







-
农作物的精准识别有助于人们及时、准确地掌握农作物的种植结构及其时空变化信息,对区域内农作物的空间格局分布、产量的预测、农业资源的调查和灾害监测等问题具有重要意义[1]。传统多光谱遥感受传感器波段少、光谱分辨率低、作物光谱相似性等问题的影响,无法获得较高的作物类型识别精度;而新兴的高光谱技术通过获取连续地物的光谱信息,能在众多窄波段范围内对作物的细微差别进行探测,进而提高作物识别精度[2]。近年来国内外学者利用高光谱数据在作物识别与分类方面已开展了大量研究工作[3-6]。高光谱数据波段多、数据量大、信息冗余严重,因此在数据的应用与处理中如何选取有效的光谱数据形式和光谱特征变量是研究的重点。目前,该类研究对作物分类识别的精度不断提升,但这些方法主要基于传统统计方法结合主观判断选择波段,主观性较大,且数据处理操作较为繁琐,难以高效简便对作物进行精确分类识别。本研究以杭州地区常见的8种作物作为研究对象,基于实测的叶片光谱反射率数据,通过不同特征提取与分类方法对作物光谱分析,探寻识别不同作物的高效方法,从而为作物高光谱遥感解译和精准分类提供参考。
-
研究区浙江省杭州市(29°11′~30°33′N,118°21′~120°30′E),地处中国东南沿海、浙江省北部。研究区属亚热带季风气候,光温同步,雨热同季,日照和无霜期较长。研究区西部连山,东部近海,地势西高东低,地形地貌复杂多样。研究区具有丰富的水稻土,约占土壤总面积的14%,主要分布在平原地区,适宜于多种作物生长。研究区主要农作物有大豆Glycine max、番茄Lycopersicon esculentum、茄Solanum melongena、水稻Oryza sativa、茶Camellia sinensis、葡萄Vitis vinifera、玉米Zea mays、山核桃Carya cathayensis、番薯Ipomoea batatas、花生Arachis hypogaea、四季豆Phaseolus vulgaris等。
-
选取杭州市常见的大豆、玉米、茄、四季豆、花生、葡萄、番薯、水稻共8种农作物作为光谱测试对象。光谱测试部位为农作物叶片,叶片反射光谱采用美国ASD FieldSpec Pro FR地物光谱仪(光谱范围350~2 500 nm)进行测量。测定时间为2018年7月上旬,每天8:00-10:00,在天空晴朗无云,无风或微风,空气湿度小的情况下,户外采集农作物叶片后立即在室内用植被探头测量8种作物的叶片反射率。每种作物选择5片叶片,每片叶片选择3处测量点,以每一测量点连续测量10次的光谱平均值作为其光谱反射值,共获得150条光谱曲线,再对每10条光谱数据取其平均值作为一个样本数据,每种作物得到15个光谱样本数据。剔除有明显异常的波段数据,以剩余数据的平均值作为该样点的光谱反射率,再对光谱曲线进行平滑处理,消除光谱曲线上存在的噪声,最终获得8种作物反射光谱曲线数据。
-
采用随机森林方法与传统的高光谱识别与分类方法处理并分析作物高光谱数据,提取识别不同作物类型的高光谱特征,对作物进行精准识别与分类,并对结果分析与比较,这些传统方法包括一阶微分、二阶微分、倒数的对数等数学变换方法、去包络线法等。
-
① 简单数学变换法:运用光谱微分方法处理光谱曲线,能够部分消除大气效应、作物环境背景(阴影、土壤等)的影响,以反映作物本身的光谱特征[7];对数据进行倒数的对数可以减少因光照等变化引起的乘性因素对光谱数据的影响,使可见光区范围内光谱数据差异增大,从而更容易识别不同的作物[8]。采用一阶微分、二阶微分以及倒数的对数变换对原始光谱进行处理,观察分析光谱特征及其区分不同作物的能力。②去包络线法:包络线(envelope)是指每条光谱曲线的外凸包曲线,去包络线(continuum removal)是一种非线性光谱变换方法[9]。去包络线法对作物光谱曲线上反射率小、光谱曲线相近的可见光波段处理有效,能在很大程度上放大作物间光谱差异性,有利于作物识别分类[10]。本研究用MATLAB软件对原始光谱曲线进行去包络线处理,提取作物间光谱差异较大的波段,再使用欧氏距离法[11]对不同作物识别与分类。③随机森林法(random forest, RF)[12]:是一种基于分类与回归决策树(classification and regression tree, C&RT)的组合算法。随机森林算法对参与分类的变量没有限定,在处理高维数据分类时,更能体现随机森林的速度快、精度高、稳定性好的优势[13]。因此用随机森林法处理作物高光谱数据时,不用提前做光谱特征提取,在实施分类的同时,就可以对高光谱变量进行筛选优化[14],并分析判断特征波段的优劣。
-
图 1为8种作物的平均光谱曲线,在可见光波段,绿峰波段反射率差异最大;在近红外波段,800~1 300 nm波段的光谱曲线差异最明显,反射率由高到低顺序为玉米、茄、四季豆、大豆、番薯、花生、葡萄、水稻,其中茄与玉米反射率较为接近,较难区分;在1 600~1 800 nm波段内不同作物的光谱曲线差异较为明显,除了葡萄与花生、茄与玉米的光谱差异较小外,剩余作物之间通过反射率差异可以区分。
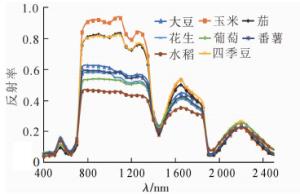
图 1 不同作物的平均光谱曲线
Figure 1. Average spectral curves of different crops
-
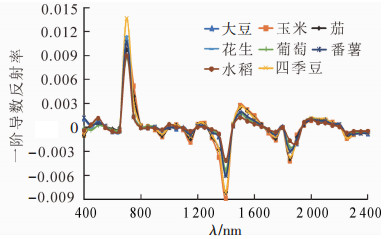
从图 2可观察到:8种作物的一阶微分光谱曲线的变化走向基本一致,在极大或极小值对应的波段,不同作物的一阶微分值差异较为明显, 其他波段内作物曲线相差不大。在685~770 nm波段内可以区分四季豆与其他作物,但不能明显区分出其余作物;在1 350~1 430 nm波段内不同作物一阶微分值有较大差异,能够区分出水稻、玉米与葡萄3种作物,其余作物无明显差别。

图 2 不同作物的一阶导数光谱曲线
Figure 2. First derivative spectral curves of different crops
-
由图 3可知:8种作物二阶导数光谱曲线在650~790、1 300~1 500和1 820~1 940 nm等3个波段区间差异较大。表 1统计了8种作物在这3个波段内的极大值及其对应波段。即便在这些差异较大的波段内,部分作物的二阶导数光谱曲线十分接近,不利于区分不同作物;比如玉米、番薯、大豆、花生、葡萄在650~790 nm波段的光谱二阶导数值相近,大豆、花生、葡萄在1 300~1 500 nm波段的光谱二阶导数值相近,番薯、大豆、花生、葡萄在1 820~1 940 nm波段的光谱二阶导数值相近,这些二阶导数值相近的作物彼此难以区分。
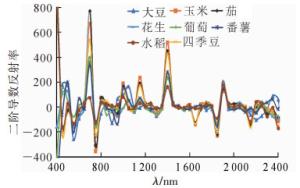
图 3 不同作物的二阶导数光谱曲线
Figure 3. Second derivative spectra of different crops
表 1 作物反射光谱二阶导数极大值及对应波段
Table 1. Maximum value of second derivative of crop reflection spectrum and corresponding band
作物类型 680~700 nm极大值对应波段 极大值×1 000 000 1 390~1 460 nm极大值对应波段 极大值×1 000 000 1 880~1 940 nm极大值对应波段 极大值×1 000 000 四季豆 693 1 101.0 1 401 431.4 1 888 416.4 茄 697 8 885.0 1 401 449.5 1 888 389.5 玉米 694 869.8 1 399 522.8 1 886 419.5 番薯 693 829.6 1 403 307.3 1 890 362.7 大豆 692 828.6 1 403 265.1 1 890 340.7 花生 691 788.7 1 403 284.2 1 890 318.9 葡萄 694 759.1 1 403 260.9 1 891 322.3 水稻 692 632.3 1 406 192.0 1 891 279.7 -
从图 4可知:与原始光谱曲线相比,波峰波谷发生了倒置,出现了“两峰一谷”,峰谷凸凹程度明显增加[15],原本相近的部分作物光谱曲线有所拉开,在800~1 300 nm波段内,水稻、玉米可以明显区分,但大豆、番薯和花生3种作物间的光谱差别过小,不易区分。
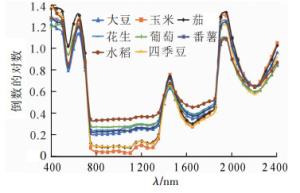
图 4 不同作物光谱倒数的对数曲线
Figure 4. Logarithmic curves of the reciprocal spectra of different crops
-
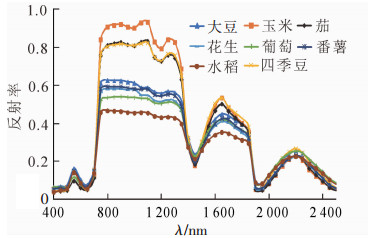
运用MATLAB软件,对作物反射光谱数据做去包络线处理。图 5可以看出:作物光谱在蓝谷,红谷,绿峰,1 170~1 190 nm,1 430~1 450 nm以及1 910~1 930 nm等波段上有较明显的差异。
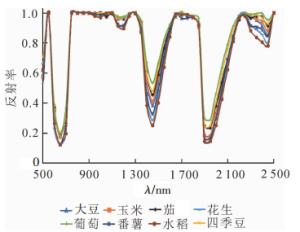
图 5 作物包络线去除后的光谱曲线
Figure 5. Spectral curve after crops envelope removal
利用这些差异较大的波段反射率,基于欧氏距离分类法,评价分析任意2种作物间的可分离性。以绿峰波段为例,任意2种作物间的欧氏距离结果见表 2。可以看出,绿峰波段,玉米、茄、花生等可以较为明显区分,其余作物较难区分。运用相同方法得出其他波段范围结果:在蓝谷波段,葡萄、茄、大豆、玉米、番薯、四季豆可以较明显区分,但水稻和花生较难以区分;红谷波段,茄、玉米、四季豆3者较难区分,其余作物较好区分;1 170~1 190 nm(水氧吸收波段)波段,葡萄、大豆、花生3种作物较难区分;1 430~1 450 nm波段,葡萄、大豆、四季豆3种作物较难区分;1 910~1 930 nm波段,葡萄、大豆两者较难区分。
表 2 绿峰波段任意2种作物间的欧氏距离表
Table 2. Significant Euclidean distance table between any two crops
作物类型 番薯 花生 大豆 葡萄 茄 水稻 四季豆 玉米 番薯 0.010 9 0.014 1 0.017 9 0.025 1 0.045 7 0.009 2 0.009 0 0.009 0 花生 0.014 1 0.015 8 0.016 3 0.039 9 0.060 5 0.017 7 0.018 0 0.018 0 大豆 0.017 9 0.016 3 0.008 1 0.043 9 0.064 6 0.020 3 0.020 9 0.020 9 葡萄 0.025 1 0.039 9 0.043 9 0.015 6 0.019 5 0.025 6 0.025 1 0.025 1 茄 0.045 7 0.060 5 0.064 6 0.019 5 0.007 2 0.042 1 0.041 5 0.041 5 水稻 0.009 2 0.017 7 0.020 3 0.025 6 0.042 1 0.016 7 0.016 8 0.016 8 四季豆 0.009 0 0.018 0 0.020 9 0.025 1 0.041 5 0.016 8 0.006 4 0.006 4 玉米 0.009 5 0.017 1 0.019 0 0.027 0 0.043 7 0.016 8 0.005 6 0.016 6 -
采用IBM SPSS modeler 18.0软件构建随机森林模型数据流,对农作物反射率数据处理。在软件中,使用C&RT算法构建随机森林的每棵树,决策树的棵数和候选分割属性集的大小设置均采用默认值,即构建500棵树,样本大小为百分百。作物在可见光波段(380~780 nm),近红外短波(780~1 100 nm),近红外长波(1 100~2 500 nm)的反射光谱特征与作物的生理生化特点有紧密联系。为了比较可见光、近红外波段的光谱对区分不同作物的效果,将作物光谱样本数据按3个不同的波段分成3组数据,与全波段(350~2 500 nm)数据构成4组不同的样本数据。样本数据一半用于随机森林法建模,提取区分不同作物的特征波段,一半用于随机森林法分类,验证所提取波段的重要性。为了减少数据处理量,将光谱反射率每10 nm取平均值用于随机森林法处理。依次将4组样本数据进行随机森林法处理,结果(表 3)表明:4组数据所建模型均都具有较高的分类准确性。随机森林法在建模的同时对参与分类的波段重要性进行了排序,表 4显示4组数据最重要的前10个波段,每个波段特征对分类精度的贡献程度不同,重要程度越高的波段对作物分类精度的影响越大。
表 3 不同波段光谱数据的随机森林分类结果
Table 3. Random forest classification results for spectral data from different bands
输入变量波段/nm 输入的波段数 模型分类准确性/% 模型误分类率/% 380~780 40 82.4 17.6 780~1 100 32 74.8 25.2 1 100~2 500 140 81.5 18.5 350~2 500 215 87.4 12.6 表 4 参与分类的波段重要性排序
Table 4. Sorting the importance of the bands involved in the classification
380~780 nm 波段重要性 780~1 100 nm 波段重要性 1 100~2 500 nm 波段重要性 350~2 500 nm 波段重要性 770 2.37 800 2.20 1 200 1.59 550 1.04 710 2.09 780 1.68 1 110 1.51 2 490 0.93 400 1.82 1 010 1.57 1 150 1.04 370 0.91 380 1.77 1 050 1.21 1 510 0.90 770 0.87 740 1.71 840 1.15 1 230 0.84 560 0.84 670 1.71 790 1.06 1 190 0.81 380 0.83 550 1.59 850 1.01 2 430 0.75 540 0.83 730 1.49 830 0.98 1 140 0.69 530 0.83 390 1.32 810 0.94 1 310 0.66 570 0.74 540 1.15 1 000 0.92 1 930 0.62 350 0.74 利用验证样本对所建随机森林法分类模型精度进行检验,结果(表 5)显示:4组样本的作物分类正确率均在84%及以上,精确度最高的样本组为350~2 500 nm全波段数据,分类精确率达99.17%。
表 5 不同作物分类准确率分析表
Table 5. Analysis table of classification accuracy of different crops
波段/nm 正确 错误 精确度/% 380~780 118 2 98.33 780~1 100 101 19 84.17 1 100~2 500 115 5 95.83 350~2 500 119 1 99.17 为了检验所筛选的特征波段对区分不同作物的效果,依据350~2 500 nm波段样本数据随机森林分类输出的前10个重要波段,从验证样本数据中选出相应波段数据为变量,在模型参数设置不变的情况下进行随机森林分类,分类准确性达100%,在完全区分不同作物的同时,数据量减少达95.34%,在节省时间的同时保证了较高的准确度。前10个波段为550、2 490、370、770、560、380、540、530、570和350 nm波段。其中350和2 490 nm这2个波段在采集反射率时有噪声,虽然经过平滑方法消除了噪声,但不能完全排除噪声对识别区分作物的影响,因此在选择使用时要慎重。重要波段中的350、370和380 nm等3个波段属于蓝光吸收谷;530、540、550、560和570 nm等5个波段处于绿光反射峰;770 nm波段处于植被反射光谱陡坡;2 490 nm波段处于作物所含水分和二氧化碳的强吸收带。不同作物在这些波段的反射率差异均与作物的叶绿素含量、水分含量、叶片海绵组织及光合作用强烈相关[16]。由此可得出结论,随机森林法筛选出的特征波段不仅区分不同作物效果显著,而且能够反映不同作物生理与生化特性的差异,从生物本身特性的角度为高光谱区分不同作物提供了印证与依据。
-
本研究发现随机森林法对350~2 500 nm全波段反射率数据处理,不仅筛选出能够区分不同作物的特征波段,而且运用所选择的波段对作物进行分类识别的效果也是最优的。不仅如此,随机森林法对筛选出的波段进行了优劣排序,其中对分类贡献较大的波段集中在蓝光、绿峰、红光等波段,反映了作物生理生化特征差异。区分8种作物的特征波段主要有350~380 nm的蓝光波段、530~570 nm的绿峰波段、770 nm的植被陡坡波段、2 490 nm的水、二氧化碳的吸收波段。运用随机森林法能够克服作物光谱相似性较高、难分类等问题,快速高效确定区分不同作物的特征波段,且分类识别精度高。而用观察法分析不同作物的反射光谱及其一阶微分、二阶微分、倒数的对数,提取同时区分识别8种作物的波段难度较大;去包络线法突出了作物光谱在红谷、蓝谷、绿峰区域等特征波段的差异,但提取的特征波段只能区分部分作物,不能同时对8种作物分类。
Crops identification based on hyperspectral data and random forest method
-
摘要:
目的 不同农作物种类光谱差异小,通过探测众多窄波段范围的细微差别,提取区分不同农作物的特征波段,是目前实现农作物高光谱遥感识别的重要途径。如何提取区分不同农作物的特征波段,进而实现农作物的精确识别是一个挑战。近来出现的随机森林方法在多变量目标的分类识别方法展现了优势,为解决这一难题提供了一个新手段。 方法 利用随机森林法与传统方法分析杭州地区8种典型农作物的反射光谱,提取特征波段并进行分类,对比不同方法的识别效果。 结果 不同作物的反射光谱及其一阶微分、二阶微分、倒数的对数、去包络线法所提取的特征波段只能区分部分作物;随机森林法无需对反射光谱预处理,直接对全波段反射光谱数据处理,不仅筛选出了区分不同作物的特征波段,且运用所选择的波段对作物进行随机森林分类的效果也是最优的。 结论 随机森林法选择的波段(550、2 490、370、770、560、380、540、530、570、350 nm)不仅能区分不同作物,还能反映农作物生化属性的不同,使得用于分类的波段及分类方法体现了不同作物间物化性质的不同,在展现高光谱遥感识别农作物优势的同时,也为大面积农作物遥感精细分类提供借鉴。 Abstract:Objective The difference in hyperspectral reflectance of different crops is small, the subtle differences of the spectral feature enable identification of different crops. This aim is to find an effective way to select spectral feature bands from hundreds of bands and indentify different crops. Method The random forest method and the traditional methods were applied to analyze the reflective hyperspectra of 8 typical crops in Hangzhou, the spectral feature bands were extracted and the crops were distinguished. The traditional methods included first-order derivative, second-order derivative, logarithm of reciprocal, and the de-enveloping line method. Result The reflective hyperspectra of the different crops and the results processed by traditional methods were limited in crops identification, the spectral feature extracted by these traditional ways could not distinguish all of 8 crops. The random forest method was the most effective one for extraction of the spectral feature. Conclusion The extracted spectral feature bands were 550, 2 490, 370, 770, 560, 380, 540, 530, 570 and 350 nm, which not only were applied effectively to distinguish 8 crops, but also reflected the biochemical properties of the crops, which made sense to explain the classification with hyperspectral remote sensing.The random forest method fully demonstrated the advantages of hyperspectral remote sensing for crop identification and spectral feature extraction. The random forest classification can provide reference for large-scale crop fine classification with hyperspectral image. -
Key words:
- forest management /
- hyperspectral remote sensing /
- spectral feature /
- crops /
- random forest /
- classification
-
图 4 不同作物光谱倒数的对数曲线
Figure 4 Logarithmic curves of the reciprocal spectra of different crops
表 1 作物反射光谱二阶导数极大值及对应波段
Table 1. Maximum value of second derivative of crop reflection spectrum and corresponding band
作物类型 680~700 nm极大值对应波段 极大值×1 000 000 1 390~1 460 nm极大值对应波段 极大值×1 000 000 1 880~1 940 nm极大值对应波段 极大值×1 000 000 四季豆 693 1 101.0 1 401 431.4 1 888 416.4 茄 697 8 885.0 1 401 449.5 1 888 389.5 玉米 694 869.8 1 399 522.8 1 886 419.5 番薯 693 829.6 1 403 307.3 1 890 362.7 大豆 692 828.6 1 403 265.1 1 890 340.7 花生 691 788.7 1 403 284.2 1 890 318.9 葡萄 694 759.1 1 403 260.9 1 891 322.3 水稻 692 632.3 1 406 192.0 1 891 279.7  下载: 导出CSV
下载: 导出CSV
表 2 绿峰波段任意2种作物间的欧氏距离表
Table 2. Significant Euclidean distance table between any two crops
作物类型 番薯 花生 大豆 葡萄 茄 水稻 四季豆 玉米 番薯 0.010 9 0.014 1 0.017 9 0.025 1 0.045 7 0.009 2 0.009 0 0.009 0 花生 0.014 1 0.015 8 0.016 3 0.039 9 0.060 5 0.017 7 0.018 0 0.018 0 大豆 0.017 9 0.016 3 0.008 1 0.043 9 0.064 6 0.020 3 0.020 9 0.020 9 葡萄 0.025 1 0.039 9 0.043 9 0.015 6 0.019 5 0.025 6 0.025 1 0.025 1 茄 0.045 7 0.060 5 0.064 6 0.019 5 0.007 2 0.042 1 0.041 5 0.041 5 水稻 0.009 2 0.017 7 0.020 3 0.025 6 0.042 1 0.016 7 0.016 8 0.016 8 四季豆 0.009 0 0.018 0 0.020 9 0.025 1 0.041 5 0.016 8 0.006 4 0.006 4 玉米 0.009 5 0.017 1 0.019 0 0.027 0 0.043 7 0.016 8 0.005 6 0.016 6
下载: 导出CSV
表 3 不同波段光谱数据的随机森林分类结果
Table 3. Random forest classification results for spectral data from different bands
输入变量波段/nm 输入的波段数 模型分类准确性/% 模型误分类率/% 380~780 40 82.4 17.6 780~1 100 32 74.8 25.2 1 100~2 500 140 81.5 18.5 350~2 500 215 87.4 12.6
下载: 导出CSV
表 4 参与分类的波段重要性排序
Table 4. Sorting the importance of the bands involved in the classification
380~780 nm 波段重要性 780~1 100 nm 波段重要性 1 100~2 500 nm 波段重要性 350~2 500 nm 波段重要性 770 2.37 800 2.20 1 200 1.59 550 1.04 710 2.09 780 1.68 1 110 1.51 2 490 0.93 400 1.82 1 010 1.57 1 150 1.04 370 0.91 380 1.77 1 050 1.21 1 510 0.90 770 0.87 740 1.71 840 1.15 1 230 0.84 560 0.84 670 1.71 790 1.06 1 190 0.81 380 0.83 550 1.59 850 1.01 2 430 0.75 540 0.83 730 1.49 830 0.98 1 140 0.69 530 0.83 390 1.32 810 0.94 1 310 0.66 570 0.74 540 1.15 1 000 0.92 1 930 0.62 350 0.74
下载: 导出CSV
表 5 不同作物分类准确率分析表
Table 5. Analysis table of classification accuracy of different crops
波段/nm 正确 错误 精确度/% 380~780 118 2 98.33 780~1 100 101 19 84.17 1 100~2 500 115 5 95.83 350~2 500 119 1 99.17
下载: 导出CSV
-
[1] 杨思睿, 薛朝辉, 张玲, 等.高光谱与LiDAR数据融合研究:以黑河中游张掖绿洲农业区精细作物分类为例[J].国土资源遥感, 2018, 30(4):33 - 40. YANG Sirui, XUE Chaohui, ZHANG Ling, et al. Fusion of hyperspectral and LiDAR data: a case study of fine crop classification in Zhangye Oasis Agricultural Area in the middle reaches of the Heihe River[J]. Remote Sensing Land Resour, 2018, 30(4): 33 - 40. [2] 舒田, 岳延滨, 李莉婕, 等.基于高光谱遥感的农作物识别[J].江苏农业学报, 2016, 32(6):1310 - 1314. SHU Tian, YUE Yanbin, LI Lijie, et al. Crops identification based on hyperspectral remote sensing[J]. Jiangsu J Agric Sci, 2016, 32(6): 1310 - 1314. [3] 陶江玥, 刘丽娟, 庞勇, 等.基于机载激光雷达和高光谱数据的树种识别方法[J].浙江农林大学学报, 2018, 35(2):314 - 323. TAO Jiangyue, LIU Lijuan, PANG Yong, et al. Automatic identification of tree species based on airborne LiDAR and hyperspectral data[J]. J Zhejiang A&F Univ, 2018, 35(2): 314 - 323. [4] 史飞飞, 高小红, 杨灵玉, 等.基于地面高光谱数据的典型作物类型识别方法:以青海省湟水流域为例[J].遥感技术与应用, 2016, 32(2):32 - 39. SHI Feifei, GAO Xiaohong, YANG Lingyu, et al. Identifying typical crop types from ground hyper-spectral data: a case study in the Huangshui River basin, Qinghai Province[J]. Geogr Geo-Inf Sci, 2016, 32(2): 32 - 39. [5] 戴建国, 张国顺, 郭鹏, 等.基于无人机遥感可见光影像的北疆主要农作物分类方法[J].农业工程学报, 2018, 34(18): 122 - 129. DAI Jianguo, ZHANG Guoshun, GUO Peng, et al. Classification method of main crops in northern Xinjiang based on UAV visible waveband images[J]. Transac Chin Soc Agric Eng, 2018, 34(18): 122 - 129. [6] 郭文茜, 任建强, 刘杏认, 等.统计数据总量约束下全局优化阈值的冬小麦分布制图[J].遥感学报, 2018, 22(6):1023 - 1041. GUO Wenqian, REN Jianqiang, LIU Xingren, et al. Winter wheat mapping with globally optimized threshold under total quantity constraint of statistical data[J]. J Remote Sensing, 2018, 22(6): 1023 - 1041. [7] 范德芹, 赵学胜, 朱文泉, 等.植物物候遥感监测精度影响因素研究综述[J].地理科学进展, 2016, 35(3):304 - 319. FAN Deqin, ZHAO Xuesheng, ZHU Wenquan, et al. Review of influencing factors of accuracy of plant phenology monitoring based on remote sensing data[J]. Prog Geogr, 2016, 35(3): 304 - 319. [8] 邱琳, 林辉, 臧卓, 等.基于均值置信区间带的湿地植被高光谱特征波段选择[J].中南林业科技大学学报, 2013, 33(1):41 - 45. QIU Lin, LIN Hui, ZANG Zhuo, et al. Hyper-spectral characteristic band selection for wetland vegetation based on mean confidence interval[J]. J Cent South Univ For Technol, 2013, 33(1): 41 - 45. [9] 张甜, 廖和平, 崔林林.包络线去除的丘陵地区遥感影像阴影信息重建[J].遥感学报, 2017, 21(4):604 - 613. ZHANG Tian, LIAO Heping, CUI Linlin. Mountainous shadow information restoration based on the continuum removed[J]. J Remote Sensing, 2017, 21(4): 604 - 613. [10] 李婵, 王俊杰, 邬国锋, 等.基于叶片光谱特征的农业区域植物分类[J].深圳大学学报(理工版), 2018, 35(3):307 - 315. LI Chan, WANG Junjie, WU Guofeng, et al. Classification of agricultural plants based on leaf spectral features[J]. J Shenzhen Univ Sci Eng, 2018, 35(3): 307 - 315. [11] 陈彦兵, 况润元, 曾帅.基于高光谱数据的鄱阳湖湿地典型植被识别分析[J].人民长江, 2018, 49(20):19 - 23. CHEN Yanbing, KUANG Runyuan, ZENG Shuai. Discriminant analysis of typical vegetation species in Poyang Lake wetland based on hyperspectral data[J]. Yangtze River, 2018, 49(20): 19 - 23. [12] BREIMAN L. Random forests[J]. Mach Learn, 2001, 45(1): 5 - 32. [13] BREIMAN L. Bagging predictors[J]. Mach Learn, 1996, 24(2): 123 - 140. [14] 杨珺雯, 张锦水, 朱秀芳, 等.随机森林在高光谱遥感数据中降维与分类的应用[J].北京师范大学学报(自然科学版), 2015, 51(增刊1):82-88. YANG Junwen, ZHANG Jinshui, ZHU Xiufang, et al. Random forest applied for dimensionreduction and classification in hyperspectral data[J]. J Beijing Norm Univ Nat Sci, 2015, 51(suppl 1): 82 - 88. [15] 刘毅, 杜培军, 郑辉, 等.基于随机森林的国产小卫星遥感影像分类研究[J].测绘科学, 2012, 37(4):194 - 196. LIU Yi, DU Peijun, ZHENG Hui, et al. Classification of China small satellite remote sensing image based on random forests[J]. Sci Surv Mapp, 2012, 37(4): 194 - 196. [16] 冯伟, 郭天财, 谢迎新, 等.作物光谱分析技术及其在生长监测中的应用[J].中国农学通报, 2009, 25(23):182 - 188. FENG Wei, GUO Tiancai, XIE Yingxin, et al. Spectrum analytical technique and its applications for the crop growth detection[J]. Chin Agric Sci Bull, 2009, 25(23): 182 - 188. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.01.018
 点击查看大图
点击查看大图
计量
- 文章访问数: 4067
- HTML全文浏览量: 1140
- PDF下载量: 191
- 被引次数: 0
