




-
城市近郊区域指市辖区范围内,与城市核心区有密切交通联系并受其辐射影响的敏感活跃区域,随城市发展逐步向外推移,其空间布局、产业结构、环境景观都不同于传统乡村地区[1]。受快速城镇化进程影响,城市近郊区域的发展面临着生态环境退化、基础设施滞后及耕地荒置等问题[2]。因此,必须寻求科学的方法,引导乡村可持续发展,从而实现“产业兴旺、生态宜居、乡风文明、治理有效、生活富裕”[3]。“田园综合体”最早于2012年提出,是继“美丽乡村”“特色小镇”后乡村振兴发展的全新概念[4]。一般认为田园综合体是集循环农业、创意农业、农事体验于一体的乡村综合体[5]。使用后评价(post occupancy evaluation,POE)被认为是环境行为和社会心理学领域的延伸[6],其核心是对已完成的项目进行综合测试和实际鉴定,用“度”来衡量建成环境对人们需求的满足。作为设计反馈机制,POE不仅能为已完成项目的管理和改造提供依据,更是类似项目在设计之初时的重要参考[7]。当前中国对于田园综合体研究主要集中在发展理念、建设模式、实践探索与设计方法等方面的探讨[8-9],对于大城市近郊田园综合体景观设计的研究较少,POE研究起步较晚。本研究以大城市近郊(杭州绕城村)田园综合体为例,采用现场观察、访谈、问卷调查等方法,分析田园综合体乡村景观氛围营造、使用者行为特征等,并利用POE得到绕城村田园综合体使用后评价报告,结合绕城田园综合体景观规划设计特点和使用者的反馈信息,研究大城市近郊乡村感知与田园综合体景观规划设计之间的关系,为今后田园综合体的规划设计提供科学依据。
-
绕城村村域面积约2.6 km2,位于杭州市西湖区三墩镇,绕城高速西线以西,杭长高速以北,北接良渚文化村,南连西湖大学,紧邻紫金港交通枢纽,地理区位良好,交通便利,是典型的大城市近郊乡村。经2 a建设,初步形成集生态农业、旅游休闲、运动养生、乡村民宿、文化创意的为一体的田园综合体。于2018年5月正式对公众开放。
-
于田园综合体开放1 a后的春、夏2季,以绕城田园综合体使用者为调查对象,调查租赁菜园、生态湿地公园、景观菜地等14个景观节点。在不干涉使用者活动状态下,观察并记录其行为活动;于3月16−17日、8月17−24日发放500份问卷,收回有效问卷454份,有效率为90.8%。
-
问卷内容包括4部分。①使用者人口统计特征,如性别、年龄、居住地等;②使用者行为特征,包来访次数、使用目的、停留时间等;③田园综合体总体满意度的评价,设计自然景观、乡村景观、田园体验及配套设施等公因子及若干相关指标作为评价使用者满意度的测量因子,采用李克特5点尺度测量所有因子,对各陈述分别计分(非常满意、满意、一般、不满意、非常不满意分别记为5、4、3、2、1分);④针对各主要景观节点进行景观特征、构成内容等方面设计问卷并打分,问卷设计采用李克特量表5点尺度进行测量(很、具有、一般、中立、无,对应记为5、4、3、2、1分)。
-
采用“自由行动野外—定点观察—时间轴—定量数据”的行动观察法[10]调查使用者行为活动。于2019年3月16−17日7:00−18:00,每隔30 min观察记录1次。记录内容包括使用者行为方式和数量,并实地测量各景观节点可活动范围的面积。采用描述性统计方法统计田园综合体受访者的人口统计特征与行为特征。采用主成分萃取分析法,使用正交旋转的方差最大法对初始因子荷载矩阵进行旋转,根据较高负载的变量来命名公因子。采取行动观察法记录使用者的行为方式与人数等。调研结束后,整理调研文字及问卷数据,用SPSS 23.0对调查数据进行统计处理。
通过SPSS 23.0内部一致性系数(α)对有效问卷进行总体信度检验,α<0.35为低信度,0.35≤α<0.70为中等信度,α≥0.70为高信度[11]。分析得到本次问卷α值为0.88,说明问卷设计合理,可以使用。运用KMO(kaiser meyer olkin)和球形Bartlett检验进行因子分析的适用性[12]。KMO检验变量间的偏相关性,KMO统计量在0~1,大于0.70时因子分析效果较好,越接近1,分析效果越好[13]。经检验,KMO统计量为0.83,球形Bartlett检验发现显著性为0.00(<0.01),适宜进行因子分析。
-
由表1可见:田园综合体的使用者中,男性为53.30%,女性为46.70%。青年人(18~35岁,63.30%)与中年人(35~65岁,26.61%)为田园综合体的使用主体,年龄结构反映了场所的使用人群,是影响设施利用及布局的重要因素。
表 1 使用者属性与使用特征分析
Table 1. User attribute and usage characteristics analysis
使用特征 特征类别 比例/% 使用特征 特征类别 比例/% 使用特征 特征类别 比例/% 性别 女性 46.70 年龄 0~18 岁 8.26 居住地 本地居民 30.28 男性 53.30 18~35 岁 63.30 市区 50.46 交通方式 步行 22.02 35~65 岁 26.61 省内(杭州市外) 10.09 自行车 5.50 ≥65 1.83 外省 9.17 电瓶车 11.93 花费时间 0~1 h 75.23 来访频次 第1次 36.70 自驾车 34.86 1~3 h 18.35 每月1次 21.10 出租车 8.26 3~5 h 3.67 每天 14.68 公共交通 17.43 ≥5 h 2.75 不定期 27.52 停留时间 0~3 h 7.35 团体构成 亲人 50.43 来访目的 体验田园生活 33.70 3~6 h 57.70 朋友 40.87 观赏田园风光 32.61 6~24 h 25.69 独自 8.70 交友交流 3.80 1~3 d 7.43 亲子教育 16.85 ≥3 d 1.83 其他 13.04 从使用者居住地来看,来自杭州市区的使用者(占50.46%)较多,多数为本地居民(30.28%),到达田园综合体的方式以自驾车(34.86%)和步行(22.02%)为主,到达绕城田园综合体时间多在1 h内,说明绕城田园综合体的影响力主要还局限于本地及其周边4.5 km范围,这可能是因为绕城田园综合体开放尚不足1 a,知名度有待提升。从使用者来访频次来看,36.70%的使用者为首次来访,多数定期或不定期多次到访绕城田园综合体的使用者为绕城村周边及本地居民。从单次停留时间来看,大多使用者(90.74%)停留时长小于1 d,短期或长期度假的比例很少。从来访团体构成来看,田园综合体的使用者多与亲人(50.43%)与朋友同行(40.87%),仅少部分(8.70%)独自来访。使用者来访目的主要为体验田园生活(33.70%)、观赏田园风光(32.61%)和开展亲子教育(16.85%)。因此认为绕城村田园综合体主要服务于绕城村周边及本地居民,用作日常聚会和田园休憩,较少用于短期或长期度假。
-
由问卷数据可知:使用者对绕城田园综合体的总体满意度为3.84,整体满意度较高。主成分萃取分析法调查发现(表2):采用的16个变量公因子中因子载荷值为0.549~0.820,均大于0.50,说明本次因子分析提取的总体效果较为理想。满意度由4个公因子构成,根据各变量因子所涵盖的内容,命名为田园体验、景观风貌、人文风貌、配套服务设施。因子分析表明:“田园体验”中“农事生产教育价值”“农村生产活动”“农田、果园采摘”“农耕体验”4项载荷值较大,说明使用者较为重视农村农业教育价值与生产活动,针对使用者对未知知识、新鲜事物感到好奇的本能,设计者可以用景墙、雕塑、标牌等进行引导、展示和说明;“景观风貌”的5项因子说明使用者希望田园综合体的植物景观风貌有乡土和地域感,因此在设计中应展示植物自然茂盛生长之美,避免修剪整齐的植物,多营造空气质量佳、湿度适宜、自然生态的亲水环境;“文化景观”中“乡村住宅风貌”载荷值最大,说明乡村建筑风貌能直观体现地方文化,乡村日常的民俗活动,如庆丰收、划龙舟、点灯等,能使使用者体验乡村节庆,感受地域非物质文化的内涵和魅力;“配套服务设施”受到使用者关注较少,提示各配套服务设施应在国家标准的基础上结合乡村现状条件,重新规划设计,以提高服务性、便捷性和地域特色。
表 2 旋转后成分矩阵
Table 2. Component matrix after rotation
公因子 变量因子 因子载荷 方差贡献率% 公因子 变量因子 因子载荷 方差贡献率% 田园体验 农事生产教育价值 0.820 19.105 景观风貌 植被覆盖率 0.767 16.684 农村生产活动 0.810 植被野趣程度 0.767 农田、果园采摘 0.649 环境舒适度 0.747 农耕体验 0.618 观赏农业景观 0.609 苗木、花卉展示科普 0.603 驳岸的野趣程度 0.549 文化景观 乡村住宅风貌 0.749 14.497 配套服务设施 游客服务中心咨询服务 0.784 12.805 人文气氛感染力 0.676 停车场、厕所等的设置 0.716 乡村节日民俗活动 0.671 配套餐饮、商店等设施 0.691 -
对454份问卷进行整理,发现各节点的乡村感知得分(表3)均大于2,其中3个景观节点得分为2~3,8个景观节点得分为3~4,3个景观节点得分大于4,综合来看绕城村田园综合体整体乡村感知较强。
表 3 景观节点乡村感知评价表
Table 3. Landscape node rural perception evaluation form
景观节点 乡村感知评价 最喜爱的乡土元素 景观节点 乡村感知评价 最喜爱的乡土元素 主入口 3.29 大樟树Cinnamomum bodinieri 杨亭湾健身公园 2.33 亭子 儿童乐园 2.53 榉树Zelkova serrata 兰里公园 3.43 荷塘 文化礼堂 3.52 竹编艺术品 游船码头 3.25 水杉Metasequoia glyptostroboides 景观农田 4.21 油菜Brassica napus花 鲜花港 3.24 盆栽植物 租赁菜园 3.93 菜园 景观菜地 4.17 菜地 生态湿地公园 3.68 茅草长廊 台湾美食街 3.12 石子路 花溪公园 2.57 芦苇Phragmites australis 田野骑行绿道 4.09 茅草屋 -
参考李克特量表对各景观节点打分,结合实地访谈与观察综合发现:最受使用者喜爱的节点为景观农田、景观菜田、租赁菜园、田野骑行绿道。将景观节点的偏爱度与乡村感知度进行叠加分析(图1),发现除儿童乐园外,其他景观节点的乡村感知度与节点偏爱度呈正相关,即景观节点体现的乡村感受越高,节点就越受使用者喜爱。这些景观节点中的农田菜地等静态元素与农民田间劳作等动态元素有机结合,呈现纯真自然的乡村风貌,是人们喜欢这里的主要原因之一。2个公园(杨亭湾健身公园和花溪公园)田园感知得分为2~3,分析发现它们属于独立设计区域的社区活动公园,其景观田园感较弱。儿童乐园的乡村感与偏爱度相关性较小,二次回访调查发现:该节点使用者类型为3~6岁孩子,未能形成有效问答,以儿童家长的问卷替代;而儿童家长(非直接使用者)对于节点的偏爱度主要依赖于孩子玩耍的愉悦程度,因此具有一定主观偏差。
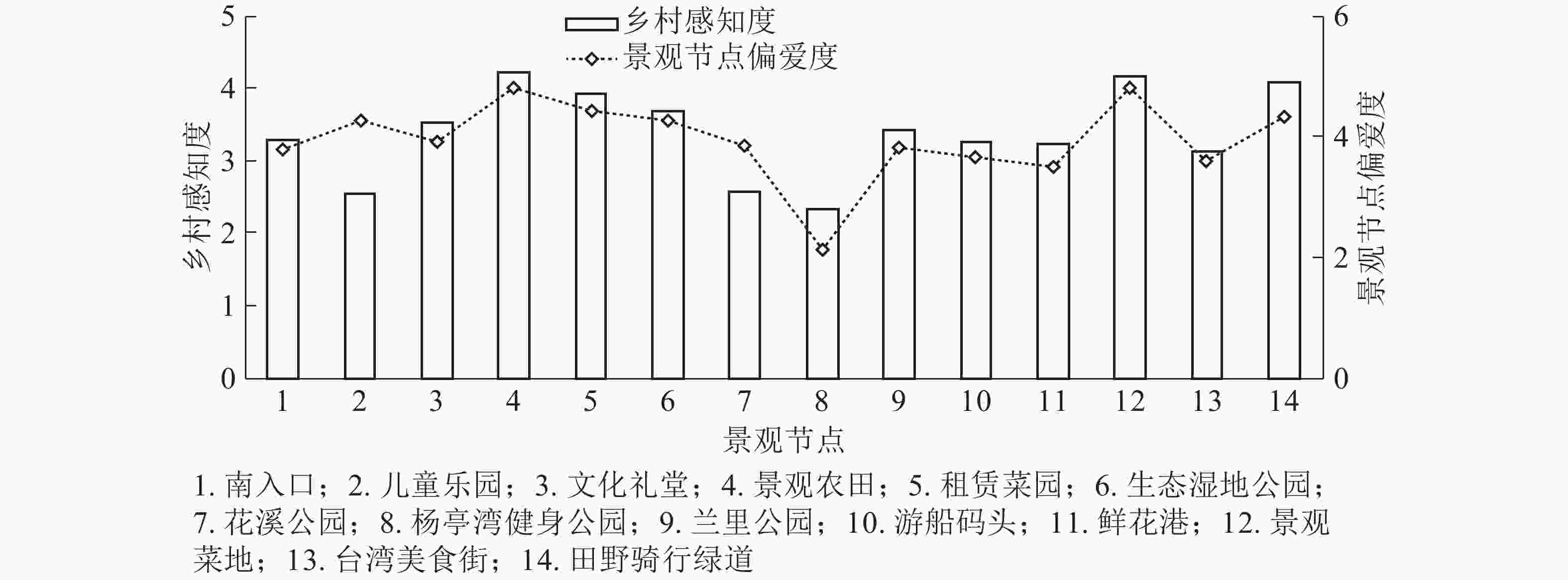
图 1 乡村感知度与景观节点偏爱度叠加分析
Figure 1. Superposition analysis of rural perception and landscape node preference
结合景观节点乡村感知情况(表3)表明:在乡村感知评价 3分以上的景观节点中,使用者最喜爱的乡土元素主要为植物景观(菜地、芦苇等)或植物材料景观(竹编艺术品、茅草长廊和茅草屋等),而田园感知得分大于4的景观节点均分布于传统农业生产区域。提示这些景观元素所构成的田园空间包含的乡村气息,唤醒了使用者对传统乡村的记忆,满足他们对乡愁的体验感知。
-
调查将使用者行为方式分成41种3种类型:①游憩行为,包括休息、交谈、野餐、露营、观景、拍照和做游戏等;②体验行为,包括耕地、拔草、种植、采摘、划船等;③移动行为,包括散步、骑行、跑步和遛狗等[14]。其中休憩行为23种,体验行为13种,移动行为5种,与农业体验相关的有采摘、耕地和种植等9种。
根据各景观节点内可活动面积、各行为方式使用者人数,计算各类行为的使用者密度。由表4可知:多数景观节点中可活动场所内体验者行为密度较其他2类行为少。结合实地考察发现,造成此种现象的原因主要为绕城田园综合体内没有可供游客进行体验性活动的设施,除团体或组织定向预约外,一般散客无法参与田园综合体内定期举办的各种田园体验类活动。
表 4 景观节点利用状况
Table 4. List of scenery node utilization situation
景观节点 节点
面积/m2使用者/
人行为
方式/种可活动
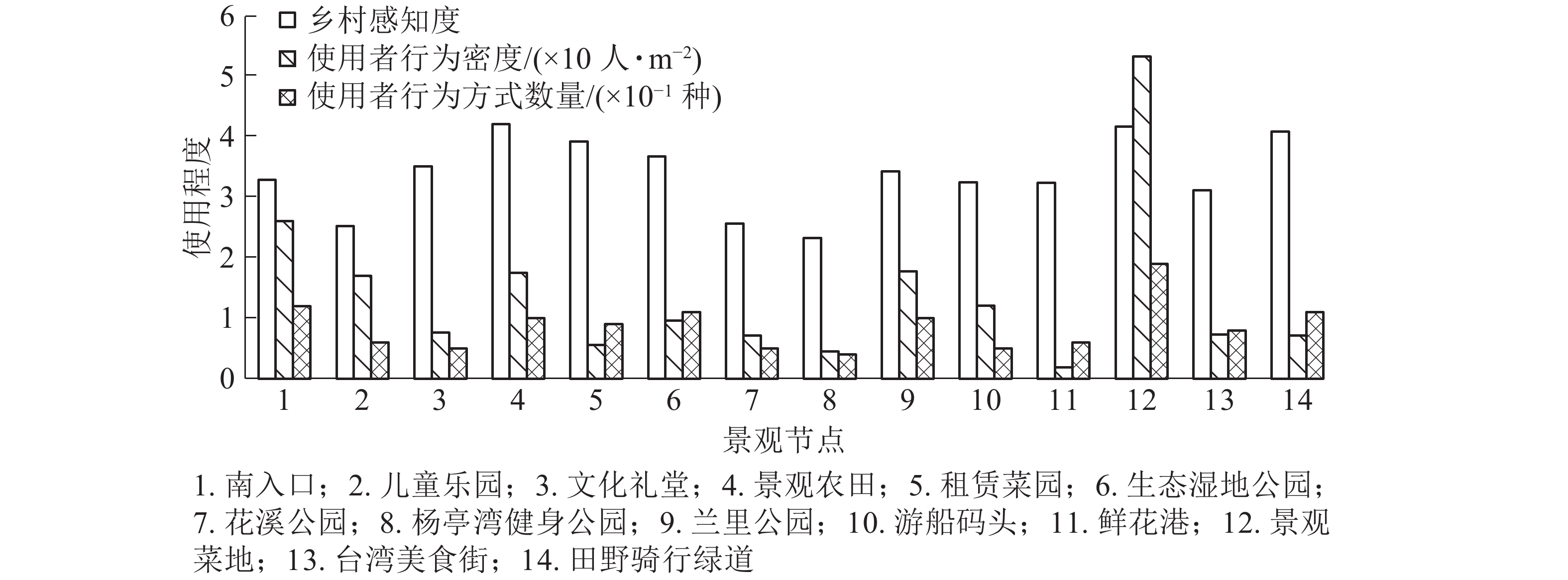
场所内容可活动范围/m2 使用者密度/(人·m−2) 游憩 移动 体验 小计 合计 主入口 5 067 304 12 可进入草坪(含景石、耐候钢板景墙) 1 660 0.148 0.025 − 0.173 0.261 水埠头 210 0.024 0.029 0.005 0.058 大樟树 166 0.024 − 0.006 0.030 儿童乐园 8 136 170 6 橡胶小广场(含设施器材、长椅) 1 057 0.060 0.083 − 0.143 0.170 橡游步道(含长凳) 710 0.007 0.020 − 0.027 文化礼堂 4 229 94 5 小广场(含景墙、竹篾、雕塑艺术品) 1 400 0.012 − − 0.012 0.076 文化展览馆 1 200 0.058 − 0.006 0.064 景观农田 124 546 934 7 可进入农田(含田埂) 11 000 0.041 0.110 0.405 0.084 0.175 景观木平台(含雕塑) 112 0.055 0.036 − 0.091 租赁菜园 49 926 237 9 可进入菜地(含景观雕塑) 5 000 0.014 0.009 0.022 0.045 0.056 水塘 662 − − 0.011 0.011 生态湿地公园 17 692 226 11 百米茅草长廊、百米南瓜长廊 1 974 0.041 0.005 − 0.046 0.095 亲水栈桥 450 0.022 0.011 0.004 0.037 水埠头(含河流、河岸、拱桥) 9 274 0.010 0.001 0.001 0.012 花溪公园 980 20 5 仿古亭(含园路) 280 0.053 0.018 − 0.071 0.071 杨亭湾健身公园 1 318 6 5 健身小广场(含健身器材、古亭) 264 0.011 − − 0.011 0.045 园路(含景墙) 86 0.011 0.023 − 0.034 兰里公园 20 395 1 033 10 石板路(含坐凳) 1 398 0.077 0.035 − 0.112 0.282 荷塘 1 195 − − 0.003 0.003 可进入花田 13 000 0.027 0.013 0.023 0.063 仿古亭廊 250 0.044 0.060 − 0.104 游船码头 3 240 85 5 水埠头(含水域) 2 278 0.023 − 0.003 0.026 0.121 小广场(含景墙、水缸、石板路) 309 0.065 − − 0.065 仿古亭廊 365 0.030 − − 0.030 鲜花港 15 618 215 6 温室大棚(含植物盆栽) 11 315 0.017 − 0.002 0.019 0.019 景观菜地 35 000 2 371 19 景观菜地 19 200 0.101 0.055 0.015 0.171 0.570 木栈道(含景观木平台) 1 127 0.093 0.037 − 0.130 说明:−表示没有该数据 对乡村感知度进行叠加分析(图2)发现:使用者行为密度与乡村感知度呈正相关,即场所的乡村感知度越高,使用者的行为数量越多。景观菜地、主入口的使用者人数较密集,参与度较高,行为活动也最丰富,结合其组成内容,发现发生在草坪上的行为方式最多,且多为团体性活动(3人及以上),如野餐、露营、打牌等,人数密度也最大,菜地上的活动类型次之,以耕种、播撒、除草等体验性活动为主。城市公园中常出现且受欢迎的可进入草坪,在近郊乡村地带依旧受到使用者的偏爱,因此田园综合体的景观规划设计中除了要保留原始乡土元素,还需考虑使用者来源和行为模式需求,适量采用现代城市化的设计手法,营造适合聚会互动、较为开敞的空间。相比而言,兰里公园、景观农田的使用者人数较密集,参与度较高,行为活动较为丰富,根据其景观构成内容发现:使用者与花田、农田的互动率最高,作为乡村特有景观,使用者在观赏农田、菜田、花田的同时也可以参与到农事劳作中,得到更加乡土的田园观感体验。总体来看,设计者在满足实用性、美观性、安全性的前提下,基于使用者的特征和行为活动需求,营造景观场所的乡村感、体验感,在追求乡村感的同时也需考虑使用者的行为习惯,是大城市近郊田园综合体区别于一般田园综合体的关键所在。
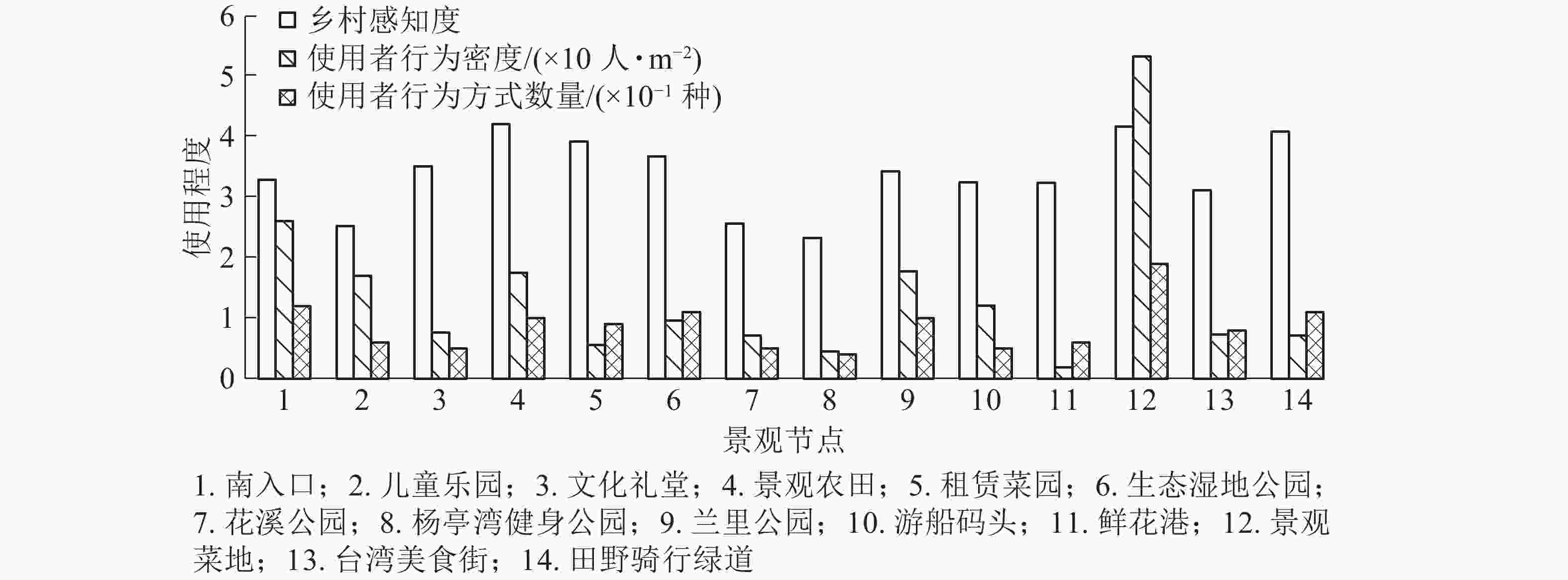
图 2 乡村感知度与使用者行为密度、行为方式数量叠加分析
Figure 2. Overlay analysis of rural perception and user population density and behavioral methods
-
本研究表明:①使用者主要来自绕城村周边和杭州市区;②使用者将绕城村田园综合体作为日常休憩场所,而非短期或长期度假;③影响使用者满意度水平的主要因子为田园体验、景观风貌、人文风貌、配套服务设施;④景观节点的乡村感知度与使用者偏爱度、使用者密度均呈正相关关系,景观规划设计不仅要保留和转译乡土元素,还需考虑使用者的来源和行为模式需求。
针对研究结果和研究区当地实际情况,提出以下几点建议:①加强宣传营销。绕城田园综合体主要服务范围为周边地区和市区,受距离、宣传力度等因素影响,远距离人群较少。因此,还需增加宣传力度与宣传途径,使得更多的使用者能参与到田园综合体中来。②完善配套设施。绕城田园综合体内餐饮、住宿购物等场所较少,应增设民宿、农家乐、商店及餐饮服务,以满足短期、长期度假或长时间使用田园综合体的使用者需求。③丰富田园体验。“田园体验”是影响满意度的重要因子,丰富有趣的田园体验是影响使用者满意度水平的重要因素。应根据绕城田园综合体实际情况定期推出丰富多彩的亲子教育、创意农业、手工制作、花卉及农产品售卖等活动,以提高绕城田园综合体体验行为的使用者密度,丰富使用者的田园体验感。④在满足使用者行为习惯的前提下提高田园综合体文化景观品质。基于使用者的特征和行为活动需求,营造集实用性、美观性、安全性于一体,且具有浓郁乡村感、丰富体验感的近郊型田园综合体景观。
Post occupancy evaluation of rural complex in the rural of large cities
-
摘要:
目的 探索田园综合体规划设计中使用者的综合要求。 方法 以杭州市西湖区绕城村田园综合体为例,以大城市近郊乡村风光和乡村生活感知营造为出发点,利用使用后评价(post occupancy evaluation,POE)方法进行调研和信息反馈,对POE结论、各景观节点使用者的空间参与度和景观偏爱度等进行相关分析。 结果 ①使用者主要来自绕城村周边和杭州市区;②绕城村田园综合体主要用于日常休憩,短期或长期度假的仅占9.26%;③影响使用者对田园综合体满意度水平的主要因子为田园体验、景观风貌、人文风貌和配套服务设施,对满意度影响较大的是田园体验和景观风貌;④景观节点的乡村感知度与使用者偏爱度、使用者密度均呈正相关关系。 结论 景观规划设计不仅要保留和转译乡土元素,还需考虑使用者的来源和行为模式需求。图2表4参14 Abstract:Objective Txplore the comprehensive requirements of users in the planning design of rural complexes. Method A case study of the rural complex in Raocheng Village of West Lake District in Hangzhou, starting with the rural scenery and the perception of rural life in the suburbs rural village of big cities, post occupancy evaluation (POE) method was used for survey and information feedback, and correlation analysis was made on POE conclusions, spatial participation and landscape preference of each landscape node in the rural surroundings complex. Result (1) The users mainly come from the villagers around Raocheng Village and residents of Hangzhou City; (2) The Raocheng village rural complex is an important place for users’ daily relaxation, however only 9.26% for short-term or long-term vacations; (3) The main factors affecting user’s satisfaction level with the rural complex are rural experience, landscape features, cultural features and supporting service facilities, and the rural experience and landscape features are the dominant ones; (4) The rural perception of landscape nodes has a positive correlation with user preference and user density. Conclusion Landscape planning design should not only retain and translate the native elements, but also need to consider the source of users and the needs of behavior patterns. [Ch, 2 fig. 4 tab. 14 ref.] -
图 1 乡村感知度与景观节点偏爱度叠加分析
Figure 1 Superposition analysis of rural perception and landscape node preference
图 2 乡村感知度与使用者行为密度、行为方式数量叠加分析
Figure 2 Overlay analysis of rural perception and user population density and behavioral methods
表 1 使用者属性与使用特征分析
Table 1. User attribute and usage characteristics analysis
使用特征 特征类别 比例/% 使用特征 特征类别 比例/% 使用特征 特征类别 比例/% 性别 女性 46.70 年龄 0~18 岁 8.26 居住地 本地居民 30.28 男性 53.30 18~35 岁 63.30 市区 50.46 交通方式 步行 22.02 35~65 岁 26.61 省内(杭州市外) 10.09 自行车 5.50 ≥65 1.83 外省 9.17 电瓶车 11.93 花费时间 0~1 h 75.23 来访频次 第1次 36.70 自驾车 34.86 1~3 h 18.35 每月1次 21.10 出租车 8.26 3~5 h 3.67 每天 14.68 公共交通 17.43 ≥5 h 2.75 不定期 27.52 停留时间 0~3 h 7.35 团体构成 亲人 50.43 来访目的 体验田园生活 33.70 3~6 h 57.70 朋友 40.87 观赏田园风光 32.61 6~24 h 25.69 独自 8.70 交友交流 3.80 1~3 d 7.43 亲子教育 16.85 ≥3 d 1.83 其他 13.04  下载: 导出CSV
下载: 导出CSV
表 2 旋转后成分矩阵
Table 2. Component matrix after rotation
公因子 变量因子 因子载荷 方差贡献率% 公因子 变量因子 因子载荷 方差贡献率% 田园体验 农事生产教育价值 0.820 19.105 景观风貌 植被覆盖率 0.767 16.684 农村生产活动 0.810 植被野趣程度 0.767 农田、果园采摘 0.649 环境舒适度 0.747 农耕体验 0.618 观赏农业景观 0.609 苗木、花卉展示科普 0.603 驳岸的野趣程度 0.549 文化景观 乡村住宅风貌 0.749 14.497 配套服务设施 游客服务中心咨询服务 0.784 12.805 人文气氛感染力 0.676 停车场、厕所等的设置 0.716 乡村节日民俗活动 0.671 配套餐饮、商店等设施 0.691
下载: 导出CSV
表 3 景观节点乡村感知评价表
Table 3. Landscape node rural perception evaluation form
景观节点 乡村感知评价 最喜爱的乡土元素 景观节点 乡村感知评价 最喜爱的乡土元素 主入口 3.29 大樟树Cinnamomum bodinieri 杨亭湾健身公园 2.33 亭子 儿童乐园 2.53 榉树Zelkova serrata 兰里公园 3.43 荷塘 文化礼堂 3.52 竹编艺术品 游船码头 3.25 水杉Metasequoia glyptostroboides 景观农田 4.21 油菜Brassica napus花 鲜花港 3.24 盆栽植物 租赁菜园 3.93 菜园 景观菜地 4.17 菜地 生态湿地公园 3.68 茅草长廊 台湾美食街 3.12 石子路 花溪公园 2.57 芦苇Phragmites australis 田野骑行绿道 4.09 茅草屋
下载: 导出CSV
表 4 景观节点利用状况
Table 4. List of scenery node utilization situation
景观节点 节点
面积/m2使用者/
人行为
方式/种可活动
场所内容可活动范围/m2 使用者密度/(人·m−2) 游憩 移动 体验 小计 合计 主入口 5 067 304 12 可进入草坪(含景石、耐候钢板景墙) 1 660 0.148 0.025 − 0.173 0.261 水埠头 210 0.024 0.029 0.005 0.058 大樟树 166 0.024 − 0.006 0.030 儿童乐园 8 136 170 6 橡胶小广场(含设施器材、长椅) 1 057 0.060 0.083 − 0.143 0.170 橡游步道(含长凳) 710 0.007 0.020 − 0.027 文化礼堂 4 229 94 5 小广场(含景墙、竹篾、雕塑艺术品) 1 400 0.012 − − 0.012 0.076 文化展览馆 1 200 0.058 − 0.006 0.064 景观农田 124 546 934 7 可进入农田(含田埂) 11 000 0.041 0.110 0.405 0.084 0.175 景观木平台(含雕塑) 112 0.055 0.036 − 0.091 租赁菜园 49 926 237 9 可进入菜地(含景观雕塑) 5 000 0.014 0.009 0.022 0.045 0.056 水塘 662 − − 0.011 0.011 生态湿地公园 17 692 226 11 百米茅草长廊、百米南瓜长廊 1 974 0.041 0.005 − 0.046 0.095 亲水栈桥 450 0.022 0.011 0.004 0.037 水埠头(含河流、河岸、拱桥) 9 274 0.010 0.001 0.001 0.012 花溪公园 980 20 5 仿古亭(含园路) 280 0.053 0.018 − 0.071 0.071 杨亭湾健身公园 1 318 6 5 健身小广场(含健身器材、古亭) 264 0.011 − − 0.011 0.045 园路(含景墙) 86 0.011 0.023 − 0.034 兰里公园 20 395 1 033 10 石板路(含坐凳) 1 398 0.077 0.035 − 0.112 0.282 荷塘 1 195 − − 0.003 0.003 可进入花田 13 000 0.027 0.013 0.023 0.063 仿古亭廊 250 0.044 0.060 − 0.104 游船码头 3 240 85 5 水埠头(含水域) 2 278 0.023 − 0.003 0.026 0.121 小广场(含景墙、水缸、石板路) 309 0.065 − − 0.065 仿古亭廊 365 0.030 − − 0.030 鲜花港 15 618 215 6 温室大棚(含植物盆栽) 11 315 0.017 − 0.002 0.019 0.019 景观菜地 35 000 2 371 19 景观菜地 19 200 0.101 0.055 0.015 0.171 0.570 木栈道(含景观木平台) 1 127 0.093 0.037 − 0.130 说明:−表示没有该数据
下载: 导出CSV
-
[1] 鄢浩. 大城市近郊区乡村居民点空间布局研究: 以重庆市南岸区为例[D]. 重庆: 重庆大学, 2016. YAN Hao. Study on Spatial Layout of Rural Residential Areas in Metropolitan Suburban Areas : A Case Study of Nan’an District in Chongqing[D]. Chong’qing: Chongqing University, 2016. [2] 徐斌, 周晓宇, 刘雷. 大城市近郊乡村更新策略: 以杭州西湖区绕城村为例[J]. 中国园林, 2018, 34(12): 63 − 67. XU Bin, ZHOU Xiaoyu, LIU Lei. Urban renewal strategy in rural areas: taking the Raocheng Village of West Lake District in Hangzhou as an example [J]. Chin Landscape Archit, 2018, 34(12): 63 − 67. [3] 习近平. 决胜全面建成小康社会夺取新时代中国特色社会主义伟大胜利: 在中国共产党第十九次全国代表大会上的报告[EB/OL]. 2017-10-18[2019-10-08]. http://news.cnr.cn/native/gd/20171027/t20171027_524003098.shtml. [4] 张毅, 王智勇. “显性”与“隐性”兼顾的田园综合体规划: 以武汉都市田园综合体为例[J]. 规划师, 2018(8): 24 − 28. ZHANG Yi, WANG Zhiyong. Rural complex planning with dominant and recessive factors: Wuhan case [J]. Planners, 2018(8): 24 − 28. [5] 中共中央、国务院. 中共中央国务院关于实施乡村振兴战略的意见[EB/OL]. 2018-01-02[2019-10-08]. http://www.audit.gov.cn/n6/n36/c119767/content.html. [6] 高喜珍, 李裕. P0E及其在公共建筑使用评价中的应用[J]. 天津理工大学学报, 2010, 26(1): 78 − 81. GAO Xizhen, LI Yu. Post occupancy evaluation and the application in the public buildings evaluating [J]. J Tianjin Univ Technol, 2010, 26(1): 78 − 81. [7] LI Xuehan. Post-Occupancy Evaluation of the Chicago Riverwalk[D]. Michigan: University of Michigan, 2018. [8] 姚杰, 程哲炀, 陈子滢, 等. 我国田园综合体文献研究进展[J]. 湖北林业科技, 2019, 48(4): 42 − 46. YAO Jie, CHENG Zheyang, CHEN Ziying, et al. Research progress of pastoral complex literature in China [J]. Hubei For Sci Technol, 2019, 48(4): 42 − 46. [9] 周敏. 新型城乡关系下田园综合体价值内涵与运行机制[J]. 规划师, 2018(8): 5 − 11. ZHOU Min. The value connotation and operation mechanism of rural complex from urban-rural relation perspective [J]. Planners, 2018(8): 5 − 11. [10] 戴菲, 章俊华. 规划设计学中的调查方法4: 行动观察法[J]. 中国园林, 2009, 25(2): 55 − 59. DAI Fei, ZHANG Junhua. The survey methods in planning and design 4: action observation [J]. Chin Landscape Archit, 2009, 25(2): 55 − 59. [11] 汪侠, 顾朝林, 梅虎. 旅游景区顾客的满意度指数模型[J]. 地理学报, 2005, 60(5): 807 − 816. WANG Xia, GU Chaolin, MEI Hu. Tourist attraction customer satisfaction index model [J]. Acta Geogr Sin, 2005, 60(5): 807 − 816. [12] 张志斌, 曹琦. 城市山体公园使用后评价: 以兰州五泉山公园为例[J]. 西北师范大学学报(自然科学版), 2010, 46(5): 114 − 119. ZHANG Zhibin, CAO Qi. Post occupancy evaluation of urban mountain park: a case study of Lanzhou Wuquan Mountain Park [J]. J Northwest Nor Univ Nat Sci, 2010, 46(5): 114 − 119. [13] 张文彤, 董伟. SPSS统计分析高级教程[M]. 北京: 高等教育出版社, 2004: 364 − 365. [14] ADINOLFI C, SUÁREZ-CÁCERES G P, CARIÑANOS P. Relation between visitors’ behavior and characteristics of green spaces in the city of Granada, south-eastern Spain [J]. Urban For Urban Greening, 2014, 13(3): 534 − 542. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.20190580
 点击查看大图
点击查看大图
计量
- 文章访问数: 2752
- HTML全文浏览量: 517
- PDF下载量: 25
- 被引次数: 0
