






-
随着经济和工业化的发展,酸雨污染日益成为中国重要的环境问题[1]。ZHAO等[2]和杨东贞等[3]发现:1999-2006年浙江省杭州市临安地区降水中除硫酸根离子(SO42-)外,硝酸根离子(NO3-)亦明显增加,尤其是2001年后NO3-对降水酸度的贡献逐步加大,降雨属于硫酸与硝酸混合型酸雨[4]。研究认为:酸雨抑制植物养分、水分的吸收和运输,导致植物无法正常生长[5-7]。酸雨可直接破坏叶肉细胞结构,使叶片细胞膜通透性增大,细胞膜保护酶的活性下降,膜脂过氧化作用加剧,植物体内清除活性氧平衡被打破,从而影响光合、呼吸以及代谢等生理生化进程而使植物受害[8-10]。秃瓣杜英Eleocarpus glabripetalus系杜英科Elaeocarpaceae杜英属Elaeocarpus常绿乔木,适于中性或微酸性的山地红壤、黄壤上生长,是亚热带地区常见树种,常在评价酸雨对亚热带地区森林系统影响时作为试材。研究发现:模拟酸雨处理下秃瓣杜英的生物量积累[11-12]、PSⅡ原初光能转化效率和PSⅡ潜在活性均有所上升[11]。俞飞等[13]则发现:不同酸雨处理下,杜英应对酸雨胁迫的途径不同。目前,有关酸雨影响植物生理与养分的研究较多,但对酸雨影响杜英生理和养分元素分配的协同探索较少,因此,本试验在研究不同酸雨梯度影响秃瓣杜英幼苗PSⅡ反应中心和幼苗养分分配的基础上,进一步探索秃瓣杜英幼苗响应酸雨胁迫的机制,为生物响应酸雨胁迫研究提供依据和参考。
-
试验地位于浙江省杭州市临安区浙江农林大学平山试验基地大棚(30°16′N,119°44′E)。属于中亚热带季风气候,温暖湿润,四季分明,年平均降水量约1 400.0 mm,全年平均气温为16.4 ℃,年无霜期250.0 d左右。区域森林覆盖率为76.5%。土壤类型为红壤。
-
选取高度为40~60 cm的1年生秃瓣杜英幼苗,于2011年1月移栽到高40 cm,直径30 cm的塑料花盆中,采用试验地红黄壤作为栽培土。幼苗随机分成10组,7株·组-1,苗木编号、挂牌后置于智能温室大棚内,保持棚内温度23~33 ℃,湿度55%~75%。缓苗期间用自来水浇灌,常规管理,2011年3月开始模拟酸雨喷洒[14]。模拟酸雨采用喷雾法,隔3 d喷洒1次,处理期间植物不浇水。
-
参照该地区酸性降水中平均离子组成和试验通用配比,按V(H2SO4):V(HNO3)=8:1配制母液,再用蒸馏水稀释成pH 2.5,pH 4.0及pH 5.6的酸雨水溶液,作为重度酸雨处理(T2.5),中度酸雨处理(T4.0)和轻度酸雨处理(T5.6)[15]。根据临安区近10 a平均降水量统计数据[13],计算每个酸雨等级所需的酸雨量,同时以等量的清水浇灌幼苗作为对照组(ck)。
-
2015年7月晴朗天气的9:00-11:00采用便携式叶绿素荧光仪(PAM-2100,Walz,德国)测定初始荧光值(Fo),黑暗中最大荧光值(Fm),实时荧光值(Ft),最大光化学量子产量光系统Ⅱ(PSⅡ)的最大光化学效率(Fv/Fm),实际光化学量子产量(Yield),光化学猝灭系数(qP)等叶绿素荧光参数。随机选取5株·处理-1,并在每株不同方位中上部随机选取5片成熟叶片进行测定。所得数据为3 d的平均值。
-
选取进行荧光测定相同的叶片,用便携式叶绿素含量测定仪(SPAD-502,日本)测定叶绿素相对含量,测定3次·叶片-1,以5个叶片作为各组的平均值。
-
2015年8月初,选择之前进行荧光实验的5株植株,各植株上中下各部位采集秃瓣杜英叶3片。幼苗叶氮元素质量分数用半微量凯式定氮法测定,磷元素质量分数用钼锑抗比色法测定[16-17];钾、镁元素利用电感耦合等离子体发射光谱仪(PerkinElmer Inc.,美国)进行测定[18]。
-
2015年11月全株收获,将黏附于秃瓣杜英幼苗上的泥土和污物用水冲洗干净,105 ℃下杀青5 min后,60 ℃下于烘箱中烘至恒量,分别称量地上部分和地下部分的生物量。
-
将取回来的土样自然风干后磨碎,过100目筛。取各处理的土壤样品0.15 g预消解过夜,移入微波消解仪中消解至变色。得到的澄清、透明溶液摇匀静置待测,同时制备空白[19]。用电感耦合等离子体发射光谱仪测定金属铜和金属锌的质量分数,重复3次·样品-1。
-
转移系数(TF)可以表征植物地上部分金属与根部中金属质量分数的比值[20],表示如下:转移系数=地上部分金属质量分数/根部金属质量分数。生物富集系数(BCF)是植物体内某种金属元素的质量分数与土壤内该种金属元素的总质量分数之比[20],表示如下:富集系数=植物体内金属元素质量分数/土壤内金属元素质量分数。
-
用SPSS 16.0软件统计秃瓣杜英幼苗的生长参数、叶绿素相对含量、荧光参数以及植株和土壤元素,对所有数据进行正态性与齐性检验后,利用单因素方差分析(one-way ANOVA)和最小显著差法(LSD)比较不同数据组间的差异,用Excel软件作图。
-
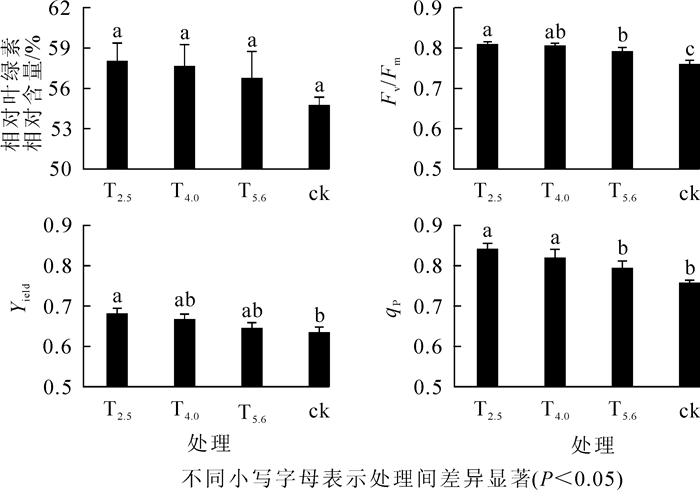
由图 1可知:酸雨处理组的叶绿素相对含量均高于对照组,且叶绿素相对含量随着酸雨pH值的降低而升高,但是各处理组与对照组间差异不显著(P>0.05)。Fv/Fm的变化跟叶绿素相对含量一致,也是随着酸雨pH值的降低而升高的,但各处理组与对照组之间均有显著差异(P<0.05),T5.6,T4.0和T2.5分别较对照组升高4.22%,6.07%和6.86%,说明酸雨有利于叶片PSⅡ原初光能转化效率的提高。同样地,Yield和qP也是随着酸雨pH的降低而升高,其中重度酸雨处理组Yield显著高于对照组(P<0.05),重、中度酸雨处理组qP显著高于对照组(P<0.05)。

图 1 不同酸雨处理下秃瓣杜英幼苗叶绿素参数
Figure 1. Changes of chlorophyll fluorescence parameters of Eleocarpus glabripetalus seedlings with different acid rain treatments
-
由图 2可知:地上部生物量最高的是对照组,随着酸雨pH值的降低,各处理组的地上部生物量相比对照组分别降低了29.27%,4.88%和26.83%;根系生物量最低的是对照组,随着酸雨pH值的降低,各处理组的根系生物量相比对照组分别提高了22.22%,5.56%和33.33%。但无论地上部或根系,各处理组与对照组之间差异均不显著(P>0.05)。

图 2 不同酸雨处理下秃瓣杜英幼苗各组织生物量变化
Figure 2. Changes of each tissue biomass of Eleocarpus glabripetalus seedlings with different acid rain treatments
-
由图 3A可知:秃瓣杜英幼苗中氮元素主要存在于叶片;与对照组相比,各处理组叶片氮质量分数均有所提高,但差异不显著(P>0.05);各处理组茎中氮质量分数与对照组相比,均无显著性差异(P>0.05);轻度酸雨处理组根氮质量分数显著大于其他组(P<0.05)。

图 3 不同酸雨梯度下秃瓣杜英幼苗器官中的元素质量分数变化
Figure 3. Changes of N, P, K and Mg in the organs of E. glabripetalus seedlings with different acid rain treatments
图 3B显示:所有器官磷质量分数总和最高的是对照组;对照组与处理组叶中的磷质量分数均无显著性差异(P>0.05);茎中磷质量分数从大到小依次为对照组,T4.0,T2.5和T5.6,除中度酸雨组外,其他处理组与对照组差异显著(P<0.05);根中磷质量分数表现为重、中度酸雨组显著高于对照组(P<0.05)。
各处理组与对照组叶片中钾质量分数无显著差异(P>0.05);茎中钾质量分数最高的是对照组,显著高于重、轻度酸雨组(P<0.05);根中钾质量分数最低的是中度酸雨组,与对照组差异显著(P<0.05)。
各器官中镁质量分数各组之间均无显著差异(P>0.05),总量最高的是对照组。叶片中镁质量分数从大到小依次为T4.0,T5.6,T2.5和对照,在根中排序为对照,T5.6,T2.5和T4.0,在茎中排序为T4.0,对照,T5.6和T2.5。
-
由表 1可知:处理组幼苗组织中铜元素和锌元素主要集中在地上部分。重、中度酸雨处理下,幼苗地上部铜质量分数显著高于对照组(P<0.05);根系铜质量分数低于对照组,其中重度酸雨组与对照组差异显著(P<0.05)。各处理组秃瓣杜英幼苗的地上部锌质量分数均高于对照组,其中,重度酸雨组与对照组之间差异显著(P<0.05)。重、中度酸雨组根系锌质量分数显著小于对照组(P<0.05)。
表 1 秃瓣杜英幼苗在不同酸雨酸度下各组织中铜和锌质量分数
Table 1. Heavy metal Cu and Zn concentrations in different tissues of Eleocarpus glabripetalus seedlings with different acid rain treatments
试验组 w铜/(mg·kg-1) w锌/(mg·kg-1) 地上部 根系 地上 根系 T2.5 13.50 ± 0.55 a 5.93 ± 0.03 c 177.16 ± 6.28 a 13.50 ± 0.77 b T4.0 15.66 ± 1.80 a 13.71 ± 1.01 b 99.55 ± 7.33 ab 6.12 ± 0.76 b T5.6 11.14 ± 1.39 ab 27.15 ± 3.38 a 138.87 ± 6.85 ab 72.07 ± 6.71 a ck 11.41 ± 0.91 b 14.83 ± 2.04 b 23.49 ± 2.21 b 68.35 ± 7.46 a 说明:表内数据为平均值±标准误。同列不同字母表示差异显著(P<0.05) 幼苗在酸雨处理下的转移系数值由表 2所示。幼苗中铜转移系数随酸雨pH值的降低而升高,重度酸雨处理下幼苗中铜转移系数最高,为2.27。各处理组锌转移系数均高于对照组,其中重、中度酸雨处理与对照组差异显著(P<0.05),中度酸雨处理下锌转移系数最高,为16.79。说明酸雨处理增加了秃瓣杜英幼苗对重金属的转移能力。
表 2 不同酸雨酸度下秃瓣杜英幼苗中铜、锌的转移系数和生物富集系数
Table 2. Translocation factor and bioconcentration factor of Cu and Zn with different acid rain treatments
试验组 转移系数 生物富集系数 铜 锌 铜 锌 T2.5 2.271 ± 0.112 a 10.837 ± 0.483 a 0.014 ± 0.001 ab 0.712 ± 0.025 a T4.0 1.169 ± 0.203 b 16.790 ± 0.240 a 0.019 ± 0.002 a 0.420 ± 0.035 a T5.6 0.423 ± 0.072 c 2.033 ± 0.081 b 0.011 ± 0.002 b 0.546 ± 0.026 a ck 0.792 ± 0.081 bc 0.409 ± 0.011 b 0.020 ± 0.002 a 0.093 ± 0.003 b 说明:表内数据为平均值±标准误。同列不同字母表示差异显著(P<0.05) 酸雨处理后铜元素的生物富集系数为0.011~0.019,均小于对照组,说明酸雨处理不利于秃瓣杜英幼苗对重金属铜的积累;处理组锌的生物富集系数均显著高于对照组(P<0.05),说明酸雨处理提高了秃瓣杜英幼苗对重金属锌的积累能力。
-
测定植物叶片的叶绿素相对含量可以间接反映植物叶片叶绿素的情况[21-24],叶绿素荧光动力学技术可以获得植物耐受环境胁迫的能力和各种胁迫已经损害光系统的程度等信息[25]。研究发现:重度酸雨下,杨梅Myrica rubra叶绿素各荧光参数均降低[26];在pH 2.5条件下,水稻Oryza sativa叶片质膜(H+-ATPase)活性,细胞渗透势和钾离子(K+)和镁离子(Mg2+)明显下降[27];五爪金龙Ipomoea cairica叶片净光合速率随酸雨pH值的下降而降低[28]。也有得出相反结论。田大伦等[29]发现:pH 3.0和pH 4.0的酸雨提高了樟树Cinnamomum camphora最大净光合速率和表观量子效率;在pH 2.5的酸雨强度下,青冈Quercus glauca幼苗的光合参数、叶绿素荧光参数与对照组相比,明显增加[30]。
本研究发现秃瓣杜英幼苗叶绿素参数随不同酸雨变化而变化。随着酸雨pH值降低,叶绿素相对含量,Fv/Fm,Yield和qP均升高,说明酸雨提高了幼苗的叶绿素含量,增强秃瓣杜英幼苗的光合能力。
对各试验组幼苗组织生物量的测定可知:所有酸雨处理组地上部生物量均小于对照组,而根系生物量大于对照组,说明酸雨导致植物生物量重新分配,使秃瓣杜英幼苗根系更加健壮;根系通过吸收土壤中的矿质元素,使植物的生长效能提高,植物对环境胁迫的抵抗力增强,由此认为:秃瓣杜英具有一定的抗酸特性,即酸雨促进秃瓣杜英幼苗的生长,提高幼苗适应不良环境的能力。
氮是构成蛋白质的重要元素,对植物的光合作用起着重要的影响。研究发现:秃瓣杜英幼苗中氮元素主要存在于叶片,酸雨处理组叶片中氮质量分数均高于对照组,说明酸雨促进秃瓣杜英叶片对氮元素的吸收,有利于秃瓣杜英的光合作用,与WOOD等[31]在北美乔松Pinus strobus的模拟酸雨试验中发现pH 4.0以下的模拟酸雨可促进幼苗生长的结论一致,并指出是由于酸雨中硝酸根(NO3-)的施肥作用的效果,即酸雨处理后对秃瓣杜英幼苗起到一定的氮补给作用。磷、钾作为植物各种生理代谢活动的重要参与者,对植物同化力的形成非常重要。酸雨处理组茎、叶部磷钾质量分数均小于对照组,而重、中度酸雨处理组根部磷钾质量分数大于对照组,表明酸雨抑制秃瓣杜英茎、叶对磷钾元素吸收的同时促进根系对磷钾元素的吸收,这可能是导致酸雨处理组的地上部生物量小于对照组,而酸雨组根部生物量大于对照组的原因。镁是组成叶绿素的重要成分,对植物光合作用非常重要,酸雨组叶片镁质量分数均大于对照组,但酸雨组与对照组各器官中镁质量分数无显著差异,这与段小华等[32]研究模拟酸雨对茶树根茎和叶中镁含量没有明显影响的结果是一致的。
铜和锌是植物正常生长和发育所必需的元素,但土壤中过量的锌和铜会抑制植物的生长。LEVY等[33]报道称,植物中铜质量分数正常范围为3~30 mg·kg-1,锌为10~150 mg·kg-1;本试验发现:各组织中铜质量分数的变化为5.93~27.15 mg·kg-1,锌为6.12~177.16 mg·kg-1,说明铜没有对秃瓣杜英产生毒害,而是作为必要元素用以构成铜蛋白,参与植物光合作用和氮素代谢,提高了叶绿素的稳定性。重度酸雨处理后,植物中铜转移系数增大,说明叶片获得了更多铜元素,从而提高叶片叶绿素含量。相比之下,酸雨处理后秃瓣杜英幼苗叶片锌质量分数超出了正常范围,根部尽管属于正常范围内,但转移系数和富集系数均高于对照组,说明秃瓣杜英通过迅速转移根部的锌到地上部分来避免根系受到锌的毒害,秃瓣杜英在酸雨刺激下对重金属锌的忍耐作用得到了提高。王菊等[34]和严明理等[35]研究均发现:酸雨促进重金属的转移和释放,但地下部重金属含量较地上部多,与本研究结果相反,说明重金属的转移和释放不仅取决于酸雨pH值,可能还与植物本身有关。
综上所述:模拟酸雨处理秃瓣杜英幼苗后,幼苗没有受到酸雨的极度危害;随着酸雨pH值的降低,酸雨各叶绿素荧光参数有所增加,说明酸雨提高了秃瓣杜英幼苗的光合能力。幼苗对酸雨的忍耐性主要表现在:浇酸之后,酸雨进入土壤,受土壤本身理化特性的缓冲作用,未对根系造成直接危害;其中的氮元素被植物逐渐吸收,使得幼苗叶片氮质量分数增加。酸雨对土壤中某些元素的活化作用,使得酸雨处理增加了幼苗叶片镁元素和地上部铜元素,对植物生长有利。随着酸雨pH值的降低,秃瓣杜英幼苗地上部生物量积累下降,根系生物量积累增加,与对照组存在显著差异;其原因首先是叶片磷、钾元素的减少,对幼苗的新陈代谢造成影响,其次则可能是为避免锌对秃瓣杜英根系产生毒害,部分能量用于转移根系锌元素。这也是秃瓣杜英耐酸的一种机理,因此,今后可加强对秃瓣杜英耐酸性机制的研究。
Chlorophyll fluorescence response and nutrient distribution of Elaeocarpus glabripetalus seedlings with simulated acid rain
-
摘要: 利用模拟酸雨喷淋试验,研究不同酸雨梯度[pH 2.5(重度酸雨)、pH 4.0(中度酸雨)和pH 5.6(轻度酸雨)]对秃瓣杜英Eleocarpus glabripetalus幼苗叶绿素参数、生物量及对土壤元素吸收的影响。结果表明:幼苗叶绿素参数随酸雨pH值的降低而升高,各处理组相对叶绿素含量均高于对照组;最大光化学量子产量(Fv/Fm)随酸雨pH值的降低而升高,各处理组较对照组升高4.22%,6.07%和6.86%,与对照组差异显著(P < 0.05)。各处理组叶片氮元素质量分数均大于对照组,但差异不显著(P>0.05);所有器官磷、钾质量分数总和均小于对照组,重、轻度酸雨组茎的磷质量分数显著低于对照组(P < 0.05);所有器官镁质量分数均小于对照组,但差异不显著(P>0.05)。重、中度酸雨组幼苗地上部铜元素质量分数和转移系数高于对照组,各处理组锌元素转移系数和富集系数均显著高于对照组(P < 0.05)。综合分析认为:酸雨增加了幼苗的相对叶绿素含量,提高了幼苗的光合能力;促进了幼苗生物量重新分配,使得根系更加健壮,从而促进根系对土壤矿质元素的吸收,增强了植物对环境胁迫的抵抗力;提高了根系锌元素转移系数,从而加大幼苗对重金属锌的忍耐作用。Abstract: To find the effects of chlorophyll parameters and biomass of Elaeocarpus glabripetalus seedlings and the absorption of soil elements for different levels of acidity, a simulated acid rain spray test with three acidity levels[pH 2.5 (severe acid rain), pH 4.0 (moderate acid), and pH 5.6 (mild acid rain)], as well as a control group (ck) was used. Results showed that the chlorophyll parameters of E. glabripetalus seedlings increased with a decrease in the pH of acid rain. The relative chlorophyll content (SPAD) in each treatment group was higher than the control group. The maximum chemical quantum yield (Fv/Fm) increased with a decrease in pH of acid rain, and there was a significant difference between the treatment groups and ck (P < 0.05). The leaf nitrogen (N) mass fraction of each treatment group was not significantly different from ck (P>0.05). The phosphorus (P) mass fraction for the heavy and mild acid rain group was significantly lower than ck (P>0.05). Seedlings in the upper part of the mass fraction and transfer coefficients of heavy and moderate acid groups were higher than ck. The zinc (Zn) transfer coefficients and enrichment coefficients of each treatment group were significantly higher than ck (P < 0.05). Based on the comprehensive analysis, acid rain increased the chlorophyll content of seedlings, and improved the photosynthetic capacity of seedlings. Acid rain also promoted the redistribution of seedlings biomass, making the root system more robust, thus promoting the absorption of mineral elements in the roots and enhancing the plant's resistance to environmental stress. In addition, acid rain increased the Zn transfer coefficient of roots thereby increasing the tolerance of seedlings to heavy metal Zn. Overall, this study provided a scientific basis for nutrient distribution characteristics and physiological response mechanism of Elaeocarpus glabripetalus seedlings with acid rain stress.
-
Key words:
- plant physiology /
- acid rain stress /
- biomass /
- chlorophyll parameters /
- Elaeocarpus glabripetalus /
- mineral element
-
图 1 不同酸雨处理下秃瓣杜英幼苗叶绿素参数
Figure 1 Changes of chlorophyll fluorescence parameters of Eleocarpus glabripetalus seedlings with different acid rain treatments
图 2 不同酸雨处理下秃瓣杜英幼苗各组织生物量变化
Figure 2 Changes of each tissue biomass of Eleocarpus glabripetalus seedlings with different acid rain treatments
图 3 不同酸雨梯度下秃瓣杜英幼苗器官中的元素质量分数变化
Figure 3 Changes of N, P, K and Mg in the organs of E. glabripetalus seedlings with different acid rain treatments
表 1 秃瓣杜英幼苗在不同酸雨酸度下各组织中铜和锌质量分数
Table 1. Heavy metal Cu and Zn concentrations in different tissues of Eleocarpus glabripetalus seedlings with different acid rain treatments
试验组 w铜/(mg·kg-1) w锌/(mg·kg-1) 地上部 根系 地上 根系 T2.5 13.50 ± 0.55 a 5.93 ± 0.03 c 177.16 ± 6.28 a 13.50 ± 0.77 b T4.0 15.66 ± 1.80 a 13.71 ± 1.01 b 99.55 ± 7.33 ab 6.12 ± 0.76 b T5.6 11.14 ± 1.39 ab 27.15 ± 3.38 a 138.87 ± 6.85 ab 72.07 ± 6.71 a ck 11.41 ± 0.91 b 14.83 ± 2.04 b 23.49 ± 2.21 b 68.35 ± 7.46 a 说明:表内数据为平均值±标准误。同列不同字母表示差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同酸雨酸度下秃瓣杜英幼苗中铜、锌的转移系数和生物富集系数
Table 2. Translocation factor and bioconcentration factor of Cu and Zn with different acid rain treatments
试验组 转移系数 生物富集系数 铜 锌 铜 锌 T2.5 2.271 ± 0.112 a 10.837 ± 0.483 a 0.014 ± 0.001 ab 0.712 ± 0.025 a T4.0 1.169 ± 0.203 b 16.790 ± 0.240 a 0.019 ± 0.002 a 0.420 ± 0.035 a T5.6 0.423 ± 0.072 c 2.033 ± 0.081 b 0.011 ± 0.002 b 0.546 ± 0.026 a ck 0.792 ± 0.081 bc 0.409 ± 0.011 b 0.020 ± 0.002 a 0.093 ± 0.003 b 说明:表内数据为平均值±标准误。同列不同字母表示差异显著(P<0.05)
下载: 导出CSV
-
[1] WANG Yanping, WANG Ya, KAI Wenbin, et al. Transcriptional regulation of abscisic acid signal core components during cucumber seed germination and under Cu2+, Zn2+, NaCl and simulated acid rain stresses[J]. Plant Physiol Biochem, 2014, 76(5):67-76. [2] ZHAO Yanxia, HOU Qing. Characteristics of the acid rain variation in China during 1993-2006 and associated causes[J]. Acta Meteorol Sin, 2010, 24(2):239-250. [3] 杨东贞, 周怀刚, 张忠华.中国区域空气污染本底站的降水化学特征[J].应用气象学报, 2002, 13(4):430-439. YANG Dongzhen, ZHOU Huaigang, ZHANG Zhonghua. Chemical characteristics of precipitation in regional air pollution background stations of China[J]. J Appl Meteorol Sci, 2002, 13(4):430-439. [4] 李正泉, 马浩, 毛裕定, 等.临安本底站酸雨长期观测特征分析[J].环境科学, 2014, 35(2):481-488. LI Zhengquan, MA Hao, MAO Yuding, et al. Analysis of acid rain characteristics of Lin'an regional background station using long-term observation data[J]. Environ Sci, 2014, 35(2):481-488. [5] 王利, 杨洪强, 张召, 等.根区酸化对平邑甜茶叶片光系统Ⅱ活性及光合速率的影响[J].林业科学, 2011, 47(10):167-171. WANG Li, YANG Hongqiang, ZHANG Zhao, et al. Effects of rhizosphere acidity on activity of photosystem Ⅱ and photosynthetic rate in Malus hupehensis leaves[J]. Sci Silv Sin, 2011, 47(10):167-171. [6] SINGH A, AGRAWAL M. Acid rain and its ecological consequences[J]. J Environ Biol, 2008, 29(1):15-24. [7] SHUKLA J B, SUNDAR S, SHIVANGI S, et al. Modeling and analysis of the acid rain formation due to precipitation and its effect on plant species[J]. Nat Res Modeling, 2013, 26(1):53-65. [8] 李永裕, 潘腾飞, 余东, 等.模拟酸雨对龙眼叶片PSⅡ反应中心和自由基代谢的影响[J].生态学报, 2012, 32(24):7866-7873. LI Yongyu, PAN Tengfei, YU Dong, et al. Effects of simulated acid rain stress on the PSⅡ reaction center and free radical metabolism in leaves of longan[J]. Acta Ecol Sin, 2012, 32(24):7866-7873. [9] 侯维, 潘远智.酸雨对勋章菊保护酶活性及叶绿素荧光参数的影响[J].核农学报, 2013, 27(7):1054-1059. HOU Wei, PAN Yuanzhi. Effects of acid rain stress on antioxidant enzyme activity and chlorophyll fluorescence in leaves of Gazania hybrids[J]. J Nucl Agric Sci, 2013, 27(7):1054-1059. [10] 金清, 江洪, 余树全, 等.酸雨胁迫对苦槠幼苗气体交换与叶绿素荧光的影响[J].植物生态学报, 2010, 34(9):1117-1124. JIN Qing, JIANG Hong, YU Shuquan, et al. Effects of acid rain stress on gas exchange and chlorophyll fluorescence of Castannopsis sclerophylla seedlings[J]. Chin J Plant Ecol, 2010, 34(9):1117-1124. [11] LIU Meihua, YI Lita, YU Shuquan, et al. Chlorophyll fluorescence characteristics and the growth response of Elaeocarpus glabripetalusto simulated acid rain[J]. Photosynthetica, 2015, 53(1):23-28. [12] 项敬银, 俞飞, 伊力塔, 等.模拟酸雨和镉复合处理对秃瓣杜英生长和荧光参数的影响[J].生态学报, 2018, 38(15):5443-5451. XIANG Jingyin, YU Fei, YI Lita, et al. Combined effects of artificial acid rain and Cd on the growth and fluorescence parameters of Elaeocarpus glabripetalus[J]. Acta Ecol Sin, 2018, 38(15):5443-5451. [13] 俞飞, 宋琦, 刘美华, 等.模拟酸雨不同处理对秃瓣杜英幼苗生物量和生理的影响[J].林业科学, 2016, 52(5):92-100. YU Fei, SONG Qi, LIU Meihua, et al. Effects of different acid rain treatments on biomass allocation and physiological characteristics in Elaeocarpus glabripetalus seedlings[J]. Sci Silv Sin, 2016, 52(5):92-100. [14] 田苏奎, 李领寰, 伊力塔, 等.模拟酸雨对杜英幼苗生长及养分元素含量的影响[J].华东森林经理, 2016, 30(3):38-42. TIAN Sukui, LI Linghuan, YI Lita, et al. Effects of simulated acid rain on Elaeocarpus decipiens seedling growth and nutrient distribution[J]. East China For Manage, 2016, 30(3):38-42. [15] 汪赛, 伊力塔, 余树全, 等.模拟酸雨对青冈光合及叶绿素荧光参数的影响[J].应用生态学报, 2014, 25(8):2183-2192. WANG Sai, YI Lita, YU Shuquan, et al. Effects of simulating acid rain on photosynthesis and chlorophyll fluorescence parameters of Quercus glauca[J]. Chin J Appl Ecol, 2014, 20(8):2183-2192. [16] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社, 2000. [17] 王沙生, 高荣孚, 吴贯明.植物生理学[M].北京:中国林业出版社, 2000. [18] 洪博.萝芙木化学成分与质量控制方法研究[D].沈阳: 沈阳药科大学, 2011. HONG Bo. Studies on Chemical Constituents and Quality Control Methods of Rauvolfia verticillata[D]. Shenyang: Shenyang Pharmaceutical University, 2011. [19] 宁寻安, 周云, 刘敬勇, 等.微波消解-火焰原子吸收光谱法测定钢铁厂废旧除尘布袋中重金属[J].光谱学与光谱分析, 2011, 31(9):2565-2568. NING Xun'an, ZHOU Yun, LIU Jingyong, et al. Determination of metals in waste bag filter of steel works by microwave digestion-flame atomic absorption spectrometry[J]. Spectrosc Spectral Anal, 2011, 31(9):2565-2568. [20] 周健民.土壤学大辞典[M].北京:科学出版社, 2013. [21] 姜丽芬, 石福臣, 王化田, 等.叶绿素计SPAD-502在林业上应用[J].生态学杂志, 2005, 24(12):1543-1548. JIANG Lifen, SHI Fuchen, WANG Huatian, et al. Application tryout of chlorophyll meter SPAD-502 in forestry[J]. Chin J Ecol, 2005, 24(12):1543-1548. [22] 王彬, 田正凤, 应彬彬, 等.高温胁迫对樟树光合性能的影响[J].浙江农林大学学报, 2019, 36(1):47-53. WANG Bin, TIAN Zhengfeng, YING Binbin, et al. Photosynthetic abilities in Cinnamomum camphora with high temperature stress[J]. J Zhejiang A & F Univ, 2019, 36(1):47-53. [23] 宋晓东, 江洪, 余树全, 等.亚热带典型常绿阔叶树种叶片叶绿素含量与其高光谱特征间的关系[J].生态学报, 2008, 28(5):1959-1963. SONG Xiaodong, JIANG Hong, YU Shuquan, et al. Relationship between chlorophyll concentrations and spectral reflectance feature of the typical evergreen hardwood species in subtropical region of China[J]. Acta Ecol Sin, 2008, 28(5):1959-1963. [24] MARKWELL J, OSTERMAN J C, MITCHELL J L. Calibration of the minoltaSPAD-502 leaf chlorophyll meter[J]. Photosynth Res, 1995, 46(3):467-472. [25] 吴根良, 何勇, 王永传, 等.不同光照强度下卡特兰和蝴蝶兰光合作用和叶绿素荧光参数日变化[J].浙江林学院学报, 2008, 25(6):733-738. WU Genliang, HE Yong, WANG Yongchuan, et al. Diurnal changes of photosynthesis and chlorophyll fluorescence in Cattleya×hybrida and Phalaenopsis amabilis with different light intensities[J]. J Zhejiang For Coll, 2008, 25(6):733-738. [26] 于晓鹏, 伊力塔, 余树全, 等.不同酸雨胁迫梯度和处理方式对杨梅幼苗叶绿素荧光特性的影响[J].生态学杂志, 2015, 34(5):1246-1252. YU Xiaopeng, YI Lita, YU Shuquan, et al. Effects of acid rainfall intensities and treatment ways on chlorophyll fluorescence parameters of Myrica rubra seedlings[J]. Chin J Ecol, 2015, 34(5):1246-1252. [27] 卜津津, 苏垒, 吕霞, 等.模拟酸雨对水稻叶片质膜H+-ATPase活性与矿质元素含量的影响[J].环境科学学报, 2015, 35(9):3020-3024. BU Jinjin, SU Lei, LÜ Xia, et al. Effect of simulated acid rain on plasma membrane H+-ATPase activity and mineral elements contents in rice leaves[J]. Acta Sci Circumstantiae, 2015, 35(9):3020-3024. [28] 赵则海.模拟酸雨对五爪金龙幼苗光合生理特性的影响[J].生态环境学报, 2014, 23(9):1498-1502. ZHAO Zehai. Effects of simulated acid rain on photosynthetic physiology characteristics of Ipomoea cairica seedling[J]. Ecol Environ Sci, 2014, 23(9):1498-1502. [29] 田大伦, 付晓萍, 方晰, 等.模拟酸雨对樟树幼苗光合特性的影响[J].林业科学, 2007, 43(8):29-35. TIAN Dalun, FU Xiaoping, FANG Xi, et al. Effect of simulated acid rain on photosynthetic characteristics in Cinnamomum camphora seedlings[J]. Sci Silv Sin, 2007, 43(8):29-35. [30] 李佳, 江洪, 余树全, 等.模拟酸雨胁迫对青冈幼苗光合特性和叶绿素荧光参数的影响[J].应用生态学报, 2009, 20(9):2092-2096. LI Jia, JIANG Hong, YU Shuquan, et al. Effects of simulated acid rain on Quercus glauca seedlings photosynthesis and chlorophyll fluorescence[J]. Chin J Appl Ecol, 2009, 20(9):2092-2096. [31] WOOD T, BORMANN F H. Short-term effects of a simulated acid rain upon the growth and nutrient relations of Pinus strobus L.[J]. Water Air Soil Pollut, 1977, 7(4):479-488. [32] 段小华, 邓泽元, 胡小飞, 等.模拟酸雨和外源铝对茶树铝及一些营养元素吸收积累的影响[J].农业环境科学学报, 2010, 29(10):1936-1942. DUAN Xiaohua, DENG Zeyuan, HU Xiaofei, et al. Effects of simulated acid rain and exogenous Al on assimilation and accumulation of Al and some nutrition elements of tea plants[J]. J Agro-environ Sci, 2010, 29(10):1936-1942. [33] LEVY D B, REDENTE E F, UPHOFF G D. Evaluating the phytotoxity of Pb-Zn tailings to big bluestem (Andropogon gerardii vitman) and switchgrass (Panicum virgatum L.)[J]. Soil Sci, 1999, 164(6):363-375. [34] 王菊, 赵秀敏, 刘禹彤, 等.模拟酸雨强度与小白菜体内Zn含量的正相关性研究[J].广东农业科学, 2013(17):21-23. WANG Ju, ZHAO Xiumin, LIU Yutong, et al. Positive correlation between simulated acid rain intensity and Zn content of pakchoi[J]. Guangdong Agric Sci, 2013(17):21-23. [35] 严明理, 冯涛, 刘丽莉, 等.模拟酸雨和Pb复合污染对芥菜型油菜的生理特性和Pb富集的影响[J].环境科学学报, 2009, 29(10):2172-2179. YAN Mingli, FENG Tao, LIU Lili, et al. Combined effects of artificial acid rain and Pb on physiological characteristics of Brassica juncea and its Pb uptake[J]. Acta Sci Circumstantiae, 2009, 29(10):2172-2179. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.03.004
 点击查看大图
点击查看大图
计量
- 文章访问数: 5232
- HTML全文浏览量: 934
- PDF下载量: 67
- 被引次数: 0
