




-
香榧Torreya grandis‘Merrillii’属裸子植物红豆杉科Taxaceae榧树属Torreya常绿乔木,为第三纪孑遗植物[1],是榧树Torreya grandis中唯一优良栽培类型,也是中国特有的珍贵经济干果树种。由于香榧具有极高的经济价值,产值超18亿元,近年香榧产业向榧树分布的省市推广扩展,发展速度位居山地经济林之首[2]。结果母枝是着生结果枝的基枝,是香榧生长发育所需要营养的主要输送通道,对种实表现出来的性状产生很大影响。减少香榧树体的营养消耗,培育重点结果母枝,对增加结果枝的结实能力、坐果率及提高种实的品质有重要的意义。叶淑媛等[3]研究发现:香榧砧苗分枝数越少,表皮越光滑,越有利于嫁接口的愈合;刘萌萌等[4-5]研究了香榧叶片中矿质元素年周期季节性变化规律及矿质元素动态变化的关系。邬玉芬等[6]研究发现:香榧枝条黄化与钙、镁、铁元素水平低以及氮、锰元素水平过高有关;詹利云等[7]研究发现:香榧叶片长度在居群间差异显著(P<0.05)。但是,目前有关香榧结果母枝性状方面的研究还未见报道。鉴于此,本研究对香榧初结果林的结果母枝分布规律与结实关系及枝叶矿质元素间的关联性进行分析,旨在为香榧科学控梢、培养结果母枝、提高种实产量提供理论依据。
-
研究区位于浙江省杭州市临安区锦源村(29°56′~30°23′N,118°51′~119°52′E)。该区属中亚热带季风气候区南缘季风型气候,温暖湿润,日照充足,雨量充沛,四季分明。6—7月为集中降雨期,年均降水量为1 613.9 mm,降水日158 d,无霜期237 d。研究区内有原生榧树分布,为适宜栽培区。
-
供试材料为2007年12月种植的3年生香榧苗,株行距4 m×5 m,树势生长健壮,无病害的初结果香榧林。林分单株地径为9.55~14.97 cm,种植后从未进行过人为树体干预,单株均已开花结果。研究区0~20 cm土壤中全氮质量分数为1 846.19 mg·kg−1,全磷质量分数为802.85 mg·kg−1,全钾质量分数为9 816.70 mg·kg−1。每年施用复合肥2~3次,平均750~1 000 g·株−1·a−1,有机肥隔年施1次。
-
用卷尺、游标卡尺分别测定9株香榧株高、冠幅(东西向和南北向的树冠直径计算冠幅)、地径、主干茎周等表观指标。株高小于2.8 m,地径小于11.0 cm为弱势树;株高2.8~3.5 m、地径11.1~13.0 cm为中庸树;株高大于3.5 m、地径大于13.0 cm为强壮树。确定3种类型香榧树作为研究对象,每种类型重复3个。
中庸树的标准:叶色绿,主枝、延长枝生长旺盛,顶芽粗壮,树体中部结果枝数量多,结果枝比例近40%。
-
于2019年5月上旬,待混合芽、营养芽全部开展后,分单株统计3种类型树体的全部混合芽(发展成结果枝)、营养芽(发展成营养枝)数量,计算混合芽比例。
-
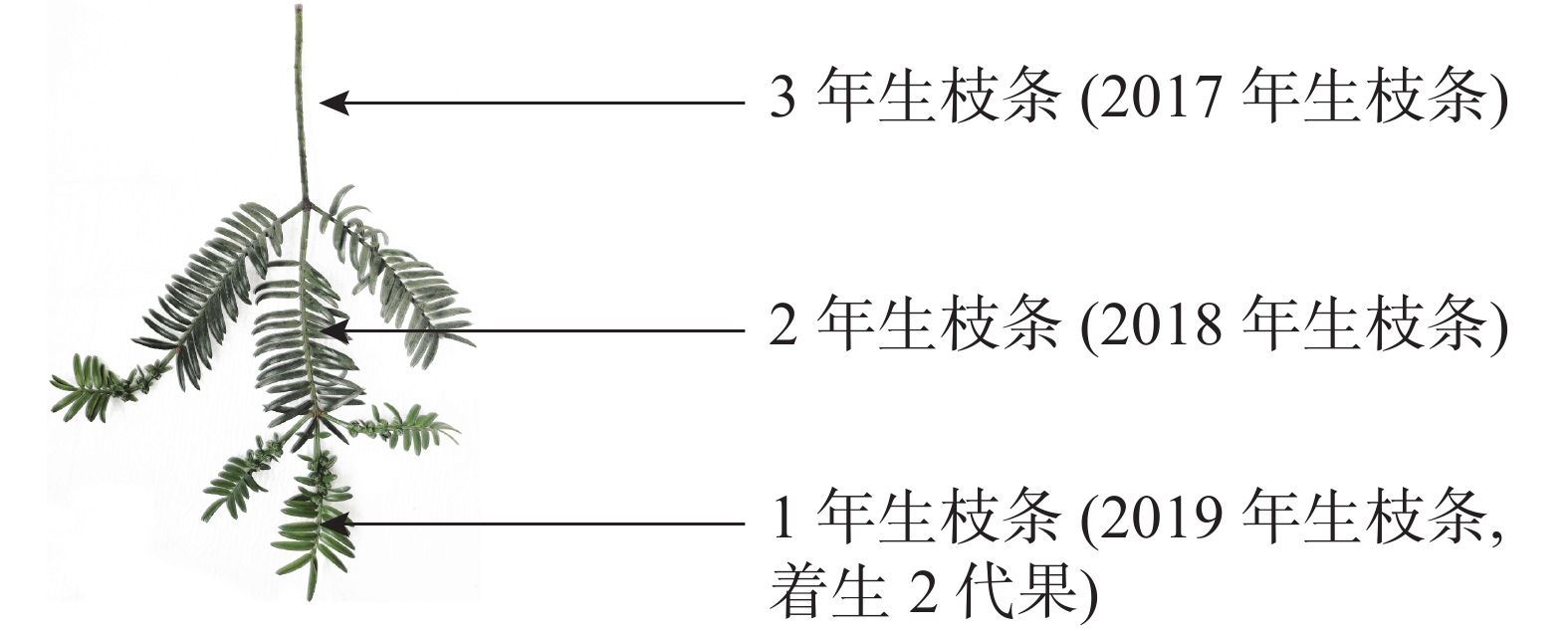
将香榧树体高度3等分,划定树体的上、中、下3个层面。于2019年3月(落果前)和6月(落果后),对3种树势树体上、中、下3个层面的东南西北方向各取5个3年生枝条,共540个枝条。图1为2017年生枝条。同时各树势植株上、中、下3个层面的东南西北方向各取1个3年生枝条,3个重复,共108个枝条,用游标卡尺测其结果母枝的粗度和长度,重复3次;根据测定结果,计算整株树结果母枝粗度和长度。
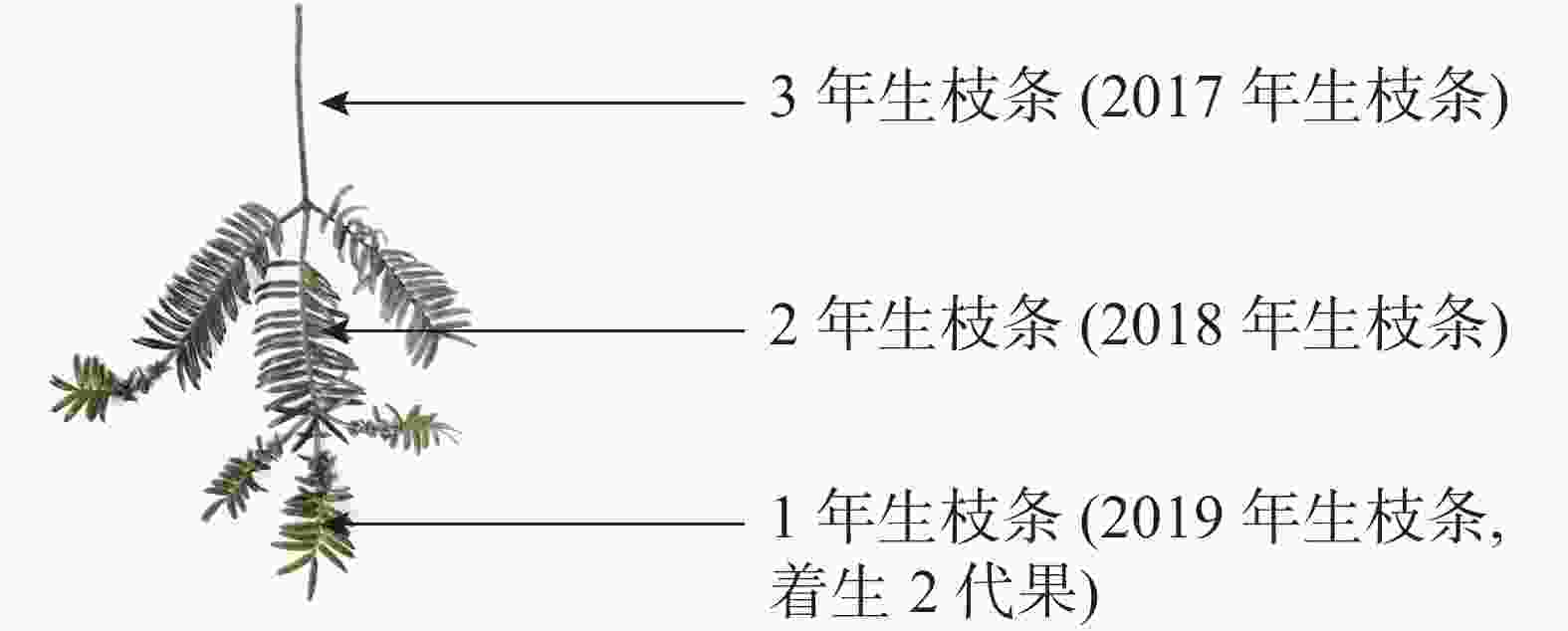
图 1 香榧3年生枝叶测量部位示意图
Figure 1. Schematic diagram of measuring parts of the 3-year-old branches and leaves of T. grandis ‘Merrillii’
-
2019年3月统计单株膨大前的2代果幼果数量,并于7月初(生理落果后),统计枝条着生的膨大果,计算全株2代果坐果率=(7月种实数量/3月种实数量)×100%。
-
将采集的样品分别用自来水、纯净水冲洗干净,再用滤纸干燥,待测。将香榧1年生枝叶去顶端和末梢,取中段枝叶进行测定。将枝条置于烘箱中,105 ℃杀青30 min,然后60 ℃左右烘干至恒量,计算枝叶样品的含水率。用植物粉碎机研磨烘干的枝条,过60目筛均匀混合。用硫酸-过氧化氢(H2SO4-H2O2)联合消煮液消煮待测样品。分别采用凯氏定氮法、钼锑抗比色法、原子吸收分光光度法[8]测定氮、磷、钾、钙、镁、锰、锌等质量分数。
-
利用SPSS 22.0开展单因素方差分析和相关分析,采用最小显著差数法(LSD法)进行平均数间多重比较,计算变异系数和回归系数,并用Excel 2007作图。
-
由表1可知:香榧单株树体营养枝数量为5 134条,结果枝数量为2 909条,结果母枝粗度2.34 mm、长度8.57 cm,膨大果数量538个,坐果率3.78%。其中冠幅、结果母枝粗度和膨大果数量均在中庸树最大,而结果母枝数量则在中庸树最少,树势越强坐果率越低,其余参数均随树势增加而增大。
表 1 9棵香榧树体基本情况与调查结果
Table 1. Basic information and investigation results of 9 T. grandis ‘Merrillii’ trees
编号 地径/cm 株高/m 冠幅/m2 营养枝
数量/条结果枝
数量/条结果母枝
粗度/mm结果母枝
长度/cm膨大果
数量/个坐果率/% 1 9.55 2.28 5.63 1 247 3 031 2.48 8.05 615 4.17 2 10.83 2.82 6.16 3 054 4 339 2.51 9.25 319 3.79 3 9.87 3.06 4.94 2 014 3 299 1.94 7.01 924 8.58 4 11.15 3.19 9.30 5 965 2 929 2.62 8.69 606 3.44 5 12.10 2.78 5.28 2 167 1 158 2.46 8.76 1 004 4.27 6 13.69 3.12 5.63 3 924 2 203 2.35 8.97 442 3.23 7 12.74 3.82 5.28 8 033 3 451 2.12 8.38 277 2.37 8 14.97 3.70 6.00 10 162 2 517 2.49 9.10 266 1.38 9 14.01 3.55 7.56 9 638 3 252 2.11 8.89 390 2.75 平均值 12.10 3.15 6.20 5 134 2 909 2.34 8.57 538 3.78 -
如表2所示:单株间的结果母枝粗度、长度、混合芽比例、膨大果数量和坐果率的性状均存在较大变幅,其中坐果率变幅最大,变异系数为53.44%;结果母枝粗度、长度变幅分别为9.85%、8.04%。表明香榧树单株间的结果母枝粗度和长度差异较小,受树势影响较小;混合芽比例、膨大果数量及坐果率在树体单株间变化较大。
表 2 结果母枝及结实性状
Table 2. Seed-bearing mother shoots and seed-bearing traits
参数 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 膨大果数量/个 坐果率/% 平均值 2.34±0.23 8.57±0.69 41.16±18.00 538.00±273.00 3.78±2.02 变异系数/% 9.85 8.04 43.74 50.81 53.44 -
如表3所示:3种树势的结果母枝数量均无显著差异(P>0.05);中庸树的结果母枝粗度最粗,与其余2种树型呈显著差异(P<0.05),强壮树与弱势树的结果母枝粗度无显著差异(P>0.05);中庸树和强壮树的结果母枝长度无显著差异(P>0.05),但两者均显著大于弱势树(P<0.05);弱势树的混合芽比例显著高于其他2种树型(P<0.05),坐果率在3种树势间均存在显著差异(P<0.05),两者从大到小依次均为弱势树、中庸树、强壮树。
表 3 树势间结果母枝及结实性状的多重比较
Table 3. A multiple comparison in seed-bearing mother shoots and seed-bearing traits among trees of various vigor
树势 结果枝数/条 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 坐果率/% 弱势树 3 556.33±690.93 a 2.31±0.32 b 8.10±1.12 b 63.88±6.27 a 5.51±2.66 a 中庸树 2 096.67±890.28 a 2.48±0.14 a 8.81±0.15 a 34.57±1.53 b 3.62±0.58 b 强壮树 3 073.33±491.97 a 2.24±0.22 b 8.79±0.37 a 25.04±5.10 b 2.07±0.69 c 说明:不同小写字母表示同一指标不同树势间差异显著(P<0.05) -
如表4所示:3种树势的结果枝数量均无显著差异(P>0.05);结果母枝粗度在树势间均为上层最粗,下层最细(P<0.05),其中中庸树的上层、中层无显著差异(P>0.05);弱势树和强壮树的结果母枝长度均是上层最长,其中弱势树中层、下层无显著差异(P>0.05),强壮树则是下层最短,中层居中(P<0.05),中庸树层面间无显著差异(P>0.05);中庸树与弱势树均是下层的坐果率最高,上层的最低(P<0.05),其中中庸树的上层、中层之间的坐果率无显著差异(P>0.05)。
表 4 层面间结果母枝及结实性状的多重比较
Table 4. A multiple comparison in seed-bearing mother shoots and seed-bearing traits among layers of a crown
树势 层面 结果枝数/条 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 坐果率/% 弱势树 上 26.00±5.00 a 2.56±0.86 a 8.54±2.89 a 56.83±27.62 b 5.29±4.00 b 中 25.33±10.26 a 1.90±0.64 b 6.79±2.45 b 71.67±33.08 ab 6.70±6.66 ab 下 26.33±3.21 a 1.55±0.82 c 5.92±3.00 b 75.83±27.97 a 8.92±9.87 a 中庸树 上 85.67±1.53 a 2.37±1.16 a 7.74±3.74 a 43.85±20.52 a 3.69±4.01 b 中 56.33±3.79 a 2.43±0.29 a 8.64±1.12 a 36.24±17.58 a 3.33±3.09 b 下 103.33±68.13 a 2.06±0.73 b 7.82±2.54 a 45.60±16.14 a 5.83±5.33 a 强壮树 上 160.33±71.11 a 2.17±084 a 8.81±3.42 a 57.40±20.95 a 1.40±1.94 a 中 67.33±3.06 a 1.81±0.95 ab 7.24±3.68 b 43.72±18.63 b 2.52±3.47 a 下 90.33±22.23 a 1.49±0.94 b 5.83±3.58 c 53.56±22.78 ab 2.03±3.61 a 说明:不同小写字母表示同一指标不同树势间差异显著(P<0.05) -
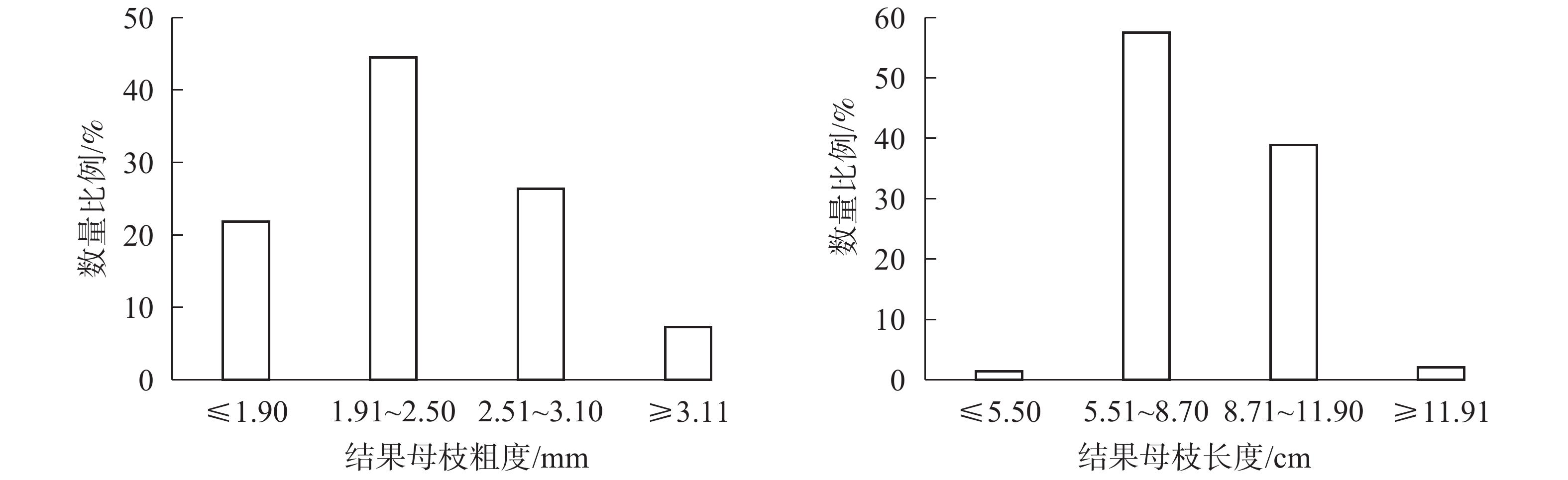
图2所示:粗度在1.91~2.50 mm的结果母枝数量最多,占44.49%;粗度在2.51~3.10 mm的结果母枝数量次之,占26.40%;粗度≤1.90 mm和≥3.11 mm的结果母枝数量最少,分别占21.83%和7.28%。长度在5.51~8.70 cm的结果母枝数量最多,占57.56%;长度在8.71~11.90的结果母枝数量次之,占38.92%;长度≤5.50和≥11.90的结果母枝数量均较少,共占3.52%。
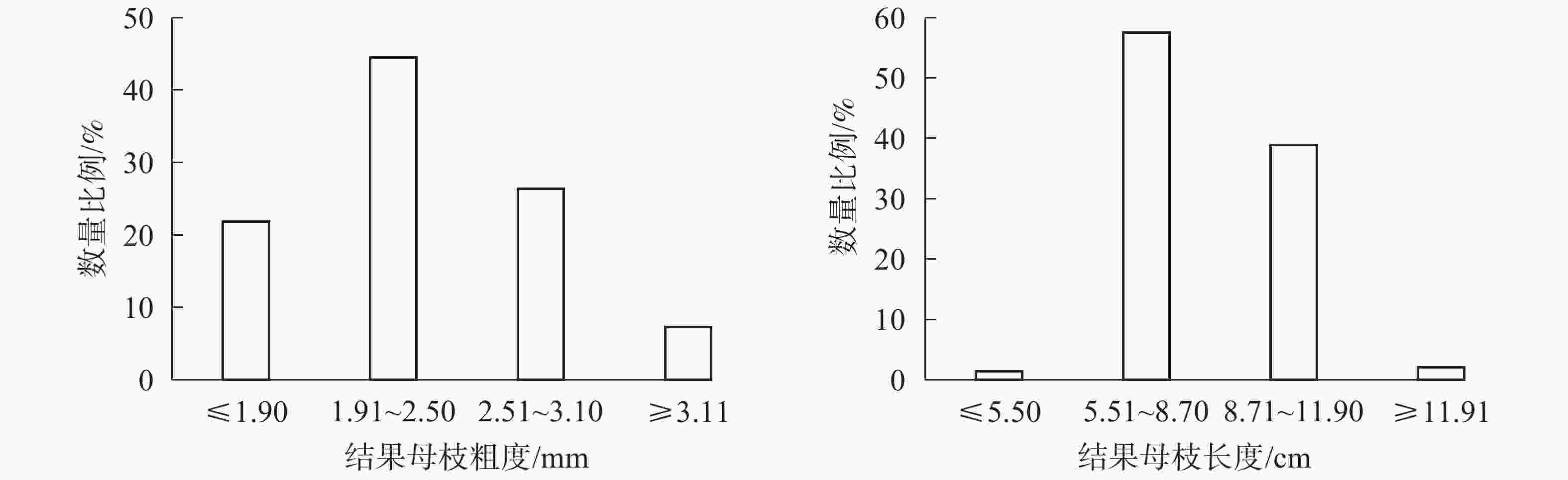
图 2 结果母枝粗度和长度分布比例
Figure 2. Distribution map of thickness and length of fruiting mother branch
-
如表5所示:结果母枝数量、2代果总数和膨大果数量在粗度1.91~2.50 mm最多,2.51~3.10 mm次之,≤1.90 mm和≥3.11 mm最少(P<0.05);此3项指标在长度为5.51~8.70 cm最多,8.70~11.90 cm次之,≤5.50 cm和≥11.90 cm最少(P<0.05);坐果率在不同粗度和长度均无显著差异(P>0.05)。
表 5 粗度、长度区间结果母枝及结实性状的多重比较
Table 5. A multiple comparison in seed-bearing mother shoots and seed-bearing traits in terms of thickness and length
指标 区间 结果母枝数/条 2代果总数/个 膨大果数量/个 坐果率/% 结果母枝粗度/mm ≤1.90 11.67±16.61 b 110.78±150.61 b 5.00±9.43 b 2.38±2.65 a 1.91~2.50 23.78±4.49 a 345.78±138.25 a 14.78±10.79 a 3.94±1.90 a 2.51~3.10 14.11±7.85 ab 209.56±124.62 a 7.78±5.65 ab 3.43±1.91 a ≥3.11 3.89±2.76 b 81.56±60.27 b 3.00±2.87 b 3.22±3.18 a 结果母枝长度/cm ≤5.50 0.78±1.99 b 8.22±21.18 b 0.44±1.01 b 1.63±3.50 a 5.51~8.70 30.89±18.20 a 424.11±242.82 a 18.22±18.19 a 3.72±1.96 a 8.71~11.90 20.89±8.13 a 294.78±158.08 a 11.78±6.42 a 4.33±2.24 a ≥11.90 1.11±1.27 b 8.33±10.77 b 0.11±0.33 b 1.11±3.33 a 说明:不同小写字母表示同一指标不同粗度、长度区间差异显著(P<0.05) -
如表6所示:结果母枝粗度、长度与混合芽比例均呈负相关,相关系数分别为−0.043、−0.552,相关性均不显著;同样,结果母枝粗度及长度与营养芽比例相关性也不显著。结果母枝粗度及长度与2代果坐果率均呈负相关,其中结果母枝长度与2代果坐果率的相关系数为−0.825,呈极显著负相关(P<0.01)。结果母枝粗度与2代果坐果率相关系数为−0.461,无显著相关;混合芽比例与坐果率的相关系数为0.680,呈显著正相关(P<0.05)。
表 6 结果母枝与结实性状的相关性分析
Table 6. Correlation between seed-bearing mother shoots and seed-bearing traits
性状 相关系数 回归方程 结果母枝粗度(x)与混合芽比例(y) −0.043 y=−2.665 3x+47.409 结果母枝长度(x)与混合芽比例(y) −0.552 y=−0.144 3x+1.647 7 结果母枝粗度(x)与坐果率(y) −0.461 y=−3.956 3x+13.045 结果母枝长度(x)与坐果率(y) −0.825** y=−2.422 2x+24.525 混合芽比例(x)与坐果率(y) 0.680* y=0.076 4x+0.632 5 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
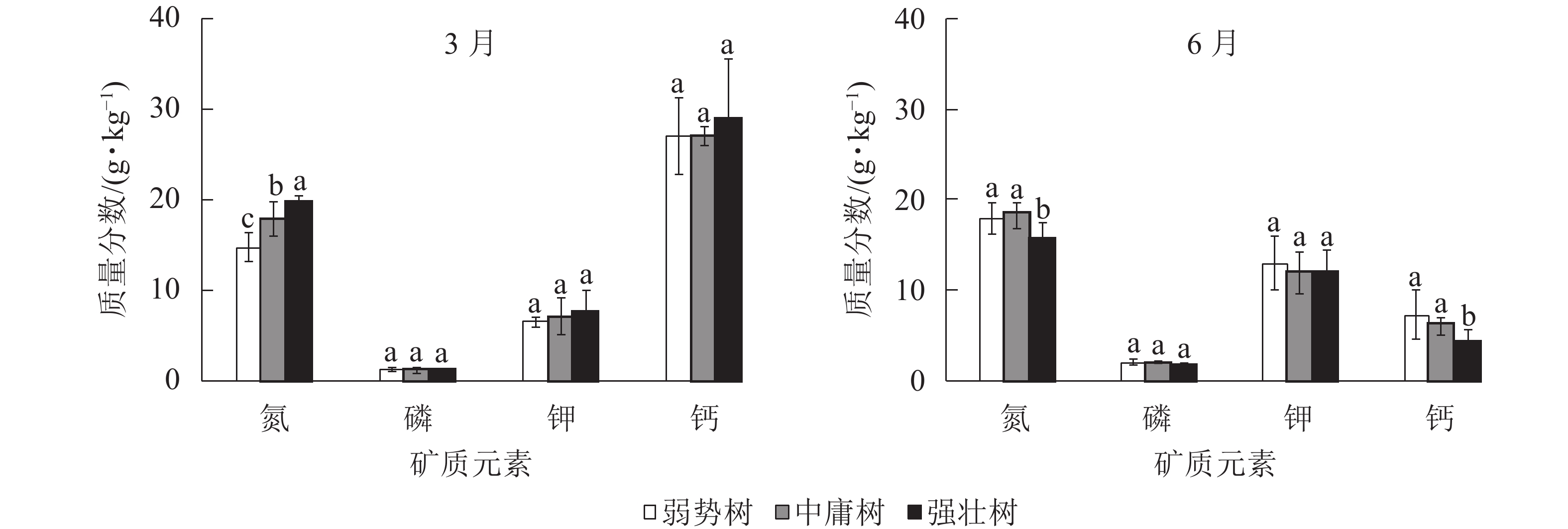
图3所示:3月香榧枝叶的氮质量分数从大到小依次为强壮树、中庸树、弱势树(P<0.05);而6月中庸树、弱势树氮质量分数极显著高于强壮树(P<0.05),弱势树与中庸树间无显著差异(P>0.05);3和6月的磷、钾质量分数在树势间均无显著差异(P>0.05),但磷、钾质量分数均为6月大于3月;3月的钙质量分数无显著差异(P>0.05),6月强壮树钙质量分数显著低于另外2种树势(P<0.05),弱势树与中庸树间没有显著差异(P>0.05)。其中3月的强壮树、中庸树、弱势树间的磷、钾质量分数接近,6月强壮树磷最低,中庸树钾最低;3种树势的钙质量分数均为3月大于6月,3月3种树势间钙质量分数接近,6月强壮树最低。
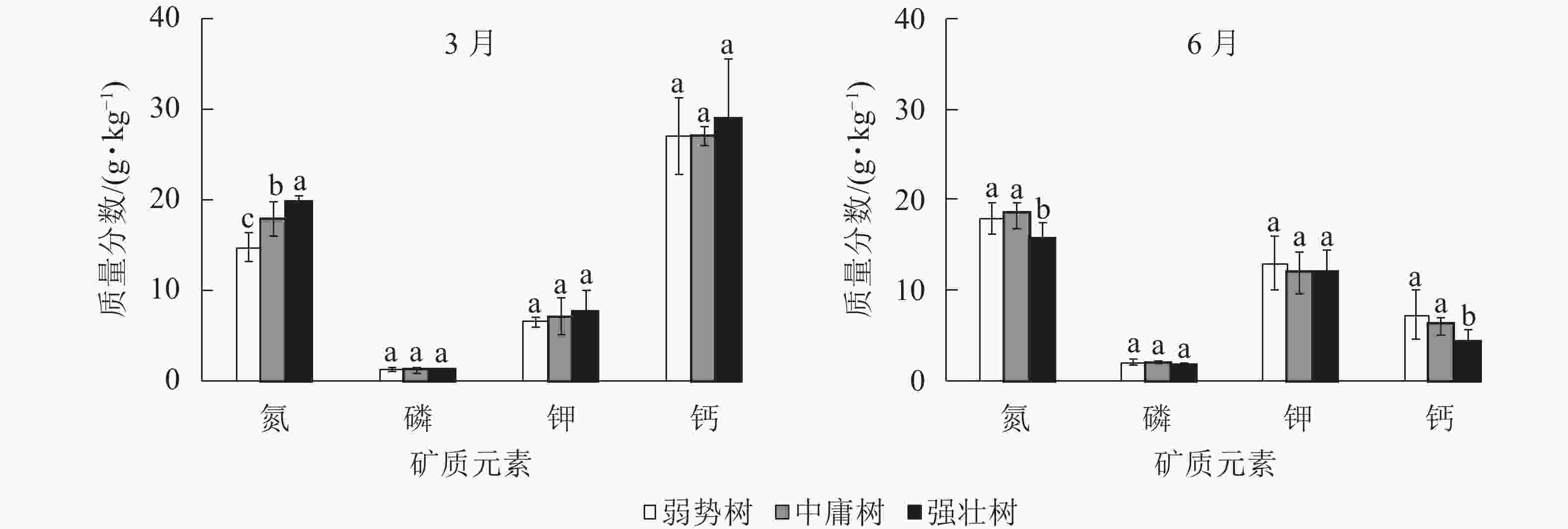
图 3 3和6月枝叶矿质元素之间的多重比较
Figure 3. Multiple comparison of mineral elements in branches and leaves of T. grandis ‘Merrillii’ in March and June
-
如表7所示:3月结果母枝粗度与钙呈显著负相关(P<0.05);混合芽比例与氮呈极显著负相关(P<0.01),与锌、锰呈显著负相关(P<0.05);其余各指标之间无显著相关。6月结果母枝粗度与氮呈极显著正相关(P<0.01),与磷呈显著正相关(P<0.05);结果母枝长度与钙呈显著负相关(P<0.05),与铁呈显著正相关(P<0.05);混合芽比例与锰呈显著的负相关(P<0.05);坐果率与钙、铁分别呈极显著正、负相关(P<0.01)。
表 7 3和6月结果母枝、结实性状与枝叶矿质元素相关性分析
Table 7. Correlation between seed-bearing mother shoots and seed-bearing traits and mineral elements in March and June
月份 参数 氮 磷 钾 钙 镁 铁 锌 锰 钠 3 结果母枝粗度 −0.009 −0.267 0.106 −0.687* 0.529 0.483 0.160 −0.045 0.400 结果母枝长度 0.133 −0.474 0.383 −0.465 −0.048 −0.091 0.326 0.243 0.240 混合芽比例 −0.804** 0.006 −0.319 −0.131 0.490 0.152 −0.756* −0.697* −0.351 坐果率 −0.470 0.251 −0.393 0.241 0.199 −0.200 −0.525 −0.485 −0.238 6 结果母枝粗度 0.847** 0.728* 0.434 −0.347 0.459 0.470 0.408 0.082 0.180 结果母枝长度 0.407 0.589 0.606 −0.791* 0.048 0.766* 0.448 0.340 0.632 混合芽比例 0.277 0.318 0.097 0.573 0.562 −0.528 −0.416 −0.774* −0.521 坐果率 −0.039 −0.205 −0.364 0.859** 0.060 −0.866** −0.551 −0.582 −0.543 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
如表8所示:营养芽与混合芽比例呈极显著负相关(P<0.01),与膨大果数量及坐果率呈显著负相关(P<0.05),相关系数分别为−0.830、−0.673和−0.692。
表 8 香榧营养生长与生殖生长相关性分析
Table 8. Analysis of correlation between vegetative growth and reproductive growth
性状 相关系数 回归方程 营养芽(x)与混合芽比例(y) −0.830** y=−0.004 4x+63.535 营养芽(x)与膨大果数量(y) −0.673* y=−0.053 7x+813.57 营养芽(x)与坐果率(y) −0.692* y=−0.000 4x+5.869 3 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
香榧结果母枝是着生结果枝的基枝,是香榧种实生长发育所需营养的主要供应源,因此结果母枝的生长状况对香榧种实性状及产量有重要影响[9-10]。变异系数的大小反映了性状离散程度的高低,可衡量性状间的变异程度[10]。本研究发现:未在人为干预情况下,结果母枝粗度与长度在单株间较为稳定,而结果枝比例、膨大果数量及坐果率的性状特征可能受树势影响较大,导致其变异程度较大。
本研究结果显示:结果母枝数量与树势、层面均无密切关系;而结果母枝粗度及长度、混合芽比例及坐果率受树势的影响较为明显。中庸树结果母枝粗度、长度均最大,混合芽比例及坐果率在中庸树中表现良好,表明初结果林分香榧树的树势越强,营养生长越强,反之,则生殖生长强于营养生长。中庸树的生殖生长和营养生长较为平衡,可根据生产实际需求,以中庸树为培育目标。同时,结果母枝粗度自下而上均越来越粗;长度在强壮树、弱势树中也是自下而上越来越长;混合芽比例弱势树为下层最高,强壮树为上层最高;坐果率在弱势树和中庸树中均是下层最高,表明树势的强弱、层面的分布均能影响结果母枝的粗度和长度。随着层面的增高,营养生长逐渐增强,这与光照强度由下向上逐渐增加的规律一致[11]。层面越低生殖生长越明显,可能与香榧树下垂枝有关。
结果母枝粗度、长度分布情况与结果母枝数量、2代果总数和膨大果数量(结果量)呈一致性,均在粗度为1.91~2.50 mm最多,长度为5.51~8.70 cm最多,坐果率不受结果母枝粗度和长度的影响。由此可见,初结果林分香榧树的结果性状在不同粗度、长度的分布状况呈现一定的规律,枝粗和枝长是影响结果性状指标的重要因素之一,是能否迅速提高早期产量的关键,也是衡量结果母枝坐果能力的重要指标,这与陈霞等[12]的研究相一致。本研究相关分析显示:与产量相关的坐果率和混合芽比例等参数受结果母枝粗度、长度的制约,结果母枝粗度越粗,长度越大,香榧树的坐果率就越低,特别是枝越长坐果率越低。结果母枝粗度、长度对混合芽、营养芽的生长也有一定的影响,但影响不明显;混合芽比例越高坐果率越高。同时,香榧初结果树的营养生长制约着生殖生长,营养生长越旺盛,越不利于香榧果实的膨大和产量的提高。因此,培养中庸树型,保持营养生长与生殖生长平衡,对提高香榧种实产量有重要意义。
氮、磷、钾是香榧种子生长发育所必需的矿质元素,对香榧苗木生长的影响作用从大到小依次为氮、钾、磷[13]。本研究表明:3和6月的氮在树势间均呈极显著差异,氮质量分数在3月从大到小依次为强壮树、中庸树、弱势树,但在6月强壮树氮质量分数最低,且3月平均值高于6月,可能与香榧种子开始生长膨大,叶片中的氮元素优先供应种子的生长发育有关。本研究结果与刘萌萌等[5]的结论相一致。而6月强壮树氮质量分数下降的幅度高于另外2种树势类型,可能与叶片中的氮元素在供应种子生长和营养枝旺盛生长的同时,消耗了大量氮元素有关,这与其他物种果实发育的研究结论一致[14]。钾能促进果实的增大和质量的增加,尤以在氮营养水平较高时,钾多则效果更为明显,因为钾可提高原生质活性,促进糖的转运,增加果实干质量[15]。磷、钾质量分数不受香榧树势影响,但6月的磷、钾质量分数均高于3月,这可能是由于6月叶片中的磷质量分数相对稳定,钾质量分数有所增加,香榧速生期和种子内部充实期,是对氮、钾需求量最大的时期[5]。香榧花芽分化受到氮、锌、锰的影响,摄入过多的氮、锌、锰会制约混合芽数量增加,这种现象在杏Armeniaca vulgaris树中也存在[16]。3、6月镁与混合芽比例无显著相关,可能是由于该时期枝叶中的镁并不是香榧花芽中镁的供应源[17],表明树体矿质元素随时间季节存在动态变化[5],因此,施肥时要考虑元素间的相互作用。
钙是果胶质中重要物质——果胶酸钙的主要组成物质,也是细胞壁构成成分,果实中钙的高低,影响果实硬度和耐储性[18]。本研究显示:6月铁质量分数的提高促进了结果母枝长度的生长,但抑制了香榧种子的膨大;而钙与枝叶长度呈显著负相关,可能是由于随着香榧种实膨大、细胞分裂迅速,大量的钙元素流向种子,钙元素促进了种子的发育。但当种子由细胞分裂转入增大时,正值新梢旺盛生长期,从而加剧了对钙的竞争。由此可见,种实膨大期适时适度增施钙肥,可以提高香榧坐果率。钙与硼对悬浮细胞的生长有显著的交互作用,增加钙可以部分减缓缺硼对细胞生长的影响[19],植物在缺硼状态下,会触发钙离子的信号转导途径,影响植物体某些蛋白的合成,从而导致植物体的代谢变化[20]。硼、钙与腐植酸的配合施用能够显著提高番茄的株高、果数、单果质量及产量[21]。因此,后续还需开展香榧初结果林中不同钙肥水平对硼等其他元素的响应研究。
-
不同树势和冠层部位的结果母枝类型、结实性状及矿质营养元素特征差异显著。生产实践中,平衡氮肥和钙肥的施用,培养中庸树型,控制香榧结果母枝长度和树体的高度,将树体修剪成开心型,控制结果母枝粗度在1.91~3.10 mm、长度5.51~11.90 cm,有利于提高香榧坐果率和混合芽的比例,对实现香榧丰产、稳产具有重要意义。
Relationship between character changing and seed-bearing capacity of initial seed-bearing mother shoots in Torreya grandis ‘Merrillii’
-
摘要:
目的 探究香榧Torreya grandis‘Merrillii’初结果林分树体结果母枝性状变化规律与结实能力的关系,为科学控梢、培养结果母枝及提高种实产量提供依据。 方法 以浙江省杭州市临安区锦源村香榧初结果林为研究对象,分析树势间和层面间的结果母枝性状、结果母枝粗度及长度区间分布、枝叶矿质元素质量分数与结果指标的关系。 结果 中庸树的结果母枝粗度最大,其结果母枝长度显著高于弱势树(P<0.05);不同树势树体的混合芽比例和坐果率差异显著(P<0.05),弱势树的混合芽比例和坐果率较高,强壮树的混合芽比例和坐果率较低;树冠上层的结果母枝粗度和长度最大,下层最小(P<0.05),混合芽比例和坐果率在不同冠层部位间差异显著(P<0.05),而且在不同树势的树体间表现不同,其中坐果率在中庸树和弱势树体的树冠下层最高;结果母枝粗度1.91 ~ 2.50 mm、长度5.51 ~ 8.70 cm的2代果总数和膨大果数量最多,且坐果率与结果母枝长度呈显著负相关(P<0.01)。此外,在枝叶矿质营养特征方面,不同树势树体枝叶的氮质量分数之间差异显著(P<0.05),6月中庸树势树体枝叶的氮质量分数最高;钙质量分数与结果母枝长度呈显著负相关,与坐果率呈显著正相关(P<0.05)。 结论 生产实践中,培养中庸树型,适当控制结果母枝长度和树体高度,有利于香榧坐果率、混合芽比例的提高;保留粗度1.91~3.10 mm、长度5.51~11.90 cm的结果母枝,对提高香榧坐果率、产量有重要意义;施钙肥,对促进香榧种子生长和提高坐果率有重要作用。图3表8参21 Abstract:Objective With an exploration of the relationship between the character changing patterns of seed-bearing mother shoots and the seed-bearing capacity of initial seed bearers in Torreya grandis ‘Merrillii’, this study is aimed to provide theoretical and technical rationale for the control of shooting, the culture of seed-bearing mother shoots, and the promotion of seed yield. Method With the T. grandis ‘Merrillii’ trees located in Jinyuan village of Lin’an District, Hangzhou City of Zhejiang Province selected as the subjects, an analysis was conducted of the characteristics of seed-bearing mother shoots either among trees of various levels of vigor or among different layers of a crown, the distribution of seed-bearing mother shoots in terms of shoot length and thickness, and the relationship between the content of mineral elements in shoots and leaves and seed-bearing parameters. Result The seed-bearing mother shoots from moderately vigorous trees were the thickest and significantly longer than those from the trees of weak vigor (P<0.05). Trees of various levels of vigor were significantly different in the percentage of mixed buds and seed-setting rate (P<0.05), with the percentage of mixed buds and seed-setting rate of weak trees being higher than those of vigorous trees. The seed-bearing mother shoots in the upper layer of the crown were the thickest and longest while those in the lower layer of the crown were the thinnest and shortest (P<0.05). The proportion of mixed buds and seed-setting rate were significantly different among layers of the crown (P<0.05) and varied with the vigor of trees, with the seed-setting rate being the highest in the lower layer of the crown of the moderately vigorous and weak trees. The mother shoots with a thickness of 1.91−2.50 mm and a length of 5.51−8.70 cm took up the highest percentage in new seeds and swollen seeds while the seed-setting rate was significantly negatively correlated with the length of the mother shoot (P<0.01). In terms of mineral nutrition, the nitrogen content in the shoots and leaves was significantly different among trees of various levels of vigor (P<0.05), with the percentage of nitrogen in the shoots and leaves sampled in June from the trees of moderate vigor being the highest whereas the Ca content was significantly positively correlated with the length of mother shoots, but significantly negatively correlated with the seed-setting rate (P<0.05). Conclusion In the process of production, the cultivation of moderately growing trees with a proper control over the length of mother shoots and the tree height would be conducive to the increase in seed setting rate and mixed bud proportion. On the other hand, it is of great significance to retain the mother shoots with a thickness of 1.91−3.10 mm and a length of 5.51−11.90 cm to achieve the promotion of seed-setting rate and yield. Also, calcium fertilizer plays an important role in promoting seed growth and improving the seed-setting rate in T. grandis ‘Merrillii’. [Ch, 3 fig. 8 tab. 21 ref.] -
图 1 香榧3年生枝叶测量部位示意图
Figure 1 Schematic diagram of measuring parts of the 3-year-old branches and leaves of T. grandis ‘Merrillii’
图 2 结果母枝粗度和长度分布比例
Figure 2 Distribution map of thickness and length of fruiting mother branch
图 3 3和6月枝叶矿质元素之间的多重比较
不同小写字母表示差异显著(P<0.05)
Figure 3 Multiple comparison of mineral elements in branches and leaves of T. grandis ‘Merrillii’ in March and June
表 1 9棵香榧树体基本情况与调查结果
Table 1. Basic information and investigation results of 9 T. grandis ‘Merrillii’ trees
编号 地径/cm 株高/m 冠幅/m2 营养枝
数量/条结果枝
数量/条结果母枝
粗度/mm结果母枝
长度/cm膨大果
数量/个坐果率/% 1 9.55 2.28 5.63 1 247 3 031 2.48 8.05 615 4.17 2 10.83 2.82 6.16 3 054 4 339 2.51 9.25 319 3.79 3 9.87 3.06 4.94 2 014 3 299 1.94 7.01 924 8.58 4 11.15 3.19 9.30 5 965 2 929 2.62 8.69 606 3.44 5 12.10 2.78 5.28 2 167 1 158 2.46 8.76 1 004 4.27 6 13.69 3.12 5.63 3 924 2 203 2.35 8.97 442 3.23 7 12.74 3.82 5.28 8 033 3 451 2.12 8.38 277 2.37 8 14.97 3.70 6.00 10 162 2 517 2.49 9.10 266 1.38 9 14.01 3.55 7.56 9 638 3 252 2.11 8.89 390 2.75 平均值 12.10 3.15 6.20 5 134 2 909 2.34 8.57 538 3.78  下载: 导出CSV
下载: 导出CSV
表 2 结果母枝及结实性状
Table 2. Seed-bearing mother shoots and seed-bearing traits
参数 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 膨大果数量/个 坐果率/% 平均值 2.34±0.23 8.57±0.69 41.16±18.00 538.00±273.00 3.78±2.02 变异系数/% 9.85 8.04 43.74 50.81 53.44
下载: 导出CSV
表 3 树势间结果母枝及结实性状的多重比较
Table 3. A multiple comparison in seed-bearing mother shoots and seed-bearing traits among trees of various vigor
树势 结果枝数/条 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 坐果率/% 弱势树 3 556.33±690.93 a 2.31±0.32 b 8.10±1.12 b 63.88±6.27 a 5.51±2.66 a 中庸树 2 096.67±890.28 a 2.48±0.14 a 8.81±0.15 a 34.57±1.53 b 3.62±0.58 b 强壮树 3 073.33±491.97 a 2.24±0.22 b 8.79±0.37 a 25.04±5.10 b 2.07±0.69 c 说明:不同小写字母表示同一指标不同树势间差异显著(P<0.05)
下载: 导出CSV
表 4 层面间结果母枝及结实性状的多重比较
Table 4. A multiple comparison in seed-bearing mother shoots and seed-bearing traits among layers of a crown
树势 层面 结果枝数/条 结果母枝粗度/mm 结果母枝长度/cm 混合芽比例/% 坐果率/% 弱势树 上 26.00±5.00 a 2.56±0.86 a 8.54±2.89 a 56.83±27.62 b 5.29±4.00 b 中 25.33±10.26 a 1.90±0.64 b 6.79±2.45 b 71.67±33.08 ab 6.70±6.66 ab 下 26.33±3.21 a 1.55±0.82 c 5.92±3.00 b 75.83±27.97 a 8.92±9.87 a 中庸树 上 85.67±1.53 a 2.37±1.16 a 7.74±3.74 a 43.85±20.52 a 3.69±4.01 b 中 56.33±3.79 a 2.43±0.29 a 8.64±1.12 a 36.24±17.58 a 3.33±3.09 b 下 103.33±68.13 a 2.06±0.73 b 7.82±2.54 a 45.60±16.14 a 5.83±5.33 a 强壮树 上 160.33±71.11 a 2.17±084 a 8.81±3.42 a 57.40±20.95 a 1.40±1.94 a 中 67.33±3.06 a 1.81±0.95 ab 7.24±3.68 b 43.72±18.63 b 2.52±3.47 a 下 90.33±22.23 a 1.49±0.94 b 5.83±3.58 c 53.56±22.78 ab 2.03±3.61 a 说明:不同小写字母表示同一指标不同树势间差异显著(P<0.05)
下载: 导出CSV
表 5 粗度、长度区间结果母枝及结实性状的多重比较
Table 5. A multiple comparison in seed-bearing mother shoots and seed-bearing traits in terms of thickness and length
指标 区间 结果母枝数/条 2代果总数/个 膨大果数量/个 坐果率/% 结果母枝粗度/mm ≤1.90 11.67±16.61 b 110.78±150.61 b 5.00±9.43 b 2.38±2.65 a 1.91~2.50 23.78±4.49 a 345.78±138.25 a 14.78±10.79 a 3.94±1.90 a 2.51~3.10 14.11±7.85 ab 209.56±124.62 a 7.78±5.65 ab 3.43±1.91 a ≥3.11 3.89±2.76 b 81.56±60.27 b 3.00±2.87 b 3.22±3.18 a 结果母枝长度/cm ≤5.50 0.78±1.99 b 8.22±21.18 b 0.44±1.01 b 1.63±3.50 a 5.51~8.70 30.89±18.20 a 424.11±242.82 a 18.22±18.19 a 3.72±1.96 a 8.71~11.90 20.89±8.13 a 294.78±158.08 a 11.78±6.42 a 4.33±2.24 a ≥11.90 1.11±1.27 b 8.33±10.77 b 0.11±0.33 b 1.11±3.33 a 说明:不同小写字母表示同一指标不同粗度、长度区间差异显著(P<0.05)
下载: 导出CSV
表 6 结果母枝与结实性状的相关性分析
Table 6. Correlation between seed-bearing mother shoots and seed-bearing traits
性状 相关系数 回归方程 结果母枝粗度(x)与混合芽比例(y) −0.043 y=−2.665 3x+47.409 结果母枝长度(x)与混合芽比例(y) −0.552 y=−0.144 3x+1.647 7 结果母枝粗度(x)与坐果率(y) −0.461 y=−3.956 3x+13.045 结果母枝长度(x)与坐果率(y) −0.825** y=−2.422 2x+24.525 混合芽比例(x)与坐果率(y) 0.680* y=0.076 4x+0.632 5 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 7 3和6月结果母枝、结实性状与枝叶矿质元素相关性分析
Table 7. Correlation between seed-bearing mother shoots and seed-bearing traits and mineral elements in March and June
月份 参数 氮 磷 钾 钙 镁 铁 锌 锰 钠 3 结果母枝粗度 −0.009 −0.267 0.106 −0.687* 0.529 0.483 0.160 −0.045 0.400 结果母枝长度 0.133 −0.474 0.383 −0.465 −0.048 −0.091 0.326 0.243 0.240 混合芽比例 −0.804** 0.006 −0.319 −0.131 0.490 0.152 −0.756* −0.697* −0.351 坐果率 −0.470 0.251 −0.393 0.241 0.199 −0.200 −0.525 −0.485 −0.238 6 结果母枝粗度 0.847** 0.728* 0.434 −0.347 0.459 0.470 0.408 0.082 0.180 结果母枝长度 0.407 0.589 0.606 −0.791* 0.048 0.766* 0.448 0.340 0.632 混合芽比例 0.277 0.318 0.097 0.573 0.562 −0.528 −0.416 −0.774* −0.521 坐果率 −0.039 −0.205 −0.364 0.859** 0.060 −0.866** −0.551 −0.582 −0.543 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 8 香榧营养生长与生殖生长相关性分析
Table 8. Analysis of correlation between vegetative growth and reproductive growth
性状 相关系数 回归方程 营养芽(x)与混合芽比例(y) −0.830** y=−0.004 4x+63.535 营养芽(x)与膨大果数量(y) −0.673* y=−0.053 7x+813.57 营养芽(x)与坐果率(y) −0.692* y=−0.000 4x+5.869 3 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
-
[1] 武栋. 安徽徽州榧树种群内种子理化性质的分析[D]. 杭州: 浙江农林大学, 2012. WU Dong. Analysis of Physico-Chemical Properties of the Seeds from a Natural Torreya Population in Huizhou, Anhui[D]. Hangzhou: Zhejiang A&F University, 2012. [2] 何祯, 王宗星, 张骏, 等. 浙江省香榧产业发展现状与对策[J]. 浙江农业科学, 2020, 61(7): 1345 − 1347. HE Zhen, WANG Zongxing, ZHANG Jun, et al. Present situation and countermeasures of Torreya grandis ‘Merrillii’ industry development in Zhejiang Province [J]. J Zhejiang Agric Sci, 2020, 61(7): 1345 − 1347. [3] 叶淑媛, 董雷鸣, 董昂, 等. 榧树半同胞子代幼苗生长性状的遗传参数估算[J]. 浙江农林大学学报, 2020, 37(4): 817 − 822. YE Shuyuan, DONG Leiming, DONG Ang, et al. Estimation of genetic parameters for juvenile growth of half-sib seedlings of Torreya grandis [J]. J Zhejiang A&F Univ, 2020, 37(4): 817 − 822. [4] 刘萌萌, 曾燕如, 江建斌, 等. 香榧生长期叶片和种子中矿质元素动态变化研究[J]. 浙江农林大学学报, 2014, 31(5): 724 − 729. LIU Mengmeng, ZENG Yanru, JIANG Jianbin, et al. Mineral elements in leaves and seeds of Torreya grandis ‘Merrillii’ during seed development [J]. J Zhejiang A&F Univ, 2014, 31(5): 724 − 729. [5] 刘萌萌, 曾燕如, 江建斌, 等. 香榧叶片中8种矿质元素年周期季节性变化规律[J]. 经济林研究, 2014, 32(2): 105 − 109. LIU Mengmeng, ZENG Yanru, JIANG Jianbin, et al. Seasonal dynamic changes of eight mineral elements in Torreya grandis leaf [J]. Non-wood For Res, 2014, 32(2): 105 − 109. [6] 邬玉芬, 刘萌萌, 储开江, 等. 香榧枝叶黄化与营养元素丰缺的相关性[J]. 福建林业科技, 2015, 42(2): 20 − 23, 66. WU Yufen, LIU Mengmeng, CHU Kaijiang, et al. Relationship between branch leaves yellowing degree and nutrimental elements levels in Torreya grandis ‘Merrillii’ [J]. J Fujian For Sci Technol, 2015, 42(2): 20 − 23, 66. [7] 詹利云, 刘琏, 曾燕如, 等. 雄性榧树天然居群表型多样性及优株初选[J]. 浙江农林大学学报, 2020, 37(6): 1120 − 1127. ZHAN Liyun, LIU Lian, ZENG Yanru, et al. Phenotypic diversity of male cones and the selection of superior plants among male populations in Torreya grandis [J]. J Zhejiang A&F Univ, 2020, 37(6): 1120 − 1127. [8] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. The Analysis Method of Soil Agricultural Chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000 [9] 朱祥枝, 陈秀萍, 蒋际谋, 等. 早钟6号枇杷结果母枝性状与果实形质的相关[J]. 福建农业学报, 2002, 17(1): 33 − 37. ZHU Xiangzhi, CHEN Xiuping, JIANG Jimou, et al. Correlationship between bearing base shoot character with form quality of fruit of Zaozhong 6 loquat [J]. Fujian J Agric Sci, 2002, 17(1): 33 − 37. [10] 甘露, 吴佳宏, 刘艳, 等. 杂交稻7个米质性状的变异及相关性分析[J]. 杂交水稻, 2019, 34(2): 63 − 68. GAN Lu, WU Jiahong, LIU Yan, et al. Variation and correlation analysis of seven grain quality traits of hybrid rice [J]. Hybrid Rice, 2019, 34(2): 63 − 68. [11] 张秀美, 王宏, 刘志, 等. ‘岳帅’苹果不同负载量光照分布与果实品质的关系[J]. 果树学报, 2017, 34(11): 1408 − 1414. ZHANG Xiumei, WANG Hong, LIU Zhi, et al. Relationships between distribution of relative light intensity and quality in ‘Yueshuai’ apple with different fruit loads [J]. J Fruit Sci, 2017, 34(11): 1408 − 1414. [12] 陈霞, 刘改霞. ‘香玲’核桃结果母枝与结果枝性状相关性研究[J]. 林业科技通讯, 2017(11): 13 − 16. CHEN Xia, LIU Gaixia. Character correlation study of ‘Xiangling’ walnut fruiting mother branch and fruiting branch [J]. For Sci Technol, 2017(11): 13 − 16. [13] 何学燕, 聂朝俊. 氮磷钾营养元素对香榧幼苗生长效果的影响[J]. 贵州林业科技, 2017, 45(1): 39 − 46. HE Xueyan, NIE Chaojun. Impact of nutritive element N, P, K on the effect of Torreya grandis seeding [J]. Guizhou For Sci Technol, 2017, 45(1): 39 − 46. [14] 陈艳秋, 曲柏宏, 牛广才, 等. 苹果梨果实矿质元素含量及其品质效应的研究[J]. 吉林农业科学, 2000, 25(6): 44 − 48. CHEN Yanqiu, QU Baihong, NIU Guangcai, et al. Study on seasonal variation of mineral elements content in pingguoli pear fruits [J]. J Jilin Agric Sci, 2000, 25(6): 44 − 48. [15] TAIZ L, ZEIGER E. 植物生理学[M]. 5版. 宋纯鹏. 译. 北京: 科学出版社, 2015. TAIZ L, ZEIGER E. Plant Physiology [M]. 5th ed. SONG Chunpeng. trans. Beijing: Science Press, 2015. [16] 李利红, 任敏. 杏树花芽分化期叶片矿质元素含量动态变化的研究[J]. 安徽农业科学, 2007, 35(21): 6361 − 6362. LI Lihong, REN Min. Studies on the dynamic variation of mineral element content in leaf during flower bud differentiation stage of apricot [J]. J Anhui Agric Sci, 2007, 35(21): 6361 − 6362. [17] 黎章矩, 戴文圣. 中国香榧[M]. 北京: 科学出版社, 2007. LI Zhangju, DAI Wensheng. Torreya grandis ‘Merrillii’ of China[M]. Beijing: Science Press, 2007. [18] 潘立忠, 张森, 王延书, 等. 常见落叶果树果实矿质营养的研究现状[J]. 安徽农学通报, 2006, 12(10): 77 − 80. PAN Lizhong, ZHANG Sen, WANG Yanshu, et al. Advances in research on mineral nutrition of fruit of common deciduous fruit tree [J]. Anhui Agric Sci Bull, 2006, 12(10): 77 − 80. [19] 王火焰, 王运华. 不同硼效率甘蓝型油菜品种悬浮细胞的硼钙营养效应[J]. 植物营养与肥料学报, 2002, 8(1): 100 − 104. WANG Huoyan, WANG Yunhua. Effects of boron-calcium nutrition on suspension-cell of rape cultivars with different boron efficiency [J]. Plant Nutr Fert Sci, 2002, 8(1): 100 − 104. [20] 刘亚林, 吴秀文, 闫磊, 等. 植物硼钙效应及其在细胞壁中互作机制的研究[J]. 植物科学学报, 2018, 36(5): 767 − 773. LIU Yalin, WU Xiuwen, YAN Lei, et al. Research progress on the effect of boron and calcium on plants and the interaction mechanism in the cell wall [J]. Plant Sci J, 2018, 36(5): 767 − 773. [21] 徐炜南. 硼对番茄生长及果实风味品质的影响[D]. 杨凌: 西北农林科技大学, 2017. XU Weinan. Effects of Boron on the Growthand Fruit Flavor of Tomato[D]. Yangling: Northwest A&F University, 2017. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200622
 点击查看大图
点击查看大图
计量
- 文章访问数: 1981
- HTML全文浏览量: 437
- PDF下载量: 43
- 被引次数: 0
