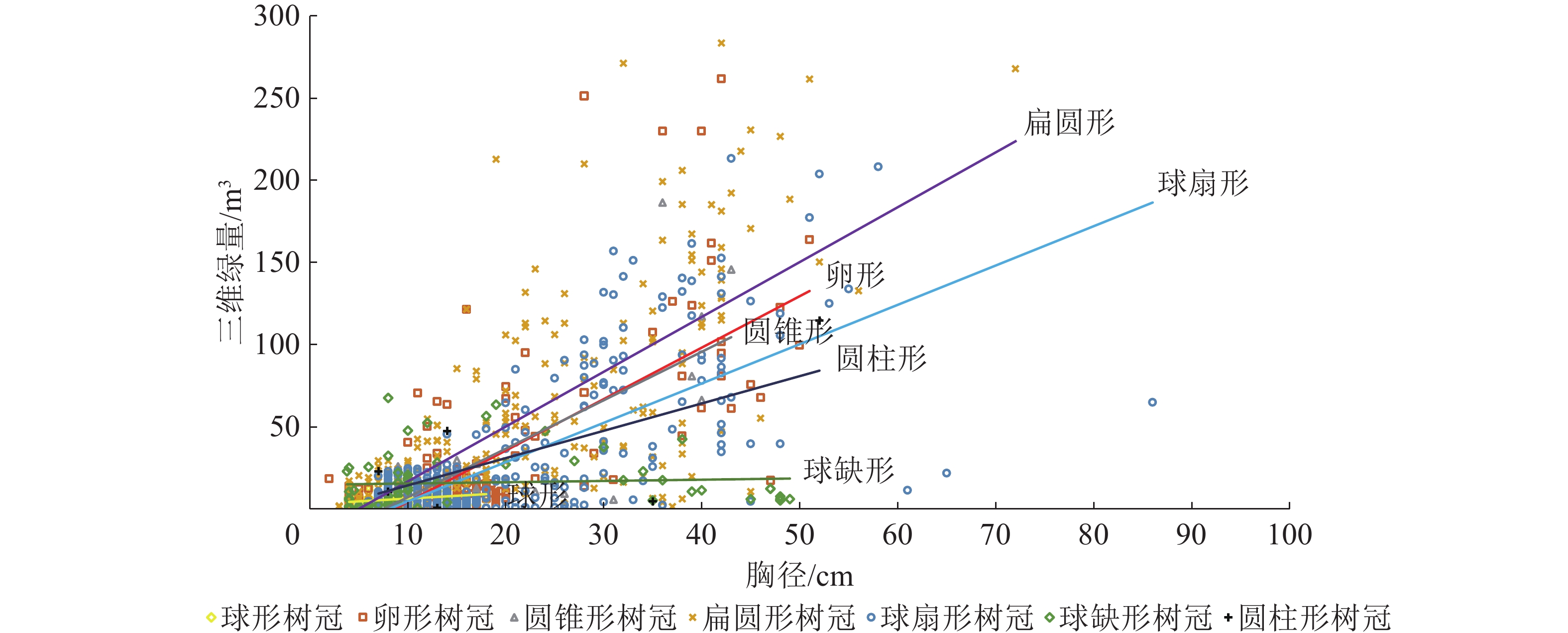
分析3个研究林地每木胸径和三维绿量的关系(图2)可知:三维绿量大于200 m3的树种,单木胸径多为30~50 cm,树冠以卵形、扁圆形和球扇形为主。随着胸径增加,扁圆形、卵形、圆锥形、球扇形和圆柱形树冠的树种三维绿量明显上升,而球缺形和球形树冠的树种三维绿量并无明显提升。不同树冠形状树种胸径与单木三维绿量的正相关系数从大到小依次为扁圆形、卵形、圆锥形、球扇形、圆柱形、球缺形、球形。

图 2 树木胸径和三维绿量相关关系
Figure 2. Relationship between the DBH and 3DGQ
-
研究得知:前山公园风景游憩林林下空间总量为5 543.75 m3,林下游憩空间量为2 148.86 m3,占比38.88%,单位面积游憩空间量0.18 m3·m−2,其中单木林下游憩空间最大的树种为樟树(66.55 m3),林下游憩空间量排名前5%的单木游憩空间量合计813.05 m3,占游憩总空间量的37.84%,主要树种为樟树、凤凰木、海南蒲桃、秋枫、刺桐等。北理工珠海学院林下空间总量为10 534.72 m3,林下游憩空间量为7 880.47 m3,占比74.80%,单位面积游憩空间量0.63 m3·m−2,其中单木林下游憩空间最大的树种为樟树(235.20 m3),林下游憩空间量排名前5%的单木游憩空间量合计2 017.77 m3,占游憩总空间量的25.60%,主要树种为樟树、木棉、非洲楝、小叶榄仁等。梅华城市公园林下空间总量为16 662.62 m3,林下游憩空间量为12 914.97 m3,占比77.51%,单位面积游憩空间量0.55 m3·m−2,其中单木林下游憩空间最大的树种为高山榕(252.00 m3),林下游憩空间量排名前5%的单木游憩空间量合计4 649.33 m3,占游憩总空间量的36.00%,主要树种为高山榕、凤凰木、美丽异木棉。
-
对3个研究林地中三维绿量和林下游憩空间前5%且种群数量较大的9个树种进行分析,比较不同树种冠层空间与林下游憩空间的分配策略。从图3中可以得出:所有9个树种,50%以上单株的三维绿量和林下游憩空间比值小于1.0,表明在城市公园风景游憩林中,树木倾向于分配更多的林下游憩空间。其中,凤凰木、美丽异木棉、木棉和秋枫的三维绿量和林下游憩空间比值均小于0.5,表明这些树种的空间分配策略,林下游憩空间占据了绝对优势;非洲楝、海南蒲桃、小叶榕和樟树的三维绿量和林下游憩空间比值总体为0.5~1.0,表明这些树种的空间分配策略为林下游憩空间稍优于三维绿量;而白兰的三维绿量和林下游憩空间比值总体为0.5~2.0,表明白兰的空间分配策略有较强的多样性。
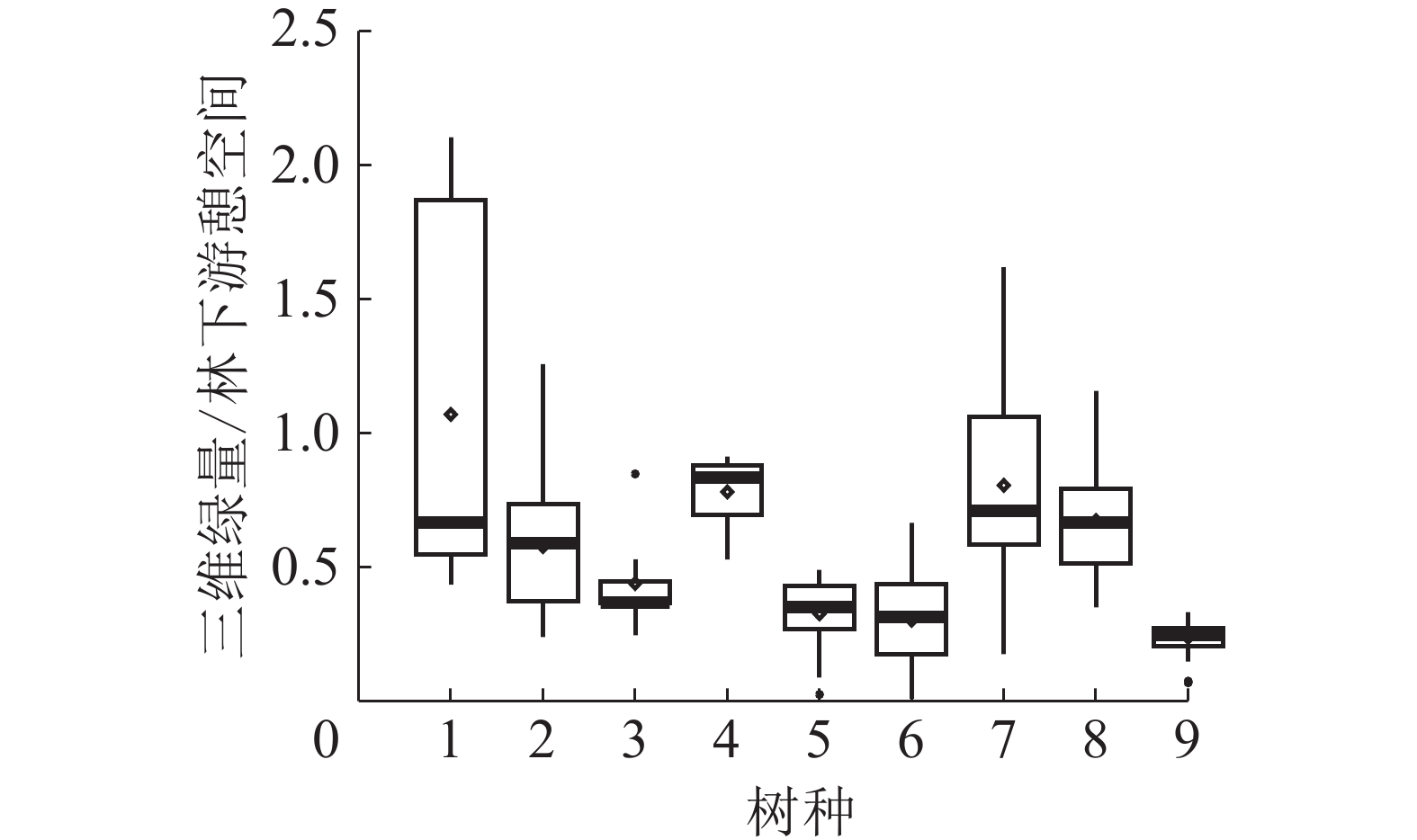
图 3 不同树种三维绿量和林下游憩空间分配
Figure 3. Allocation strategy of 3DGQ and recreation space of different tree species
-
城市森林的三维绿量一定程度上反映城市森林生态服务功能的强弱。LIANG等[24]对上海公共绿地乔木的研究结果表明:乔木三维绿量与树冠投影面积呈现显著正相关;李晓婷[25]研究发现:随着研究林地的地理位置由城市核心区向城市近郊区过渡,乔木平均冠幅呈现逐渐减小趋势,单位面积三维绿量逐渐减小,绿量分布趋于集中。本研究中,0~300 m3绿量区间内,梅华城市公园绿量分布密度呈现“平缓均衡”的趋势,北理工珠海学院呈现“中间均衡,两头集中”的趋势,前山公园呈现“由集中急速向均衡”改变的趋势。与郑绍伟等[26]得出的大径级乔木数量随绿量级增大逐渐减小的结论略有差异,极有可能是树种形态不同导致的。本研究发现:每木胸径和三维绿量的正相关关系受树冠形态影响;相同胸径的球形乔木和扁圆形乔木,三维绿量差异较大。因此推测,相同绿量级的乔木径级很有可能处于不同区间,这也是北理工珠海学院绿量分布密度“中间均衡,两头集中”的原因。
林冠空间的三维绿量提供了生态服务功能,林下空间提供了有效的游憩功能。城市公园风景游憩林三维绿量和林下游憩空间量的比值不同,反映出不同树种在空间分配策略上的差异。凤凰木、美丽异木棉、木棉属于典型的观花景观树种,林冠空间是决定景观效果的关键性因子;然而从空间分配策略上看,这3个树种的林下游憩空间占据绝对优势,与树种自身的特点有矛盾。林下游憩空间的主要影响因子为树冠投影面积与最低分支点。然而在实际城市环境中,游憩空间主要由林下空旷地和林下少量植被共同构成[27],林下的小乔木和灌木对林下游憩的真实活动空间具有直接影响;平衡林下游憩真实的活动空间和林下植被量,关系到风景游憩林群落中下层的配置和游憩功能的发挥。因此,今后需构建反映林下游憩真实的活动空间和林下植被量的综合评价指标参数,从而更加真实地反映和评价城市风景游憩林林下游憩空间配置。
林木树冠三维绿量和林下游憩空间的分布格局,演替、生长、物候等引起的时间动态变化,森林群落学与生态学等相关学科的融合交叉,是进一步深入研究城市森林、城市植被提供生态与景观服务功能的有效途径[28]。本研究针对城市风景游憩林的林冠空间和林下空间展开,重点集中在体量、空间分布、分配策略等方面,是一种静态研究结果;而树冠三维绿量和林下游憩空间实则为一个动态过程,群落演替、生长、物候等都将引起时间、空间的动态变化。因此,构建反映动态过程的指标参数,可以更加科学地反映和评价城市风景游憩林的景观游憩功能。本研究基于树冠、树形、枝下高等数据测算出三维绿量和林下游憩空间,人力、时间成本巨大。今后可以利用搭载多光谱镜头的无人机监测城市森林斑块的冠层动态演变[29],运用孔径雷达估测大面积城市森林地上生物量的空间格局[30],从而快速获取更高精度的包括树冠结构、林下空间、生长演替、物候变迁在内的城市森林多维时空尺度数据,提升研究者对城市森林生态与景观功能的认知。
-
珠海城市公园风景游憩林单位面积绿量由城市核心区向近郊区逐渐降低。从三维绿量密度分布上看,处在城市近郊区的前山公园风景游憩林绿量分布较为集中,而处在城市核心区的梅华城市公园绿量分布较为均衡。单木绿量较大的树种有白兰、小叶榕、樟树、荔枝、非洲楝、高山榕、凤凰木、阴香。从每木胸径和三维绿量的关系分析,不同树冠形状树种胸径和三维绿量的正相关系数从大到小依次为扁圆形、卵形、圆锥形、球扇形、圆柱形、球缺形、球形。三维绿量和林下游憩空间量的比例关系一定程度上反映树木对提供观赏和生态服务功能的冠层空间和提供游憩功能的林下空间的分配策略;在风景游憩林构建过程中,应结合不同树种的自身特点分配冠层空间和林下空间。
Characteristics of the three-dimensional green quantity and recreation space for landscape-recreational forests in Zhuhai City
-
摘要:
目的 明确不同生态位风景游憩林的三维绿量(3DGQ)和林下游憩空间量(FRS)特征,揭示风景游憩林构建的规律和存在的问题。 方法 采用群落生态学的方法分别在广东省珠海市前山公园(13个样方)、北理工珠海学院(14个样方)和梅华城市公园(26个样方)设置风景游憩林群落研究林地,结合高分遥感影像,进行每木调查。测算三维绿量和林下游憩空间量,分析不同区位风景游憩林的差异。 结果 位于城市近郊区(UFA)的前山公园风景游憩林单位面积绿量最低。单木绿量较大的树种有白兰Michelia × alba、小叶榕Ficus microcarpa、樟树Cinnamomum camphora、荔枝Litchi chinensis、非洲楝Khaya senegalensis、高山榕Ficus altissima、凤凰木Delonix regia、阴香Cinnamomum burmannii。3个研究林地三维绿量分布有一定差异,位于城市核心区(CUA)的梅华城市公园三维绿量分布较均匀;树木胸径与其三维绿量呈正相关,相关系数与树种树冠形状有关。凤凰木、美丽异木棉Ceiba speciosa、木棉Bombax ceiba和秋枫Bischofia javanica的空间分配策略为林下游憩空间占据绝对优势,白兰的空间分配策略有较强的多样性。 结论 随着研究林地地理位置由城市核心向城市近郊过渡,单位面积三维绿量逐渐减小,绿量分布迅速趋于集中;胸径与三维绿量的正相关关系受树冠形态影响;风景游憩林构建过程中,冠层空间和林下游憩空间的分配应结合树种特点进行。图3表1参30 Abstract:Objective This study is aimed to clarify the three-dimensional green quantity (3DGQ) and the characteristics of forest recreation space (FRS) of landscape-recreational forests at different niches, and reveal the rules of constructing landscape-recreation forests and the existing problems. Method On the basis of the community ecology theory, with 13, 14 and 26 sample plots selected from Qianshan Park, Zhuhai Institute of BIT, and Meihua City Park respectively as the subjects, each tree was surveyed using high-resolution remote sensing images before 3DGQ and FRS were calculated to analyze the differences between different location of landscape-recreation forests. Result Qianshan Park, located in the core urban area (CUA), had the lowest 3DGQ density. Michelia × alba, Ficus microcarpa, Cinnamomum camphora, Litchi chinensis, Khaya senegalensis, Ficus altissima, Delonix regia and Cinnamomum burmannii displayed high values of 3DGQ. The three sites showed different spatial distributions of 3DGQ, and Meihua City Park which was located in the core urban area had relatively even distribution of 3DGQ than the others. There was a positive correlation between DBH and 3DGQ, which was related to the crown shapes of trees. In terms of the spatial allocation, for D. regia, Ceiba speciose, Bombax ceiba and C. camphora, superiority was given to FRS, while for M. ×alba, various spatial allocation strategies were adopted. Conclusion The 3DGQ density decreases while the distribution of 3DGQ showed a strong central tendency with the location of landscape-recreational forests shifting from the core urban area to the sub-urban area (SUA) and finally to urban fringe area (UFA). The correlation coefficient between DBH and 3DGQ is related to the crown shapes of tree species and the features of different tree species in the progress of the construction of landscape-recreation forest should be taken into consideration in the allocation of canopy space and recreation space. [Ch, 3 fig. 1 tab. 30 ref.] -
图 3 不同树种三维绿量和林下游憩空间分配
1. 白兰;2. 非洲楝;3. 凤凰木;4. 海南蒲桃;5. 美丽异木棉;6. 木棉;7. 小叶榕;8. 樟树;9. 秋枫
Figure 3 Allocation strategy of 3DGQ and recreation space of different tree species
表 1 树冠形态分类及树冠体积
Table 1. Classification of crown morphology and calculation of crown volume
树冠形状 树种 体积公式 球形 四季桂Osmanthus fragrans、黄金榕Ficus microcarpa V=πx2y/6 卵形 白兰Michelia × alba、垂柳Salix babylonica、翻白叶树Pterospermum heterophyllum、 红苞木Rhodoleia championii、黄金香柳Melaleuca bracteata、黄皮Clausena lansium、木荷 Schima superba、桑Morus alba、铁冬青Ilex rotunda、土蜜树Bridelia tomentosa、樟树 Cinnamomum camphora、海南蒲桃Syzygium hainanense、蒲葵Livistona chinensis V=πx2y/6 圆锥形 罗汉松Podocarpus macrophyllus、竹柏Nageia nagi、马占相思Acacia mangium、台湾相思Acacia confusa、马尾松Pinus massoniana、南洋杉Araucaria columnaris、鱼尾葵Caryota maxima、红 车Syzygium rehderianum V=πx2y/12 半球形/扁圆形 紫薇Lagerstroemia indica、黄槿Hibiscus tiliaceus、大花紫薇Lagerstroemia speciosa、鸡蛋花 Plumeria rubra、黄槐决明Senna surattensis、小叶榕Ficus microcarpa、潺槁木姜子Litsea glutinosa、 刺桐Erythrina variegata、非洲楝Khaya senegalensis、莲雾Syzygium samarangense、蒲桃 Sterculia monosperma、阴香Cinnamomum burmannii、印度紫檀Pterocarpus indicus、高山榕 Ficus altissima、荔枝Litchi chinensis V=πx2y/6 球扇形 大叶相思Acacia auriculiformis、洋紫荆Bauhinia variegata、红花紫荆Bauhinia×blakeana、蓝花 楹Jacaranda mimosifolia、柳叶榕Ficus binnendijkii、窿缘桉Eucalyptus exserta、麻楝 Chukrasia tabularis、杧果Mangifera indica、美丽针葵Phoenix canariensis、秋枫Bischofia javanica、盆架子Alstonia rostrata、散尾葵Chrysalidocarpus lutescens、小叶榄仁Terminalia neotaliala、凤凰木Delonix regia、番木瓜 Carica papaya、美丽异木棉Ceiba speciosa、木棉 Bombax ceiba、黄花风铃木Handroanthus chrysanthus V=π[2y3−y2(4y2−x2)1/2]/3 球缺形 澳洲火焰木Brachychiton acerifolius 、澳洲鸭脚木Schefflera actinophylla、大王椰子Roystonea regia、假槟榔Archontophoenix alexandrae、老人葵Coccothrinax crinita V=π(3xy2−2y3)/6 圆柱形 火焰木Spathodea campanulata、苹婆Sterculia monosperma、白千层Melaleuca cajuputi V=πx2y/4 说明:V为树冠体积(m3);x为平均冠层(m);y为冠高(m)  下载: 导出CSV
下载: 导出CSV
-
[1] PIRNAT J. Conservation and management of forest patches and corridors in suburban landscapes [J]. Landscape Urban Plann, 2000, 52(2/3): 135 − 143. [2] NOVÁKOVÁ J. Vegetation changes in Prague’s suburban forest during the last 40 years: human impact and legislative protection [J]. Urban For Urban Greening, 2008, 7(4): 301 − 314. [3] 陈鑫峰, 沈国舫. 森林游憩的几个重要概念辨析[J]. 世界林业研究, 2000, 13(1): 69 − 73. CHEN Xinfeng, SHEN Guofang. Dissection of some concepts about forest recreation [J]. World For Res, 2000, 13(1): 69 − 73. [4] 国家市场监督管理总局, 中国国家标准化管理委员会. 国家森林城市评价指标: GB T37342-2019[S]. 北京: 中国标准出版社, 2019. [5] 赵庆, 唐洪辉, 魏丹, 等. 基于绿视率的城市绿道空间绿量可视性特征[J]. 浙江农林大学学报, 2016, 33(2): 288 − 294. ZHAO Qing, TANG Honghui, WEI Dan, et al. Spatial visibility of green areas of urban greenway using the green appearance percentage [J]. J Zhejiang A&F Univ, 2016, 33(2): 288 − 294. [6] MILLER R W. Urban Forestry[M]. New Jersey: Prentice Hall, 1996. [7] 陈丹, 彭蓉. 台湾地区国家公园环境教育体系浅析: 以金门国家公园为例[J]. 林产工业, 2019, 46(5): 62 − 64. CHEN Dan, PENG Rong. Analysis on environmental education system in China’s Taiwan National Parks: take Jinmen National Park as an example [J]. China For Prod Ind, 2019, 46(5): 62 − 64. [8] 梁星权. 城市林业[M]. 北京: 中国林业出版社, 2001. [9] 刘常富, 李海梅, 何兴元, 等. 城市森林概念探析[J]. 生态学杂志, 2003, 22(5): 146 − 149. LIU Changfu, LI Haimei, HE Xingyuan, et al. Concept discussion and analysis of urban forest [J]. Chin J Ecol, 2003, 22(5): 146 − 149. [10] 王孟欣. 福建华安九龙江湿地公园保护与利用研究[J]. 林产工业, 2019, 46(4): 61 − 64. WANG Mengxin. Protection and utilization of Jiulongjiang Wetland Park in Fujian Huaan [J]. China For Prod Ind, 2019, 46(4): 61 − 64. [11] BREUSTE J H. Decision making, planning and design for the conservation of indigenous vegetation within urban development [J]. Landscape Urban Plann, 2004, 68(4): 439 − 452. [12] 宋永昌. 植被生态学[M]. 上海: 华东师范大学出版社, 2001. [13] 何兴元, 金莹杉, 朱文泉, 等. 城市森林生态学的基本理论与研究方法[J]. 应用生态学报, 2002, 13(12): 1679 − 1683. HE Xingyuan, JIN Yingshan, ZHU Wenquan, et al. Basic theory and research method of urban forest ecology [J]. Chin J Appl Ecol, 2002, 13(12): 1679 − 1683. [14] 周红敏, 惠刚盈, 赵中华, 等. 森林结构调查中最适样方面积和数量的研究[J]. 林业科学研究, 2009, 22(4): 485 − 495. ZHOU Hongmin, HUI Gangying, ZHAO Zhonghua, et al. Studies on the area and the number of the sample for forest structure [J]. For Res, 2009, 22(4): 485 − 495. [15] 郭雪艳. 南京市常见园林植物的绿量研究[D]. 南京: 南京林业大学, 2009. GUO Xueyan. Research on the Living Vegetation Volume of Common Landscape Plants in Nanjing[D]. Nanjing: Nanjing Forestry University, 2009. [16] 王东良, 金荷仙, 范丽琨, 等. 疗养院人工绿地三维绿量分布特征及影响因子[J]. 浙江农林大学学报, 2013, 30(4): 529 − 535. WANG Dongliang, JIN Hexian, FAN Likun, et al. Distribution and influencing factor of three-dimensional green biomass for artificial greenbelts in sanatoriums [J]. J Zhejiang A&F Univ, 2013, 30(4): 529 − 535. [17] 王帆. 深圳坝光区域滨海河溪红树植物群落生态与景观特性研究[D]. 广州: 仲恺农业工程学院, 2017. WANG Fan. The Study on The Ecology and Landscape of Mangrove Communities in Coastal Rivers of Baguang Community, Shenzhen[D]. Guangzhou: Zhongkai University of Agriculture and Engineering, 2017. [18] 祖若川. 海口市公园抗风园林植物的选择与应用[D]. 海口: 海南大学, 2016. ZU Ruochuan. Haikou Park Selection and Application of Wind-Resistant Garden Plants[D]. Haikou: Hainan University, 2016. [19] 乔小菊. 南京城区园林绿化中常见阔叶乔木树种的光合特性及相关生态功能的研究[D]. 南京: 南京农业大学, 2016. QIAO Xiaoju. Nanjing City Landscaping Broadleaf Tree Species of Photosynthetic Characteristics and Associated Ecosystem Functions[D]. Nanjing: Nanjing Agriculture University, 2016. [20] 吴显坤. 台风灾害对深圳城市园林树木的影响和对策[D]. 南京: 南京林业大学, 2007. WU Xiankun. Effect of Typhoon Disasters to Urban Landscape Trees and Typhoon Disaster-Reducing Strategies in Shenzhen[D]. Nanjing: Nanjing Forestry University, 2007. [21] 王燕. 广东省生态景观林带生态效益和景观评价[D]. 广州: 华南农业大学, 2016. WANG Yan. Ecological Benefit and Landscape Aesthetic Value of Eco-Landscape Forest Belt in Guangdong Province: A Case Study of Huizhou City and Meizhou City[D]. Guangzhou: South China Agricultural University, 2016. [22] 陈莉. 福州市棕榈科植物景观特性及评价[D]. 福州: 福建农林大学, 2015. CHEN Li. Analysis on the Landscaping Characteristics of Palms and Ecaluation of Its Application in Fuzhou[D]. Fuzhou: Fujian Agriculture and Forestry University, 2015. [23] 北京市园林绿化局. 公园设计规范: GB51192-2016[S]. 北京: 中国标准出版社, 2019. [24] LIANG Huilin, LI Weizheng, ZHANG Qingping, et al. Using unmanned aerial vehicle data to assess the three-dimension green quantity of urban green space: a case study in Shanghai, China [J]. Landscape Urban Plann, 2017, 164: 81 − 90. [25] 李晓婷. 北京城区校园与医院林木树冠覆盖与城市森林结构研究[D]. 北京: 中国林业科学研究院, 2018. LI Xiaoting. A Study on Urban Tree Canopy and Urban Forest Structure of Schools and Hospitals in Beijing Urban Area[D]. Beijing: Chinese Academy of Forestry, 2018. [26] 郑绍伟, 李隽, 黎燕琼, 等. 利用形态因子建立城市森林主要乔木树种三维绿量预测模型[J]. 四川林业科技, 2017, 38(1): 6 − 10. ZHENG Shaowei, LI Jun, LI Yanqiong, et al. Establishing the three dimensional green quantity forescast model of main tree species in urban forest by using morphological factors [J]. J Sichuan For Sci Technol, 2017, 38(1): 6 − 10. [27] ZHAO Qing, XU Dongxian, QIAN Wanhui, et al. Ecological and landscape perspectives on urban forest planning and construction: a case study in guangdong-hongkong-macao greater bay area of China [J]. Front Sustainable Cities, 2020, 2: 44. doi: 10.3389/frsc.2020.00044. [28] ZHAO Q, TANG H, GAO C J, et al. Evaluation of urban forest landscape health: a case study of the Nanguo Peach Garden, China [J]. For Biogeosci For, 2020, 13: 175. doi: 10.3832/ifor3206-013. [29] TIAN Jinyan, WANG Le, LI Xiaojuan, et al. Comparison of UAV and WorldView-2 imagery for mapping leaf area index of mangrove forest [J]. Int J Appl Earth Observ Geoinf, 2017, 61: 22 − 31. [30] RANSON K J, SUN G, WEISHAMPEL J F, et al. Forest biomass from combined ecosystem and radar backscatter modeling [J]. Remote Sensing Environ, 1997, 59(1): 118 − 133. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200455
 点击查看大图
点击查看大图

图(3) / 表(1)
计量
- 文章访问数: 2091
- HTML全文浏览量: 499
- PDF下载量: 79
- 被引次数: 0

